
A Novel Isaindigotone Derivative Displays Better Anti-Proliferation Activities and Induces Apoptosis in Gastric Cancer Cells

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Identification, Characterization, and Chemical Synthesis of BLG26
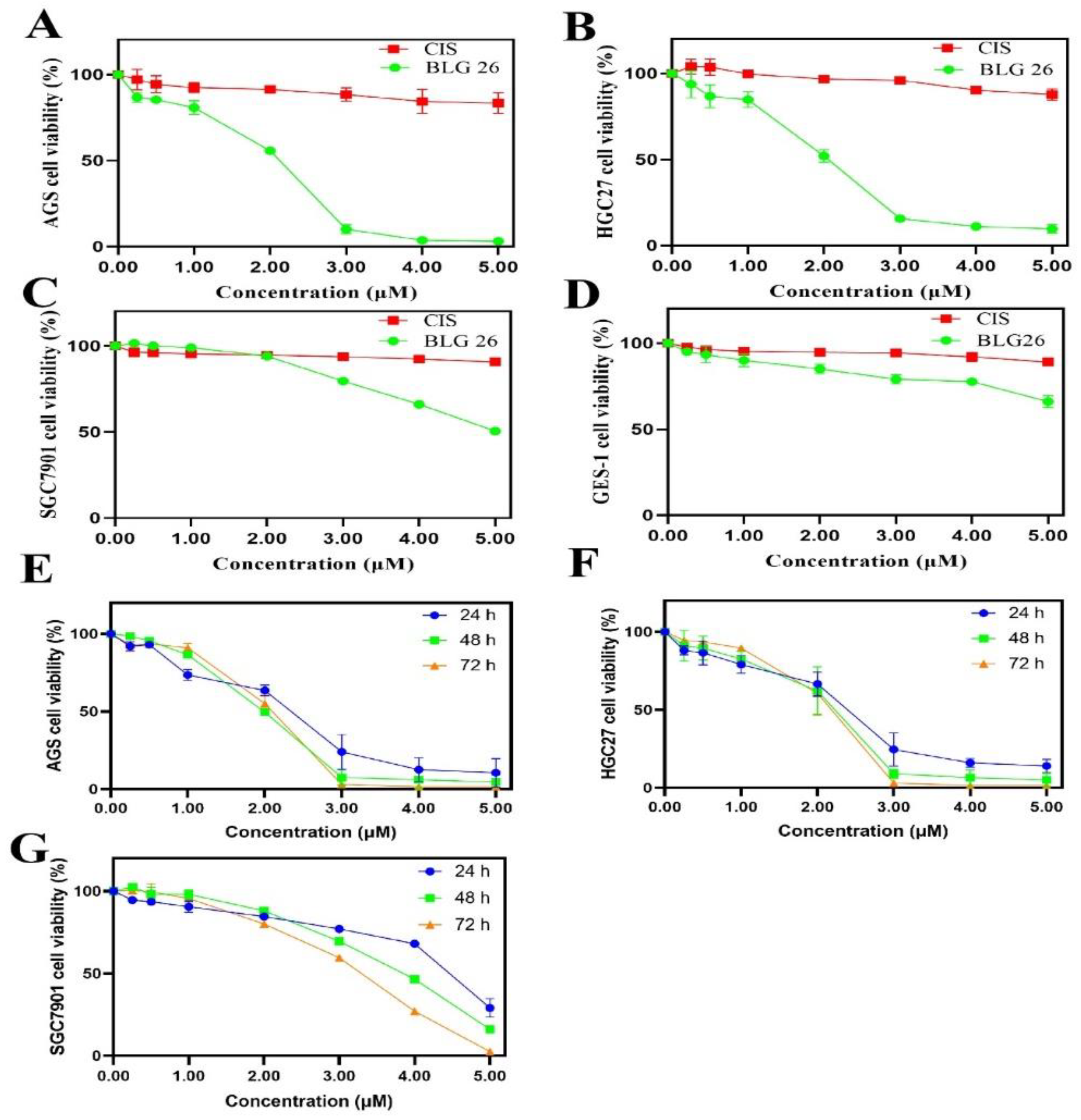
2.2. Anti-Proliferation Activity of BLG26
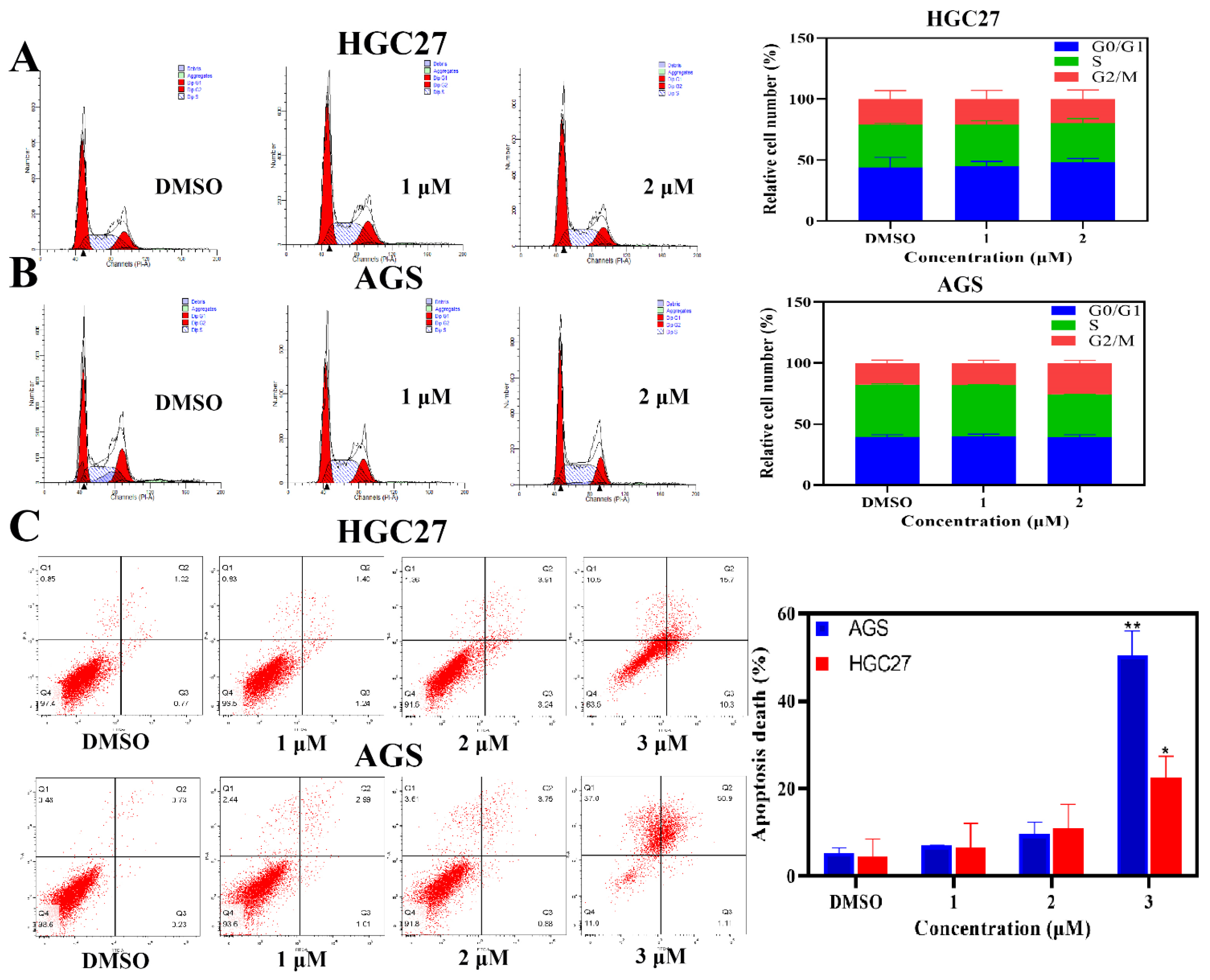
2.3. BLG26-Induced Slight Cell Cycle Arrest
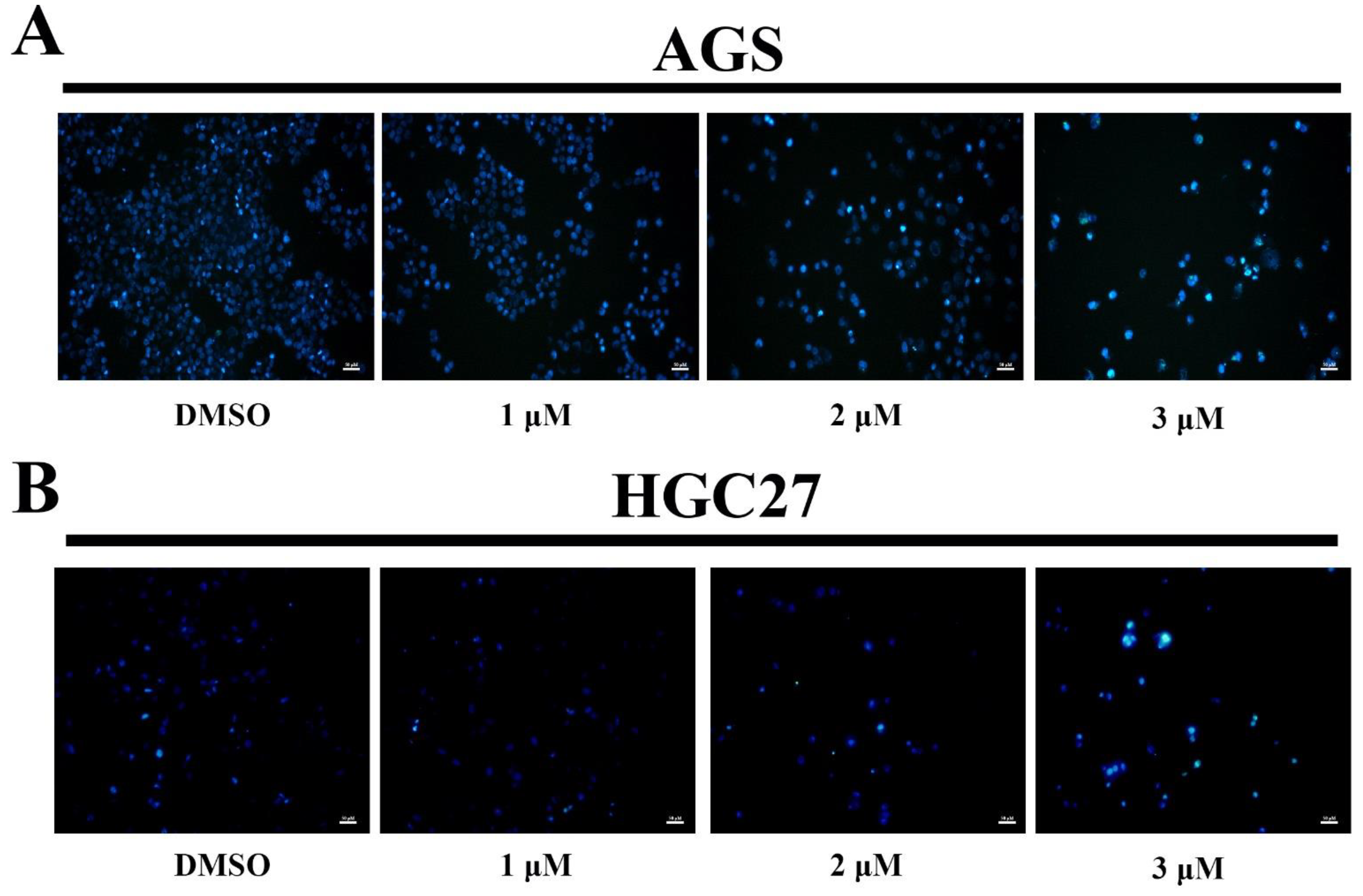
2.4. Morphological Analysis by Hoechst 33258 Staining
2.5. BLG26-Induced Apoptosis in AGS and HGC27 Cells
2.6. BLG26-Triggered Mitochondrial Membrane Potential (MMP) Loss in AGS Cells
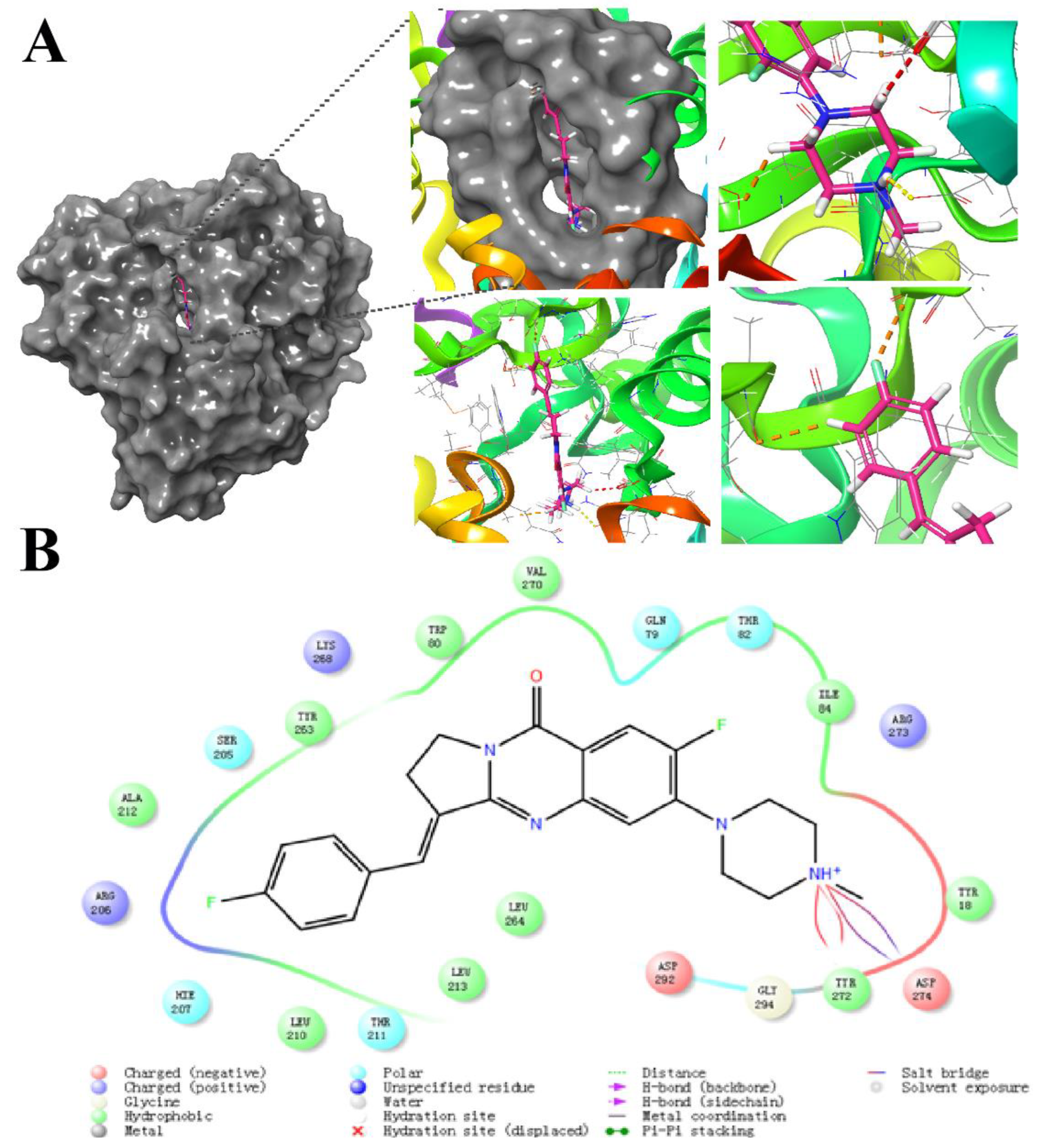
2.7. Molecular Docking Study
2.8. Apoptosis-Related Proteins Regulated by BLG26
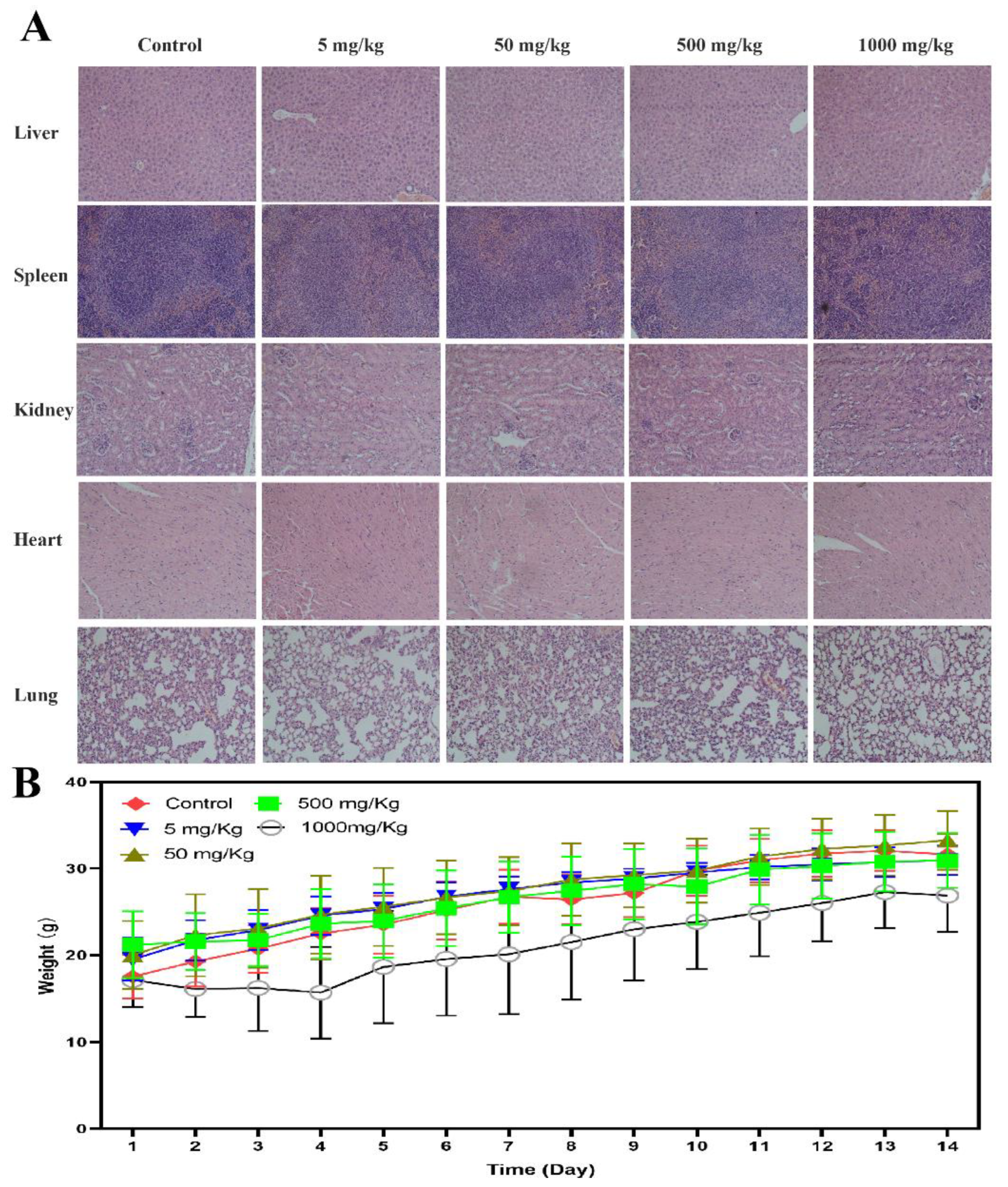
2.9. The Preliminary Acute Toxicity
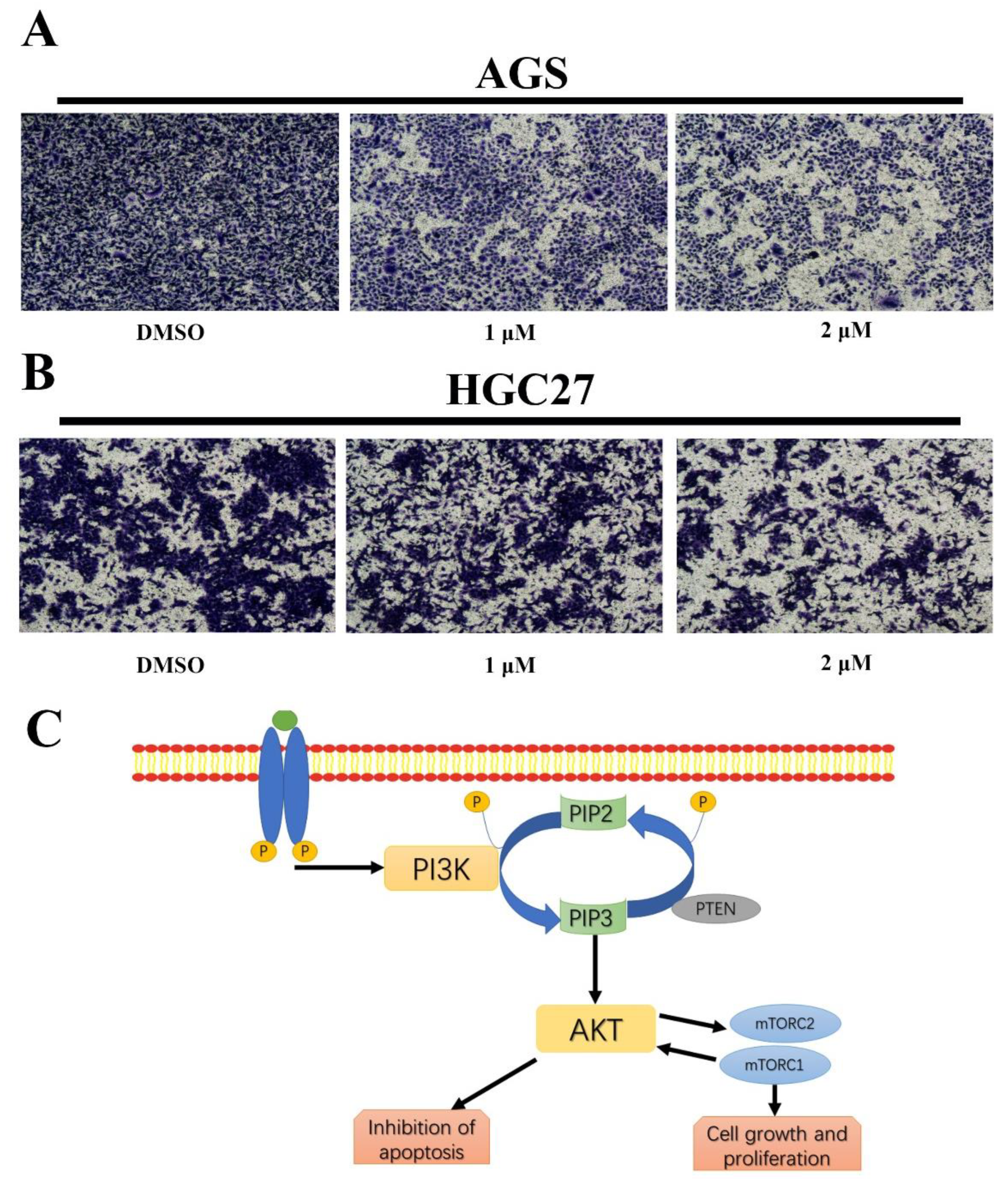
2.10. BLG26 Inhibited Cell Migration
3. Discussion
4. Materials and Methods
4.1. Chemistry
4.1.1. Synthetic Procedure for Target Compound 2
4.1.2. Synthetic Procedure for Target Compound 3
4.1.3. Synthetic Procedure for Target Compound BLG26
4.2. Cell Lines and Cell Culture
4.3. In Vitro Cytotoxicity Evaluation
4.4. Cell Cycle Analysis
4.5. Hoechst 33258 Staining
4.6. Apoptosis Analysis by Flow Cytometry
4.7. Measurement of the Mitochondrial Membrane Potential (MMP) with JC-1
4.8. Docking Studies
4.9. Western Blot Analysis
4.10. The Preliminary Acute Toxicity Study of BLG26 Using Kunming Mice
4.11. Transwell Cell Migration Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation
| GC | Gastric |
| MMP | Mitochondrial membrane potential |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| DMSO | Dimethyl sulfoxide |
| PI | Propidium iodide |
| FITC | FluoresceinIsothiocyanate |
| CCCP | Carbonyl cyanide-m-chlorophenylhydrazone |
| WB | Western Blot |
| Thr | Threonine |
| Lys | Lysine |
