
Virus Elimination from Naturally Infected Field Cultivars of Potato (Solanum tuberosum) by Transgenic RNA Interference

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of Intron-Hairpin RNA Expression Cassettes
2.2. Virus Detection by ELISA
2.3. Potato Transformation
2.4. Molecular Analysis of Putative Transgenic Plants
2.5. RNA Extraction and Blot Hybridization Analysis of Transgenic Plants
2.6. Illumina Sequencing Analysis of Transgenic, Viral, and Plant sRNAs
3. Results and Discussion
3.1. Virus Elimination from Field Potato Cultivars Using Transgenic RNAi
3.2. Field Trials of Transgenic Plants for Virus Resistance
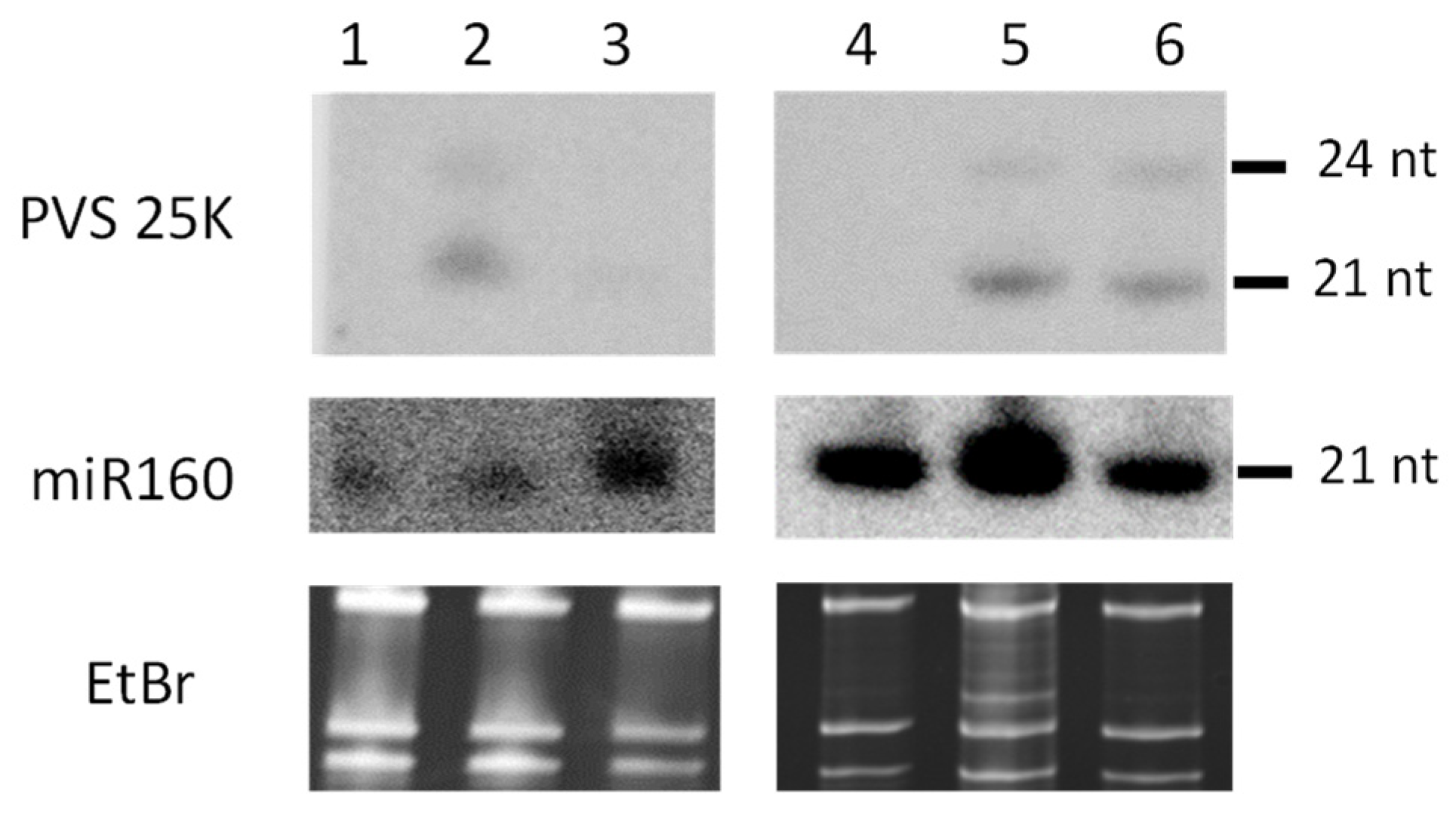
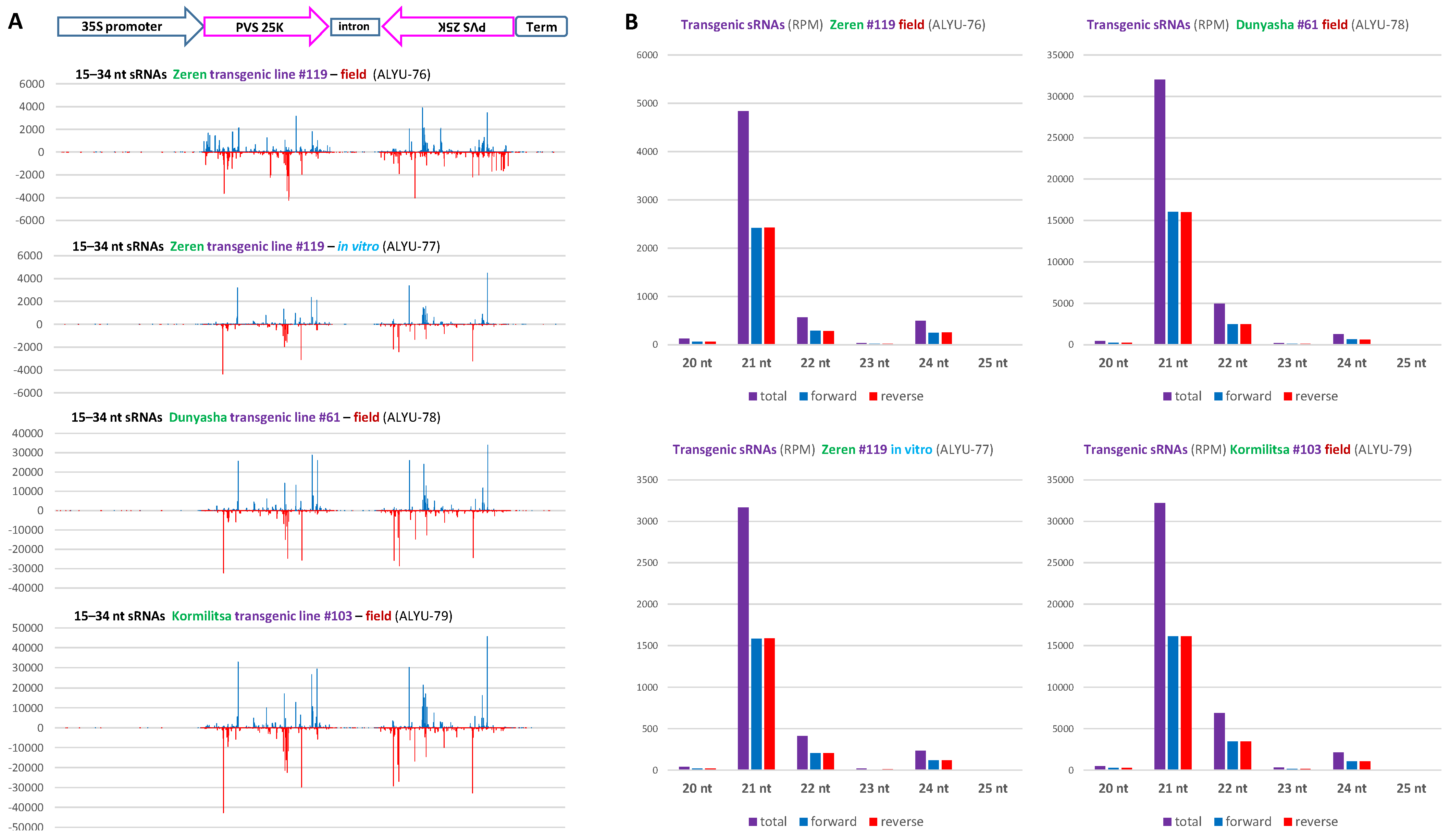
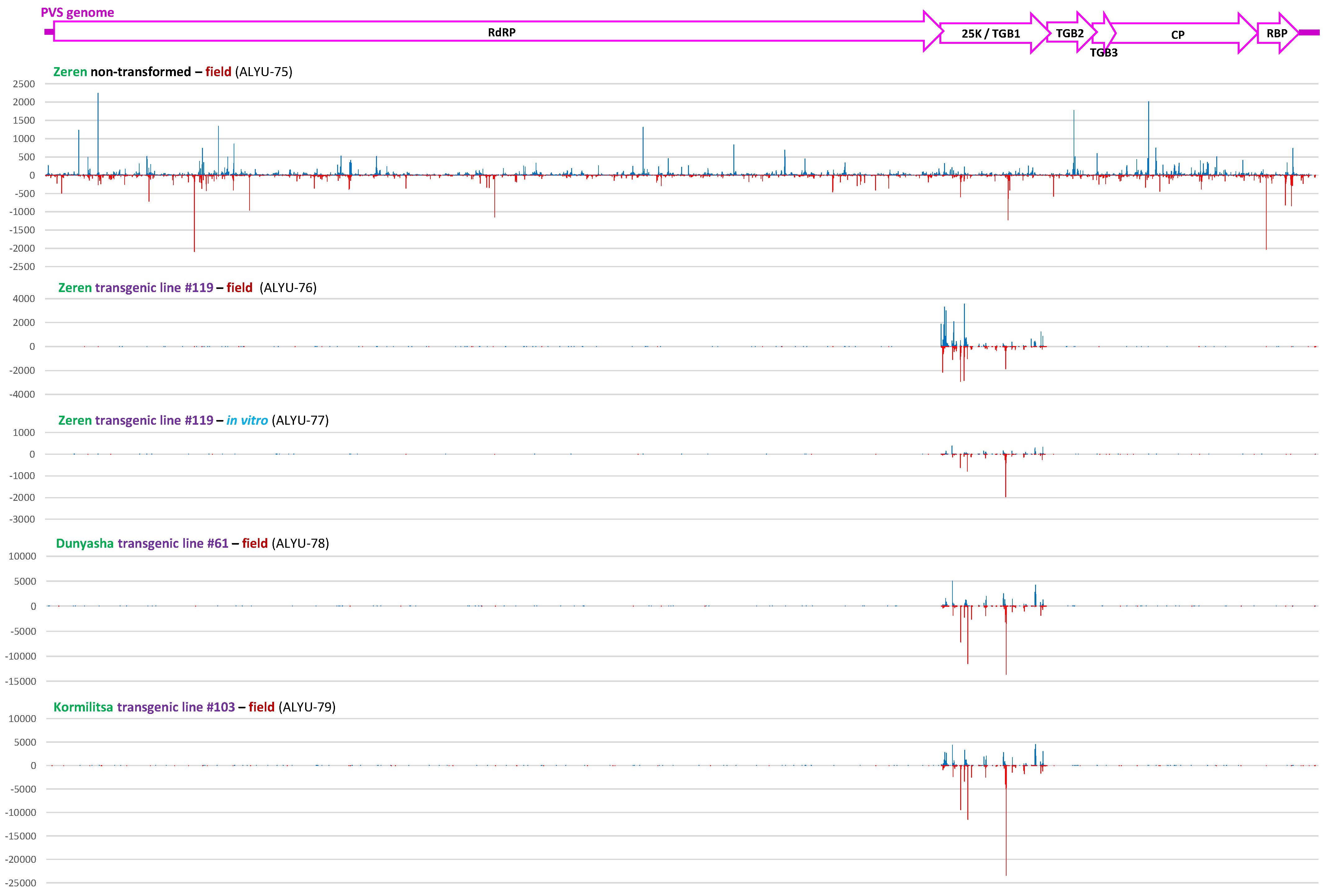
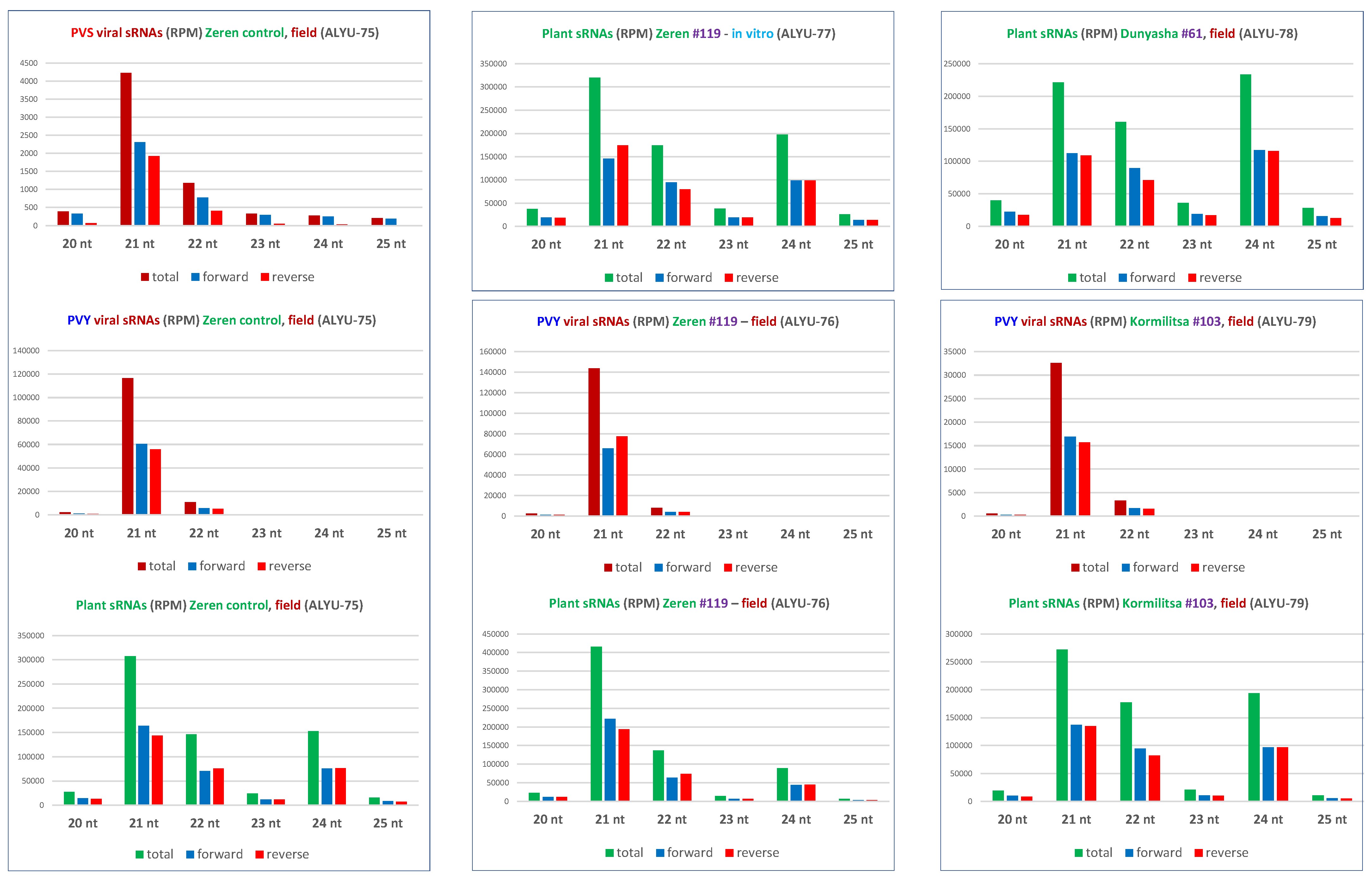
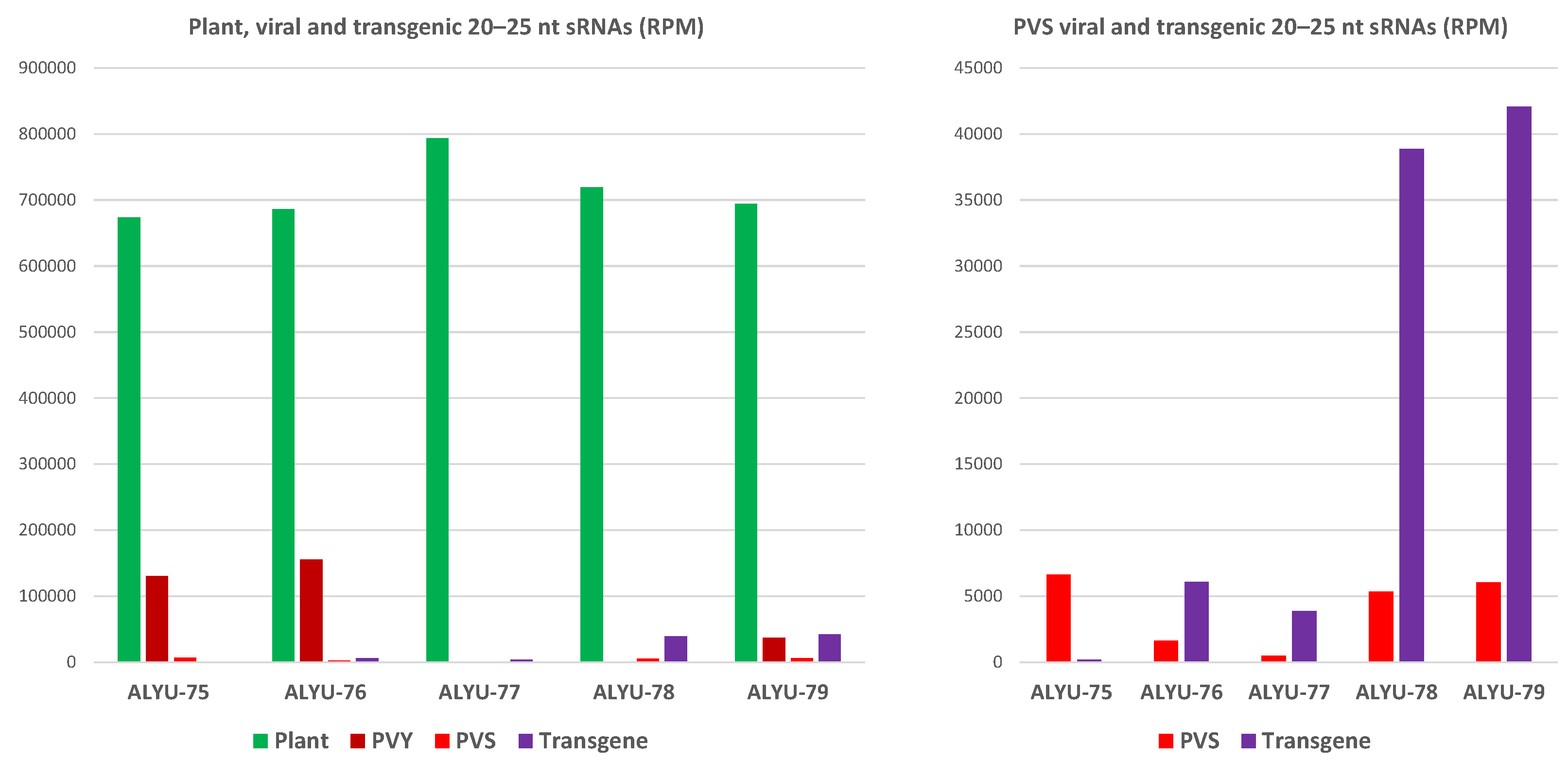
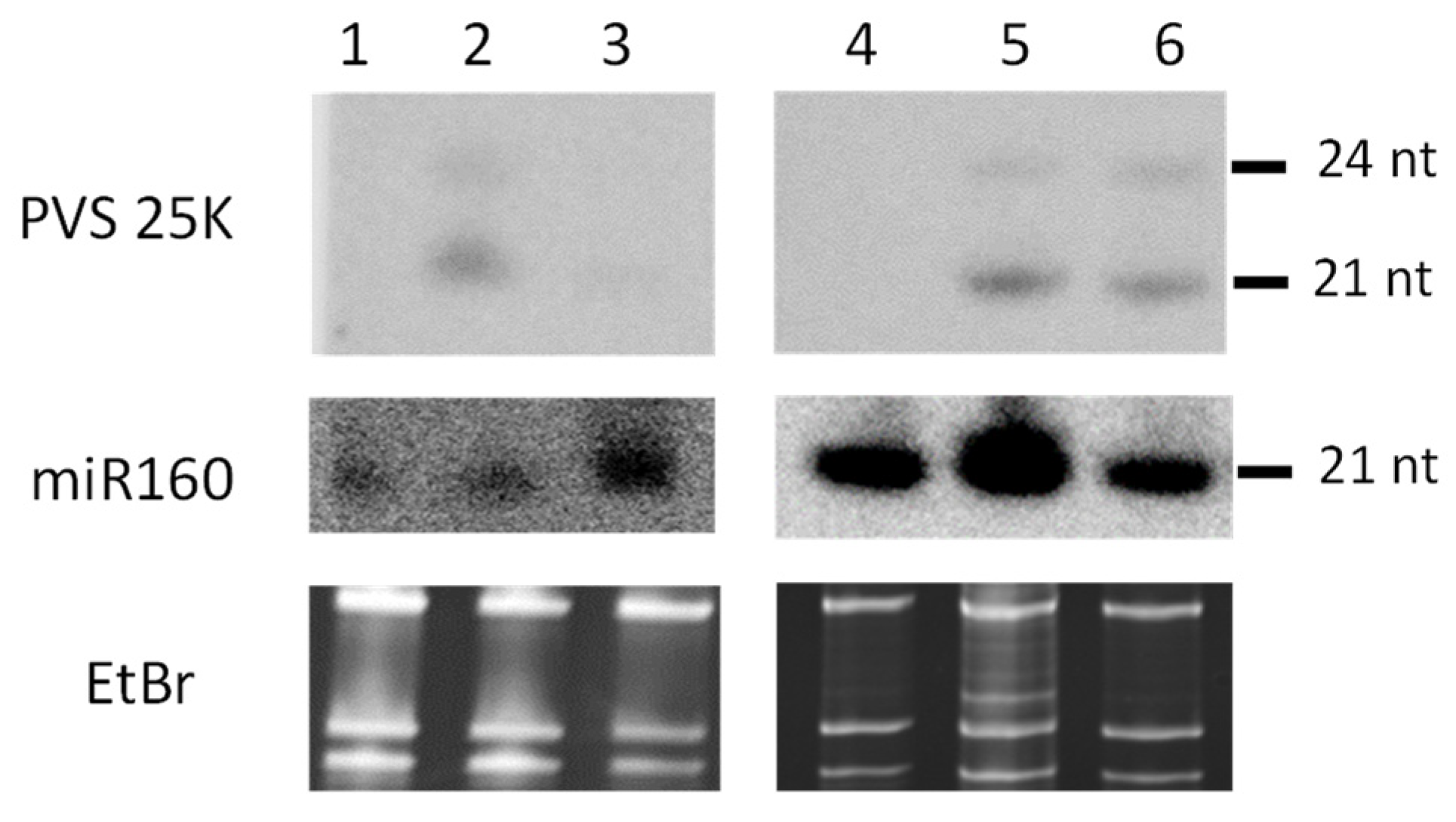
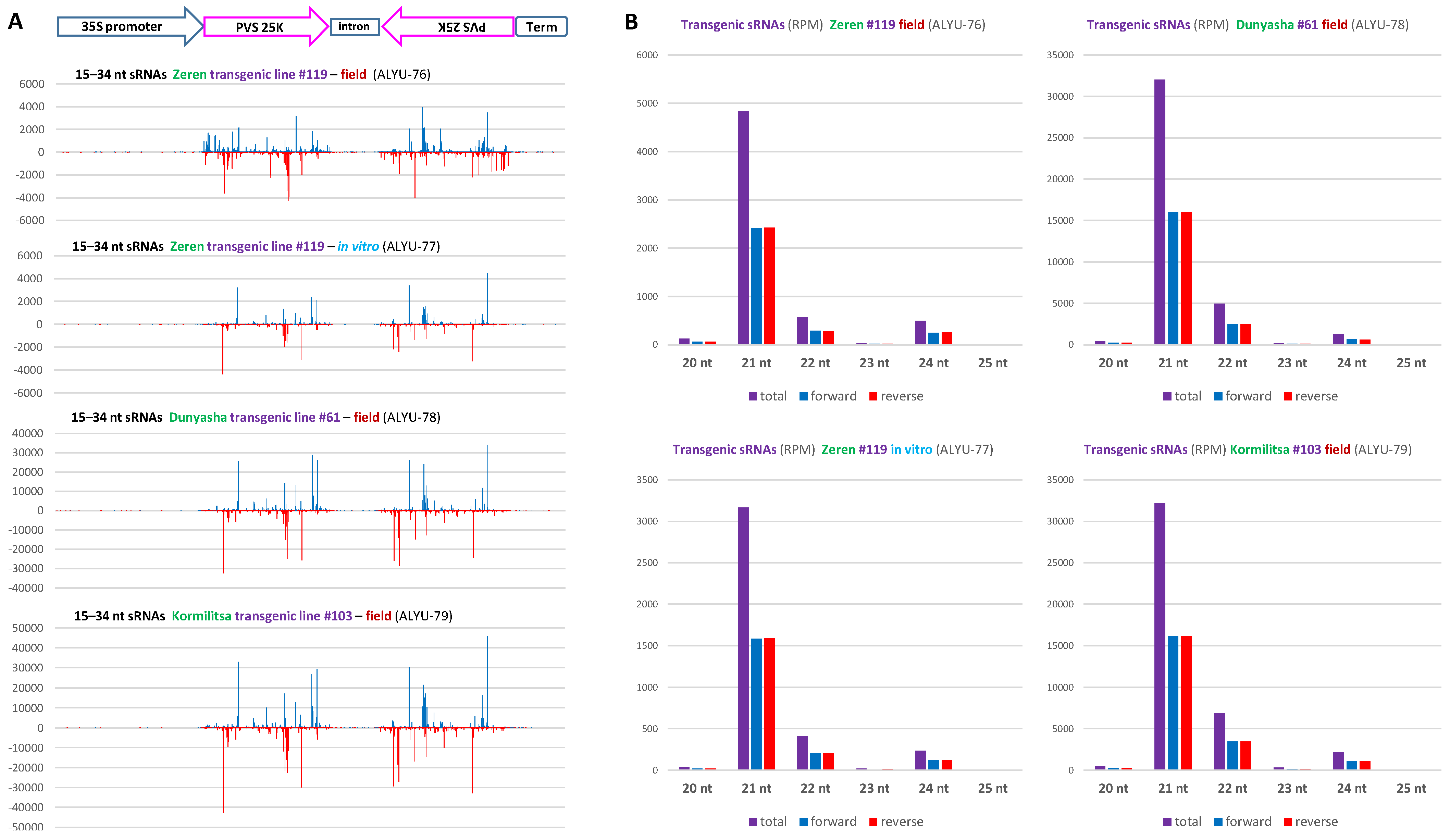
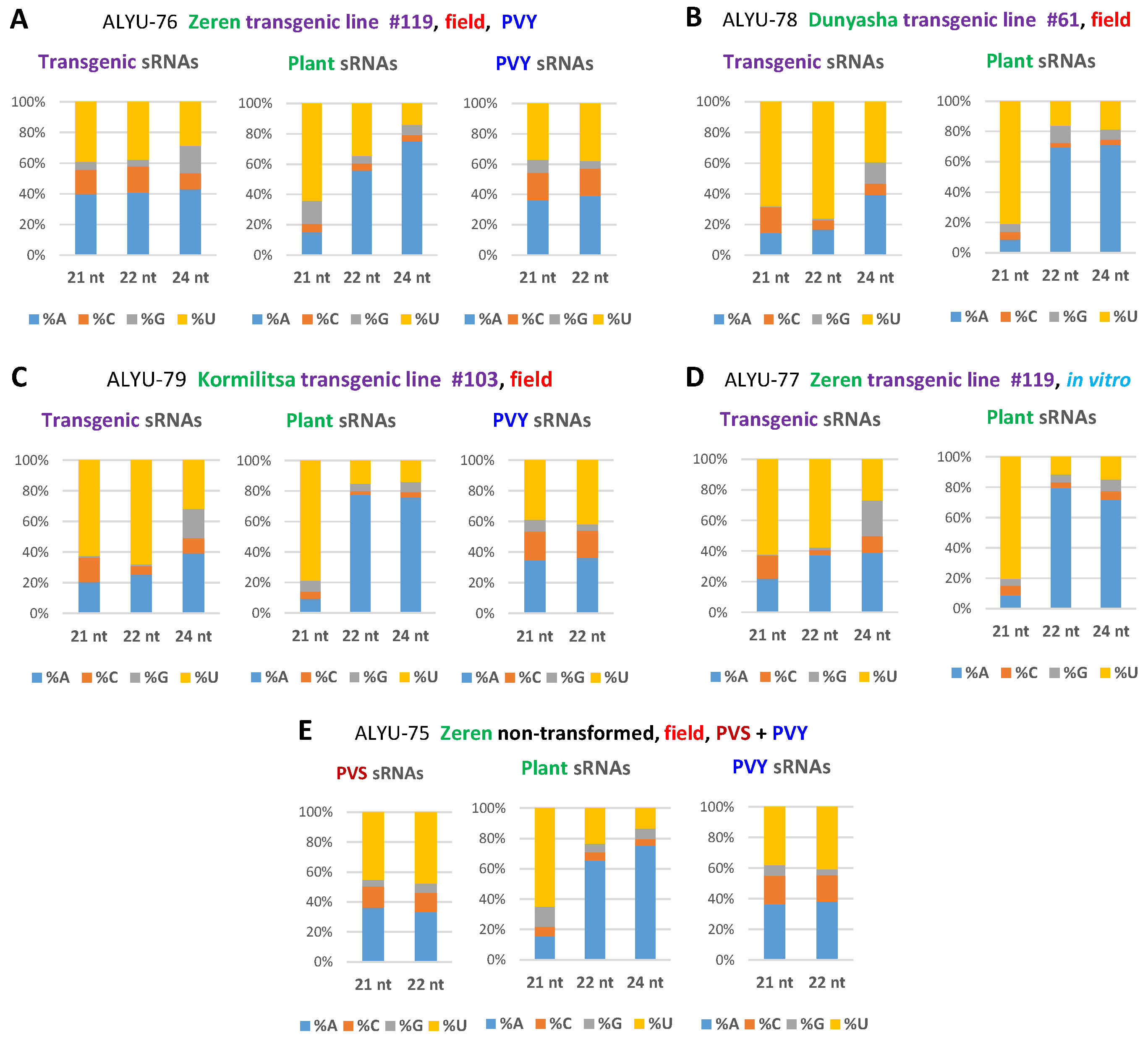
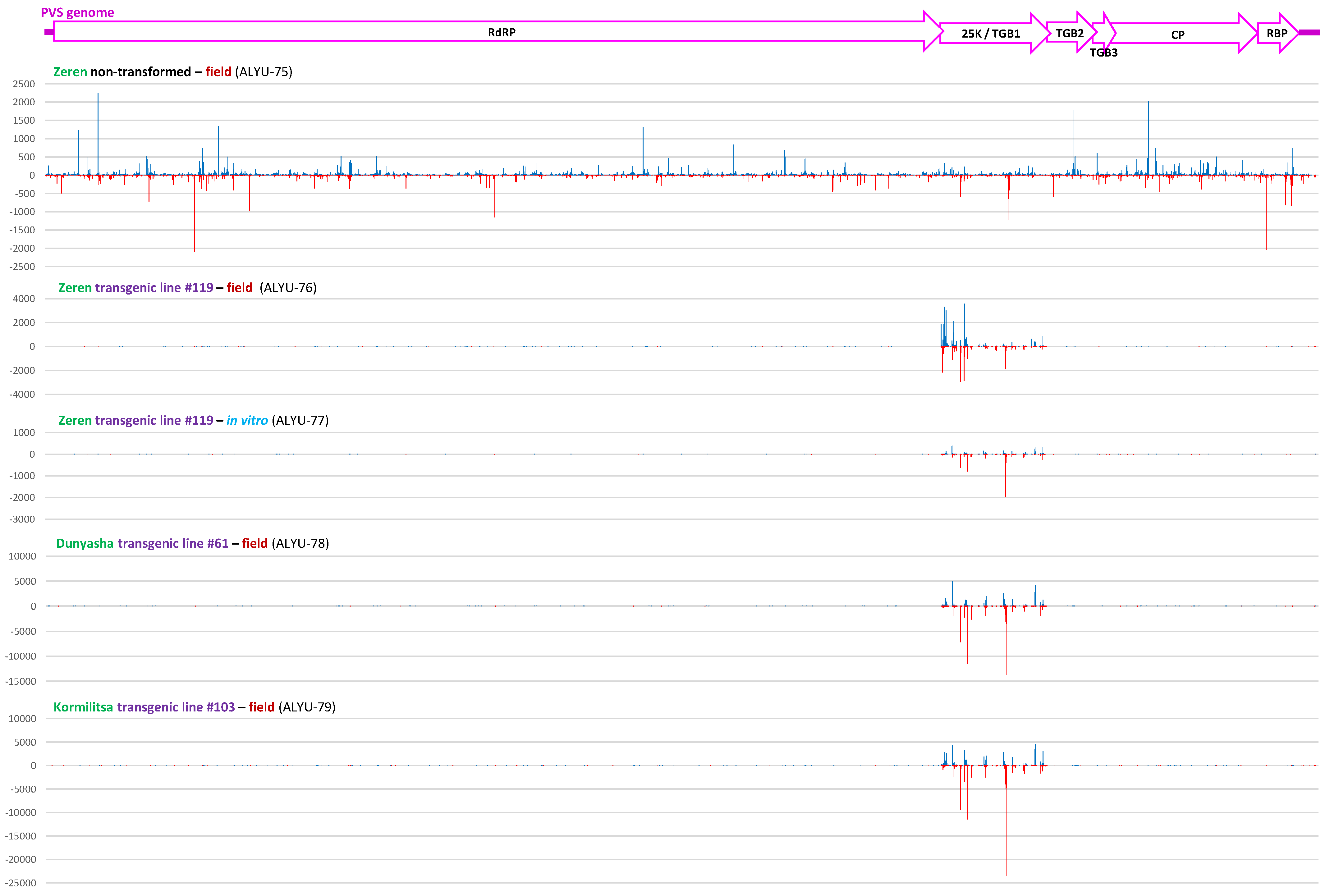
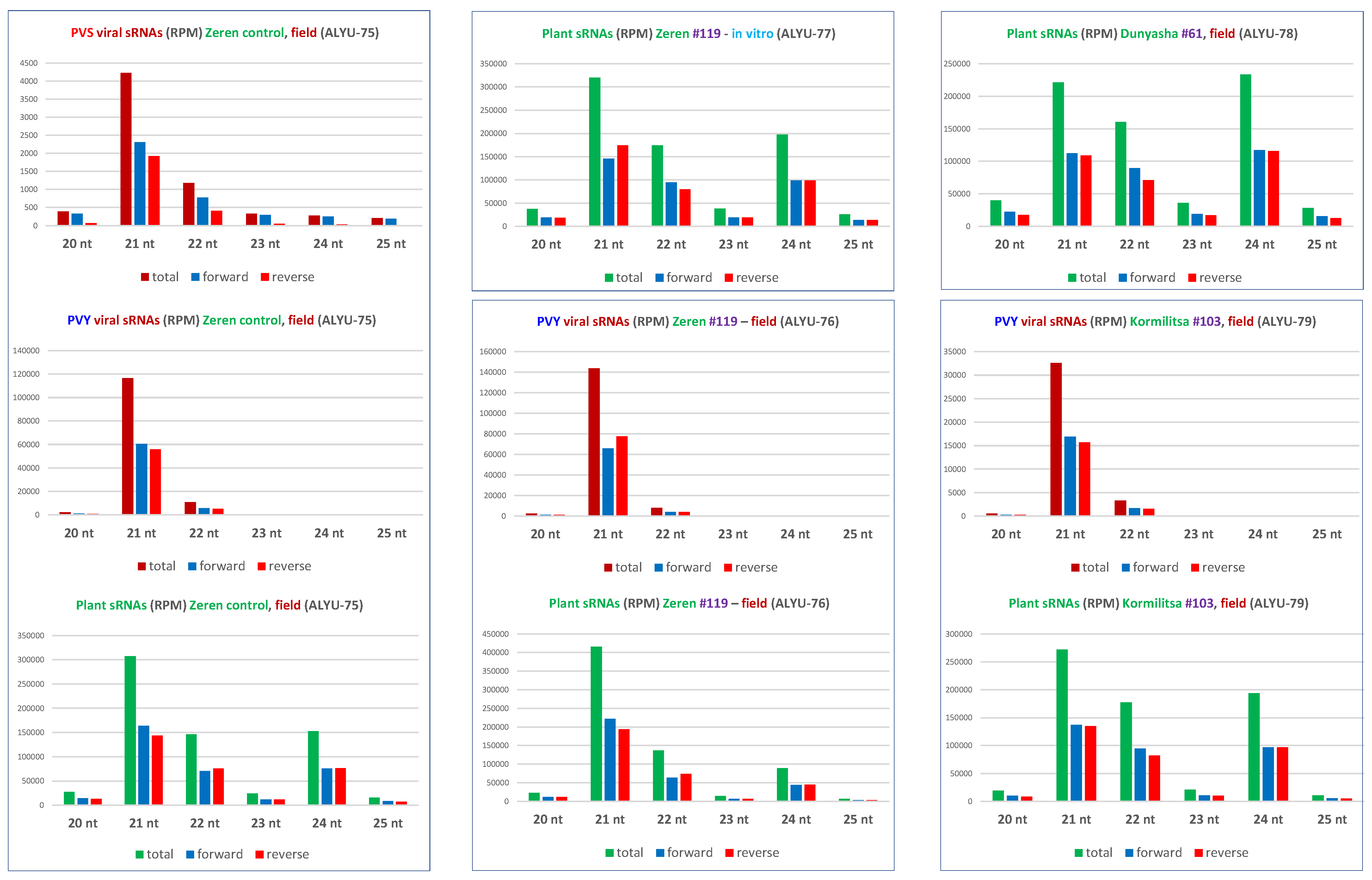
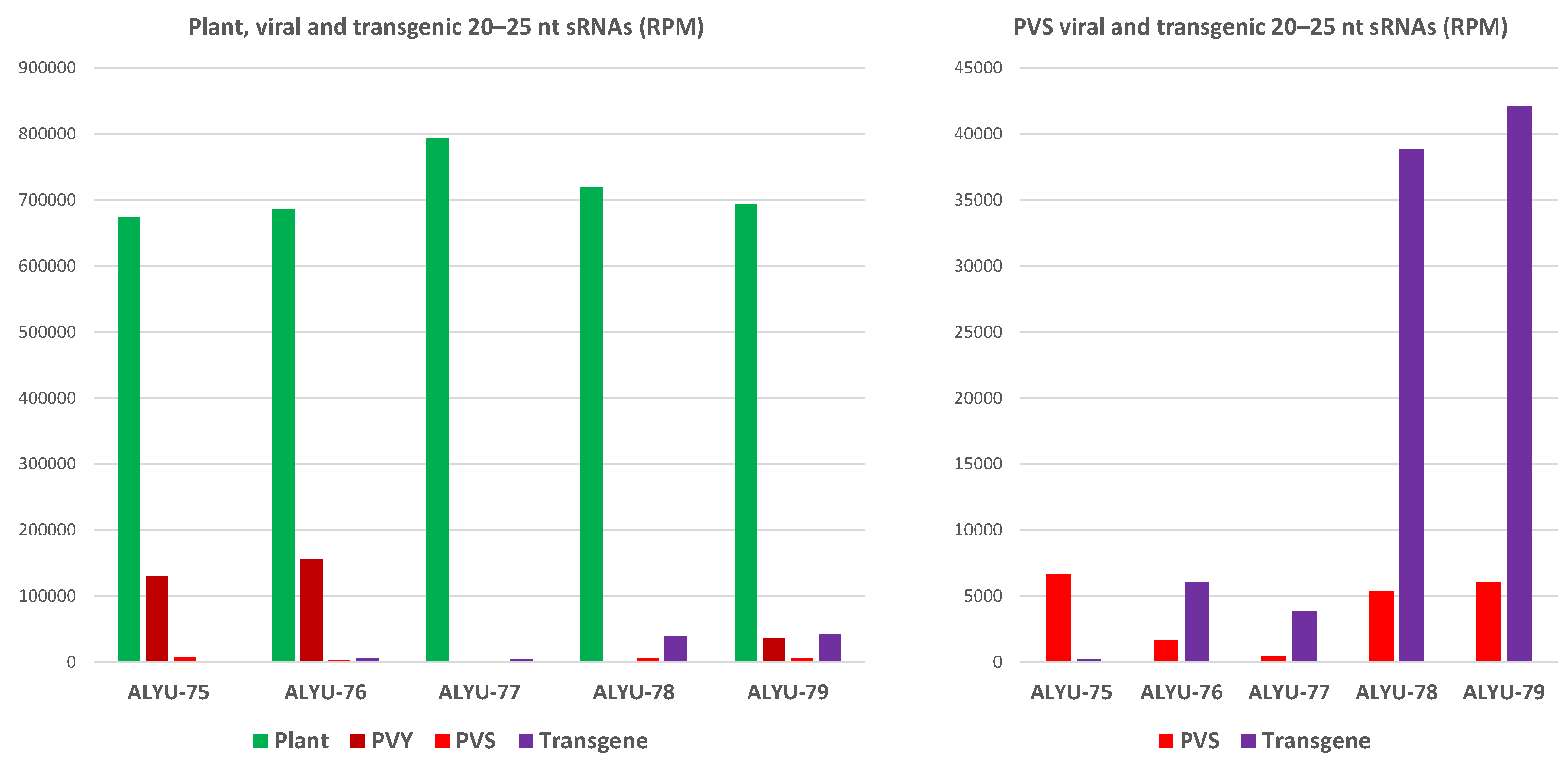
3.3. Small RNA Blot Hybridization and Illumina Sequencing Analyses of Transgenic Plants
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Devaux, A.; Goffart, J.P.; Petsakos, A.; Kromann, P.; Gatto, M.; Okello, J.; Suarez, V.; Hareau, G. Global food security, contributions from sustainable potato agri-food systems. In The Potato Crop; Campos, H., Ortiz, O., Eds.; Springer: Cham, Switzerland, 2020; pp. 3–35. [Google Scholar]
- Gong, H.; Igiraneza, C.; Dusengemungu, L. Major in vitro techniques for potato virus elimination and post eradication detection methods. Am. J. Potato Res. 2019, 96, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Awasthi, L.P.; Verma, H.N. Current status of viral diseases of potato and their ecofriendly management—A critical review. Virol. Res. Rev. 2017, 1, 1–16. [Google Scholar]
- Kreuze, J.F.; Souza-Dias, J.A.C.; Jeevalatha, A.; Figueira, A.R.; Valkonen, J.P.T.; Jones, R.A.C. Viral diseases in potato. In The Potato CROP; Campos, H., Ortiz, O., Eds.; Springer: Cham, Switzerland, 2020; pp. 389–430. [Google Scholar]
- Alexandrova, A.M.; Karpova, O.V.; Nargilova, R.M.; Kryldakov, R.V.; Nizkorodova, A.S.; Zhigaylov, A.V.; Yekaterinskaya, E.M.; Kushnarenko, S.V.; Akbergenov, R.Z.; Iskakov, B.K. Distribution of potato (Solanum tuberosum) viruses in Kazakhstan. Int. J. Biol. Chem. 2018, 11, 33–40. [Google Scholar] [CrossRef]
- Wardrop, E.A.; Gray, A.B.; Singh, R.P.; Peterson, J.F. Aphid transmission of potato virus S. Am. Potato J. 1989, 66, 449–459. [Google Scholar] [CrossRef]
