
Uptake of Phosphate, Calcium, and Vitamin D by the Pregnant Uterus of Sheep in Late Gestation: Regulation by Chorionic Somatomammotropin Hormone

 , , and
, , and 
Abstract
1. Introduction
2. Results
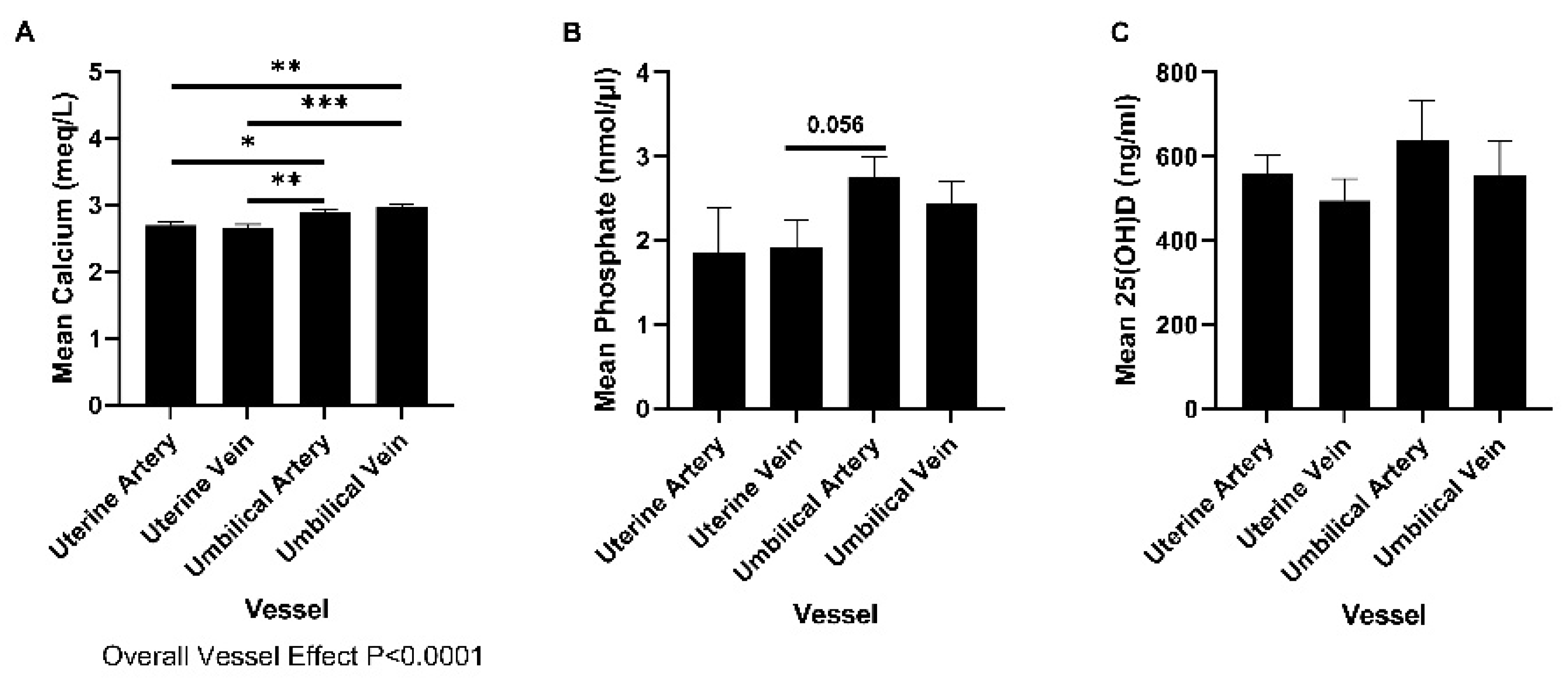
2.1. Concentrations of Phosphate, Calcium, and 25(OH)D in Uterine and Umbilical Blood
2.2. The Effect of CSH RNAi on Calcium, Phosphate, and Vitamin D in Plasma
2.2.1. Calcium
2.2.2. Phosphate
2.2.3. 25(OH)D
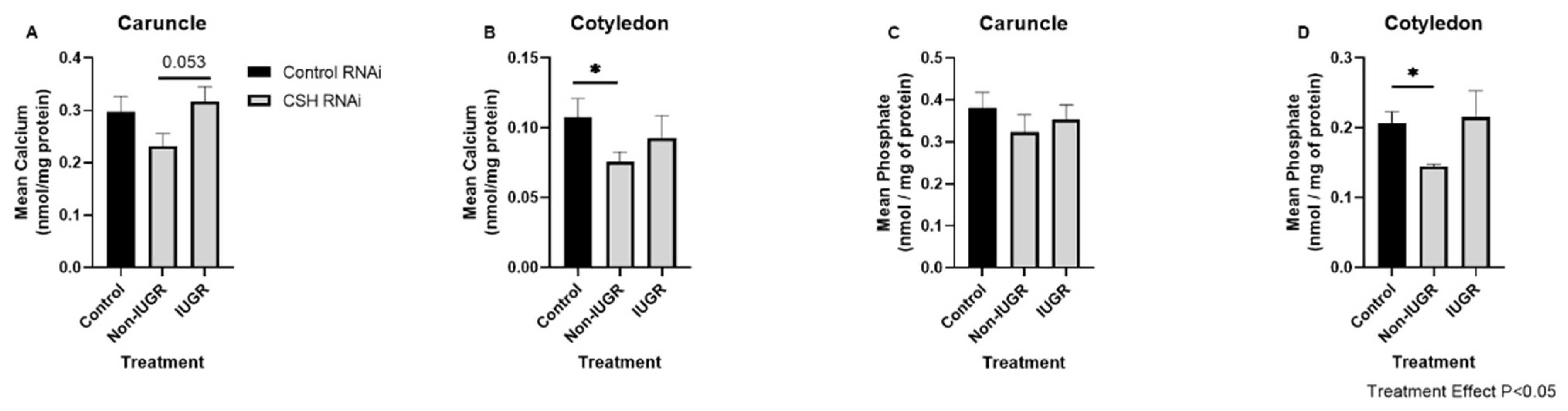
2.3. Effect of CSH RNAi on Abundances of Calcium and Phosphate in Cotyledons and Caruncles
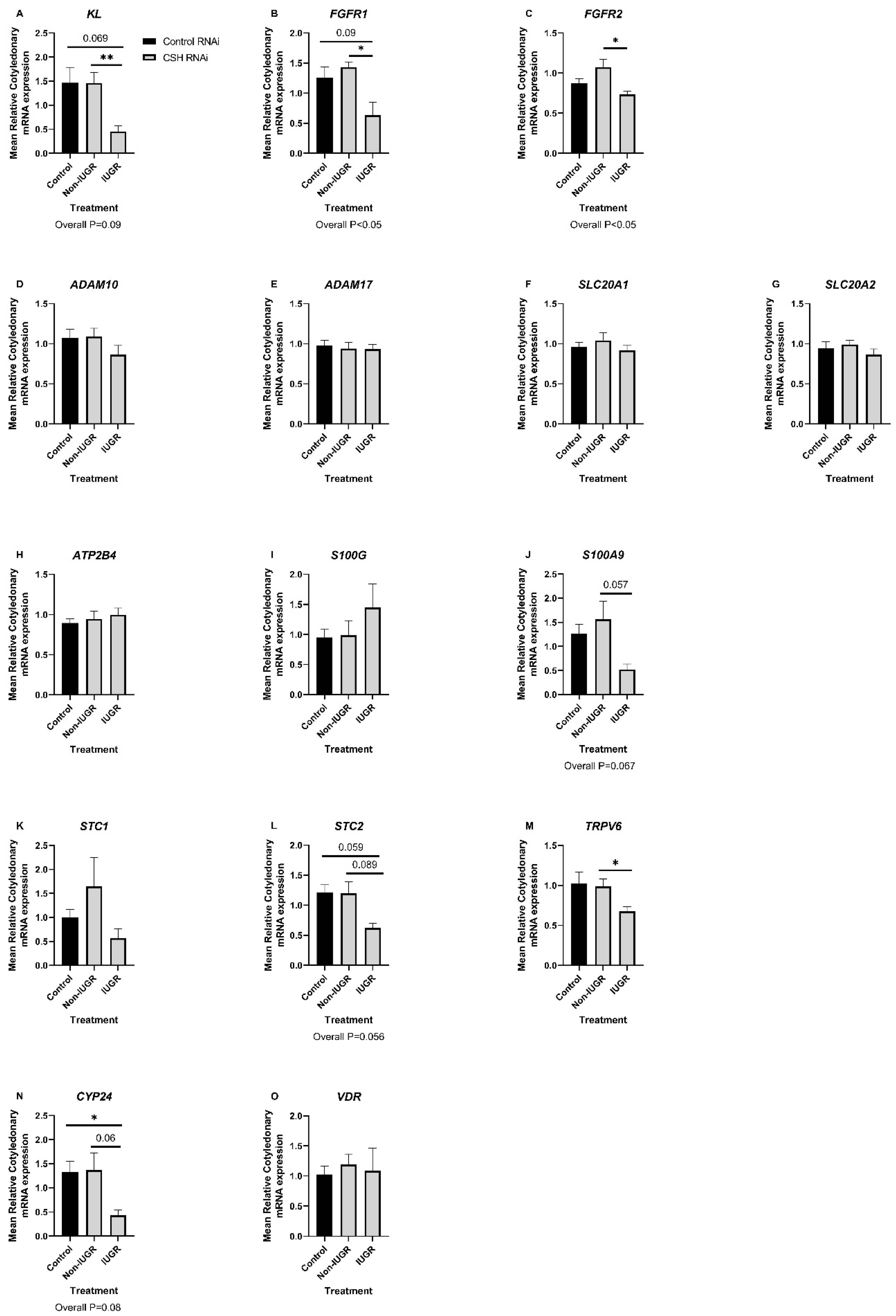
2.4. Effect of CSH RNAi on the Expression of Candidate mRNAs with Roles in the Regulation of Calcium, Phosphate, and Vitamin D Signaling in Caruncles and Cotyledons
2.5. Correlations between Biometric Data and Caruncular and Cotyledonary Phosphate and Calcium Abundance
2.6. Correlations between Biometric Data and Calcium, Phosphate, and Vitamin D Utilization
2.7. Correlations between Biometric Data and the Caruncular and Cotyledonary mRNA Expression of Calcium, Phosphate, and Vitamin D Regulatory Molecules
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Sample Collection
4.2. Quantification of Calcium, Phosphate, and 25(OH)D in Uterine Flushings, Plasma, and Endometrial Homogenates
4.3. Analysis of Candidate Gene Expression by qPCR
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kovacs, C.S. Bone Development and Mineral Homeostasis in the Fetus and Neonate: Roles of the Calciotropic and Phosphotropic Hormones. Physiol. Rev. 2014, 94, 1143–1218. [Google Scholar] [CrossRef] [PubMed]
- Santella, L. The Role of Calcium in the Cell Cycle: Facts and Hypotheses. Biochem. Biophys. Res. Commun. 1998, 244, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Brostrom, M.A.; Brostrom, C.O. Calcium Dynamics and Endoplasmic Reticular Function in the Regulation of Protein Synthesis: Implications for Cell Growth and Adaptability. Cell Calcium 2003, 34, 345–363. [Google Scholar] [CrossRef]
- Chin, K.V.; Cade, C.; Brostrom, C.O.; Galuska, E.M.; Brostrom, M.A. Calcium-Dependent Regulation of Protein Synthesis at Translational Initiation in Eukaryotic Cells. J. Biol. Chem. 1987, 262, 16509–16514. [Google Scholar] [CrossRef]
- Glancy, B.; Balaban, R.S. Role of Mitochondrial Ca2+ in the Regulation of Cellular Energetics. Biochemistry 2012, 51, 2959–2973. [Google Scholar] [CrossRef]
- Jeon, U.S. Kidney and Calcium Homeostasis. Electrolyte Blood Press. 2008, 6, 68–76. [Google Scholar] [CrossRef]
- Penido, M.G.M.G.; Alon, U.S. Phosphate Homeostasis and Its Role in Bone Health. Pediatr. Nephrol. 2012, 27, 2039–2048. [Google Scholar] [CrossRef]
- Nabeshima, Y.I.; Imura, H. A-Klotho: A Regulator That Integrates Calcium Homeostasis. Am. J. Nephrol. 2008, 28, 455–464. [Google Scholar] [CrossRef]
- Khalil, R.; Ri, N.; Jardi, F.; Vanderschueren, D.; Claessens, F.; Decallonne, B. Sex Steroids and the Kidney: Role in Renal Calcium and Phosphate Handling. Mol. Cell. Endocrinol. 2018, 465, 61–72. [Google Scholar] [CrossRef]
- Bagur, R.; Hajnoczky, G. Intracellular Ca2+ Sensing: Role in Calcium Homeostasis and Signaling. Mol. Cell 2017, 66, 780–788. [Google Scholar] [CrossRef]
- Stafford, N.; Wilson, C.; Oceandy, D.; Neyses, L.; Cartwright, E.J. The Plasma Membrane Calcium ATPases and Their Role as Major New Players in Human Disease. Physiol. Rev. 2017, 97, 1089–1125. [Google Scholar] [CrossRef] [PubMed]
- Hermann, A.; Donato, R.; Weiger, T.M.; Chazin, W.J. S100 Calcium Binding Proteins and Ion Channels. Front. Pharmacol. 2012, 3, 67. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S. Transient Receptor Potential Channels. Adv. Exp. Med. Biol. 2011, 704, 218. [Google Scholar] [CrossRef]
- Segawa, H.; Shiozaki, Y.; Kaneko, I.; Miyamoto, K.I. The Role of Sodium-Dependent Phosphate Transporter in Phosphate Homeostasis. J. Nutr. Sci. Vitaminol. 2015, 61, S119–S121. [Google Scholar] [CrossRef] [PubMed]
- Lederer, E. Renal Phosphate Transporters. Curr. Opin. Nephrol. Hypertens. 2014, 23, 502–506. [Google Scholar] [CrossRef]
- Stenhouse, C.; Halloran, K.M.; Newton, M.G.; Gaddy, D.; Suva, L.J.; Bazer, F.W. Novel Mineral Regulatory Pathways in Ovine Pregnancy: 1. Phosphate, Klotho Signaling, and Sodium Dependent Phosphate Transporters. Biol. Reprod. 2021, 104, 1084–1096. [Google Scholar] [CrossRef]
- Stenhouse, C.; Halloran, K.M.; Newton, M.G.; Gaddy, D.; Suva, L.J.; Bazer, F.W. Novel Mineral Regulatory Pathways in Ovine Pregnancy: II. Calcium Binding Proteins, Calcium Transporters, and Vitamin D Signaling. Biol. Reprod. 2021, 105, 232–243. [Google Scholar] [CrossRef]
- Stenhouse, C.; Suva, L.J.; Gaddy, D.; Wu, G.; Bazer, F.W. Phosphate, Calcium, and Vitamin D: Key Regulators of Fetal and Placental Development in Mammals. Adv. Exp. Med. Biol. 2022, 1354, 77–107. [Google Scholar] [CrossRef]
- Stenhouse, C.; Halloran, K.; Moses, R.; Seo, H.; Gaddy, D.; Johnson, G.; Wu, G.; Suva, L.; Bazer, F. Effects of Progesterone and Interferon Tau on Ovine Endometrial Phosphate, Calcium, and Vitamin D Signaling. Biol. Reprod. 2022, 106, 888–899. [Google Scholar] [CrossRef]
- Stenhouse, C.; Halloran, K.; Hoskins, E.; Newton, M.; Moses, R.; Seo, H.; Dunlap, K.; Satterfield, M.; Gaddy, D.; Johnson, G.; et al. Effects of Exogenous Progesterone on Expression of Mineral Regulatory Molecules by Ovine Endometrium and Placentomes. Biol. Reprod. 2022, 106, 1126–1142. [Google Scholar] [CrossRef]
- Martal, J.; Djiane, J. The Production of Chorionic Somatomammotrophin in Sheep. J. Reprod. Fertil. 1977, 49, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Wooding, F.B.P.; Morgan, G.; Forsyth, I.A.; Butcher, G.; Hutchings, A.; Billingsley, S.A.; Gluckman, P.D. Light and Electron Microscopic Studies of Cellular Localization of OPL with Monoclonal and Polyclonal Antibodies. J. Histochem. Cytochem. 1992, 40, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.; Robertson, H.; Friesen, H. Maternal and Fetal Concentrations of Ovine Placental Lactogen Measured by Radioimmunoassay. Endocrinology 1978, 102, 1606–1613. [Google Scholar] [CrossRef] [PubMed]
- Tanner, A.R.; Lynch, C.S.; Kennedy, V.C.; Ali, A.; Winger, Q.A.; Rozance, P.J.; Anthony, R.V. CSH RNA Interference Reduces Global Nutrient Uptake and Umbilical Blood Flow Resulting in Intrauterine Growth Restriction. Int. J. Mol. Sci. 2021, 22, 8150. [Google Scholar] [CrossRef]
- Tanner, A.R.; Lynch, C.S.; Ali, A.; Winger, Q.A.; Rozance, P.J.; Anthony, R.V. Impact of Chorionic Somatomammotropin RNA Interference on Uterine Blood Flow and Placental Glucose Uptake in the Absence of Intrauterine Growth Restriction. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2021, 320, 138–148. [Google Scholar] [CrossRef]
- Weatherley, A.J.; Ross, R.; Pickard, D.W.; Care, A.D. The Transfer of Calcium during Perfusion of the Placenta in Intact and Thyroparathyroidectomized Sheep. Placenta 1983, 4, 271–278. [Google Scholar] [CrossRef]
- Care, A.; Caple, I.; Abbas, S.; Pickard, D. The Effect of Fetal Thyroparathyroidectomy on the Transport of Calcium across the Ovine Placenta to the Fetus. Placenta 1986, 7, 417–424. [Google Scholar] [CrossRef]
- Symonds, H.; Sansom, B.; Twardock, A. The Measurement of the Transfer of Calcium and Phosphorus from Foetus to Dam in the Sheep Using a Whole Body Counter. Res. Vet. Sci. 1972, 13, 272–275. [Google Scholar] [CrossRef]
- Burstyn, P.; Lloyd, I.; McKillop, W. The Effect of Human Placental Lactogen on the Renal Excretion of Calcium in the Rabbit. IRCS J. Med. Sci. 1975, 3, 30. [Google Scholar]
- Dvir, R.; Golander, A.; Jaccard, N.; Yedwab, G.; Otremski, I.; Spirer, Z.; Weisman, Y. Amniotic Fluid and Plasma Levels of Parathyrod Hormone-Related Protein and Hormonal Modulation of Its Secretion by Amniotic Fluid Cells. Eur. J. Endocrinol. 1995, 133, 277–282. [Google Scholar] [CrossRef]
- Song, G.; Bazer, F.W.; Wagner, G.F.; Spencer, T.E. Stanniocalcin (STC) in the Endometrial Glands of the Ovine Uterus: Regulation by Progesterone and Placental Hormones. Biol. Reprod. 2006, 74, 913–922. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lea, R.G.; Wooding, P.; Stewart, I.; Hannah, L.T.; Morton, S.; Wallace, K.; Aitken, R.P.; Milne, J.S.; Regnault, T.R.; Anthony, R.V.; et al. The Expression of Ovine Placental Lactogen, StAR and Progesterone-Associated Steroidogenic Enzymes in Placentae of Overnourished Growing Adolescent Ewes. Reproduction 2007, 133, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, Z.; Bick, T.; Perlman, R.; Lahav, M.; Barzilai, D. The Modulation of Placental Lactogen Secretion by Calcium: Studies with Cultured Human Term Trophoblast. Mol. Cell. Biol. 1984, 37, 359–362. [Google Scholar] [CrossRef]
- Burke, J.; Rice, G.; Ralph, M.; Thorburn, G. Effects of Calcium Availability on the Release of Ovine Choriomammoropin from Cotyledonary Cells Incubated in Vitro. Comp. Biochem. Physiol. 1989, 93A, 489–492. [Google Scholar] [CrossRef]
- Renkema, K.Y.; Alexander, R.T.; Bindels, R.J.; Hoenderop, J.G. Calcium and Phosphate Homeostasis: Concerted Interplay of New Regulators. Ann. Med. 2008, 40, 82–91. [Google Scholar] [CrossRef]
- Anthony, R.V.; Limesand, S.W.; Fanning, M.D.; Liang, R. Placental Lactogen and Growth Hormone. Regulation and Action. In Endocrinology of Pregnancy; Bazer, F., Ed.; Humana Press: Totowa, NJ, USA, 1998; pp. 461–490. [Google Scholar]
- Devaskar, U.P.; Ho, M.; Devaskar, S.U.; Tsang, R.C. 25-Hydroxy- and 1 Alpha,25-Dihydroxyvitamin D. Maternal-Fetal Relationship and the Transfer of 1,25-Dihydroxyvitamin D3 across the Placenta in an Ovine Model. Dev. Pharmacol. Ther. 1984, 7, 213–220. [Google Scholar] [CrossRef]
- Ross, R.; Halbert, K.; Tsang, R.C. Determination of the Production and Metabolic Clearance Rates of 1,25-Dihydroxyvitamin D3 in the Pregnant Sheep and Its Chronically Catheterized Fetus by Primed Infusion Technique. Pediatr. Res. 1989, 26, 633–634. [Google Scholar] [CrossRef]
- Jang, H.; Choi, Y.; Yoo, I.; Han, J.; Hong, J.S.; Kim, Y.Y.; Ka, H. Vitamin D-Metabolic Enzymes and Related Molecules: Expression at the Maternal-Conceptus Interface and the Role of Vitamin D in Endometrial Gene Expression in Pigs. PLoS ONE 2017, 12, e0187221. [Google Scholar] [CrossRef]
- Shahbazi, M.; Jeddi-tehrani, M.; Zareie, M.; Salek-moghaddam, A.; Akhondi, M.M.; Bahmanpoor, M.; Sadeghi, M.R.; Zarnani, A.H. Expression Profiling of Vitamin D Receptor in Placenta, Decidua and Ovary of Pregnant Mice. Placenta 2011, 32, 657–664. [Google Scholar] [CrossRef]
- O’Brien, K.O.; Li, S.; Cao, C.; Kent, T.; Young, B.V.; Queenan, R.A.; Pressman, E.K.; Cooper, E.M. Placental CYP27B1 and CYP24A1 Expression in Human Placental Tissue and Their Association With Maternal and Neonatal Calcitropic Hormones. Endocr. Res. 2014, 99, 1348–1356. [Google Scholar] [CrossRef][Green Version]
- Bergada, L.; Pallares, J.; Arcidiacono, M.; Cardus, A.; Santacan, M.; Valls, J.; Cao, G.; Fernandez, E.; Dolcet, X.; Dusso, A.; et al. Role of Local Bioactivation of Vitamin D by CYP27A1 and CYP2R1 in the Control of Cell Growth in Normal Endometrium and Endometrial Carcinoma. Lab. Investig. 2014, 94, 608–622. [Google Scholar] [CrossRef] [PubMed]
- Spanos, E.; Brown, D.; Stevenson, J.; Macintyre, I. Stimulation of 1,25-Dihydroxycholecalciferol Production by Prolactin and Related Peptides in Intact Renal Cell Preparations in Vitro. Biochim. Biophys. Acta 1981, 672, 7–15. [Google Scholar] [CrossRef]
- Takeuchi, K.; Morikawa, H.; Ueda, Y.; Mochizuki, M. Studies on the Effects of Placental Lactogen on Calcium Metabolism during Pregnancy. Nihon Naibunpi Gakkai Zasshi 1988, 64, 1175–1186. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bikle, D. Vitamin D Metabolism, Mechanism of Action, and Clinical Applications. Chem. Biol. 2014, 21, 319–329. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| CSH RNAi | Overall p Value | |||
|---|---|---|---|---|
| Control RNAi | Non-IUGR | IUGR | ||
| Uterine | ||||
| Uterine Arterial Calcium (meq/L) | 2.694 ± 0.054 | 2.594 ± 0.026 | 2.776 ± 0.029 | >0.10 |
| Uterine Venous Calcium (meq/L) | 2.654 ± 0.052 | 2.538 ± 0.026 | 2.718 ± 0.016 | 0.09 |
| Uterine Artery-Vein Calcium Gradient (meq/L) | 0.040 ± 0.007 | 0.056 ± 0.002 | 0.058 ± 0.016 | >0.10 |
| Uterine Calcium Uptake (meq/min) | 0.071 ± 0.012 ab | 0.080 ± 0.004 a | 0.057 ± 0.010 b | >0.10 |
| Uterine Calcium Uptake per kg of Uterus (meq/min) | 0.109 ± 0.023 | 0.133 ± 0.004 | 0.112 ± 0.014 | >0.10 |
| Uterine Calcium Uptake per kg Fetus (meq/min) | 0.019 ± 0.003 | 0.021 ± 0.001 | 0.021 ± 0.003 | >0.10 |
| Uterine Calcium Uptake per kg Placenta (meq/min) | 0.161 ± 0.025 | 0.209 ± 0.009 | 0.172 ± 0.032 | >0.10 |
| Umbilical | ||||
| Umbilical Arterial Calcium (meq/L) | 2.896 ± 0.042 | 2.878 ± 0.064 | 2.899 ± 0.104 | >0.10 |
| Umbilical Venous Calcium (meq/L) | 2.977 ± 0.042 | 2.937 ± 0.064 | 2.984 ± 0.107 | >0.10 |
| Umbilical Vein-Artery Gradient (meq/L) | 0.081 ± 0.005 a | 0.060 ± 0.009 b | 0.086 ± 0.004 ab | <0.05 |
| Umbilical Calcium Uptake (meq/min) | 0.063 ± 0.006 a | 0.048 ± 0.008 ab | 0.038 ± 0.008 b | 0.089 |
| Umbilical Calcium Uptake per kg of Uterus (meq/min) | 0.089 ± 0.011 | 0.078 ± 0.013 | 0.075 ± 0.014 | >0.10 |
| Umbilical Calcium Uptake per kg Fetus (meq/min) | 3.759 ± 0.7171 | 2.325 ± 0.4301 | 2.078 ± 0.695 | >0.10 |
| Umbilical Calcium Uptake per kg Placenta (meq/min) | 0.145 ± 0.016 | 0.123 ± 0.019 | 0.105 ± 0.007 | >0.10 |
| Uteroplacental | ||||
| Uteroplacental Calcium Utilization (meq/L) | 0.008 ± 0.016 | 0.032 ± 0.006 | 0.019 ± 0.012 | >0.10 |
| CSH RNAi | Overall p Value | |||
|---|---|---|---|---|
| Control RNAi | Non-IUGR | IUGR | ||
| Uterine | ||||
| Uterine Arterial 25(OH)D (ng/mL) | 560.5 ± 43.74 a | N/A | 322.9 ± 68.68 b | <0.05 |
| Uterine Venous 25(OH)D (ng/mL) | 495.0 ± 51.87 a | 485.3 ± 52.62 ab | 267.2 ± 28.82 b | <0.05 |
| Uterine Artery-Vein 25(OH)D Gradient (ng/mL) | 61.63 ± 26.96 | N/A | 59.07 ± 28.82 | >0.10 |
| Uterine 25(OH)D Uptake (µg/min) | 38.77 ± 12.81 | N/A | 84.47 ± 33.26 | >0.10 |
| Uterine 25(OH)D Uptake per kg of Uterus (µg/min) | 85.59 ± 36.63 | N/A | 157.7 ± 58.69 | >0.10 |
| Uterine 25(OH)D Uptake per kg Fetus (µg/min) | 21.74 ± 12.31 | N/A | 25.71 ± 9.403 | >0.10 |
| Uterine 25(OH)D Uptake per kg Placenta (µg/min) | 192.0 ± 113.6 | N/A | 204.2 ± 87.33 | >0.10 |
| Umbilical | ||||
| Umbilical Arterial 25(OH)D (ng/mL) | 639.7 ± 93.39 | 683.3 ± 127.6 | 830.3 ± 230.2 | >0.10 |
| Umbilical Venous 25(OH)D (ng/mL) | 555.4 ± 80.09 | 573.0 ± 74.25 | 586.9 ± 170.4 | >0.10 |
| Umbilical Vein-Artery 25(OH)D Gradient (ng/mL) | −84.26 ± 71.34 | −110.3 ± 132.6 | −243.4 ± 104.3 | >0.10 |
| Umbilical 25(OH)D Uptake (µg/min) | −63.13 ± 51.85 | −63.28 ± 90.15 | −227.4 ± 105.2 | >0.10 |
| Umbilical 25(OH)D Uptake per kg of Uterus (µg/min) | −99.51 ± 62.51 | −118.7 ± 150.5 | −468.5 ± 226.8 | >0.10 |
| Umbilical 25(OH)D Uptake per kg Fetus (µg/min) | −16.92 ± 13.12 | −14.89 ± 22.05 | −98.13 ± 55.18 | 0.09 |
| Umbilical 25(OH)D Uptake per kg Placenta (µg/min) | −162.9 ± 122.0 | −165.6 ± 234.2 | −723.6 ± 434.9 | >0.10 |
| Uteroplacental | ||||
| Uteroplacental 25(OH)D Utilization (µg/min) | 155.3 ± 84.27 | N/A | 252.9 ± 139.7 | >0.10 |
| Biometric Parameter | mRNA | Caruncle | Cotyledon | ||
|---|---|---|---|---|---|
| r | p-Value | r | p-Value | ||
| Maternal Weight | KL | 0.098 | 0.205 | 0.083 | 0.232 |
| Maternal Weight | FGFR1 | 0.001 | 0.896 | 0.116 | 0.142 |
| Maternal Weight | FGFR2 | 0.070 | 0.261 | 0.389 | 0.006 |
| Maternal Weight | ADAM10 | 0.003 | 0.815 | 0.158 | 0.083 |
| Maternal Weight | ADAM17 | 0.021 | 0.538 | 0.005 | 0.783 |
| Maternal Weight | SLC20A1 | 0.060 | 0.299 | 0.195 | 0.059 |
| Maternal Weight | SLC20A2 | 0.072 | 0.251 | 0.019 | 0.586 |
| Maternal Weight | ATP2B4 | 0.007 | 0.731 | 0.039 | 0.433 |
| Maternal Weight | PTHrP | 0.262 | 0.025 | N/A | N/A |
| Maternal Weight | S100G | 0.049 | 0.346 | 0.024 | 0.522 |
| Maternal Weight | S100A9 | N/A | N/A | 0.086 | 0.223 |
| Maternal Weight | STC1 | 0.233 | 0.042 | 0.105 | 0.205 |
| Maternal Weight | STC2 | 0.059 | 0.314 | 0.093 | 0.203 |
| Maternal Weight | TRPV6 | 0.022 | 0.535 | 0.091 | 0.210 |
| Maternal Weight | CYP24 | 0.050 | 0.341 | 0.115 | 0.168 |
| Maternal Weight | VDR | 0.014 | 0.615 | 0.012 | 0.645 |
| Uterine Weight | KL | 0.058 | 0.338 | 0.184 | 0.067 |
| Uterine Weight | FGFR1 | 0.025 | 0.519 | 0.009 | 0.699 |
| Uterine Weight | FGFR2 | 0.002 | 0.851 | 0.004 | 0.803 |
| Uterine Weight | ADAM10 | 0.226 | 0.034 | 0.008 | 0.714 |
| Uterine Weight | ADAM17 | 0.140 | 0.105 | 0.001 | 0.880 |
| Uterine Weight | SLC20A1 | 0.085 | 0.214 | 0.001 | 0.879 |
| Uterine Weight | SLC20A2 | 0.008 | 0.710 | 0.127 | 0.147 |
| Uterine Weight | ATP2B4 | 0.185 | 0.058 | 0.045 | 0.398 |
| Uterine Weight | PTHrP | 0.177 | 0.072 | N/A | N/A |
| Uterine Weight | S100G | 0.0003 | 0.952 | 7.535 × 10−5 | 0.972 |
| Uterine Weight | S100A9 | N/A | N/A | 0.060 | 0.312 |
| Uterine Weight | STC1 | 0.199 | 0.06 | 0.058 | 0.351 |
| Uterine Weight | STC2 | 0.0003 | 0.942 | 0.018 | 0.584 |
| Uterine Weight | TRPV6 | 0.023 | 0.524 | 0.094 | 0.201 |
| Uterine Weight | CYP24 | 0.024 | 0.511 | 0.039 | 0.432 |
| Uterine Weight | VDR | 0.080 | 0.226 | 0.091 | 0.197 |
| Fetal Weight | KL | 0.06 | 0.326 | 0.222 | 0.041 |
| Fetal Weight | FGFR1 | 0.055 | 0.333 | 0.194 | 0.05 |
| Fetal Weight | FGFR2 | 0.041 | 0.394 | 0.120 | 0.160 |
| Fetal Weight | ADAM10 | 0.161 | 0.079 | 0.076 | 0.238 |
| Fetal Weight | ADAM17 | 0.005 | 0.767 | 0.002 | 0.854 |
| Fetal Weight | SLC20A1 | 0.241 | 0.028 | 0.004 | 0.806 |
| Fetal Weight | SLC20A2 | 0.167 | 0.074 | 0.090 | 0.227 |
| Fetal Weight | ATP2B4 | 0.09 | 0.20 | 0.115 | 0.170 |
| Fetal Weight | PTHrP | 0.281 | 0.02 | N/A | N/A |
| Fetal Weight | S100G | 0.007 | 0.725 | 0.002 | 0.854 |
| Fetal Weight | S100A9 | N/A | N/A | 0.225 | 0.04 |
| Fetal Weight | STC1 | 0.130 | 0.142 | 0.209 | 0.065 |
| Fetal Weight | STC2 | 0.002 | 0.866 | 0.202 | 0.053 |
| Fetal Weight | TRPV6 | 4.328 × 10−5 | 0.978 | 0.002 | 0.863 |
| Fetal Weight | CYP24 | 0.139 | 0.105 | 0.105 | 0.190 |
| Fetal Weight | VDR | 5.289 × 10−6 | 0.992 | 0.224 | 0.035 |
| Placental Weight | KL | 0.049 | 0.376 | 0.045 | 0.383 |
| Placental Weight | FGFR1 | 0.015 | 0.615 | 0.045 | 0.369 |
| Placental Weight | FGFR2 | 8.461 × 10−6 | 0.990 | 0.031 | 0.486 |
| Placental Weight | ADAM10 | 0.159 | 0.08 | 3.854 × 10−5 | 0.979 |
| Placental Weight | ADAM17 | 0.054 | 0.325 | 0.002 | 0.848 |
| Placental Weight | SLC20A1 | 0.288 | 0.015 | 0.034 | 0.451 |
| Placental Weight | SLC20A2 | 0.004 | 0.789 | 0.002 | 0.878 |
| Placental Weight | ATP2B4 | 0.084 | 0.215 | 0.068 | 0.296 |
| Placental Weight | PTHrP | 0.099 | 0.190 | N/A | N/A |
| Placental Weight | S100G | 0.032 | 0.451 | 0.014 | 0.625 |
| Placental Weight | S100A9 | N/A | N/A | 0.022 | 0.548 |
| Placental Weight | STC1 | 0.032 | 0.478 | 0.081 | 0.270 |
| Placental Weight | STC2 | 0.028 | 0.492 | 0.077 | 0.249 |
| Placental Weight | TRPV6 | 0.128 | 0.122 | 0.018 | 0.585 |
| Placental Weight | CYP24 | 0.01 | 0.678 | 0.007 | 0.736 |
| Placental Weight | VDR | 0.0006 | 0.918 | 0.208 | 0.043 |
| Parameter | Formula |
|---|---|
| Uterine uptake | uterine blood flow (mL/min) × ([UTA] − [UTV]) |
| Umbilical uptake | umbilical blood flow (mL/min) × ([UMV] − [UMA]) |
| Uteroplacental utilization | uterine uptake–umbilical uptake |
| Uterine uptake relative to uterine, fetal, or placental weight | uterine uptake/uterine, fetal, or placental weight |
| Umbilical uptake relative to uterine, fetal, or placental weight | umbilical uptake/uterine, fetal, or placental weight |
| Uterine artery–uterine vein gradient | [UTA] − [UTV] |
| Umbilical vein–umbilical artery gradient | [UMV] − [UMA] |
| Uterine artery-umbilical artery gradient | [UTA] − [UMA] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stenhouse, C.; Halloran, K.M.; Tanner, A.R.; Suva, L.J.; Rozance, P.J.; Anthony, R.V.; Bazer, F.W. Uptake of Phosphate, Calcium, and Vitamin D by the Pregnant Uterus of Sheep in Late Gestation: Regulation by Chorionic Somatomammotropin Hormone. Int. J. Mol. Sci. 2022, 23, 7795. https://doi.org/10.3390/ijms23147795
Stenhouse C, Halloran KM, Tanner AR, Suva LJ, Rozance PJ, Anthony RV, Bazer FW. Uptake of Phosphate, Calcium, and Vitamin D by the Pregnant Uterus of Sheep in Late Gestation: Regulation by Chorionic Somatomammotropin Hormone. International Journal of Molecular Sciences. 2022; 23(14):7795. https://doi.org/10.3390/ijms23147795
Chicago/Turabian StyleStenhouse, Claire, Katherine M. Halloran, Amelia R. Tanner, Larry J. Suva, Paul J. Rozance, Russell V. Anthony, and Fuller W. Bazer. 2022. "Uptake of Phosphate, Calcium, and Vitamin D by the Pregnant Uterus of Sheep in Late Gestation: Regulation by Chorionic Somatomammotropin Hormone" International Journal of Molecular Sciences 23, no. 14: 7795. https://doi.org/10.3390/ijms23147795
APA StyleStenhouse, C., Halloran, K. M., Tanner, A. R., Suva, L. J., Rozance, P. J., Anthony, R. V., & Bazer, F. W. (2022). Uptake of Phosphate, Calcium, and Vitamin D by the Pregnant Uterus of Sheep in Late Gestation: Regulation by Chorionic Somatomammotropin Hormone. International Journal of Molecular Sciences, 23(14), 7795. https://doi.org/10.3390/ijms23147795