
Effect of Polymeric Nanoparticles with Entrapped Fish Oil or Mupirocin on Skin Wound Healing Using a Porcine Model

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
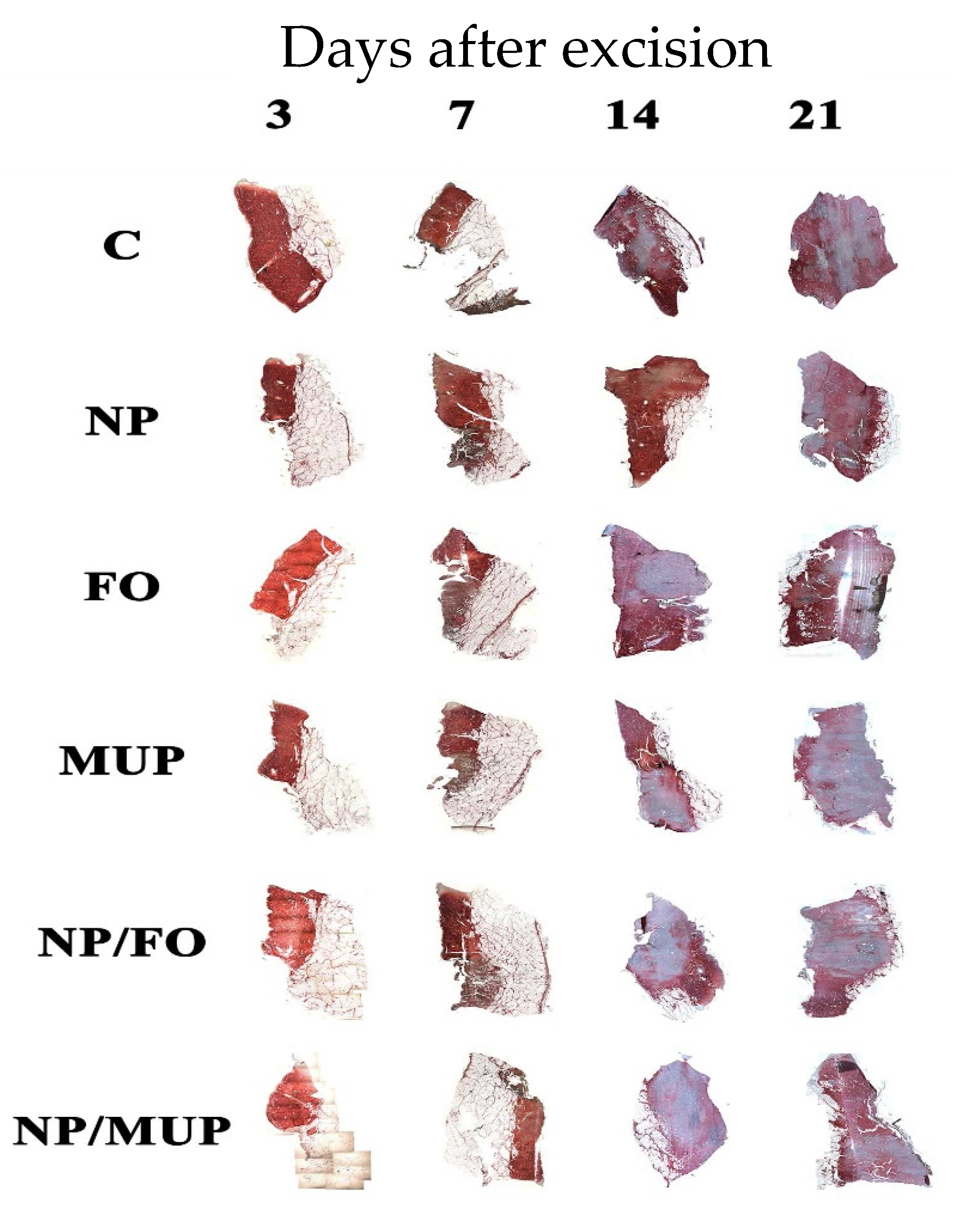
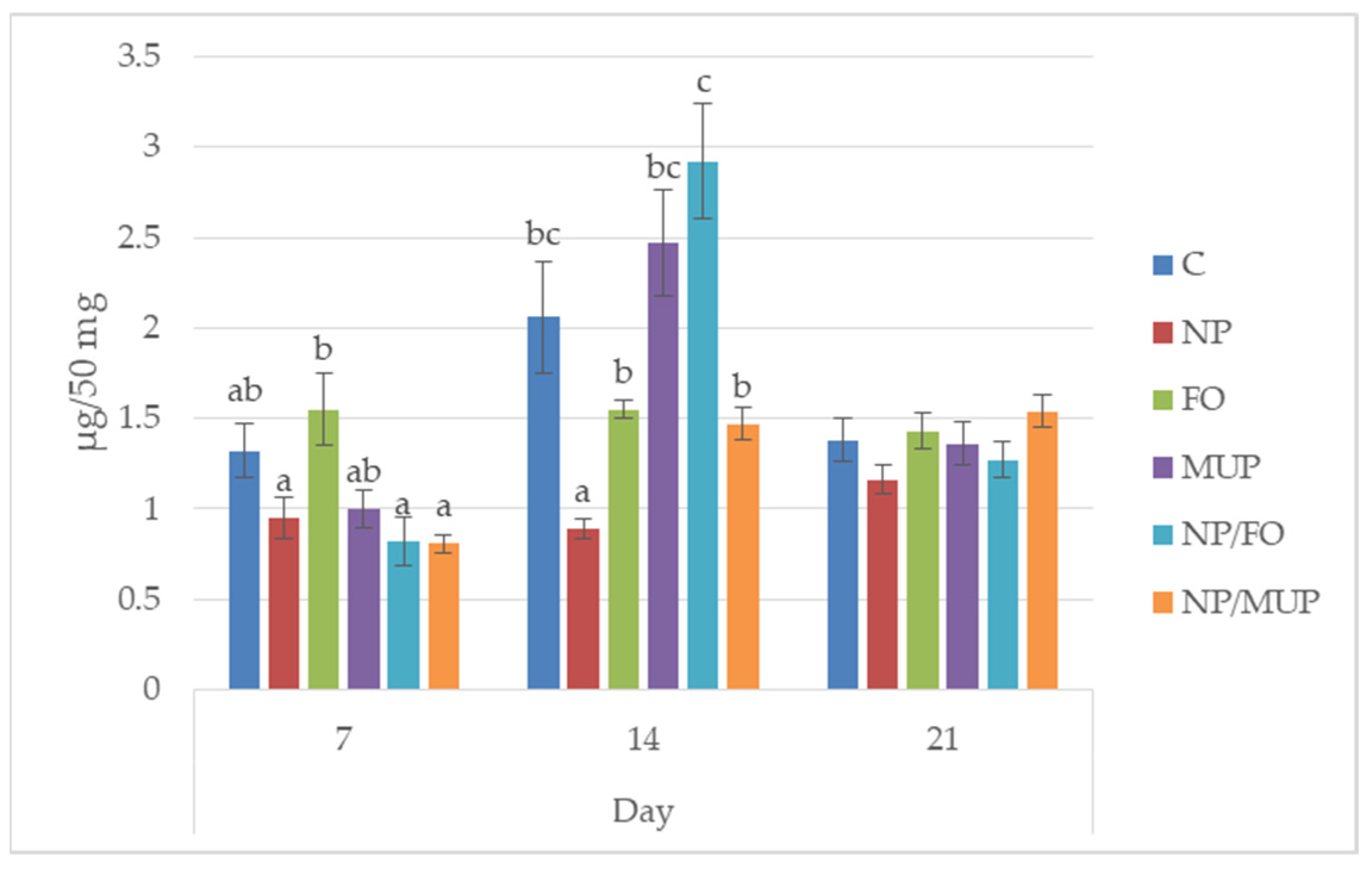
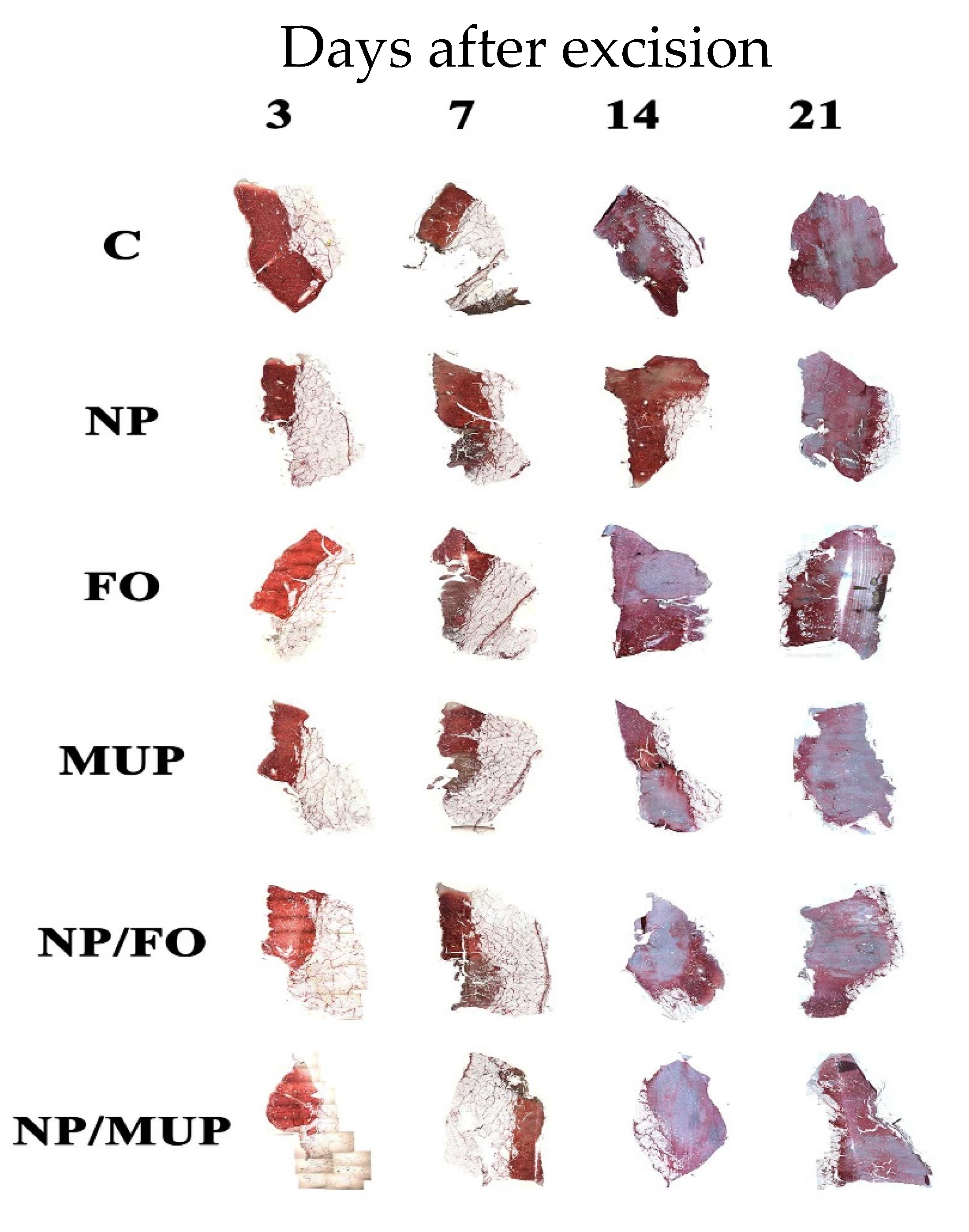
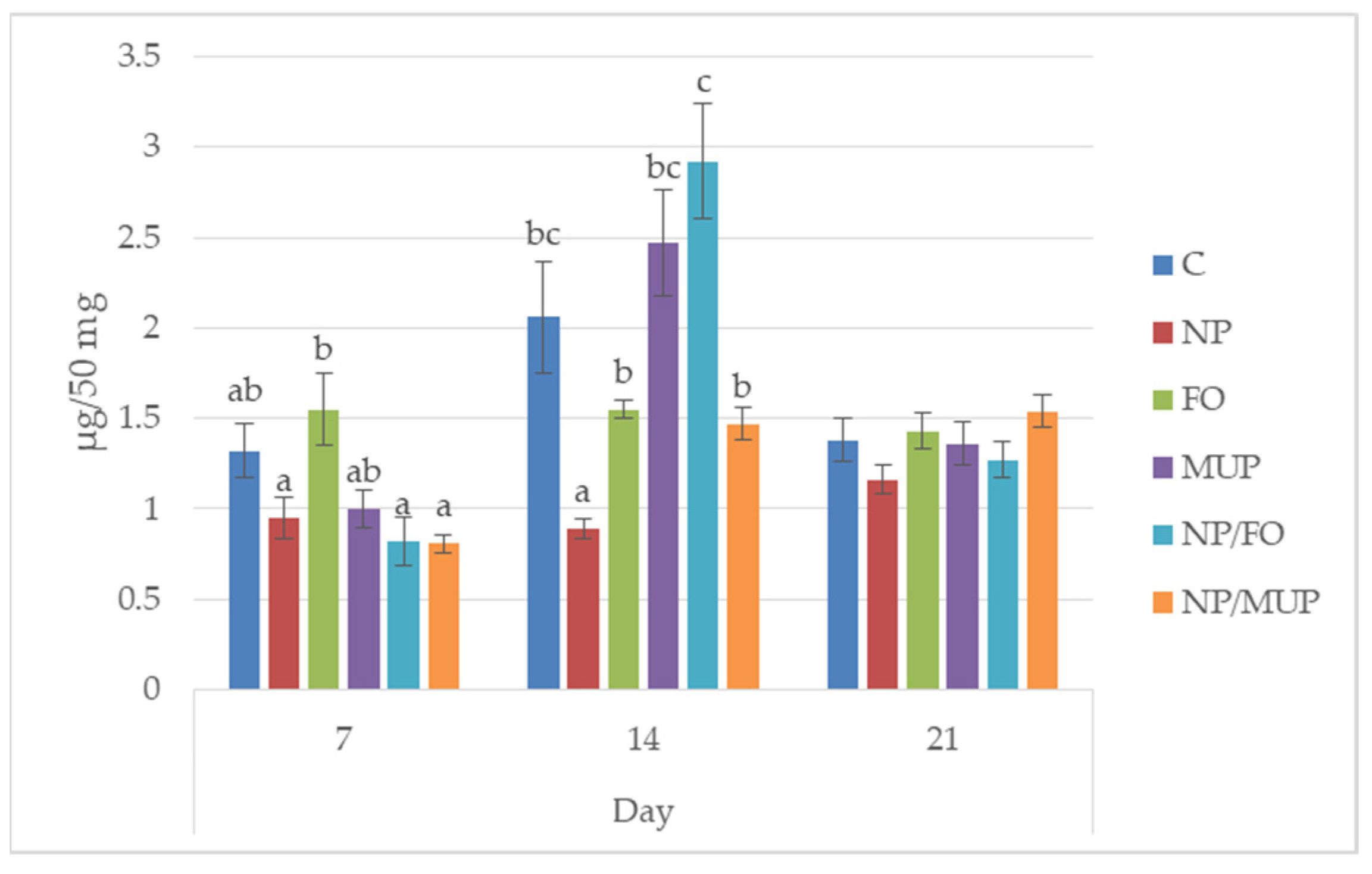
2.1. Collagenous Tissue Maturation
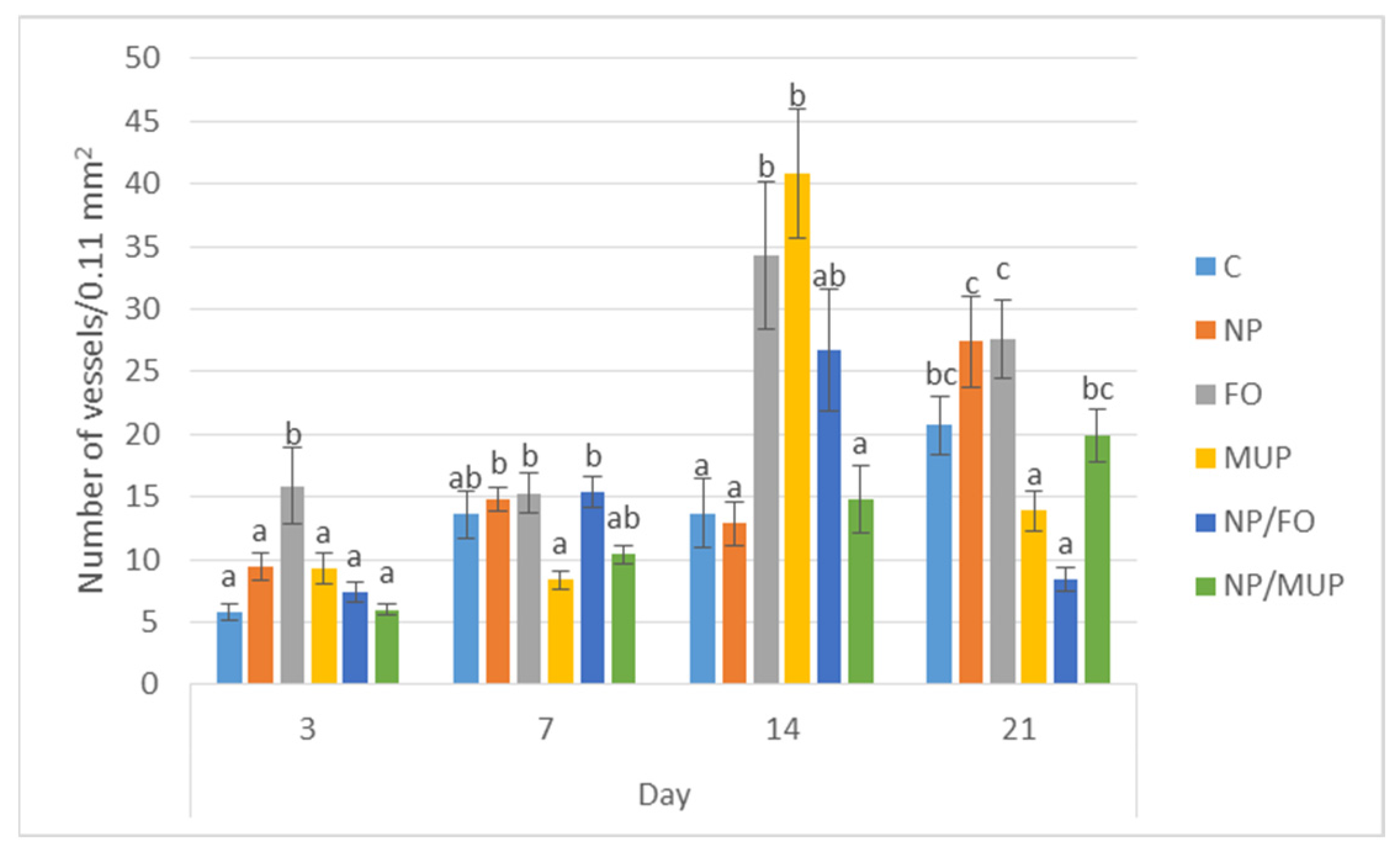
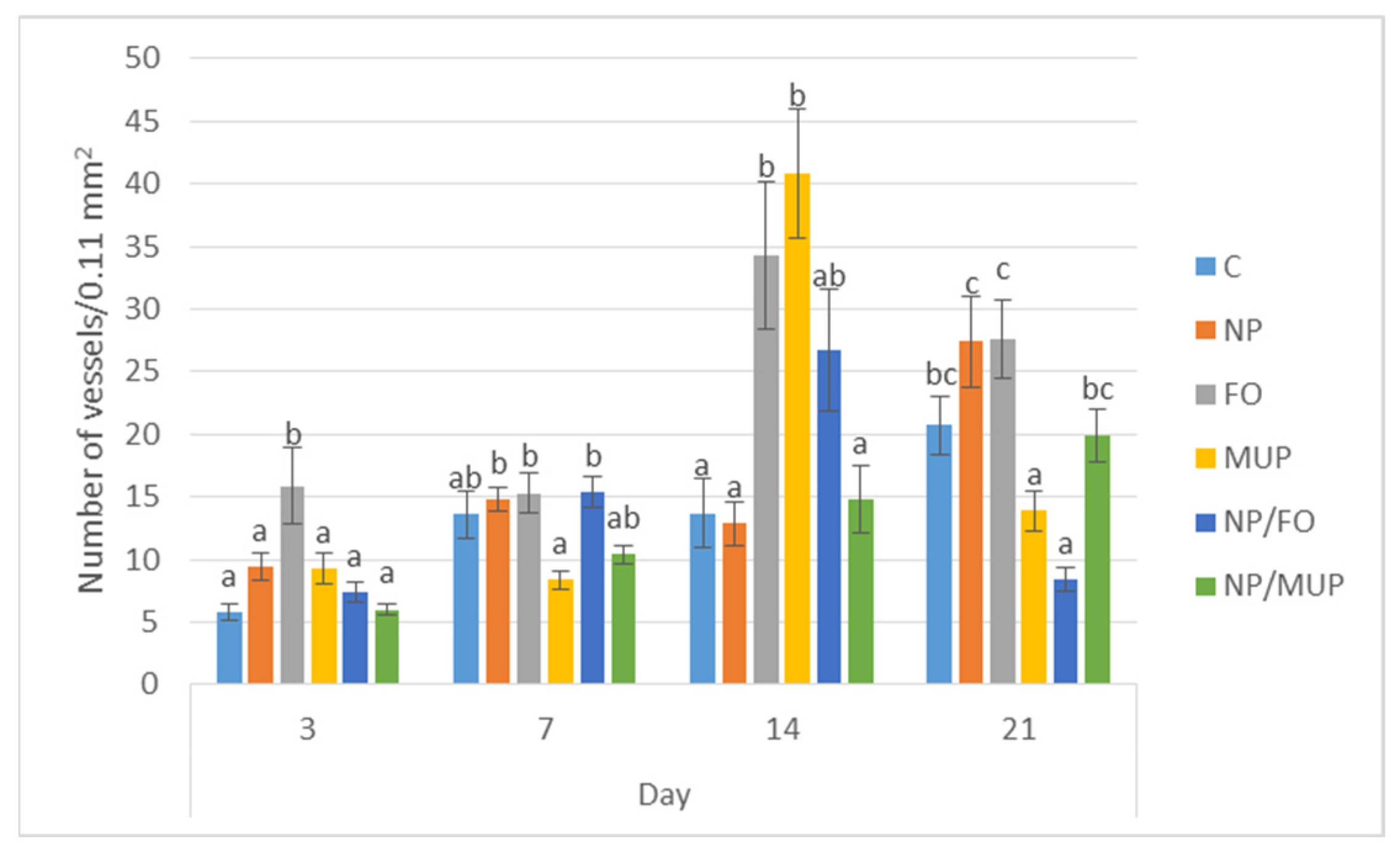
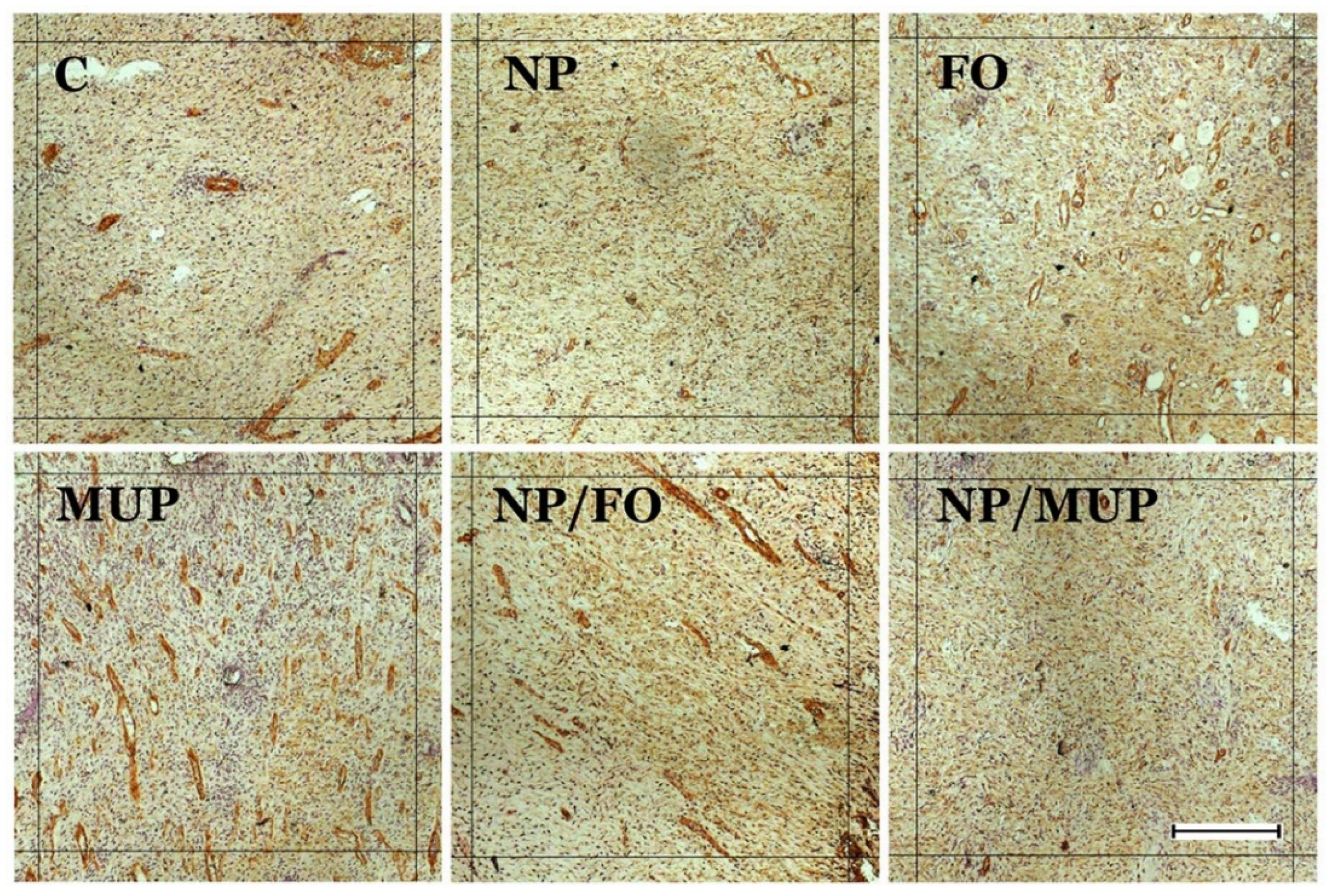
2.2. Angiogenesis
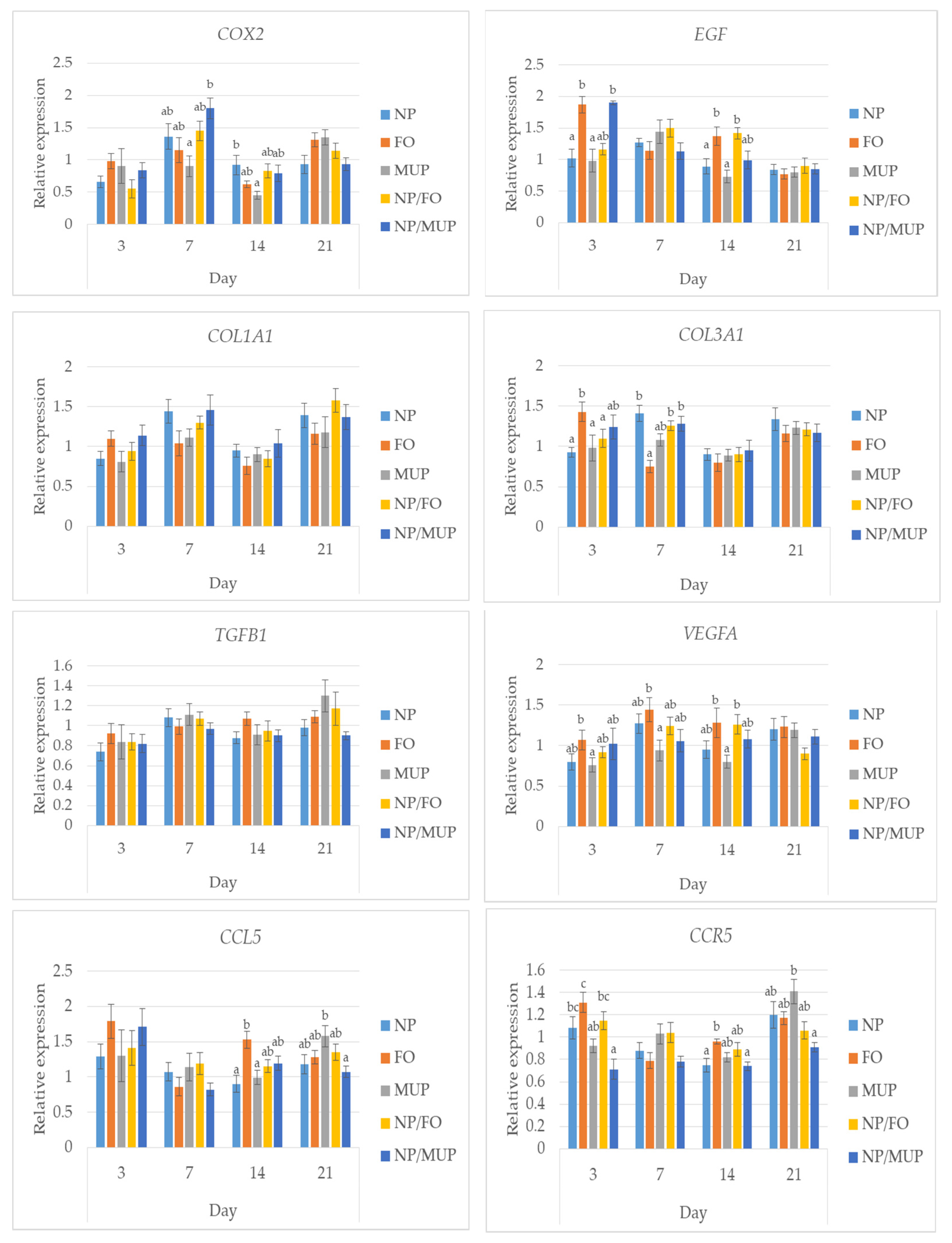
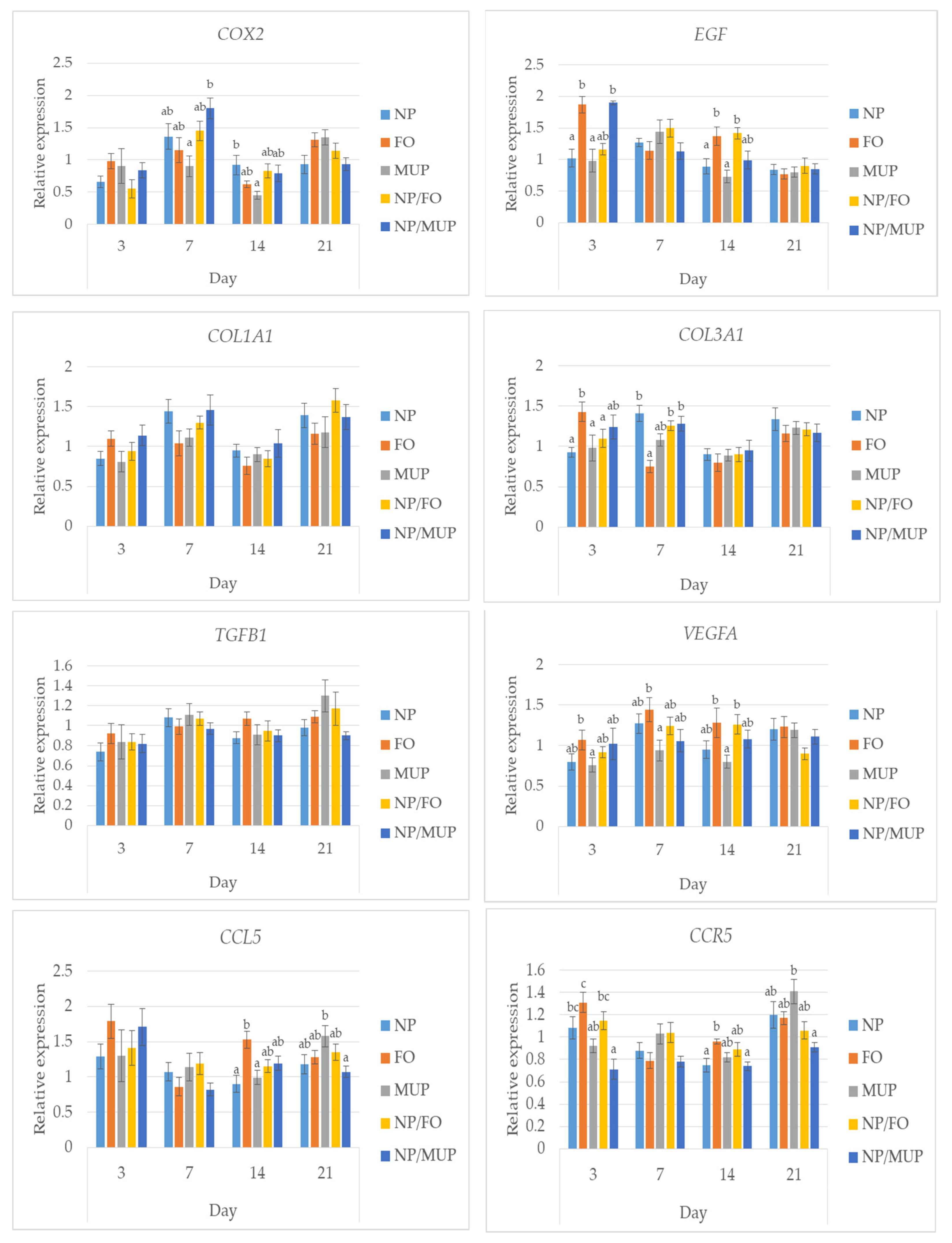
2.3. Gene Expression
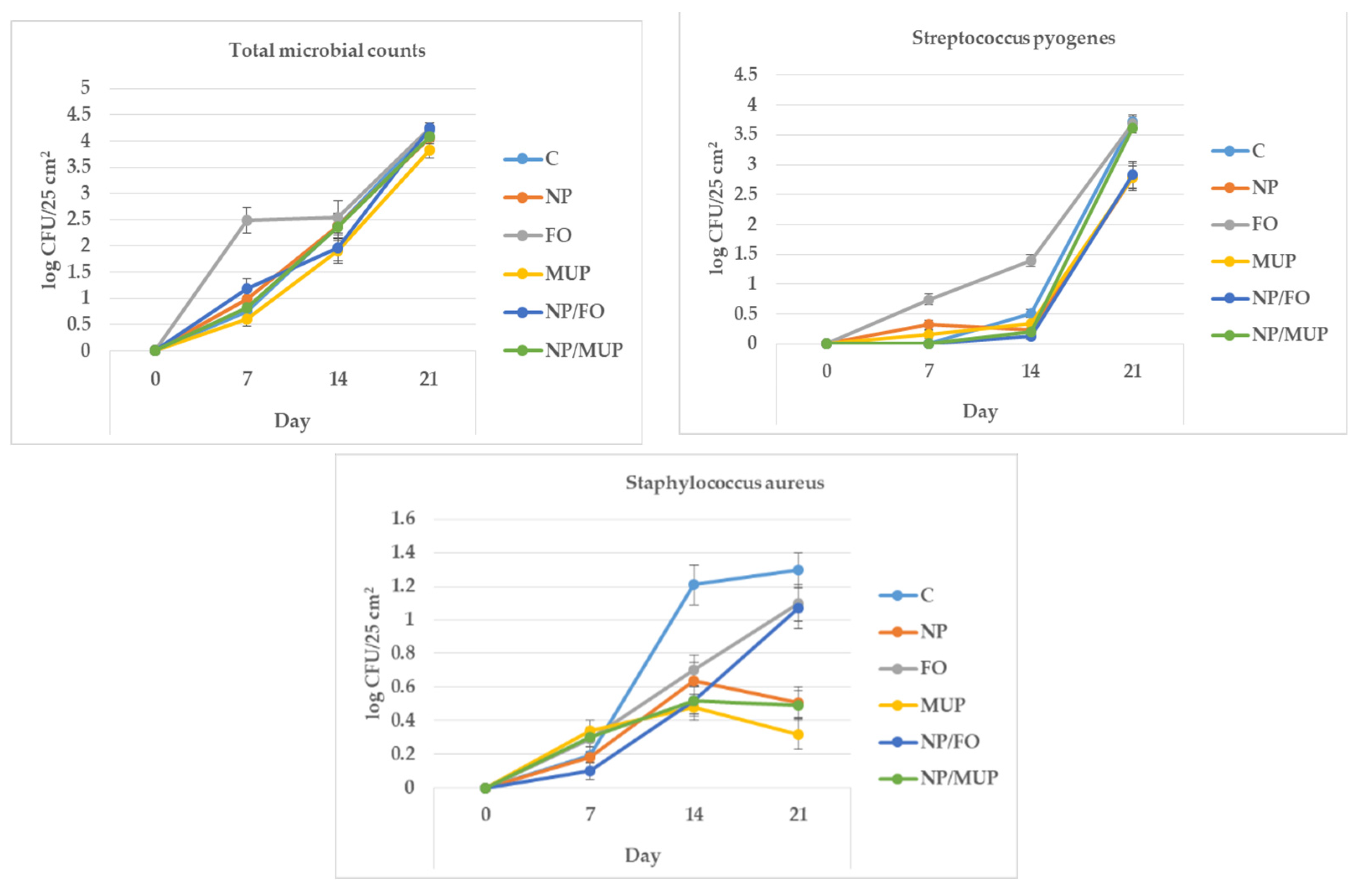
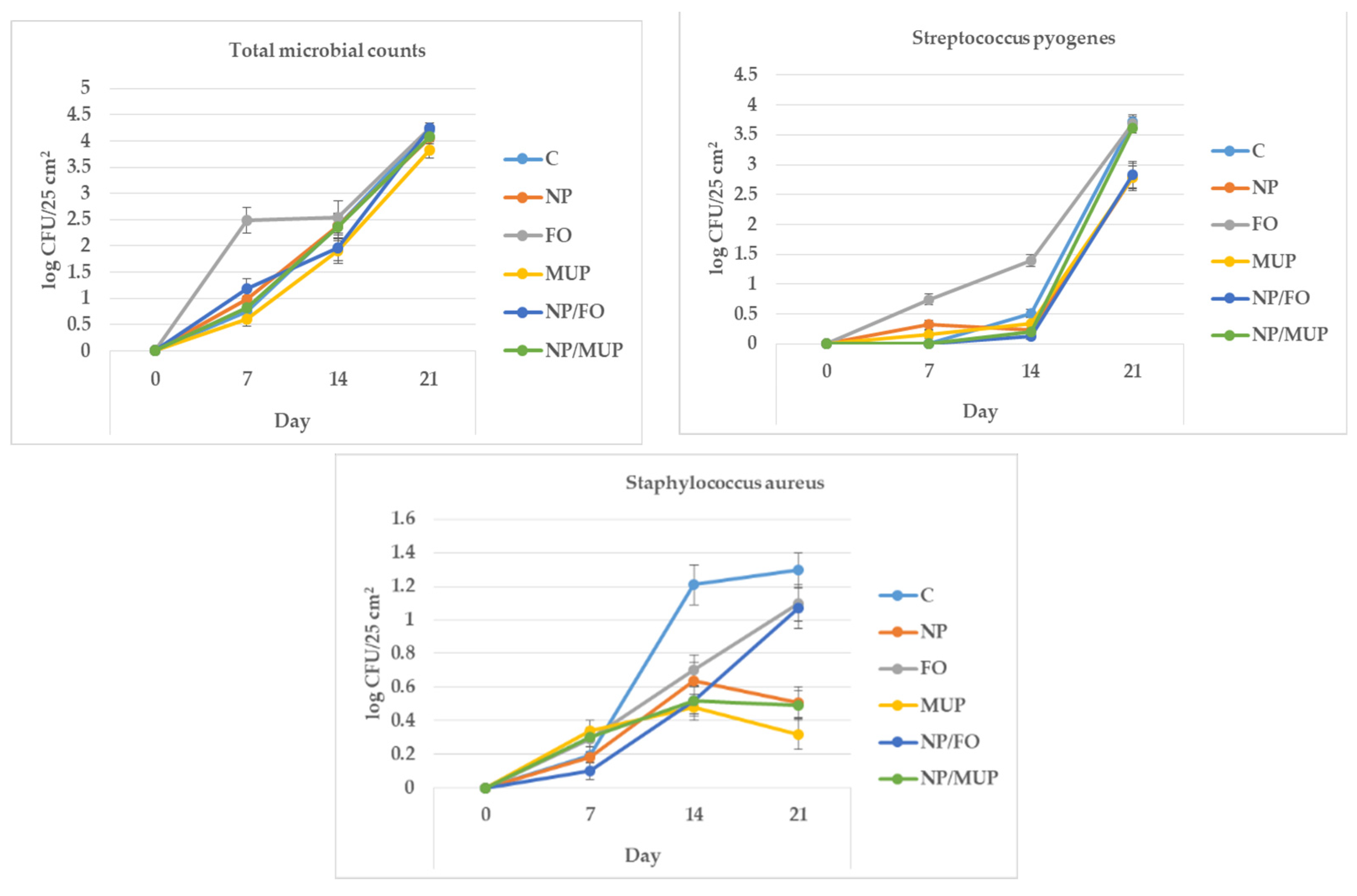
2.4. Microbiological Analysis
3. Discussion
3.1. Collagenous Tissue Maturation
3.2. Angiogenesis
3.3. Gene Expression
3.4. Microbiological Markers
4. Methods and Materials
4.1. Animals
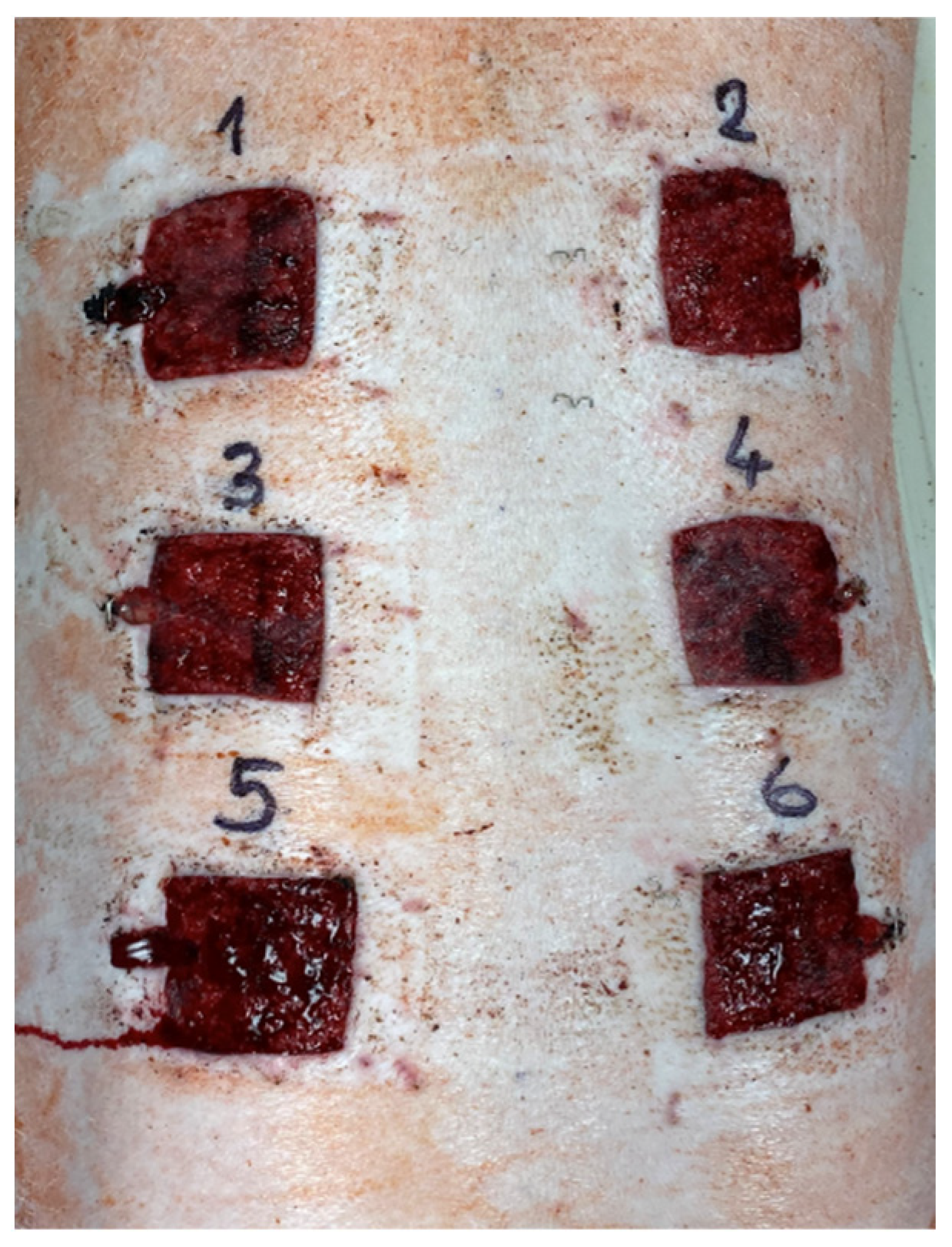
4.2. Experimental Design
4.3. Production of Nanoparticles
4.4. Dressings
4.5. Excisions and Sample Collection
4.6. Analysis of Total Collagenous Tissue Maturation
4.7. Determination of Hydroxyproline
4.8. Evaluation of the Extent of Angiogenesis
4.9. Quantification of the Gene Expression
4.10. Microbiological Analysis
4.11. Statistical Evaluations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Gercek, A.; Yildirim, O.; Konya, D.; Bozkurt, S.; Ozgen, S.; Kilic, T. Effects of parenteral fish-oil emulsion (Omegaven) on cutaneous wound healing in rats treated with dexamethasone. J. Parenter. Enter. Nutr. 2007, 31, 161–166. [Google Scholar] [CrossRef]
- Caetano, G.F.; Fronza, M.; Leite, M.N.; Gomes, A.; Cipriani Frade, M.A. Comparison of collagen content in skin wounds evaluated by biochemical assay and by computer-aided histomorphometric analysis. Pharm. Biol. 2016, 54, 2555–2559. [Google Scholar] [CrossRef]