- Martelli, G.P.; Adams, M.J.; Kreuze, J.F.; Dolja, V.V. Family Flexiviridae: A case study in virion and genome plasticity. Annu. Rev. Phytopathol. 2007, 45, 73–100. [Google Scholar] [CrossRef] [Green Version]
- Senshu, H.; Yamaji, Y.; Minato, N.; Shiraishi, T.; Maejima, K.; Hashimoto, M.; Miura, C.; Neriya, Y.; Namba, S. A dual strategy for the suppression of host antiviral silencing: Two distinct suppressors for viral replication and viral movement encoded by potato virus M. J. Virol. 2011, 85, 10269–10278. [Google Scholar] [CrossRef] [Green Version]
- Palukaitis, P. Resistance to viruses of potato and their vectors. Plant Pathol. J. 2012, 28, 248–258. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yang, X.; Zhou, G.; Zhang, T. Engineering plant virus resistance: From RNA silencing to genome editing strategies. Plant Biotechnol. J. 2020, 18, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Kuo, Y.W.; Falk, B.W. RNA interference approaches for plant disease control. Biotechniques 2020, 69, 469–477. [Google Scholar] [CrossRef]
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef]
- Ghildiyal, M.; Zamore, P.D. Small silencing RNAs: An expanding universe. Nat. Rev. Genet. 2009, 10, 94–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- TenOever, B.R. The Evolution of Antiviral Defense Systems. Cell Host Microbe 2016, 19, 142–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pooggin, M.M. Small RNA-omics for plant virus identification, virome reconstruction, and viral defense characterization. Front. Microbiol. 2018, 9, 2779. [Google Scholar] [CrossRef]
- Borges, F.; Martienssen, R.A. The expanding world of small RNAs in plants. Nat. Rev. Mol. Cell. Biol. 2015, 16, 727–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matzke, M.A.; Kanno, T.; Matzke, A.J. RNA-Directed DNA Methylation: The Evolution of a Complex Epigenetic Pathway in Flowering Plants. Annu. Rev. Plant Biol. 2015, 66, 243–267. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Qi, Y. RNAi in Plants: An Argonaute-Centered View. Plant Cell 2016, 28, 272–285. [Google Scholar] [CrossRef] [Green Version]
- Carbonell, A.; Carrington, J.C. Antiviral roles of plant ARGONAUTES. Curr. Opin. Plant Biol. 2015, 27, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Zvereva, A.S.; Pooggin, M.M. Silencing and innate immunity in plant defense against viral and non-viral pathogens. Viruses 2012, 4, 2578–2597. [Google Scholar] [CrossRef] [Green Version]
- Pooggin, M.M. How can plant DNA viruses evade siRNA-directed DNA methylation and silencing? Int. J. Mol. Sci. 2013, 14, 15233–15259. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, A.; Carlos, N.; Ruiz, Y.; Callard, D.; Sánchez, Y.; Ochagavía, M.E.; Seguin, J.; Malpica-López, N.; Hohn, T.; Lecca, M.R.; et al. Field Trial and Molecular Characterization of RNAi-Transgenic Tomato Plants That Exhibit Resistance to Tomato Yellow Leaf Curl Geminivirus. Mol. Plant Microbe Interact. 2016, 29, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Pooggin, M.M. RNAi-mediated resistance to viruses: A critical assessment of methodologies. Curr. Opin. Virol. 2017, 26, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Golyaev, V.; Candresse, T.; Rabenstein, F.; Pooggin, M.M. Plant virome reconstruction and antiviral RNAi characterization by deep sequencing of small RNAs from dried leaves. Sci. Rep. 2019, 9, 19268. [Google Scholar] [CrossRef] [PubMed]
- Malpica-López, N.; Rajeswaran, R.; Beknazariants, D.; Seguin, J.; Golyaev, V.; Farinelli, L.; Pooggin, M.M. Revisiting the Roles of Tobamovirus Replicase Complex Proteins in Viral Replication and Silencing Suppression. Mol. Plant Microbe Interact. 2018, 31, 125–144. [Google Scholar] [CrossRef] [Green Version]
- Turco, S.; Golyaev, V.; Seguin, J.; Gilli, C.; Farinelli, L.; Boller, T.; Schumpp, O.; Pooggin, M.M. Small RNA-omics for virome reconstruction and antiviral defense characterization in mixed infections of cultivated Solanum plants. Mol. Plant Microbe Interact. 2018, 31, 707–723. [Google Scholar] [CrossRef] [Green Version]
- Seguin, J.; Otten, P.; Baerlocher, L.; Farinelli, L.; Pooggin, M.M. MISIS-2: A bioinformatics tool for in-depth analysis of small RNAs and representation of consensus master genome in viral quasispecies. J. Virol. Methods 2016, 233, 37–40. [Google Scholar] [CrossRef] [Green Version]
- Seguin, J.; Rajeswaran, R.; Malpica-López, N.; Martin, R.R.; Kasschau, K.; Dolja, V.V.; Otten, P.; Farinelli, L.; Pooggin, M.M. De novo reconstruction of consensus master genomes of plant RNA and DNA viruses from siRNAs. PLoS ONE 2014, 9, e88513. [Google Scholar] [CrossRef]
- Leibman, D.; Wolf, D.; Saharan, V.; Zelcer, A.; Arazi, T.; Yoel, S.; Gaba, V.; Gal-On, A. A high level of transgenic viral small RNA is associated with broad Potyvirus resistance in cucurbits. Mol. Plant-Microbe Interact. 2011, 24, 1220–1238. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Song, G. High-throughput sequencing as an effective approach in profiling small RNAs derived from a hairpin RNA expression vector in woody plants. Plant Sci. 2014, 228, 39–47. [Google Scholar] [CrossRef]
- Zhao, D.; Song, G. Rootstock-to-scion transfer of transgene-derived small interfering RNAs and their effect on virus resistance in nontransgenic sweet cherry. Plant Biotechnol. J. 2014, 12, 1319–1328. [Google Scholar] [CrossRef] [Green Version]
- Montes, C.; Castro, Á.; Barba, P.; Rubio, J.; Sánchez, E.; Carvajal, D.; Aguirre, C.; Tapia, E.; Dell´Orto, P.; Decroocq, V.; et al. Differential RNAi responses of Nicotiana benthamiana individuals transformed with a hairpin-inducing construct during Plum pox virus challenge. Virus Genes 2014, 49, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, H.; Carbonell, A.; Hoyer, J.S.; Fahlgren, N.; Gilbert, K.B.; Takeda, A.; Giampetruzzi, A.; Garcia Ruiz, M.T.; McGinn, M.G.; Lowery, N.; et al. Roles and programming of Arabidopsis ARGONAUTE proteins during Turnip Mosaic Virus infection. PLoS Pathog. 2015, 11, e1004755. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Virus Infection Status before Transformation | Number of Km-Resistant Regenerants | Number of PCR-Positive Regenerants | Number of Carlavirus-free Regenerants | Number of Carlavirus-Free Lines after 3 Years in Field |
|---|---|---|---|---|---|
| Albinka | PVM, PVY | 19 | 8 | 7 | 3 |
| Dunyasha | PVM | 24 | 11 | 4 | 9 |
| Fortuna | PVS, PVM | 10 | 3 | 0 | 2 |
| Kormilitsa | PVS | 5 | 1 | 1 | 1 |
| Orbita | PVM | 6 | 1 | 0 | not tested |
| Picasso | PVM, PVY | 34 | 9 | 3 | 4 |
| Sante | PVM, PVY | 36 | 3 | 1 | 1 |
| Tokhtar | PVM | 80 | 9 | 0 | not tested |
| Tian-Shanskiy | PVM | 25 | 0 | 0 | not tested |
| Ushkonyr | PVS | 0 | 0 | 0 | not tested |
| Zeren | PVS, PVM, PVY | 20 | 5 | 2 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alexandrova, A.; Karpova, O.; Kryldakov, R.; Golyaev, V.; Nargilova, R.; Iskakov, B.; Pooggin, M.M. Virus Elimination from Naturally Infected Field Cultivars of Potato (Solanum tuberosum) by Transgenic RNA Interference. Int. J. Mol. Sci. 2022, 23, 8020. https://doi.org/10.3390/ijms23148020
Alexandrova A, Karpova O, Kryldakov R, Golyaev V, Nargilova R, Iskakov B, Pooggin MM. Virus Elimination from Naturally Infected Field Cultivars of Potato (Solanum tuberosum) by Transgenic RNA Interference. International Journal of Molecular Sciences. 2022; 23(14):8020. https://doi.org/10.3390/ijms23148020
Chicago/Turabian StyleAlexandrova, Alyona, Oxana Karpova, Ruslan Kryldakov, Victor Golyaev, Rufina Nargilova, Bulat Iskakov, and Mikhail M. Pooggin. 2022. "Virus Elimination from Naturally Infected Field Cultivars of Potato (Solanum tuberosum) by Transgenic RNA Interference" International Journal of Molecular Sciences 23, no. 14: 8020. https://doi.org/10.3390/ijms23148020
APA StyleAlexandrova, A., Karpova, O., Kryldakov, R., Golyaev, V., Nargilova, R., Iskakov, B., & Pooggin, M. M. (2022). Virus Elimination from Naturally Infected Field Cultivars of Potato (Solanum tuberosum) by Transgenic RNA Interference. International Journal of Molecular Sciences, 23(14), 8020. https://doi.org/10.3390/ijms23148020