
An Integrated Proteomic and Glycoproteomic Investigation Reveals Alterations in the N-Glycoproteomic Network Induced by 2-Deoxy-D-Glucose in Colorectal Cancer Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
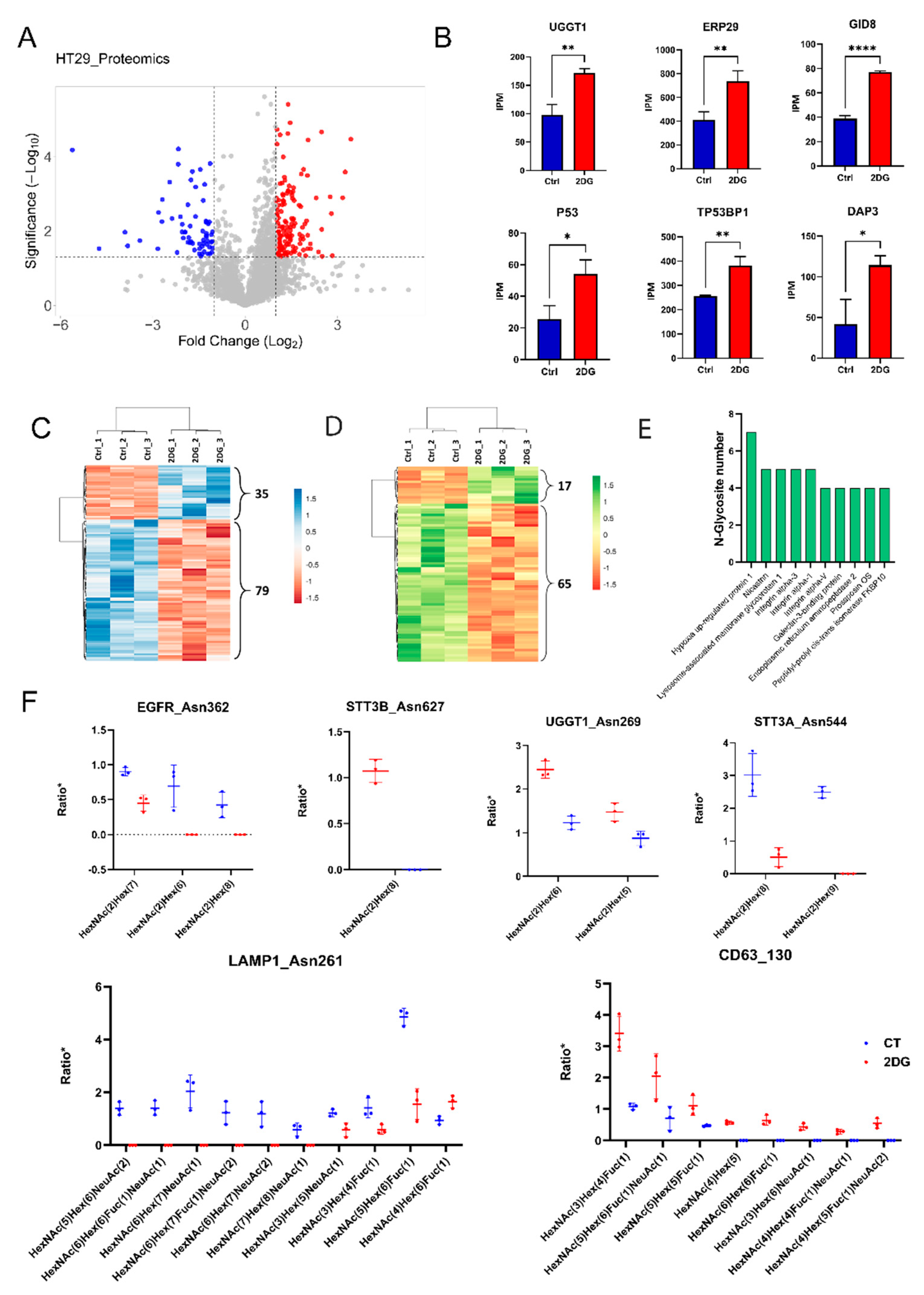
2.1. Proteomic and N-Glycoproteomic Analyses of the 2DG-Treated HT29 Cells
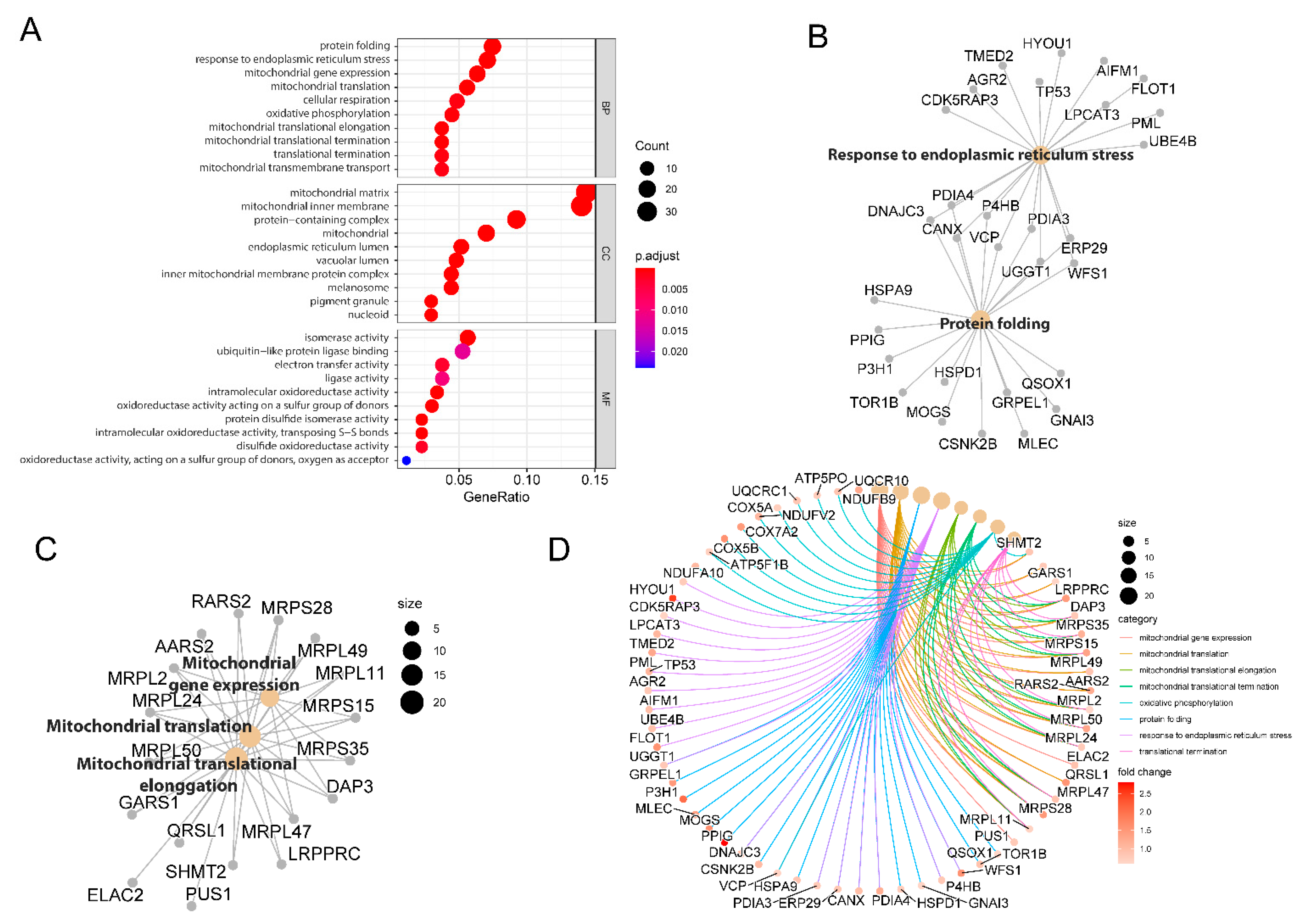
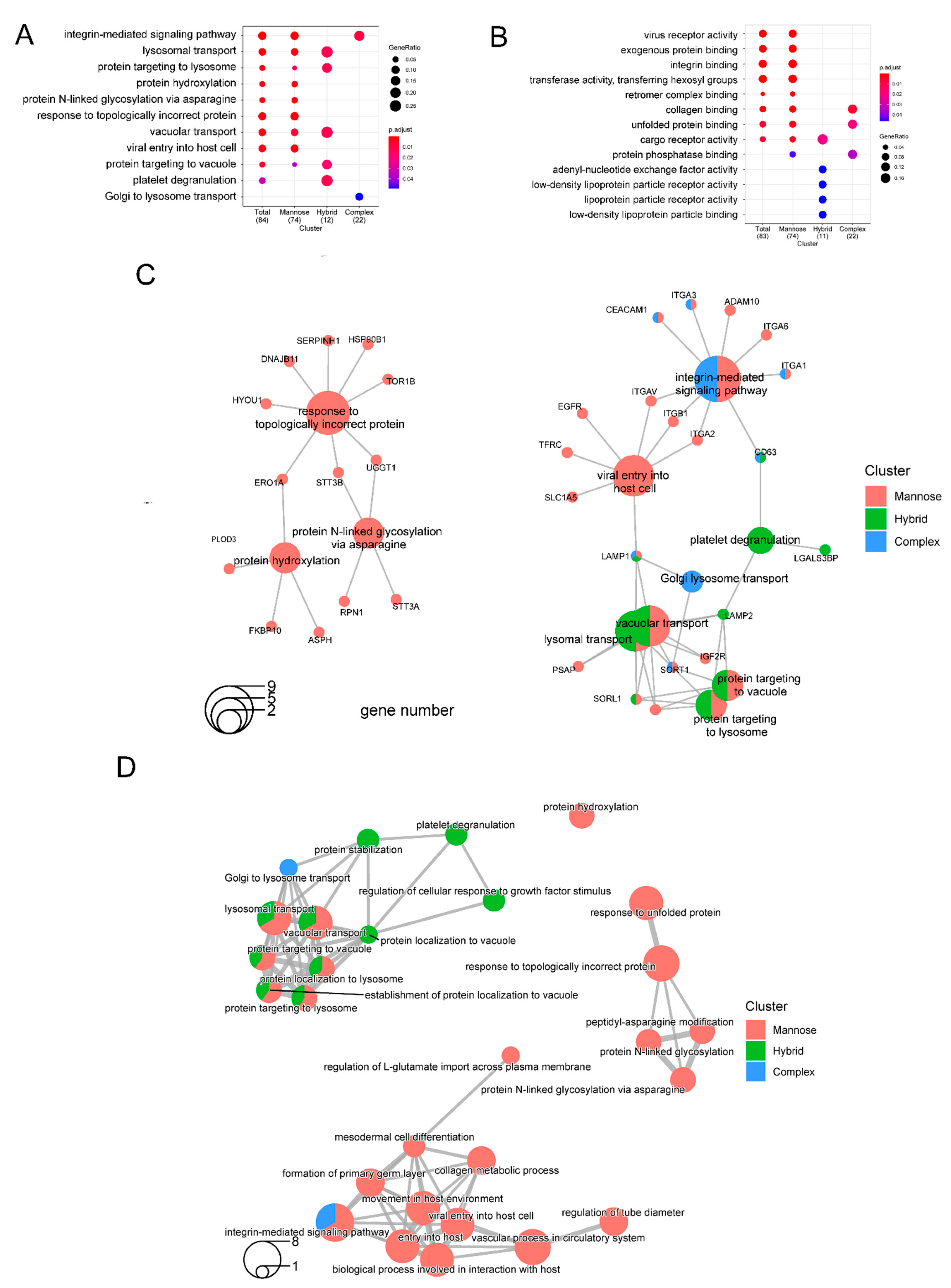
2.2. Functional Clustering of Dysregulated Proteins Induced by 2DG
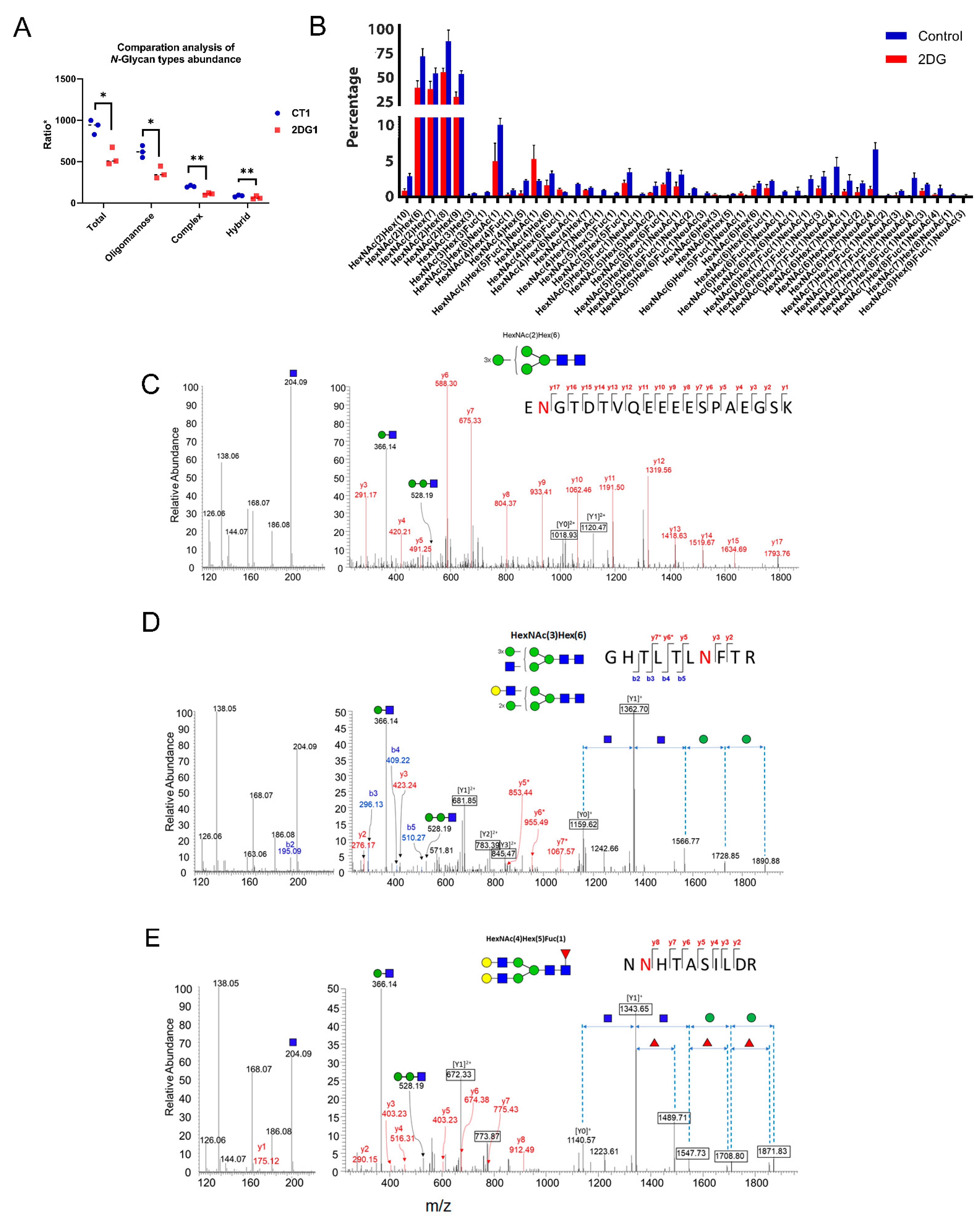
2.3. Altered N-Glycoforms with High-Mannose, Hybrid, and Complex Types
2.4. Biological Theme Comparison among Different N-Glycosylation Types
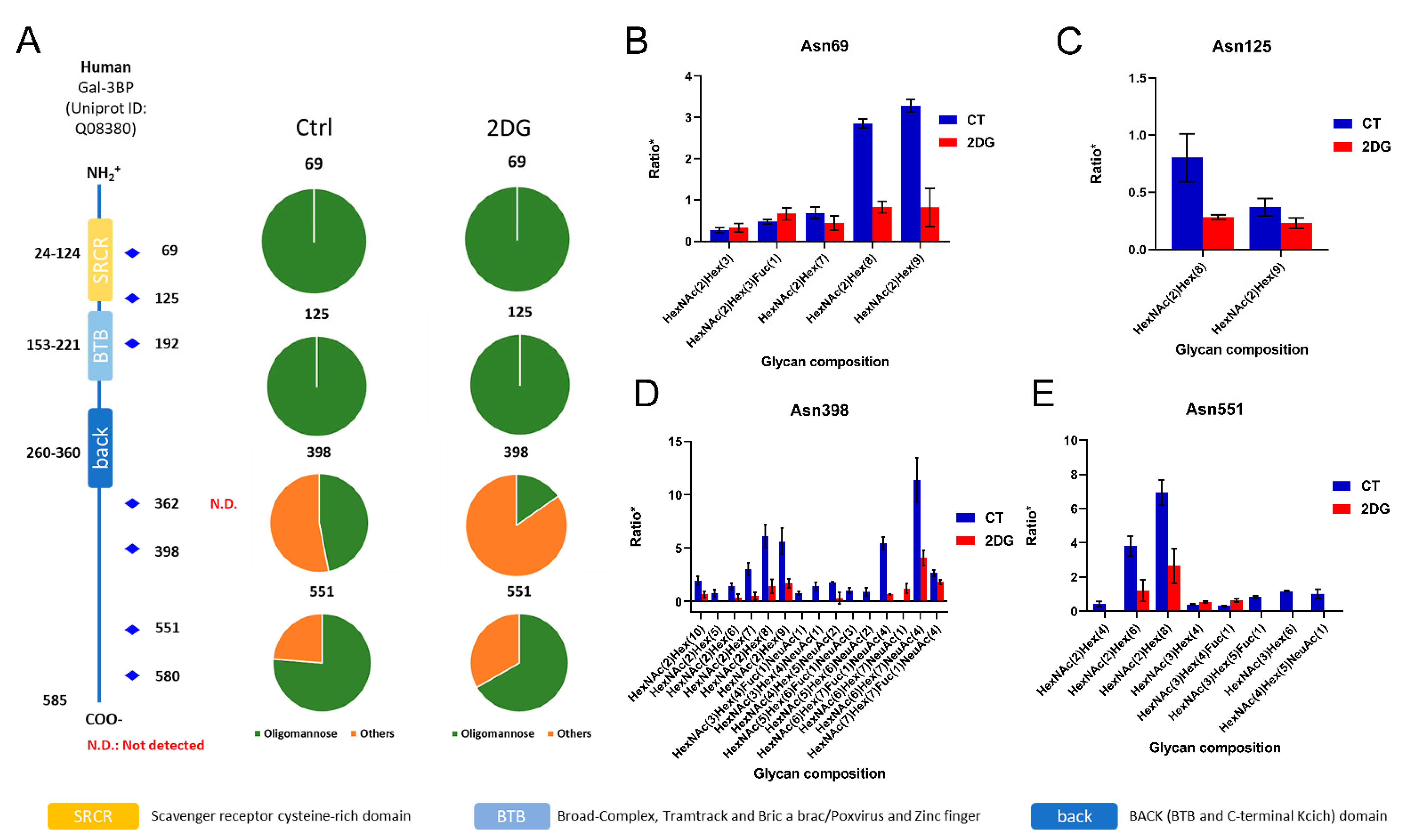
2.5. Interactive Influences of 2DG on Global Proteome and N-Glycoproteome
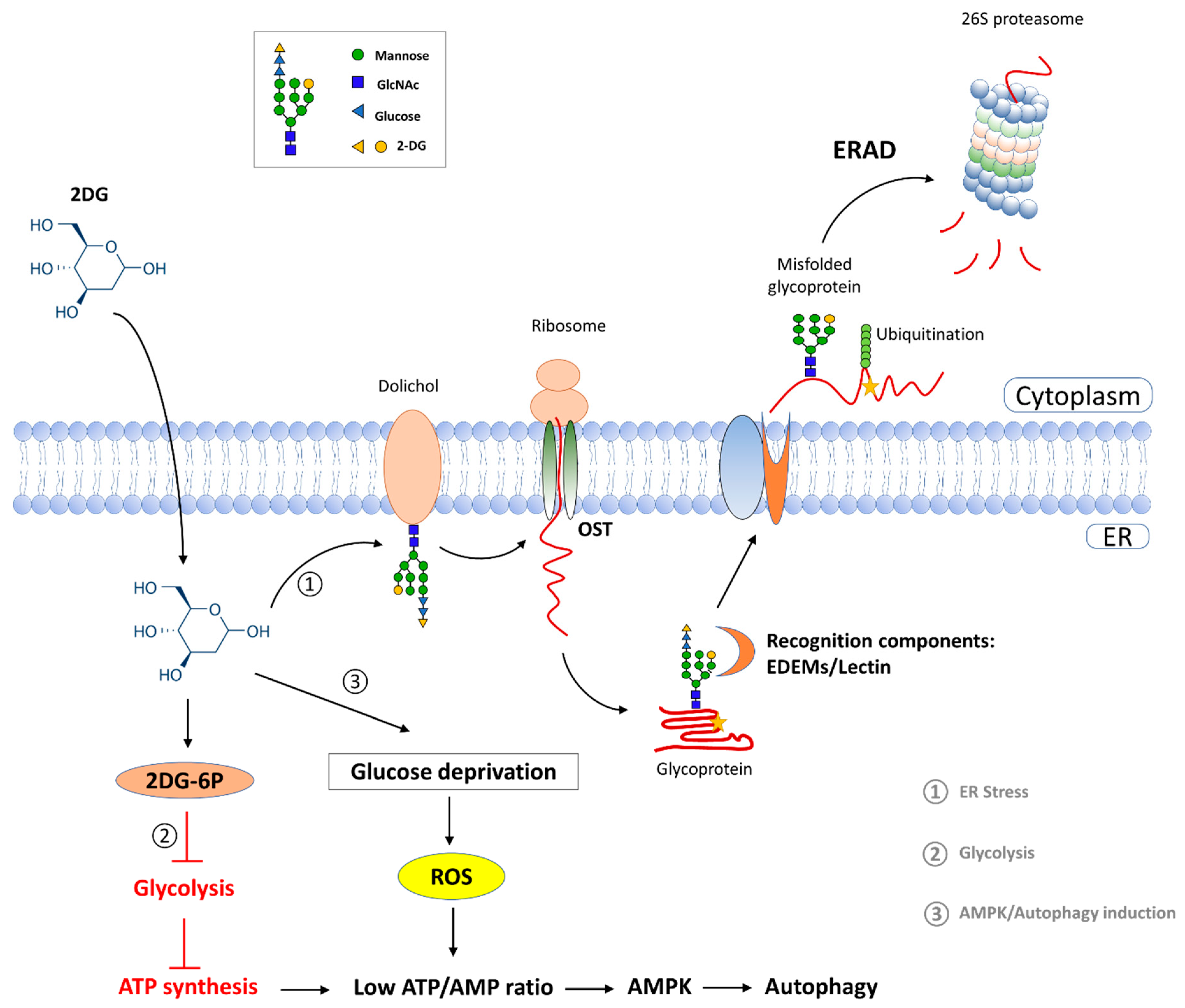
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatment
4.2. Protein Purification and Trypsin Digestion
4.3. Enrichment of Intact N-Glycopeptides
4.4. LC–MS/MS Analysis
4.5. Database Searching and Data Analysis
4.6. Gene Ontology Enrichment Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pelicano, H.; Martin, D.S.; Xu, R.-H.; Huang, P. Glycolysis inhibition for anticancer treatment. Oncogene 2006, 25, 4633–4646. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.-H.; Pelicano, H.; Zhou, Y.; Carew, J.S.; Feng, L.; Bhalla, K.N.; Keating, M.J.; Huang, P. Inhibition of Glycolysis in Cancer Cells: A Novel Strategy to Overcome Drug Resistance Associated with Mitochondrial Respiratory Defect and Hypoxia. Cancer Res. 2005, 65, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.L.; Dinsdale, D.; MacFarlane, M.; Cain, K. Switching from aerobic glycolysis to oxidative phosphorylation modulates the sensitivity of mantle cell lymphoma cells to TRAIL. Oncogene 2012, 31, 4996–5006. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, J.; Wang, F.; Hu, J.; Wang, S.; Sun, Y. 2-Deoxy-D-glucose targeting of glucose metabolism in cancer cells as a potential therapy. Cancer Lett. 2014, 355, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G.; Lin, S.-C. AMP-activated protein kinase—Not just an energy sensor. F1000Research 2017, 6, 1724. [Google Scholar] [CrossRef]
- Kurtoglu, M.; Gao, N.; Shang, J.; Maher, J.C.; Lehrman, M.A.; Wangpaichitr, M.; Savaraj, N.; Lane, A.N.; Lampidis, T.J. Under normoxia, 2-deoxy-d-glucose elicits cell death in select tumor types not by inhibition of glycolysis but by interfering with N-linked glycosylation. Mol. Cancer Ther. 2007, 6, 3049–3058. [Google Scholar] [CrossRef]
- Berthe, A.; Zaffino, M.; Muller, C.; Foulquier, F.; Houdou, M.; Schulz, C.; Bost, F.; De Fay, E.; Mazerbourg, S.; Flament, S. Protein N-glycosylation alteration and glycolysis inhibition both contribute to the antiproliferative action of 2-deoxyglucose in breast cancer cells. Breast Cancer Res. Treat. 2018, 171, 581–591. [Google Scholar] [CrossRef]
- Shah, S.S.; Rodriguez, G.A.; Musick, A.; Walters, W.M.; de Cordoba, N.; Barbarite, E.; Marlow, M.M.; Marples, B.; Prince, J.S.; Komotar, R.J.; et al. Targeting Glioblastoma Stem Cells with 2-Deoxy-D-Glucose (2-DG) Potentiates Radiation-Induced Unfolded Protein Response (UPR). Cancers 2019, 11, 159. [Google Scholar] [CrossRef]
- Bieberich, E. Synthesis, Processing, and Function of N-glycans in N-glycoproteins. Adv. Neurobiol. 2014, 9, 47–70. [Google Scholar] [CrossRef]
- Dubé, S.; Fisher, J.W.; Powell, J.S. Glycosylation at specific sites of erythropoietin is essential for biosynthesis, secretion, and biological function. J. Biol. Chem. 1988, 263, 17516–17521. [Google Scholar] [CrossRef]
- Wright, A.; Morrison, S.L. Effect of glycosylation on antibody function: Implications for genetic engineering. Trends Biotechnol. 1997, 15, 26–32. [Google Scholar] [CrossRef]
- Jennewein, M.F.; Alter, G. The Immunoregulatory Roles of Antibody Glycosylation. Trends Immunol. 2017, 38, 358–372. [Google Scholar] [CrossRef] [PubMed]
- Kurtoglu, M.; Maher, J.C.; Lampidis, T.J. Differential toxic mechanisms of 2-deoxy-D-glucose versus 2-fluorodeoxy-D-glucose in hypoxic and normoxic tumor cells. Antioxid. Redox Signal. 2007, 9, 1383–1390. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, R.T.; Schmidt, M.F.G.; Datema, R. Inhibition of Glycosylation of Viral Glycoproteins. Biochem. Soc. Trans. 1979, 7, 322–326. [Google Scholar] [CrossRef]
- Maschek, G.; Savaraj, N.; Priebe, W.; Braunschweiger, P.; Hamilton, K.; Tidmarsh, G.F.; De Young, L.R.; Lampidis, T.J. 2-Deoxy-d-glucose Increases the Efficacy of Adriamycin and Paclitaxel in Human Osteosarcoma and Non-Small Cell Lung Cancers in Vivo. Cancer Res. 2004, 64, 31–34. [Google Scholar] [CrossRef]
- Xi, H.; Kurtoglu, M.; Liu, H.; Wangpaichitr, M.; You, M.; Liu, X.; Savaraj, N.; Lampidis, T.J. 2-Deoxy-d-glucose activates autophagy via endoplasmic reticulum stress rather than ATP depletion. Cancer Chemother. Pharmacol. 2010, 67, 899–910. [Google Scholar] [CrossRef]
- Andresen, L.; Skovbakke, S.L.; Persson, G.; Hagemann-Jensen, M.; Hansen, K.A.; Jensen, H.; Skov, S. 2-Deoxy d-Glucose Prevents Cell Surface Expression of NKG2D Ligands through Inhibition of N-Linked Glycosylation. J. Immunol. 2012, 188, 1847–1855. [Google Scholar] [CrossRef] [PubMed]
- Harada, Y.; Ohkawa, Y.; Maeda, K.; Taniguchi, N. Glycan quality control in and out of the endoplasmic reticulum of mammalian cells. FEBS J. 2021. Epub ahead of print. [Google Scholar] [CrossRef]
- Ishino, K.; Kudo, M.; Peng, W.-X.; Kure, S.; Kawahara, K.; Teduka, K.; Kawamoto, Y.; Kitamura, T.; Fujii, T.; Yamamoto, T.; et al. 2-Deoxy- d -glucose increases GFAT1 phosphorylation resulting in endoplasmic reticulum-related apoptosis via disruption of protein N -glycosylation in pancreatic cancer cells. Biochem. Biophys. Res. Commun. 2018, 501, 668–673. [Google Scholar] [CrossRef]
- Lee, N.; Jang, W.-J.; Seo, J.H.; Lee, S.; Jeong, C.-H. 2-Deoxy-d-Glucose-Induced Metabolic Alteration in Human Oral Squamous SCC15 Cells: Involvement of N-Glycosylation of Axl and Met. Metabolites 2019, 9, 188. [Google Scholar] [CrossRef]
- Defenouillère, Q.; Verraes, A.; Laussel, C.; Friedrich, A.; Schacherer, J.; Léon, S. The induction of HAD-like phosphatases by multiple signaling pathways confers resistance to the metabolic inhibitor 2-deoxyglucose. Sci. Signal. 2019, 12, eaaw8000. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, W.; Li, Y.; Wang, W.; Wang, T.; Li, L.; Han, Z.; Wang, S.; Ma, D.; Wang, H. Proteomics Analysis of Normal and Senescent NG108-15 Cells: GRP78 Plays a Negative Role in Cisplatin-Induced Senescence in the NG108-15 Cell Line. PLoS ONE 2014, 9, e90114. [Google Scholar] [CrossRef] [PubMed]
- Urakami, K.; Zangiacomi, V.; Yamaguchi, K.; Kusuhara, M. Impact of 2-deoxy-D-glucose on the target metabolome profile of a human endometrial cancer cell line. Biomed. Res. 2013, 34, 221–229. [Google Scholar] [CrossRef]
- Bern, M.; Kil, Y.J.; Becker, C. Byonic: Advanced Peptide and Protein Identification Software. Curr. Protoc. Bioinform. 2012, 40, 13–20. [Google Scholar] [CrossRef]
- Klein, J.; Carvalho, L.; Zaia, J. Application of network smoothing to glycan LC-MS profiling. Bioinformatics 2018, 34, 3511–3518. [Google Scholar] [CrossRef]
- Polasky, D.A.; Yu, F.; Teo, G.C.; Nesvizhskii, A.I. Fast and comprehensive N- and O-glycoproteomics analysis with MSFragger-Glyco. Nat. Methods 2020, 17, 1125–1132. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Pan, S.; Chen, R.; Tamura, Y.; Crispin, D.A.; Lai, L.A.; May, D.; McIntosh, M.W.; Goodlett, D.R.; Brentnall, T.A. Quantitative Glycoproteomics Analysis Reveals Changes in N-Glycosylation Level Associated with Pancreatic Ductal Adenocarcinoma. J. Proteome Res. 2014, 13, 1293–1306. [Google Scholar] [CrossRef]
- Nigjeh, E.N.; Chen, R.; Allen-Tamura, Y.; Brand, R.E.; Brentnall, T.A.; Pan, S. Spectral library-based glycopeptide analysis-detection of circulating galectin-3 binding protein in pancreatic cancer. Proteom.–Clin. Appl. 2017, 11, 1700064. [Google Scholar] [CrossRef]
- Pan, S.; Brand, R.E.; Lai, L.A.; Dawson, D.W.; Donahue, T.R.; Kim, S.; Khalaf, N.I.; Othman, M.O.; Fisher, W.E.; Bronner, M.P.; et al. Proteome heterogeneity and malignancy detection in pancreatic cyst fluids. Clin. Transl. Med. 2021, 11, e506. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Mueckler, M.; Makepeace, C. Transmembrane Segment 6 of the Glut1 Glucose Transporter Is an Outer Helix and Contains Amino Acid Side Chains Essential for Transport Activity. J. Biol. Chem. 2008, 283, 11550–11555. [Google Scholar] [CrossRef]
- Mueckler, M.; Makepeace, C. Model of the Exofacial Substrate-Binding Site and Helical Folding of the Human Glut1 Glucose Transporter Based on Scanning Mutagenesis. Biochemistry 2009, 48, 5934–5942. [Google Scholar] [CrossRef]
- Lee, E.E.; Ma, J.; Sacharidou, A.; Mi, W.; Salato, V.K.; Nguyen, N.; Jiang, Y.; Pascual, J.M.; North, P.E.; Shaul, P.W.; et al. A Protein Kinase C Phosphorylation Motif in GLUT1 Affects Glucose Transport and is Mutated in GLUT1 Deficiency Syndrome. Mol. Cell 2015, 58, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, K.; Finer-Moore, J.S.; Pedersen, B.P.; Caboni, L.; Waight, A.; Hillig, R.C.; Bringmann, P.; Heisler, I.; Müller, T.; Siebeneicher, H.; et al. Mechanism of inhibition of human glucose transporter GLUT1 is conserved between cytochalasin B and phenylalanine amides. Proc. Natl. Acad. Sci. USA 2016, 113, 4711–4716. [Google Scholar] [CrossRef]
- Pajak, B.; Siwiak, E.; Sołtyka, M.; Priebe, A.; Zieliński, R.; Fokt, I.; Ziemniak, M.; Jaśkiewicz, A.; Borowski, R.; Domoradzki, T.; et al. 2-Deoxy-d-Glucose and Its Analogs: From Diagnostic to Therapeutic Agents. Int. J. Mol. Sci. 2019, 21, 234. [Google Scholar] [CrossRef]
- Riek, U.; Scholz, R.; Konarev, P.; Rufer, A.; Suter, M.; Nazabal, A.; Ringler, P.; Chami, M.; Müller, S.A.; Neumann, D.; et al. Structural properties of AMP-activated protein kinase: Dimerization, molecular shape, and changes upon ligand binding. J. Biol. Chem. 2008, 283, 18331–18343. [Google Scholar] [CrossRef]
- Stapleton, D.; Mitchelhill, K.I.; Gao, G.; Widmer, J.; Michell, B.J.; Teh, T.; House, C.M.; Fernandez, C.S.; Cox, T.; Witters, L.A.; et al. Mammalian AMP-activated Protein Kinase Subfamily. J. Biol. Chem. 1996, 271, 611–614. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Y. AMPK and Autophagy. Adv. Exp. Med. Biol. 2019, 1206, 85–108. [Google Scholar]
- Cassim, S.; Vučetić, M.; Ždralević, M.; Pouyssegur, J. Warburg and Beyond: The Power of Mitochondrial Metabolism to Collaborate or Replace Fermentative Glycolysis in Cancer. Cancers 2020, 12, 1119. [Google Scholar] [CrossRef]
- Chevet, E.; Cameron, P.H.; Pelletier, M.F.; Thomas, D.; Bergeron, J.J. The endoplasmic reticulum: Integration of protein folding, quality control, signaling and degradation. Curr. Opin. Struct. Biol. 2001, 11, 120–124. [Google Scholar] [CrossRef]
- Kaufman, R.J. Orchestrating the unfolded protein response in health and disease. J. Clin. Investig. 2002, 110, 1389–1398. [Google Scholar] [CrossRef]
- Słomińska-Wojewódzka, M.; Sandvig, K. The Role of Lectin-Carbohydrate Interactions in the Regulation of ER-Associated Protein Degradation. Molecules 2015, 20, 9816–9846. [Google Scholar] [CrossRef]
- Arnold, S.M.; Fessler, L.I.; Fessler, J.H.; Kaufman, R.J. Two Homologues Encoding Human UDP-Glucose:Glycoprotein Glucosyltransferase Differ in mRNA Expression and Enzymatic Activity. Biochemistry 2000, 39, 2149–2163. [Google Scholar] [CrossRef]
- Cheretis, C.; Dietrich, F.; Chatzistamou, I.; Politi, K.; Angelidou, E.; Kiaris, H.; Mkrtchian, S.; Koutselini, H. Expression of ERp29, an Endoplasmic Reticulum Secretion Factor in Basal-Cell Carcinoma. Am. J. Dermatopathol. 2006, 28, 410–412. [Google Scholar] [CrossRef]
- Ramírez, A.S.; Kowal, J.; Locher, K.P. Cryo–electron microscopy structures of human oligosaccharyltransferase complexes OST-A and OST-B. Science 2019, 366, 1372–1375. [Google Scholar] [CrossRef]
- Chen, R.; Lai, L.A.; Sullivan, Y.; Wong, M.; Wang, L.; Riddell, J.; Jung, L.; Pillarisetty, V.G.; Brentnall, T.A.; Pan, S. Disrupting glutamine metabolic pathways to sensitize gemcitabine-resistant pancreatic cancer. Sci. Rep. 2017, 7, 7950. [Google Scholar] [CrossRef]
- Senavirathna, L.; Ma, C.; Chen, R.; Pan, S. Proteomic Investigation of Glyceraldehyde-Derived Intracellular AGEs and Their Potential Influence on Pancreatic Ductal Cells. Cells 2021, 10, 1005. [Google Scholar] [CrossRef]
- Peng, H.; Pan, S.; Yan, Y.; Brand, R.E.; Petersen, G.M.; Chari, S.T.; Lai, L.A.; Eng, J.K.; Brentnall, T.A.; Chen, R. Systemic Proteome Alterations Linked to Early Stage Pancreatic Cancer in Diabetic Patients. Cancers 2020, 12, 1534. [Google Scholar] [CrossRef]
- Peng, H.; Chen, R.; Brentnall, T.A.; Eng, J.K.; Picozzi, V.J.; Pan, S. Predictive proteomic signatures for response of pancreatic cancer patients receiving chemotherapy. Clin. Proteom. 2019, 16, 31. [Google Scholar] [CrossRef]
- Orsburn, B. Proteome Discoverer—A Community Enhanced Data Processing Suite for Protein Informatics. Proteomes 2021, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Kerr, E.D.; Caboche, C.H.; Schulz, B.L. Posttranslational Modifications Drive Protein Stability to Control the Dynamic Beer Brewing Proteome. Mol. Cell. Proteom. 2019, 18, 1721–1731. [Google Scholar] [CrossRef] [PubMed]
- Pegg, C.L.; Phung, T.K.; Caboche, C.H.; Niamsuphap, S.; Bern, M.; Howell, K.; Schulz, B.L. Quantitative Data-Independent Acquisition Glycoproteomics of Sparkling Wine. Mol. Cell. Proteom. 2021, 20, 100020. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, C.; Tsai, H.-Y.; Zhang, Q.; Senavirathna, L.; Li, L.; Chin, L.-S.; Chen, R.; Pan, S. An Integrated Proteomic and Glycoproteomic Investigation Reveals Alterations in the N-Glycoproteomic Network Induced by 2-Deoxy-D-Glucose in Colorectal Cancer Cells. Int. J. Mol. Sci. 2022, 23, 8251. https://doi.org/10.3390/ijms23158251
Ma C, Tsai H-Y, Zhang Q, Senavirathna L, Li L, Chin L-S, Chen R, Pan S. An Integrated Proteomic and Glycoproteomic Investigation Reveals Alterations in the N-Glycoproteomic Network Induced by 2-Deoxy-D-Glucose in Colorectal Cancer Cells. International Journal of Molecular Sciences. 2022; 23(15):8251. https://doi.org/10.3390/ijms23158251
Chicago/Turabian StyleMa, Cheng, Hong-Yuan Tsai, Qi Zhang, Lakmini Senavirathna, Lian Li, Lih-Shen Chin, Ru Chen, and Sheng Pan. 2022. "An Integrated Proteomic and Glycoproteomic Investigation Reveals Alterations in the N-Glycoproteomic Network Induced by 2-Deoxy-D-Glucose in Colorectal Cancer Cells" International Journal of Molecular Sciences 23, no. 15: 8251. https://doi.org/10.3390/ijms23158251
APA StyleMa, C., Tsai, H.-Y., Zhang, Q., Senavirathna, L., Li, L., Chin, L.-S., Chen, R., & Pan, S. (2022). An Integrated Proteomic and Glycoproteomic Investigation Reveals Alterations in the N-Glycoproteomic Network Induced by 2-Deoxy-D-Glucose in Colorectal Cancer Cells. International Journal of Molecular Sciences, 23(15), 8251. https://doi.org/10.3390/ijms23158251