| Asp | Asparticacid |
| Asn | Asparagine |
| Glu | Glutamicacid |
| TFA | Trifluoroacetic acid |
| DMF | N,N-dimethyllformamide |
| FBS | Fetal bovine serum |
| PBS | Phosphate belanced solution |
| PVDF | polyvinylidene fluoride |
| SDS-PAGE | dodecyl sulfate, sodium salt-Polyacrylamide gel electrophoresis |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Xu, Z.Y.; Ruan, S.M.; Mo, S.; Qin, J.J.; Cheng, X.D. Long non-coding RNAs towards precision medicine in gastric cancer: Early diagnosis, treatment, and drug resistance. Mol. Cancer 2020, 19, 96. [Google Scholar] [CrossRef]
- Kang, M.; Wu, H.; Yang, Q.; Huang, L.; Hu, Q.; Ma, T.; Li, Z.; Liu, J. A chromosome-scale genome assembly of Isatis indigotica, an important medicinal plant used in traditional Chinese medicine: An Isatis genome. Hortic. Res. 2020, 7, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, E.K.; Kim, D.H.; Lim, H.; Shin, H.K.; Kim, J.K. The anti-inflammatory effects of a methanolic extract from Radix Isatidis in murine macrophages and mice. Inflammation 2010, 33, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Ruan, D.; Liu, W.; Shi, Y.; Tan, M.; Yang, L.; Wang, Z.; Zhou, Y.; Wang, R. Protective Effects of Aqueous Extract of Radix Isatidis on Lipopolysaccharide-Induced Sepsis in C57BL/6J Mice. J. Med. Food 2020, 23, 79–89. [Google Scholar] [CrossRef]
- Jiang, L.; Lu, Y.; Jin, J.; Dong, L.; Xu, F.; Chen, S.; Wang, Z.; Liang, G.; Shan, X. n-Butanol extract from Folium isatidis inhibits lipopolysaccharide-induced inflammatory cytokine production in macrophages and protects mice against lipopolysaccharide-induced endotoxic shock. Drug Des. Dev. Ther. 2015, 9, 5601–5609. [Google Scholar] [CrossRef] [Green Version]
- Kido, S.; Kitadai, Y.; Hattori, N.; Haruma, K.; Kido, T.; Ohta, M.; Tanaka, S.; Yoshihara, M.; Sumii, K.; Ohmoto, Y.; et al. Interleukin 8 and vascular endothelial growth factor--prognostic factors in human gastric carcinomas? Eur. J. Cancer 2001, 37, 1482–1487. [Google Scholar] [CrossRef]
- Echizen, K.; Hirose, O.; Maeda, Y.; Oshima, M. Inflammation in gastric cancer: Interplay of the COX-2/prostaglandin E2 and Toll-like receptor/MyD88 pathways. Cancer Sci. 2016, 107, 391–397. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.T.; Son, D.J.; Lee, C.K.; Yoon, D.Y.; Lee, D.H.; Park, M.H. Interleukin 32, inflammation and cancer. Pharmacol. Ther. 2017, 174, 127–137. [Google Scholar] [CrossRef]
- Murata, M. Inflammation and cancer. Environ. Health Prev. Med. 2018, 23, 860–867. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.W.; Li, Y.P.; Ye, W.J.; Chen, S.B.; Hou, J.Q.; Tan, J.H.; Ou, T.M.; Li, D.; Gu, L.Q.; Huang, Z.S. Design, synthesis and evaluation of isaindigotone derivatives as dual inhibitors for acetylcholinesterase and amyloid beta aggregation. Bioorg. Med. Chem. 2012, 20, 2527–2534. [Google Scholar] [CrossRef] [PubMed]
- Shakhidoyatov, K.M.; Elmuradov, B.Z. Tricyclic Quinazoline Alkaloids: Isolation, Synthesis, Chemical Modification, and Biological Activity. Chem. Nat. Compd. 2014, 50, 781–800. [Google Scholar] [CrossRef]
- Xiao, H.; Xu, J. Isaindigotone as an inhibitor of the lipopolysaccharide-induced inflammatory reaction of BV-2 cells and corresponding mechanisms. Mol. Med. Rep. 2019, 19, 2890–2896. [Google Scholar] [CrossRef]
- Shan, C.; Yan, J.W.; Wang, Y.Q.; Che, T.; Huang, Z.L.; Chen, A.C.; Yao, P.F.; Tan, J.H.; Li, D.; Ou, T.M.; et al. Design, Synthesis, and Evaluation of Isaindigotone Derivatives To Downregulate c-myc Transcription via Disrupting the Interaction of NM23-H2 with G-Quadruplex. J. Med. Chem. 2017, 60, 1292–1308. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Q.; Huang, Z.L.; Chen, S.B.; Wang, C.X.; Shan, C.; Yin, Q.K.; Ou, T.M.; Li, D.; Gu, L.Q.; Tan, J.H.; et al. Design, Synthesis, and Evaluation of New Selective NM23-H2 Binders as c-MYC Transcription Inhibitors via Disruption of the NM23-H2/G-Quadruplex Interaction. J. Med. Chem. 2017, 60, 6924–6941. [Google Scholar] [CrossRef]