- Nascimento, A.P.; Costa, A.M. Overweight induced by high-fat diet delays rat cutaneous wound healing. Br. J. Nutr. 2006, 96, 1069–1077. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zhao, Y.; Simonenko, V.; Xu, J.J.; Liu, K.; Wang, D.; Shi, J.; Zhong, T.; Zhang, L.; Zeng, J.; et al. Simultaneous silencing of TGF-ß1 and COX-2 reduces human skin hypertrophic scar through activation of fibroblast apoptosis. Oncotarget 2017, 8, 80651–80665. [Google Scholar] [CrossRef] [Green Version]
- Ramanathan, G.; Muthukumar, T.; Sivagnanan, U.T. In vivo efficiency of the collagen nanofibrous scaffold and their effect on growth factors and pro-inflammatory cytokines in wound healing. Eur. J. Pharmacol. 2017, 814, 44–45. [Google Scholar] [CrossRef]
- Pastor-Clerigues, A.; Marti-Bonmati, E.; Milara, J.; Almudever, P.; Cortijo, J. Anti-inflammatory and anti-fibrotic profile of fish oil emulsions used in parenteral nutrition-associated liver disease. PLoS ONE 2014, 9, e115404. [Google Scholar] [CrossRef]
- Ridiandries, A.; Bursill, C.; Tan, J. Broad-spectrum inhibition of the CC-chemokine class improves wound healing and wound angiogenesis. Int. J. Mol. Sci. 2017, 18, 155. [Google Scholar] [CrossRef] [Green Version]
- Ishida, Y.; Kimura, A.; Kuninaka, Y.; Inui, M.; Matsushima, K.; Mukaida, N.; Kondo, T. Pivotal role of the CCL5/CCR5 interaction for recruitment of endothelial progenitor cells in mouse wound healing. J. Clin. Investig. 2012, 122, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef]
- Vojtová, L.; Pavliňáková, V.; Muchová, J.; Kacvinská, K.; Brtníková, J.; Knoz, M.; Lipový, B.; Faldyna, M.; Göpfert, E.; Holoubek, J.; et al. Healing and Angiogenic Properties of Collagen/Chitosan Scaffolds Enriched with Hyperstable FGF2-STAB® Protein: In Vitro, Ex Ovo and In Vivo Comprehensive Evaluation. Biomedicines 2021, 9, 590. [Google Scholar] [CrossRef]
- Pensalfini, M.; Haertel, E.; Hopf, R.; Wietecha, M.; Werner, S.; Mazza, E. The mechanical fingerprint of murine excisional wounds. Acta Biomater. 2018, 65, 226–236. [Google Scholar] [CrossRef]
- Seaton, M.; Hocking, A.; Gibran, N.S. Porcine models of cutaneous wound healing. ILAR J. 2015, 56, 127–138. [Google Scholar] [CrossRef]
- Chereddy, K.K.; Vandermeulen, G.; Préat, V. PLGA based drug delivery systems: Promising carriers for wound healing activity. Wound Repair Regen. 2016, 24, 223–236. [Google Scholar] [CrossRef]
- Berthet, M.; Gauthier, Y.; Lacroix, C.; Verrier, B.; Monge, C. Nanoparticle-based dressing: The future of wound treatment? Trends Biotechnol. 2017, 35, 770–784. [Google Scholar] [CrossRef]
- Kemme, M.; Heinzel-Wieland, R. Quantitative assessment of antimicrobial activity of PLGA films loaded with 4-hexylresorcinol. J. Funct. Biomater. 2018, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- Sritharadol, R.; Nakpheng, T.; Heng, P.W.S.; Srichana, T. Development of a topical mupirocin spray for antibacterial and wound-healing applications. Drug Dev. Ind. Pharm. 2017, 43, 1715–1728. [Google Scholar] [CrossRef]
- Budhiraja, M.; Zafar, S.; Akhter, S.; Alrobaian, M.; Rashid, M.A.; Barkat, M.A.; Beg, S.; Ahmad, F.J. Mupirocin-loaded chitosan microspheres embedded in Piper betle extract containing collagen scaffold accelerate wound healing activity. AAPS Pharm. Sci. Tech. 2022, 23, 77. [Google Scholar] [CrossRef]
- Solak, E.K.; Kaya, S.; Asman, G. Preparation, characterization, and antibacterial properties of biocompatible material for wound healing. J. Macromol. Sci. A Pure Appl. Chem. 2021, 58, 709–716. [Google Scholar] [CrossRef]
- Giustina, A.D.; de Souza Goldim, M.P.; Gainski Danielski, L.; Garbossa, L.; Oliveira Junior, A.N.; Cidreira, T.; Denico, T.; Bonfante, S.; da Rosa, N.; Fortunato, J.J.; et al. Lipoic acid and fish oil combination potentiates neuroinflammation and oxidative stress regulation and prevents cognitive decline of rats after sepsis. Mol. Neurobiol. 2020, 57, 4451–4466. [Google Scholar] [CrossRef]
- Rakotoarisoa, M.; Angelov, B.; Garamus, V.M.; Angelova, A. Curcumin- and fish oil-loaded spongosome and cubosome nanoparticles with neuroprotective potential against H2O2-induced oxidative stress in differentiated human SH-SY5Y cells. ACS Omega 2019, 4, 3061–3073. [Google Scholar] [CrossRef] [Green Version]
- Komprda, T. Effect of n-3 long-chain polyunsaturated fatty acids on wound healing using animal models—A review. Acta Vet. Brno 2018, 87, 309–320. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, G.; Ecker, J. The opposing effects of n-3 and n-6 fatty acids. Prog. Lipid Res. 2008, 47, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Komprda, T. Eicosapentaenoic and docosahexaenoic acids as inflammation-modulating and lipid homeostasis influencing nutraceuticals: A review. J. Funct. Foods 2012, 4, 25–38. [Google Scholar] [CrossRef]
- Chereddy, K.K.; Coco, R.; Memvanga, P.B.; Ucakar, B.; Rieux, A.; Vandermeulen, G.; Préat, V. Combined effect of PLGA and curcumin on wound healing activity. J. Control. Release 2013, 171, 208–215. [Google Scholar] [CrossRef]
- Liu, S.-J.; Kau, Y.-C.; Chou, C.-Y.; Chen, J.-K.; Wu, R.-C.; Yeh, W.-L. Electrospun PLGA/collagen nanofibrous membrane as early-stage wound dressing. J. Membr. Sci. 2010, 355, 53–59. [Google Scholar] [CrossRef]
- Candreva, T.; Kühl, C.M.C.; Burger, B.; dos Anjos, M.B.P.; Torsoni, M.A.; Consonni, S.R.; Crisma, A.R.; Fisk, H.L.; Calder, P.C.; de Mato, F.C.P.; et al. Docosahexaenoic acid slows inflammation resolution and impairs the quality of healed skin tissue. Clin. Sci. 2019, 133, 2345–2360. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, C.R.; Favoreto, S.; Oliveira, L.L.; Vancim, J.O.; Barban, G.B.; Ferraz, D.B.; Silva, J.S. Oleic acid modulation of the immune response in wound healing: A new approach for skin repair. Immunobiology 2011, 216, 409–415. [Google Scholar] [CrossRef]
- Bradberry, J.C.; Hilleman, D.E. Overview of omega-3 fatty acid therapies. Pharm. Ther. 2013, 38, 681–691. [Google Scholar]
- Turk, H.F.; Monk, J.M.; Fan, Y.Y.; Callaway, E.S.; Weeks, B.; Chapkin, R.S. Inhibitory effects of omega-3 fatty acids on injury-induced epidermal growth factor receptor transactivation contribute to delayed wound healing. Am. J. Physiol. Cell Physiol. 2013, 304, C905–C917. [Google Scholar] [CrossRef] [Green Version]
- McDaniel, J.C.; Belury, M.; Ahijevych, K.; Blakely, W. Omega-3 fatty acids effects on wound healing. Wound Repair Regen. 2008, 16, 337–345. [Google Scholar] [CrossRef] [Green Version]
- Arantes, E.L.; Dragano, N.; Ramalho, A.; Vitorino, D.; de Souza, G.F.; Lima, M.H.M.; Velloso, L.A.; Araúcho, E.P. Topical docosahexaenoic acid accelerates skin wound healing in rats and activates GPR120. Biol. Res. Nurs. 2016, 18, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Kadler, K.E.; Holmes, D.F.; Trotter, J.A.; Chapman, J.A. Collagen fibril formation. Biochem. J. 1996, 316, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wolfram, D.; Tzankov, A.; Pülzl, P.; Piza-Katzer, H. Hypertrophic scars and keloids a review of their pathophysiology, risk factors, and therapeutic management. Dermatol. Surg. 2009, 35, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, T.; Hashimoto, Y.; Myokei, Y.; Aoyama, H.; Izawa, Y. Changes in type of collagen during the development of human post-burn hypertrophic scars. Clin. Chim. Acta 1979, 93, 119–125. [Google Scholar] [CrossRef]
- Wang, J.F.; Olson, M.E.; Reno, C.R.; Kulyk, W.; Wright, J.B.; Hart, D.A. Molecular and cell biology of skin wound healing in a pig model. Connect. Tissue Res. 2000, 41, 195–211. [Google Scholar] [CrossRef]
- Kawai, K.; Larson, B.J.; Ishise, H.; Carre, A.L.; Nishimoto, S.; Longaker, M.; Lorenz, H.P. Calcium-Based Nanoparticles Accelerate Skin Wound Healing. PLoS ONE 2021, 6, e27106. [Google Scholar] [CrossRef] [Green Version]
- Kwan, K.H.L.; Liu, X.; To, M.K.T.; Yeung, K.W.K.; Ho, C.H.; Wong, K.K.Y. Modulation of collagen alignment by silver nanoparticles results in better mechanical properties in wound healing. Nanomedicine 2011, 7, 497–504. [Google Scholar] [CrossRef]
- Tanideh, N.; Abdordide, E.; Yousefabad, S.L.A.; Daneshi, S.; Hosseinabadi, O.K.; Samani, S.M. A comparison of the effects of honey, fish oil and their combination on wound healing in rat. J. Coast. Life Med. 2016, 4, 683–688. [Google Scholar]
- Rittié, L. Cellular mechanisms of skin repair in humans and other mammals. J. Cell Commun. Signal. 2016, 10, 103–120. [Google Scholar] [CrossRef] [Green Version]
- Turgeon, J.; Dussault, S.; Maingrette, F.; Groleau, J.; Haddad, P.; Perez, G.; Rivard, A. Fish oil-enriched diet protects against ischemia by improving angiogenesis, endothelial progenitor cell function and postnatal neovascularization. Atherosclerosis 2013, 229, 295–303. [Google Scholar] [CrossRef]
- Ali, M.; Radad, K. Cod liver oil/honey mixture: An effective treatment of equine complicated lower leg wounds. Vet. World 2011, 4, 304–310. [Google Scholar]
- Shingel, K.I.; Faure, M.P.; Azoulay, L.; Roberge, C.; Deckelbaum, R.J. Solid emulsion gel as a vehicle for delivery ofpolyunsaturated fatty acids: Implications for tissuerepair, dermal angiogenesis and wound healing. J. Tissue Eng. Regen. Med. 2008, 2, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Okur, N.Ü.; Hökenek, N.; Okur, M.H.; Ayla, S.; Yoltaş, A.; Siafaka, P.I.; Cevherg, E. An alternative approach to wound healing field; new composite films from natural polymers for mupirocin dermal delivery. Saudi Pharm. J. 2019, 27, 738–752. [Google Scholar] [CrossRef] [PubMed]
- Farahpoura, M.R.; Pirkhezrb, E.; Ashrafianc, A.; Sonboli, A. Accelerated healing by topical administration of Salvia officinalis essentialoil on Pseudomonas aeruginosa and Staphylococcus aureus infected wound model. Biomed. Pharmacother. 2020, 128, 110–120. [Google Scholar]
- Khezri, K.; Farahpour, M.R.; Rad, S.M. Accelerated infected wound healing by topical application of encapsulated Rosemary essential oil into nanostructured lipid carriers. Artif. Cells Nanomed. Biotechnol. 2019, 47, 980–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golmohammadi, R.; Najar-Peerayeh, S.; Moghadam, T.T.; Hosseini, S.M.J. Synergistic antibacterial activity and wound healing properties of selenium-chitosan-mupirocin nanohybrid System: An in vivo study on rat diabetic Staphylococcus aureus wound infection model. Sci. Rep. 2020, 10, 2854. [Google Scholar] [CrossRef] [Green Version]
- Porporato, P.; Payen, V.; Saedeleer, C.; Préat, V.; -Thissen, P.J.; Feron, O. Lactate stimulates angiogenesis and accelerates the healing of superficial and ischemic wounds in mice. Angiogenesis 2012, 15, 581–592. [Google Scholar] [CrossRef]
- Barui, A.K.; Nethi, S.K.; Haque, S.; Basuthakur, P.; Patra, C.H.R. Recent Development of metal nanoparticles for angiogenesis study and their therapeutic applications. ACS Appl. Bio Mater. 2019, 2, 5492–5511. [Google Scholar] [CrossRef]
- Kargozar, S.; Baino, F.; Hamzehlou, S.; Hamblin, M.R.; Mozafari, M. Nanotechnology for angiogenesis: Opportunities and challenges. Chem. Soc. Rev. 2020, 49, 5008–5057. [Google Scholar] [CrossRef]
- Bluff, J.E.; O'Ceallaigh, S.; O'Kane, S.; Ferguson, M.W.; Ireland, G. The microcirculation in acute murine cutaneous incisional wounds shows a spatial and temporal variation in the functionality of vessels. Wound Repair Regen. 2006, 14, 434–442. [Google Scholar] [CrossRef]
- Pan, S.-C.; Li, C.-Y.; Kuo, C.-Y.; Kuo, Y.-Z.; Fang, W.-Y.; Huang, Y.-H.; Hsieh, T.-C.; Kao, H.-Y.; Kuo, Y.; Kang, Y.-R.; et al. The p53-S100A2 positive feedback loop negatively regulates epithelialization in cutaneous wound healing. Sci. Rep. 2018, 8, 5458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardwicke, J.; Schmaljohann, D.; Boyce, D.; Thomas, D. Epidermal growth factor therapy and wound healing–past, present and future perspectives. Surgeon 2008, 6, 172–177. [Google Scholar] [CrossRef]
- Cheng, W.; Xu, R.; Li, D.; Bortolini, C.; He, J.; Dong, M.; Besenbacher, F.; Huang, Y.; Chen, M. Artificial extracellular matrix delivers TGFb1 regulating myofibroblast differentiation. RSC Adv. 2016, 6, 21922–21928. [Google Scholar] [CrossRef]
- Khalaf, A.A.; Hassanen, E.I.; Zaki, A.R.; Tohamy, A.F.; Ibrahim, M.A. Histopathological, immunohistochemical, and molecular studies for determination of wound age and vitality in rats. Int. