
Regulation of CD4+ and CD8+ T Cell Biology by Short-Chain Fatty Acids and Its Relevance for Autoimmune Pathology

 , , ,
, , ,  and
and
Abstract
:1. Introduction
What Are Short-Chain Fatty Acids?
2. Effect of SCFAs on General Function of T Cells in the Gut and Periphery
2.1. Effect on Tregs
2.2. Effect on Effector Th17 Cells
2.3. Effect on CD8+ Cells
2.4. Anti-Inflammatory Effect in Periphery Is Dose-Dependent
3. SCFA Interventions in Inflammatory Disorders
3.1. Multiple Sclerosis
3.2. Type I Diabetes
3.3. Rheumatoid Arthritis and Short Chain Fatty Acids
3.4. Inflammatory Bowel Disease
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dopkins, N.; Nagarkatti, P.S.; Nagarkatti, M. The Role of Gut Microbiome and Associated Metabolome in the Regulation of Neuroinflammation in Multiple Sclerosis and Its Implications in Attenuating Chronic Inflammation in Other Inflammatory and Autoimmune Disorders. Immunology 2018, 154, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Corrêa-Oliveira, R.; Fachi, J.L.; Vieira, A.; Sato, F.T.; Vinolo, M.A.R. Regulation of immune cell function by short-chain fatty acids. Clin. Transl. Immunol. 2016, 5, e73. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Qie, Y.; Park, J.; Kim, C.H. Gut Microbial Metabolites Fuel Host Antibody Responses. Cell Host Microbe 2016, 20, 202–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, T.; Miyauchi, E.; Kanaya, T.; Kato, T.; Nakanishi, Y.; Watanabe, T.; Kitami, T.; Taida, T.; Sasaki, T.; Negishi, H. Acetate Differentially Regulates IgA Reactivity to Commensal Bacteria. Nature 2021, 595, 560–564. [Google Scholar] [CrossRef]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly-Y, M.; Glickman, J.N.; Garrett, W.S. The Microbial Metabolites, Short-Chain Fatty Acids, Regulate Colonic T Reg Cell Homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J.; Kim, C.H. Short-Chain Fatty Acids Induce Both Effector and Regulatory T Cells by Suppression of Histone Deacetylases and Regulation of the MTOR–S6K Pathway. Mucosal Immunol. 2015, 8, 80–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummings, J.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; MacFarlane, G. Short Chain Fatty Acids in Human Large Intestine, Portal, Hepatic and Venous Blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [Green Version]
- Blaak, E.E.; Canfora, E.E.; Theis, S.; Frost, G.; Groen, A.K.; Mithieux, G.; Nauta, A.; Scott, K.; Stahl, B.; Van Harsselaar, J. Short Chain Fatty Acids in Human Gut and Metabolic Health. Benef. Microbes 2020, 11, 411–455. [Google Scholar] [CrossRef]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; De Los Reyes-gavilán, C.G.; Salazar, N. Intestinal Short Chain Fatty Acids and Their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deleu, S.; Machiels, K.; Raes, J.; Verbeke, K.; Vermeire, S. Short Chain Fatty Acids and Its Producing Organisms: An Overlooked Therapy for IBD? eBioMedicine 2021, 66, 103293. [Google Scholar] [CrossRef] [PubMed]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donohoe, D.R.; Garge, N.; Zhang, X.; Sun, W.; O’Connell, T.M.; Bunger, M.K.; Bultman, S.J. The Microbiome and Butyrate Regulate Energy Metabolism and Autophagy in the Mammalian Colon. Cell Metab. 2011, 13, 517–526. [Google Scholar] [CrossRef] [Green Version]
- Iraporda, C.; Errea, A.; Romanin, D.E.; Cayet, D.; Pereyra, E.; Pignataro, O.; Sirard, J.C.; Garrote, G.L.; Abraham, A.G.; Rumbo, M. Lactate and Short Chain Fatty Acids Produced by Microbial Fermentation Downregulate Proinflammatory Responses in Intestinal Epithelial Cells and Myeloid Cells. Immunobiology 2015, 220, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; McKenzie, C.I.; Hijikata, A.; Wong, C. Metabolite-Sensing Receptors GPR43 and GPR109A Facilitate Dietary Fibre-Induced Gut Homeostasis through Regulation of the Inflammasome. Nat. Commun. 2015, 6, 6734. [Google Scholar] [CrossRef] [Green Version]
- Sakata, T. Pitfalls in Short-Chain Fatty Acid Research: A Methodological Review. Anim. Sci. J. 2019, 90, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsson, A.; Gustavsen, S.; Nguyen, T.D.; Nyman, M.; Langkilde, A.R.; Hansen, T.H.; Sellebjerg, F.; Oturai, A.B.; Bach Søndergaard, H. Serum Short-Chain Fatty Acids and Associations With Inflammation in Newly Diagnosed Patients With Multiple Sclerosis and Healthy Controls. Front. Immunol. 2021, 12, 661493. [Google Scholar] [CrossRef]
- Jakobsdottir, G.; Bjerregaard, J.H.; Skovbjerg, H.; Nyman, M. Fasting Serum Concentration of Short-Chain Fatty Acids in Subjects with Microscopic Colitis and Celiac Disease: No Difference Compared with Controls, but between Genders. Scand. J. Gastroenterol. 2013, 48, 696–701. [Google Scholar] [CrossRef]
- Pomare, E.W.; Branch, W.J.; Cummings, J.H. Carbohydrate Fermentation in the Human Colon and Its Relation to Acetate Concentrations in Venous Blood. J. Clin. Investig. 1985, 75, 1448–1454. [Google Scholar] [CrossRef] [Green Version]
- Holling, T.M.; Schooten, E.; van Den Elsen, P.J. Function and Regulation of MHC Class II Molecules in T-Lymphocytes: Of Mice and Men. Hum. Immunol. 2004, 65, 282–290. [Google Scholar] [CrossRef]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites Produced by Commensal Bacteria Promote Peripheral Regulatory T-Cell Generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal Microbe-Derived Butyrate Induces the Differentiation of Colonic Regulatory T Cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-G.; Mathur, R.; Long, M.; Hosh, N.; Hao, L.; Hayden, M.S.; Ghosh, S. T Regulatory Cells Maintain Intestinal Homeostasis by Suppressing Γδ T Cells. Immunity 2010, 33, 791–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosovanu, C.; Neumann, C. The Many Functions of Foxp3+ Regulatory T Cells in the Intestine. Front. Immunol. 2020, 11, 600973. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, W.; Zenewicz, L.A.; Flavell, R.A. The Dual Nature of TH 17 Cells: Shifting the Focus to Function. Nat. Immunol. 2010, 11, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Bank, I. The Role of Gamma Delta T Cells in Autoimmune Rheumatic Diseases. Cells 2020, 9, 462. [Google Scholar] [CrossRef] [Green Version]
- Vantourout, P.; Hayday, A. Six-of-the-Best: Unique Contributions of Γδ T Cells to Immunology. Nat. Rev. Immunol. 2013, 13, 88–100. [Google Scholar] [CrossRef] [Green Version]
- Carding, S.R.; Egan, P.J. Γδ T Cells: Functional Plasticity and Heterogeneity. Nat. Rev. Immunol. 2002, 2, 336–345. [Google Scholar] [CrossRef]
- Littman, D.R.; Rudensky, A.Y. Th17 and Regulatory T Cells in Mediating and Restraining Inflammation. Cell 2010, 140, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Atarashi, K.; Tanoue, T.; Ando, M.; Kamada, N.; Nagano, Y.; Narushima, S.; Suda, W.; Imaoka, A.; Setoyama, H.; Nagamori, T.; et al. Th17 Cell Induction by Adhesion of Microbes to Intestinal Epithelial Cells. Cell 2015, 163, 367–380. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, I.I.; Atarashi, K.; Manel, N.; Brodie, E.L.; Shima, T.; Karaoz, U.; Wei, D.; Goldfarb, K.C.; Santee, C.A.; Lynch, S.V.; et al. Induction of Intestinal Th17 Cells by Segmented Filamentous Bacteria. Cell 2009, 139, 485–498. [Google Scholar] [CrossRef] [Green Version]
- Chung, H.; Pamp, S.J.; Hill, J.A.; Surana, N.K.; Edelman, S.M.; Troy, E.B.; Reading, N.C.; Villablanca, E.J.; Wang, S.; Mora, J.R.; et al. Gut Immune Maturation Depends on Colonization with a Host-Specific Microbiota. Cell 2012, 149, 1578–1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muranski, P.; Restifo, N.P. Essentials of Th17 Cell Commitment and Plasticity. Blood 2013, 121, 2402–2414. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Chen, F.; Liu, Z.; Cong, Y. Microbiota-Specific Th17 Cells: Yin and Yang in Regulation of Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2016, 22, 1473–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Sun, M.; Wu, W.; Chen, F.; Huang, X.; Xiao, Y.; Yao, S.; Cong, Y. Short Chain Fatty Acids Differentially Regulate Differentiation and Function of Th1 and Th17 Cells in the Intestines. J. Immunol. 2017, 198, 16. [Google Scholar] [CrossRef]
- Dupraz, L.; Magniez, A.; Rolhion, N.; Richard, M.L.; Da Costa, G.; Touch, S.; Mayeur, C.; Planchais, J.; Agus, A.; Danne, C. Gut Microbiota-Derived Short-Chain Fatty Acids Regulate IL-17 Production by Mouse and Human Intestinal Γδ T Cells. Cell Rep. 2021, 36, 109332. [Google Scholar] [CrossRef]
- Duscha, A.; Gisevius, B.; Hirschberg, S.; Yissachar, N.; Stangl, G.I.; Eilers, E.; Bader, V.; Haase, S.; Kaisler, J.; David, C. Propionic Acid Shapes the Multiple Sclerosis Disease Course by an Immunomodulatory Mechanism. Cell 2020, 180, 1067–1080. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Pattaroni, C.; Lopez-Mejia, I.C.; Riva, E.; Pernot, J.; Ubags, N.; Fajas, L.; Nicod, L.P.; Marsland, B.J. Dietary Fiber Confers Protection against Flu by Shaping Ly6c− Patrolling Monocyte Hematopoiesis and CD8+ T Cell Metabolism. Immunity 2018, 48, 992–1005. [Google Scholar] [CrossRef] [Green Version]
- Bachem, A.; Makhlouf, C.; Binger, K.J.; de Souza, D.P.; Tull, D.; Hochheiser, K.; Whitney, P.G.; Fernandez-Ruiz, D.; Dähling, S.; Kastenmüller, W. Microbiota-Derived Short-Chain Fatty Acids Promote the Memory Potential of Antigen-Activated CD8+ T Cells. Immunity 2019, 51, 285–297. [Google Scholar] [CrossRef]
- Donohoe, D.R.; Holley, D.; Collins, L.B.; Montgomery, S.A.; Whitmore, A.C.; Hillhouse, A.; Curry, K.P.; Renner, S.W.; Greenwalt, A.; Ryan, E.P. A Gnotobiotic Mouse Model Demonstrates That Dietary Fiber Protects against Colorectal Tumorigenesis in a Microbiota-and Butyrate-Dependent Manner. Cancer Discov. 2014, 4, 1387–1397. [Google Scholar] [CrossRef] [Green Version]
- Williams, E.A.; Coxhead, J.M.; Mathers, J.C. Anti-Cancer Effects of Butyrate: Use of Micro-Array Technology to Investigate Mechanisms. Proc. Nutr. Soc. 2003, 62, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, J.; Villa, M.; Sanin, D.E.; Buck, M.D.; O’Sullivan, D.; Ching, R.; Matsushita, M.; Grzes, K.M.; Winkler, F.; Chang, C.-H. Acetate Promotes T Cell Effector Function during Glucose Restriction. Cell Rep. 2019, 27, 2063–2074. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Fu, L.; Li, Y.; Wang, W.; Gong, M.; Zhang, J.; Dong, X.; Huang, J.; Wang, Q.; Mackay, C.R. Gut Microbial Metabolites Facilitate Anticancer Therapy Efficacy by Modulating Cytotoxic CD8+ T Cell Immunity. Cell Metab. 2021, 33, 988–1000. [Google Scholar] [CrossRef]
- Luu, M.; Riester, Z.; Baldrich, A.; Reichardt, N.; Yuille, S.; Busetti, A.; Klein, M.; Wempe, A.; Leister, H.; Raifer, H. Microbial Short-Chain Fatty Acids Modulate CD8+ T Cell Responses and Improve Adoptive Immunotherapy for Cancer. Nat. Commun. 2021, 12, 4077. [Google Scholar] [CrossRef]
- Park, E.M.; Chelvanambi, M.; Bhutiani, N.; Kroemer, G.; Zitvogel, L.; Wargo, J.A. Targeting the Gut and Tumor Microbiota in Cancer. Nat. Med. 2022, 28, 690–703. [Google Scholar] [CrossRef]
- Qi Hui, S.; Hua, L.; Wen Shan, Y.; Zhaohong, T.; Sharada, R.; Matthew Wook, C.; Louis Yi Ann, C. The Divergent Immunomodulatory Effects of Short Chain Fatty Acids and Medium Chain Fatty Acids. Int. J. Mol. Sci. 2021, 22, 6453. [Google Scholar] [CrossRef]
- Coutzac, C.; Jouniaux, J.-M.; Paci, A.; Schmidt, J.; Mallardo, D.; Seck, A.; Asvatourian, V.; Cassard, L.; Saulnier, P.; Lacroix, L.; et al. Systemic Short Chain Fatty Acids Limit Antitumor Effect of CTLA-4 Blockade in Hosts with Cancer. Nat. Commun. 2020, 11, 2168. [Google Scholar] [CrossRef]
- Cavaglieri, C.R.; Nishiyama, A.; Fernandes, L.C.; Curi, R.; Miles, E.A.; Calder, P.C. Differential Effects of Short-Chain Fatty Acids on Proliferation and Production of pro- and Anti-Inflammatory Cytokines by Cultured Lymphocytes. Life Sci. 2003, 73, 1683–1690. [Google Scholar] [CrossRef]
- Magnusson, M.K.; Vidal, A.; Maasfeh, L.; Isaksson, S.; Malhotra, R.; Olsson, H.K.; Öhman, L. Impaired Butyrate Induced Regulation of T Cell Surface Expression of CTLA-4 in Patients with Ulcerative Colitis. Int. J. Mol. Sci. 2021, 22, 3084. [Google Scholar] [CrossRef]
- Haghikia, A.; Jörg, S.; Duscha, A.; Berg, J.; Manzel, A.; Waschbisch, A.; Hammer, A.; Lee, D.-H.; May, C.; Wilck, N. Dietary Fatty Acids Directly Impact Central Nervous System Autoimmunity via the Small Intestine. Immunity 2015, 43, 817–829. [Google Scholar] [CrossRef] [Green Version]
- Kespohl, M.; Vachharajani, N.; Luu, M.; Harb, H.; Pautz, S.; Wolff, S.; Sillner, N.; Walker, A.; Schmitt-Kopplin, P.; Boettger, T.; et al. The Microbial Metabolite Butyrate Induces Expression of Th1-Associated Factors in CD4+ T Cells. Front. Immunol. 2017, 8, 1036. [Google Scholar] [CrossRef]
- Dalile, B.; Vervliet, B.; Bergonzelli, G.; Verbeke, K.; Van Oudenhove, L. Colon-Delivered Short-Chain Fatty Acids Attenuate the Cortisol Response to Psychosocial Stress in Healthy Men: A Randomized, Placebo-Controlled Trial. Neuropsychopharmacology 2020, 45, 2257–2266. [Google Scholar] [CrossRef] [PubMed]
- Hemmer, B.; Kerschensteiner, M.; Korn, T. Role of the Innate and Adaptive Immune Responses in the Course of Multiple Sclerosis. Lancet Neurol. 2015, 14, 406–419. [Google Scholar] [CrossRef]
- Park, J.; Wang, Q.; Wu, Q.; Mao-Draayer, Y.; Kim, C.H. Bidirectional Regulatory Potentials of Short-Chain Fatty Acids and Their G-Protein-Coupled Receptors in Autoimmune Neuroinflammation. Sci. Rep. 2019, 9, 8837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trend, S.; Leffler, J.; Jones, A.P.; Cha, L.; Gorman, S.; Brown, D.A.; Breit, S.N.; Kermode, A.G.; French, M.A.; Ward, N.C.; et al. Associations of Serum Short-Chain Fatty Acids with Circulating Immune Cells and Serum Biomarkers in Patients with Multiple Sclerosis. Sci. Rep. 2021, 11, 5244. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.; Abuazab, M.; Schwiertz, A.; Walter, S.; Faßbender, K.C.; Fousse, M.; Unger, M.M. Short-Chain Fatty Acids and Intestinal Inflammation in Multiple Sclerosis: Modulation of Female Susceptibility by Microbial Products? Autoimmun. Highlights 2021, 12, 7. [Google Scholar] [CrossRef]
- Pérez-Pérez, S.; Domínguez-Mozo, M.I.; Alonso-Gómez, A.; Medina, S.; Villarrubia, N.; Fernández-Velasco, J.I.; García-Martínez, M.Á.; García-Calvo, E.; Estévez, H.; Costa-Frossard, L.; et al. Acetate Correlates with Disability and Immune Response in Multiple Sclerosis. PeerJ 2020, 8, e10220. [Google Scholar] [CrossRef]
- Zeng, Q.; Gong, J.; Liu, X.; Chen, C.; Sun, X.; Li, H.; Zhou, Y.; Cui, C.; Wang, Y.; Yang, Y.; et al. Gut Dysbiosis and Lack of Short Chain Fatty Acids in a Chinese Cohort of Patients with Multiple Sclerosis. Neurochem. Int. 2019, 129, 104468. [Google Scholar] [CrossRef]
- Huang, J.; Pearson, J.A.; Peng, J.; Hu, Y.; Sha, S.; Xing, Y.; Huang, G.; Li, X.; Hu, F.; Xie, Z.; et al. Gut Microbial Metabolites Alter IgA Immunity in Type 1 Diabetes. JCI Insight 2020, 5, e135718. [Google Scholar] [CrossRef]
- Winther, S.A.; Mannerla, M.M.; Frimodt-Møller, M.; Persson, F.; Hansen, T.W.; Lehto, M.; Hörkkö, S.; Blaut, M.; Forsblom, C.; Groop, P.-H.; et al. Faecal Biomarkers in Type 1 Diabetes with and without Diabetic Nephropathy. Sci. Rep. 2021, 11, 15208. [Google Scholar] [CrossRef]
- Martinsson, K.; Dürholz, K.; Schett, G.; Zaiss, M.M.; Kastbom, A. Higher Serum Levels of Short-Chain Fatty Acids Are Associated with Non-Progression to Arthritis in Individuals at Increased Risk of RA. Ann. Rheum. Dis. 2022, 81, 445–447. [Google Scholar] [CrossRef]
- Rosser, E.C.; Piper, C.J.M.; Matei, D.E.; Blair, P.A.; Rendeiro, A.F.; Orford, M.; Alber, D.G.; Krausgruber, T.; Catalan, D.; Klein, N. Microbiota-Derived Metabolites Suppress Arthritis by Amplifying Aryl-Hydrocarbon Receptor Activation in Regulatory B Cells. Cell Metab. 2020, 31, 837–851. [Google Scholar] [CrossRef]
- Yao, Y.; Cai, X.; Zheng, Y.; Zhang, M.; Fei, W.; Sun, D.; Zhao, M.; Ye, Y.; Zheng, C. Short-chain Fatty Acids Regulate B Cells Differentiation via FFAR2 to Alleviate Rheumatoid Arthritis. Br. J. Pharmacol. 2022, 1–15. [Google Scholar] [CrossRef]
- Dürholz, K.; Hofmann, J.; Iljazovic, A.; Häger, J.; Lucas, S.; Sarter, K.; Strowig, T.; Bang, H.; Rech, J.; Schett, G. Dietary Short-Term Fiber Interventions in Arthritis Patients Increase Systemic SCFA Levels and Regulate Inflammation. Nutrients 2020, 12, 3207. [Google Scholar] [CrossRef]
- Häger, J.; Bang, H.; Hagen, M.; Frech, M.; Träger, P.; Sokolova, M.V.; Steffen, U.; Tascilar, K.; Sarter, K.; Schett, G. The Role of Dietary Fiber in Rheumatoid Arthritis Patients: A Feasibility Study. Nutrients 2019, 11, 2392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haase, S.; Mäurer, J.; Duscha, A.; Lee, D.-H.; Balogh, A.; Gold, R.; Müller, D.N.; Haghikia, A.; Linker, R.A. Propionic Acid Rescues High-Fat Diet Enhanced Immunopathology in Autoimmunity via Effects on Th17 Responses. Front. Immunol. 2021, 12, 701626. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef] [PubMed]
- Burrack, A.L.; Martinov, T.; Fife, B.T. T Cell-Mediated Beta Cell Destruction: Autoimmunity and Alloimmunity in the Context of Type 1 Diabetes. Front. Endocrinol. 2017, 8, 343. [Google Scholar] [CrossRef] [PubMed]
- Mariño, E.; Richards, J.L.; McLeod, K.H.; Stanley, D.; Yap, Y.A.; Knight, J.; McKenzie, C.; Kranich, J.; Oliveira, A.C.; Rossello, F.J. Gut Microbial Metabolites Limit the Frequency of Autoimmune T Cells and Protect against Type 1 Diabetes. Nat. Immunol. 2017, 18, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Bell, K.J.; Saad, S.; Tillett, B.J.; McGuire, H.M.; Bordbar, S.; Yap, Y.A.; Nguyen, L.T.; Wilkins, M.R.; Corley, S.; Brodie, S. SCFA Supplementation Is Associated with Microbiota and Immune Modulation in Diabetes. Res. Sq. 2022, 10, 9. [Google Scholar] [CrossRef]
- de Groot, P.F.; Nikolic, T.; Imangaliyev, S.; Bekkering, S.; Duinkerken, G.; Keij, F.M.; Herrema, H.; Winkelmeijer, M.; Kroon, J.; Levin, E. Oral Butyrate Does Not Affect Innate Immunity and Islet Autoimmunity in Individuals with Longstanding Type 1 Diabetes: A Randomised Controlled Trial. Diabetologia 2020, 63, 597–610. [Google Scholar] [CrossRef]
- Ismail, H.M.; Evans-Molina, C.; DiMeglio, L.A. 57437 Effects of Prebiotics on the Gut Microbiome Profile, Beta-Cell Function and Immune Markers in Newly-Diagnosed Type 1 Diabetes. J. Clin. Transl. Sci. 2021, 5, 38. [Google Scholar] [CrossRef]
- Kamali, A.N.; Noorbakhsh, S.M.; Hamedifar, H.; Jadidi-Niaragh, F.; Yazdani, R.; Bautista, J.M.; Azizi, G. A Role for Th1-like Th17 Cells in the Pathogenesis of Inflammatory and Autoimmune Disorders. Mol. Immunol. 2019, 105, 107–115. [Google Scholar] [CrossRef]
- Petrelli, A.; van Wijk, F. CD8+ T Cells in Human Autoimmune Arthritis: The Unusual Suspects. Nat. Rev. Rheumatol. 2016, 12, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Weavers, H.; Martin, P. The Cell Biology of Inflammation: From Common Traits to Remarkable Immunological Adaptations. J. Cell Biol. 2020, 219. [Google Scholar] [CrossRef]
- Takahashi, D.; Hoshina, N.; Kabumoto, Y.; Maeda, Y.; Suzuki, A.; Tanabe, H.; Isobe, J.; Yamada, T.; Muroi, K.; Yanagisawa, Y. Microbiota-Derived Butyrate Limits the Autoimmune Response by Promoting the Differentiation of Follicular Regulatory T Cells. EBioMedicine 2020, 58, 102913. [Google Scholar] [CrossRef] [PubMed]
- Lucas, S.; Omata, Y.; Hofmann, J.; Böttcher, M.; Iljazovic, A.; Sarter, K.; Albrecht, O.; Schulz, O.; Krishnacoumar, B.; Krönke, G. Short-Chain Fatty Acids Regulate Systemic Bone Mass and Protect from Pathological Bone Loss. Nat. Commun. 2018, 9, 55. [Google Scholar] [CrossRef] [Green Version]
- Wallace, K.L.; Zheng, L.-B.; Kanazawa, Y.; Shih, D.Q. Immunopathology of Inflammatory Bowel Disease. World J. Gastroenterol. WJG 2014, 20, 6. [Google Scholar] [CrossRef]
- Xu, H.-M.; Zhao, H.-L.; Guo, G.-J.; Xu, J.; Zhou, Y.-L.; Huang, H.-L.; Nie, Y.-Q. Characterization of Short-Chain Fatty Acids in Patients with Ulcerative Colitis: A Meta-Analysis. BMC Gastroenterol. 2022, 22, 117. [Google Scholar] [CrossRef] [PubMed]
- Scheppach, W. Treatment of Distal Ulcerative Colitis with Short-Chain Fatty Acid Enemas a Placebo-Controlled Trial. Dig. Dis. Sci. 1996, 41, 2254–2259. [Google Scholar] [CrossRef]
- Vernia, P.; Cittadini, M.; Caprilli, R.; Torsoli, A. Topical Treatment of Refractory Distal Ulcerative Colitis with 5-ASA and Sodium Butyrate. Dig. Dis. Sci. 1995, 40, 305–307. [Google Scholar] [CrossRef]
- Steinhart, A.H.; Hiruki, T.; Brzeznski, A.; Baker, J.P. Treatment of Left-Sided Ulcerative Colitis with Butyrate Enemas: A Controlled Trial. Aliment. Pharmacol. Ther. 1996, 10, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Breuer, R.I.; Soergel, K.H.; Lashner, B.A.; Christ, M.L.; Hanauer, S.B.; Vanagunas, A.; Harig, J.M.; Keshavarzian, A.; Robinson, M.; Sellin, J.H.; et al. Short Chain Fatty Acid Rectal Irrigation for Left-Sided Ulcerative Colitis: A Randomised, Placebo Controlled Trial. Gut 1997, 40, 485–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamka, M.; Kokot, M.; Kaczmarek, N.; Bermagambetova, S.; Nowak, J.K.; Walkowiak, J. The Effect of Sodium Butyrate Enemas Compared with Placebo on Disease Activity, Endoscopic Scores, and Histological and Inflammatory Parameters in Inflammatory Bowel Diseases: A Systematic Review of Randomised Controlled Trials. Complement. Med. Res. 2021, 28, 344–356. [Google Scholar] [CrossRef]
- Berndt, B.E.; Zhang, M.; Owyang, S.Y.; Cole, T.S.; Wang, T.W.; Luther, J.; Veniaminova, N.A.; Merchant, J.L.; Chen, C.-C.; Huffnagle, G.B. Butyrate Increases IL-23 Production by Stimulated Dendritic Cells. Am. J. Physiol. Liver Physiol. 2012, 303, G1384–G1392. [Google Scholar] [CrossRef] [Green Version]
- Trapecar, M.; Communal, C.; Velazquez, J.; Maass, C.A.; Huang, Y.-J.; Schneider, K.; Wright, C.W.; Butty, V.; Eng, G.; Yilmaz, O.; et al. Gut-Liver Physiomimetics Reveal Paradoxical Modulation of IBD-Related Inflammation by Short-Chain Fatty Acids. Cell Syst. 2020, 10, 223–239. [Google Scholar] [CrossRef]
- Lee, J.G.; Lee, J.; Lee, A.; Jo, S.V.; Park, C.H.; Han, D.S.; Eun, C.S. Impact of Short-Chain Fatty Acid Supplementation on Gut Inflammation and Microbiota Composition in a Murine Colitis Model. J. Nutr. Biochem. 2022, 101, 108926. [Google Scholar] [CrossRef]
- Sun, M.; Wu, W.; Chen, L.; Yang, W.; Huang, X.; Ma, C. Microbiota-Derived Short-Chain Fatty Acids Promote Th1 Cell IL-10 Production to Maintain Intestinal Homeostasis. Nat Commun. 2018, 9, 3555. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Goergen, C.J.; HogenEsch, H.; Kim, C.H. Chronically Elevated Levels of Short-Chain Fatty Acids Induce T Cell–Mediated Ureteritis and Hydronephrosis. J. Immunol. 2016, 196, 2388–2400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzocco, S.; Fazeli, G.; Di Micco, L.; Autore, G.; Adesso, S.; Dal Piaz, F.; Heidland, A.; Di Iorio, B. Supplementation of Short-Chain Fatty Acid, Sodium Propionate, in Patients on Maintenance Hemodialysis: Beneficial Effects on Inflammatory Parameters and Gut-Derived Uremic Toxins, a Pilot Study (PLAN Study). J. Clin. Med. 2018, 7, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, L.W.; Sun, X.N.; Ma, Z.C.; Fu, J.; Liu, F.J.; Huang, B.L.; Liang, D.C.; Sun, D.M.; Lan, C. Increased Vδ1γδT Cells Predominantly Contributed to IL-17 Production in the Development of Adult Human Post-Infectious Irritable Bowel Syndrome. BMC Gastroenterol. 2021, 21, 271. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, M.K.; Isaksson, S.; Öhman, L. The Anti-Inflammatory Immune Regulation Induced by Butyrate Is Impaired in Inflamed Intestinal Mucosa from Patients with Ulcerative Colitis. Inflammation 2020, 43, 507–517. [Google Scholar] [CrossRef] [Green Version]
- Meyer, F.; Seibert, F.S.; Nienen, M.; Welzel, M.; Beisser, D.; Bauer, F.; Rohn, B.; Westhoff, T.H.; Stervbo, U.; Babel, N. Propionate Supplementation Promotes the Expansion of Peripheral Regulatory T-Cells in Patients with End-Stage Renal Disease. J. Nephrol. 2020, 33, 817–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
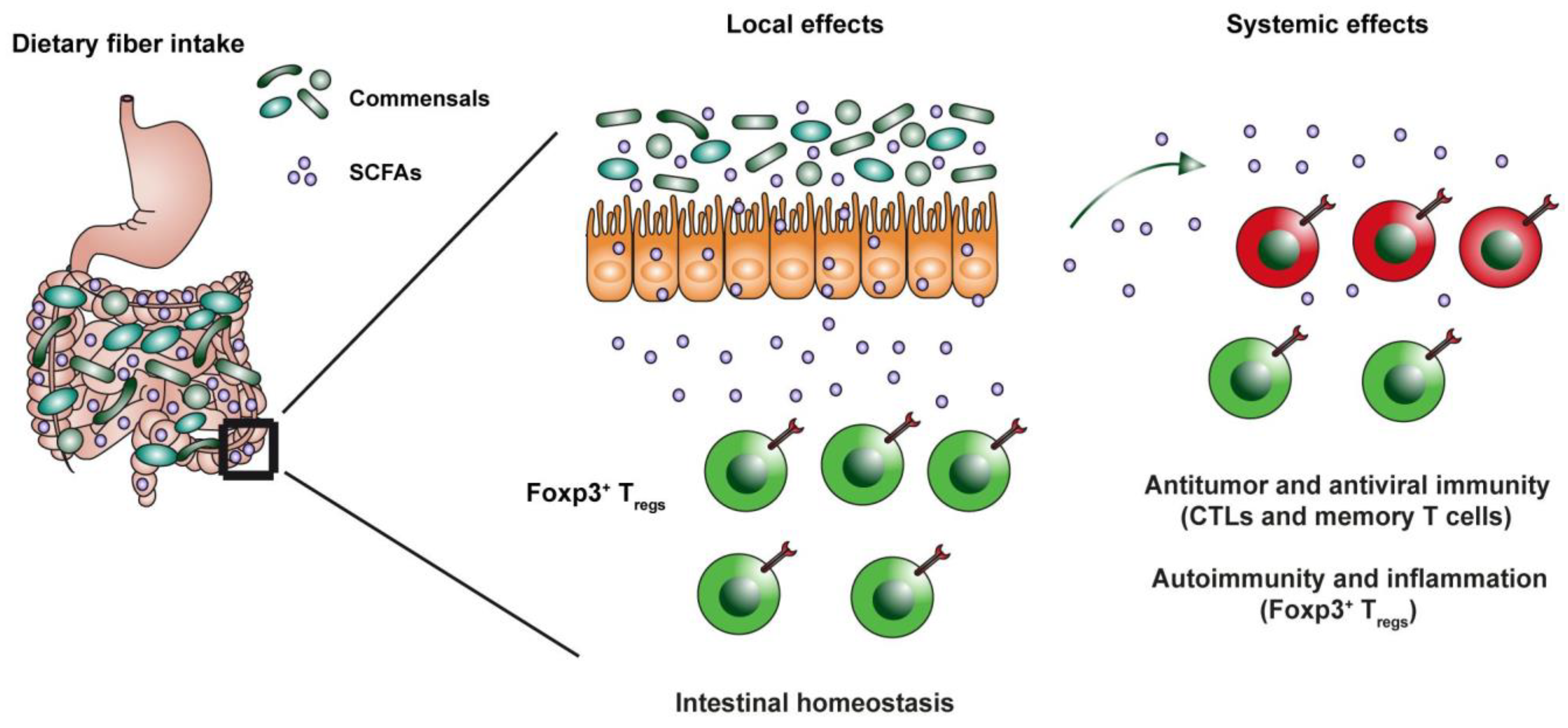

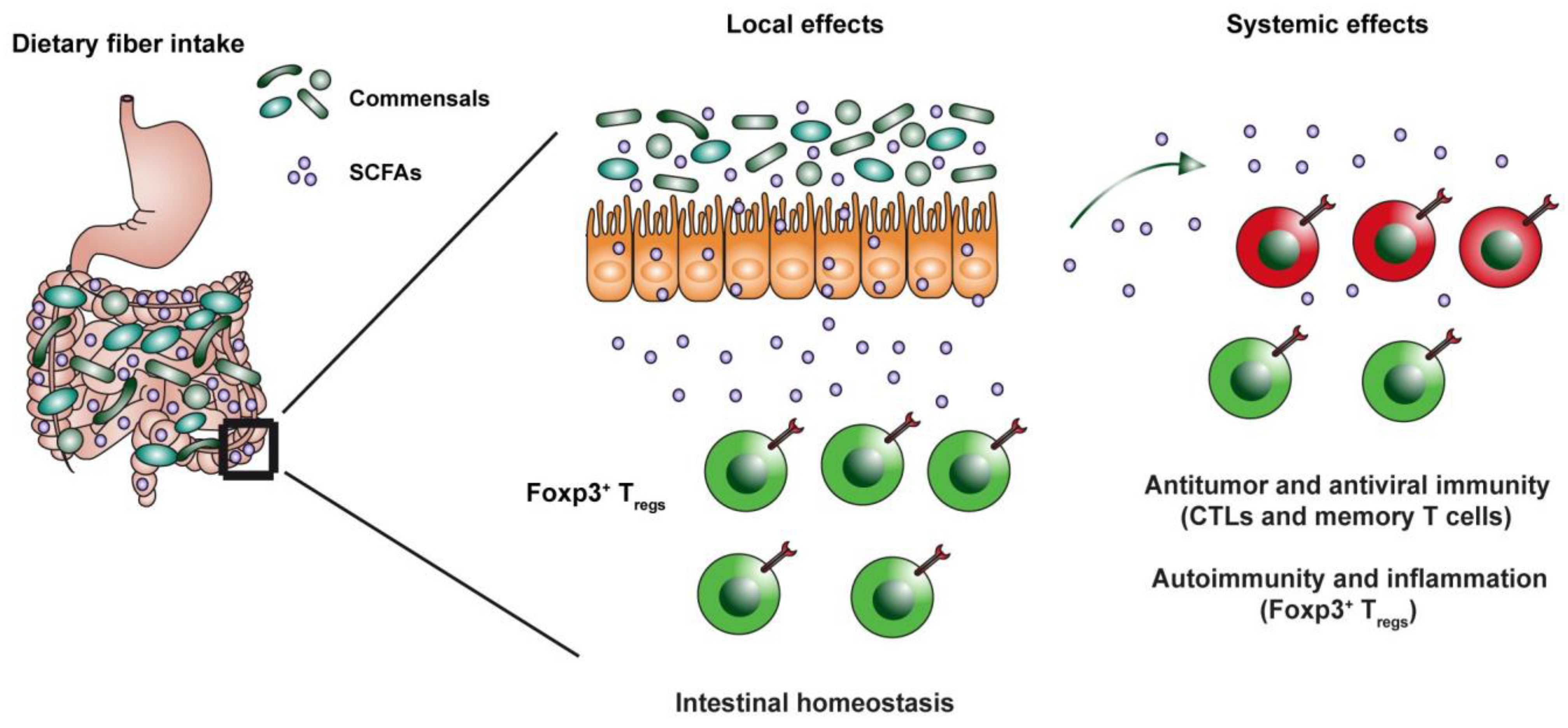{kind=link}
| SCFA | Colon (Caecum) | Blood (Serum/Plasma, Fasted) | References | |
|---|---|---|---|---|
| Healthy Humans | Total SCFA | 131 mmol/kg | ~79–510 μmol/L | [8,17] |
| Acetate | 44–69 mmol/kg | ~5–402 μmol/L | [8,9,17] | |
| Propionate | 14–25 mmol/kg | 3.3–ca.15 μmol/L | [9,17,18] | |
| Butyrate | 15–26 mmol/kg | 2.1–10 μmol/L | [9,18] | |
| Valerate | ~4.5 mmol/kg | 1.3–ca 4 μmol/L | [8,17,18] |
| Author | Experiment | Cells Used for Assay | SCFA Used | Dose Used | Effect For | No Effect For | Outcome |
|---|---|---|---|---|---|---|---|
| [48] | Lymphocyte proliferation assay, cytokine production | rat lymphocytes (from lymphnodes) | acetate, butyrate, propionate | acetate 10 mM, butyrate 1.5 mM, propionate 2 mM | Butyrate at 0.25 mM–1.5 mM | acetate, propionate (Il-2) | ↓ Thymidine incorporation ↑ IL-10 production after 48 h for acetate, propionate or a combination of both/with butyrate; ↓ IL-10 after 24 h for butyrate alone buyrate inhibited IL-2/IFN- γ production |
| [22] | Induction of Treg cells in vitro | splenic naïve (CD44lo CD62Lhi) CD4+ T cells | (Sodium) acetate, butyrate propionate | 0.1 mM | Butyrate, propionate | acetate | ↑ Tregs for butyrate, moderately for propionate, no effect for acetate |
| [21] | Induction of extrathymic Tregs in vitro | Peripheral naïve (CD44loCD62LhiCD25−) CD4+ T cells | Butyrate, isovalerate, acetate, propionate | 0–1024 μM | Butyrate, isovalerate, propionate | acetate | ↑ FOXP3+ cells |
| [6] | Purified cTregs from GF mice cultured for 24 h | cTregs from GF mice | propionate | 0.1 mM | propionate | -- | ↑ IL-10 expression; ↑ IL-10 protein; ↑ FOXP3 expression, no effect for TGF-β |
| [7] | Effect of SCFAs under; anti-CD3/CD28 in a Th17 or Th1 condition | naive CD4+ T cells | acetate, propionate, butyrate | acetate: 0–20 mM propionate: 0–1 mM | Optimal: acetate 5–20 mM; propionate: 0.5–1 mM | acetate (<1 mM), propionate (<0.5 mM) | ↑ Th1 and Th17 differentiation ↑ expression of IL-17A, IL-17F, RORα, RORγt, T-bet, and IFN-γ ↑ FOXP3 in low anti-CD3 (1 μg mL−1) activation condition ↑ high IL-10 expression ↑ IL-10, IFN-γ, and IL-17 in CD8+ T cells |
| [49] | peripheral blood mononuclear cells (PBMCs)/ | Human PBMCS | butyrate, propionate, acetate | Propionate+ acetate: 0–6.4 mM; butyrate: 0–1.6 mM | butyrate, propionate | acetate | ↓ expression of CD25 in CD4+ and CD8+ cells, butyrate (0.1–0.4 mM) ↑ CTLA-4 |
| [50] | Treg & Th17 polarizing conditions | human naive CD4+ T cells (CD45RA−CD45RO+CD25−CD127+) | propionate | 1 μM–10 mM | 1 μM–1 mM (Treg), 150 μM Th17 | 10mM propionate | ↑ CD4+CD25+Foxp3+ cells; ↑ proliferation of differentiated Tregs; ↓ CD4+IL-17A+ T cells |
| [47] | CD3 stimulation | Human PBMCS | butyrate | 0–100 μM, | 50 μM and 100 μM | 10 μM butyrate | ↑ % of Tregs in PBMCs |
| [51] | CD3 stimulation/Treg polarizing conditions | Murine CD4+ cells from lymph nodes/spleen | butyrate, propionate, acetate | 0.1–10 mM | Increased regulatory expression: 0.1–0.25 mM; supressed regulation: 0.5 mM–1 mM | acetate, butyrate >0.5 mM induced IFN- γ but not Tregs | ≤0.25 mM: ↑ FOXP3 and inducible Treg differentiation only when TGF-ß was suboptimal; absence or presence of high TGF-ß abolished effects of butyrate; >0.25 mM: abolished effect of Treg differentiation, induction of IFN- γ,(not IL-17A or IL-4) |
| Reference | Groups | SCFA Studied | SCFA Measured In | Concentration in Patient Group | Findings |
|---|---|---|---|---|---|
| Multiple Sclerosis | |||||
| [55] | 30 MS/CIS 10 HC | acetate, butyrate, propionate | blood | concentrations in nmol/mL: acetate: ~1600; butyrate: ~14; propionate: ~160 | lower propionate in MS than controlsbutyrate positively correlates with Tfh cells no association of SCFAs with Treg, Th1 or Th17 counts |
| [17] | 58 (RR)MS 50 HC | acetate, propionate, butyrate, valerate | blood | concentrations in μmol/L: total SCFA 453; acetate 349; butyrate: ~6.4; propionate ~15 valerate: ~3.8 | lower total SCFAs concentration in MS compared to controls no individual difference for acetate butyrate, propionate or valerate between MS and controls after multiple testing correction no significant correlation of SCFAs with inflammatory cytokines in patients with MS positive association of butyrate with TNF and IFN-γ in HC, negative correlation of acetate and IFN-γ SCFA levels not related to clinical follow up at 12 months |
| [57] | 95 MS 54 HC | acetate | blood | approximate mean values in µM/L: acetate: ~25; propionate: ~3.9; butyrate: ~3.8 | higher acetate levels in blood of patients with MS; positive association with CD4+IL-17+ (not significant after correction) and CD8+ IL-17* cells, negative association with naive CD4+ cells |
| [58] | 34 MS/12 MS for cells 34 HC | propionate, butyrate, acetate | fecal samples | na | depletion of fecal acetate, propionate, and butyrate in MS; fecal SCFA level positively correlated with pTreg frequency |
| [56] | 41 MS/35 controls | propionate, butyrate, acetate | fecal samples | median concentrations in mmol/g: acetate 41.70; propionate 5.51/butyrate 1.25/valerate | non-significantly lower SCFA concentrations in MS compared to controls |
| Type 1 Diabetes | |||||
| [59] | 19 patients with T1D and matched controls+ animal model | acetate, butyrate | feces | total μg/g: acetate ~1700; butyrate: ~1600 | ·significantly lower SCFA concentrations (butyrate, propionate acetate) in T1D; valerate levels similar No assessment of T cells in patients; higher frequencies of CD4+FN-γ+ and CD8+TNF-α+ T cells in the mice colonized with fecal bacteria from patients with T1D vs. controls |
| [60] | 132 T1D; 40 HC | propionate, butyrate, acetate | feces | total µmol/g: acetate: 39; propionate: 9, butyrate: 7.8, valerate: 1.8 | propionate and butyrate significantly lower in T1D, valerate and acetate not different; higher hsCRP and neutrophil count, correlation with SCFAs not assessed |
| Rheumatoid arthritis | |||||
| [61] | 82 people with increased risk for RA | acetate, butyrate, propionate or pentanoate | blood | serum in uM: acetate: ~80; butyrate: ~3; pentanoate: ~4.5; Proprionate ~8 | higher total sum of SCFAs and/or butyrate/acetate in people who did not develop RA;·butyrate correlated negatively with serum IgA-anti-citrullinated protein antibody levels but not with IgG or IgM isotypes; T cells were not assessed |
| [62] | 19 RA, 20 HC stool/13 HC 14RA blood | acetate, butyrate, propionate | stool and blood | concentrations in stool, μmol/g: acetate: ~7; propionate ~2, butyrate ~1.8; in blood μmol/g: acetate: ~30, proprionate ~2, butyrate ~10 | stool: significant reduction in butyrate/propionate, no difference in acetate; serum: no difference in propionate/butyrate but significant increase in acetate; butyrate correlates positively with circulating IL-10+ B cells in serum. |
| [63] | 9HC 10RA | acetate, butyrate, propionate, valerate | stool | concentrations in umol/g: acetate ~9, propionate ~1.5, butyrate ~5; valproate ~0.19 | lower acetate, propionate, butyrate and valerate in patients compared to controls |
| [64,65] | 10 HC 36/29 RA | acetate, butyrate, propionate | stool and serum | in mM: in serum: total in mM: ~0.2, acetate: ~0.2, propionate: ~0.015, butyrate ~0.004 | no comparison between HC and RA patients regarding SCFAs, ·inflammatory cytokines were not higher at baseline, but patients received DMARD therapy |
| Reference | Disease | n Cases/Controls | Intervention (RCT/Animal/IV) | Intervention+Dose | Duration | Cell Type Studied | Measured SCFA Concentration Post Intervention | Outcome |
|---|---|---|---|---|---|---|---|---|
| [37] | MS | 91 MS, 24 HC | Short-term propionate | 1 g oral propionate daily | 14 days | Treg, Th17 | in CSF (n = 3) ↑ propionate after mean of 2.8 months | ↓ Th17, ↑ Treg count, ↑ metabolism and function after 14 days and 90 days; ↑ Tregs suppressive capacity ex vivo; ↓ IL-10 ex vivo cell culture of propionate treated patients, mixed results for IL-17 |
| MS | 52 MS | Long-term propionate | 1 g propionate daily/1–3 years | 1–3 years | na | na | improved clinical outcome (disability status and relapse rate) | |
| RA | 20 RA | Short-term propionate | 1 g propionate daily | 14/28 days | Treg | na | ↑ Tregs after 14 days | |
| [93] | end stage renal disease | 10 ESRD patients/7HC | Open-label intervention study propionate | 2 × 500 mg daily sodium propionate | 30 days | Treg, CD25+CD127- | na | ↑ Tregs over time, but not due to proliferation; ↓ after treatment with propionate; no consistent difference for granzyme, IFN-γ, IL-2, IL-17, and TNF-α producing CD4+ and CD8+T-cells |
| [35], sample from [55] | MS | 34 non obese MS (22 at fu); 6 obese MS (5 at fu) | propionate | 500 mg sodium-propionate (PA) capsules twice daily | 90 days | Treg, Th17 | na | ↑ Tregs in non-obese group, non-significant ↑ in obese group; ↓ Th17 in obese group, non-significant ↓ in non-obese group |
| [70] | T1D | 20 T1D | HAMSAB dietary supplement | 40 g/day HAMSAB | 6 weeks | T cells, B cells, monocytes | plasma: ↑ acetate after 3 weeks, maximum at 6 weeks: 58% subjects acetate ↑ >2-fold; similar but less strong observations for propionate + butyrate; | ↑ (naive)B cells at week 6; ↓ CD86 on B cells after washout period of 6 weeks; ↑ CD3+ cells after washout period week 12; ↑ CTL4A expression; ↓ plasma IL-8, MIP-1α, and bFGF at week 12 compared to baseline |
| [71] | T1D | 30 T1D | RCT with butyrate/placebo | 4 g sodium butyrate or placebo | 1 month | PBMCs (monocytes, autoreactive CD8+ T cells) | ← fecal acetate, ↓ fecal propionate and butyrate | ← CRP after butyrate treatment; ← monocyte subsets (compared to placebo) ← ex vivo cytokine production; ↓ islet autoreactive CD8 T cells (assay only possible in in 13/30); ← lymphocyte subsets |
| [64,65] | RA | 36 RA | dietary intervention high fiber bars | high-fiber bars 15–30 g/day or cereals | 28 days (14 + 14)/30 days | Tregs, Th1, Th17 | ↑ acetate and butyrate and propionate after 30 days | ↑ Tregs, ↑ Th1/Th17 ratio; ← CD4 and CD8 numbers. Improved physical & mental quality of life and disability scores, ↓ IgA; ← anti-citrullinated vimentin p18 peptide antibody levels; ← for IL-1β, IFN-a2, IFN-γ, TNF-α, IL-6, CXCL-8 (IL-8), IL-10, IL-12p70, IL-17A and IL-23; ↓ for CCL-2, IL-18 and IL-33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiweck, C.; Edwin Thanarajah, S.; Aichholzer, M.; Matura, S.; Reif, A.; Vrieze, E.; Weigert, A.; Visekruna, A. Regulation of CD4+ and CD8+ T Cell Biology by Short-Chain Fatty Acids and Its Relevance for Autoimmune Pathology. Int. J. Mol. Sci. 2022, 23, 8272. https://doi.org/10.3390/ijms23158272
Schiweck C, Edwin Thanarajah S, Aichholzer M, Matura S, Reif A, Vrieze E, Weigert A, Visekruna A. Regulation of CD4+ and CD8+ T Cell Biology by Short-Chain Fatty Acids and Its Relevance for Autoimmune Pathology. International Journal of Molecular Sciences. 2022; 23(15):8272. https://doi.org/10.3390/ijms23158272
Chicago/Turabian StyleSchiweck, Carmen, Sharmili Edwin Thanarajah, Mareike Aichholzer, Silke Matura, Andreas Reif, Elske Vrieze, Andreas Weigert, and Alexander Visekruna. 2022. "Regulation of CD4+ and CD8+ T Cell Biology by Short-Chain Fatty Acids and Its Relevance for Autoimmune Pathology" International Journal of Molecular Sciences 23, no. 15: 8272. https://doi.org/10.3390/ijms23158272
APA StyleSchiweck, C., Edwin Thanarajah, S., Aichholzer, M., Matura, S., Reif, A., Vrieze, E., Weigert, A., & Visekruna, A. (2022). Regulation of CD4+ and CD8+ T Cell Biology by Short-Chain Fatty Acids and Its Relevance for Autoimmune Pathology. International Journal of Molecular Sciences, 23(15), 8272. https://doi.org/10.3390/ijms23158272