- Barnes, P.J. Mechanisms of development of multimorbidity in the elderly. Eur. Respir. J. 2015, 45, 790–806. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wei, W. Anti-inflammatory and immunoregulatory effects of paeoniflorin and total glucosides of paeony. Pharmacol. Ther. 2020, 207, 107452. [Google Scholar] [CrossRef]
- Ala, M.; Ala, M. Metformin for Cardiovascular Protection, Inflammatory Bowel Disease, Osteoporosis, Periodontitis, Polycystic Ovarian Syndrome, Neurodegeneration, Cancer, Inflammation and Senescence: What Is Next? ACS Pharmacol. Transl. Sci. 2021, 4, 1747–1770. [Google Scholar] [CrossRef]
- Deng, J.; Ma, Y.; He, Y.; Yang, H.; Chen, Y.; Wang, L.; Huang, D.; Qiu, S.; Tao, X.; Chen, W. A Network Pharmacology-Based Investigation to the Pharmacodynamic Material Basis and Mechanisms of the Anti-Inflammatory and Anti-Viral Effect of Isatis indigotica. Drug Des. Dev. Ther. 2021, 15, 3193–3206. [Google Scholar] [CrossRef]
- Chen, W.; Zheng, R.; Baade, P.D.; Zhang, S.; Zeng, H.; Bray, F.; Jemal, A.; Yu, X.Q.; He, J. Cancer statistics in China, 2015. CA Cancer J. Clin. 2016, 66, 115–132. [Google Scholar] [CrossRef] [Green Version]
- Cragg, G.M.; Pezzuto, J.M. Natural Products as a Vital Source for the Discovery of Cancer Chemotherapeutic and Chemopreventive Agents. Med. Princ. Pract. 2016, 25 (Suppl. 2), 41–59. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Zhao, X.; Yang, D.; Cao, X.; Yu, Y.; Jiang, H.; Zhou, X.; Li, C.; Guo, Q. LFG-500, a newly synthesized flavonoid, induces apoptosis in human ovarian carcinoma SKOV3 cells with involvement of the reactive oxygen species-mitochondria pathway. Exp. Ther. Med. 2017, 13, 2819–2827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Bai, D.; Du, J.; Ren, H.; Zhang, Z.; Jiang, C.; Lu, N. Induction of cell cycle arrest and apoptosis by CPUC002 through stabilization of p53 and suppression of STAT3 signaling pathway in multiple myeloma. Cell Biol. Toxicol. 2021, 37, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Bock, F.J.; Tait, S.W.G. Mitochondria as multifaceted regulators of cell death. Nat. Rev. Mol. Cell Biol. 2020, 21, 85–100. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, X. Cytochrome c promotes caspase-9 activation by inducing nucleotide binding to Apaf-1. J. Biol. Chem. 2000, 275, 31199–31203. [Google Scholar] [CrossRef] [Green Version]
- Antonsson, B.; Montessuit, S.; Lauper, S.; Eskes, R.; Martinou, J.C. Bax oligomerization is required for channel-forming activity in liposomes and to trigger cytochrome c release from mitochondria. Biochem. J. 2000, 345 Pt 2, 271–278. [Google Scholar] [CrossRef]
- Gaumer, S.; Guénal, I.; Brun, S.; Théodore, L.; Mignotte, B. Bcl-2 and Bax mammalian regulators of apoptosis are functional in Drosophila. Cell Death Differ. 2000, 7, 804–814. [Google Scholar] [CrossRef] [Green Version]
- Ediriweera, M.K.; Tennekoon, K.H.; Samarakoon, S.R. Role of the PI3K/AKT/mTOR signaling pathway in ovarian cancer: Biological and therapeutic significance. Semin. Cancer Biol. 2019, 59, 147–160. [Google Scholar] [CrossRef]
- Uko, N.E.; Güner, O.F.; Matesic, D.F.; Bowen, J.P. Akt Pathway Inhibitors. Curr. Top. Med. Chem. 2020, 20, 883–900. [Google Scholar] [CrossRef]
- Nitulescu, G.M.; Margina, D.; Juzenas, P.; Peng, Q.; Olaru, O.T.; Saloustros, E.; Fenga, C.; Spandidos, D.; Libra, M.; Tsatsakis, A.M. Akt inhibitors in cancer treatment: The long journey from drug discovery to clinical use (Review). Int. J. Oncol. 2016, 48, 869–885. [Google Scholar] [CrossRef] [Green Version]
- Staal, S.P. Molecular cloning of the akt oncogene and its human homologues AKT1 and AKT2: Amplification of AKT1 in a primary human gastric adenocarcinoma. Proc. Natl. Acad. Sci. USA 1987, 84, 5034–5037. [Google Scholar] [CrossRef] [Green Version]
- Smith, U.; Carvalho, E.; Mosialou, E.; Beguinot, F.; Formisano, P.; Rondinone, C. PKB inhibition prevents the stimulatory effect of insulin on glucose transport and protein translocation but not the antilipolytic effect in rat adipocytes. Biochem. Biophys. Res. Commun. 2000, 268, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Van Ummersen, L.; Binger, K.; Volkman, J.; Marnocha, R.; Tutsch, K.; Kolesar, J.; Arzoomanian, R.; Alberti, D.; Wilding, G. A phase I trial of perifosine (NSC 639966) on a loading dose/maintenance dose schedule in patients with advanced cancer. Clin. Cancer Res. 2004, 10, 7450–7456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Zhou, Q.; Qiu, Q.; Hou, L.; Wu, M.; Li, J.; Li, X.; Lu, B.; Cheng, X.; Liu, P.; et al. CircPLEKHM3 acts as a tumor suppressor through regulation of the miR-9/BRCA1/DNAJB6/KLF4/AKT1 axis in ovarian cancer. Mol. Cancer 2019, 18, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calleja, V.; Laguerre, M.; Parker, P.J.; Larijani, B. Role of a novel PH-kinase domain interface in PKB/Akt regulation: Structural mechanism for allosteric inhibition. PLoS Biol. 2009, 7, e17. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Jabeen, I. Pharmacoinformatic Approaches to Design Novel Inhibitors of Protein Kinase B Pathways in Cancer. Curr. Cancer Drug Targets 2018, 18, 830–846. [Google Scholar] [CrossRef]
- Merckaert, T.; Zwaenepoel, O.; Gevaert, K.; Gettemans, J. An AKT2-specific nanobody that targets the hydrophobic motif induces cell cycle arrest, autophagy and loss of focal adhesions in MDA-MB-231 cells. Biomed. Pharmacother. Biomed. Pharmacother. 2021, 133, 111055. [Google Scholar] [CrossRef]
- Yu, X.; Xu, J.; Xie, L.; Wang, L.; Shen, Y.; Cahuzac, K.M.; Chen, X.; Liu, J.; Parsons, R.E.; Jin, J. Design, Synthesis, and Evaluation of Potent, Selective, and Bioavailable AKT Kinase Degraders. J. Med. Chem. 2021, 64, 18054–18081. [Google Scholar] [CrossRef]
- Hua, H.; Zhang, H.; Chen, J.; Wang, J.; Liu, J.; Jiang, Y. Targeting Akt in cancer for precision therapy. J. Hematol. Oncol. 2021, 14, 128. [Google Scholar] [CrossRef]
- Fattahi, S.; Amjadi-Moheb, F.; Tabaripour, R.; Ashrafi, G.H.; Akhavan-Niaki, H. PI3K/AKT/mTOR signaling in gastric cancer: Epigenetics and beyond. Life Sci. 2020, 262, 118513. [Google Scholar] [CrossRef]
- Ahmmed, B.; Khan, M.N.; Nisar, M.A.; Kampo, S.; Zheng, Q.; Li, Y.; Yan, Q. Tunicamycin enhances the suppressive effects of cisplatin on lung cancer growth through PTX3 glycosylation via AKT/NF-κB signaling pathway. Int. J. Oncol. 2019, 54, 431–442. [Google Scholar] [CrossRef] [Green Version]
- Klein, G.; Vellenga, E.; Fraaije, M.W.; Kamps, W.A.; de Bont, E.S. The possible role of matrix metalloproteinase (MMP)-2 and MMP-9 in cancer, e.g. acute leukemia. Crit. Rev. Oncol./Hematol. 2004, 50, 87–100. [Google Scholar] [CrossRef]
- Tan, J.H.; Ou, T.M.; Hou, J.Q.; Lu, Y.J.; Huang, S.L.; Luo, H.B.; Wu, J.Y.; Huang, Z.S.; Wong, K.Y.; Gu, L.Q. Isaindigotone derivatives: A new class of highly selective ligands for telomeric G-quadruplex DNA. J. Med. Chem. 2009, 52, 2825–2835. [Google Scholar] [CrossRef]
- Yin, Q.K.; Wang, C.X.; Wang, Y.Q.; Guo, Q.L.; Zhang, Z.L.; Ou, T.M.; Huang, S.L.; Li, D.; Wang, H.G.; Tan, J.H.; et al. Discovery of Isaindigotone Derivatives as Novel Bloom’s Syndrome Protein (BLM) Helicase Inhibitors That Disrupt the BLM/DNA Interactions and Regulate the Homologous Recombination Repair. J. Med. Chem. 2019, 62, 3147–3162. [Google Scholar] [CrossRef]
- Pan, L.; Tan, J.H.; Hou, J.Q.; Huang, S.L.; Gu, L.Q.; Huang, Z.S. Design, synthesis and evaluation of isaindigotone derivatives as acetylcholinesterase and butyrylcholinesterase inhibitors. Bioorg. Med. Chem. Lett. 2008, 18, 3790–3793. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | PANC-1 | HCT116 | SMMC7721 | AGS | GES-1 |
|---|---|---|---|---|---|
| BLG26 | 44.61 ± 2.36 | 3.25 ± 0.05 | 20.25 ± 1.37 | 1.45 ± 0.09 | 14.06 ± 1.97 |
| CIS | >50.00 | >50.00 | >50.00 | >50.00 | >50.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, K.; Yang, C.; Zhou, Z.; Ma, Y.; Tian, Y.; Zhang, R.; Zhang, H.; Jiang, X.; Zhu, H.; Liu, H.; et al. A Novel Isaindigotone Derivative Displays Better Anti-Proliferation Activities and Induces Apoptosis in Gastric Cancer Cells. Int. J. Mol. Sci. 2022, 23, 8028. https://doi.org/10.3390/ijms23148028
Du K, Yang C, Zhou Z, Ma Y, Tian Y, Zhang R, Zhang H, Jiang X, Zhu H, Liu H, et al. A Novel Isaindigotone Derivative Displays Better Anti-Proliferation Activities and Induces Apoptosis in Gastric Cancer Cells. International Journal of Molecular Sciences. 2022; 23(14):8028. https://doi.org/10.3390/ijms23148028
Chicago/Turabian StyleDu, Kangjia, Chengjie Yang, Zhongkun Zhou, Yunhao Ma, Yanan Tian, Rentao Zhang, Hao Zhang, Xinrong Jiang, Hongmei Zhu, Huanxiang Liu, and et al. 2022. "A Novel Isaindigotone Derivative Displays Better Anti-Proliferation Activities and Induces Apoptosis in Gastric Cancer Cells" International Journal of Molecular Sciences 23, no. 14: 8028. https://doi.org/10.3390/ijms23148028
APA StyleDu, K., Yang, C., Zhou, Z., Ma, Y., Tian, Y., Zhang, R., Zhang, H., Jiang, X., Zhu, H., Liu, H., Chen, P., & Liu, Y. (2022). A Novel Isaindigotone Derivative Displays Better Anti-Proliferation Activities and Induces Apoptosis in Gastric Cancer Cells. International Journal of Molecular Sciences, 23(14), 8028. https://doi.org/10.3390/ijms23148028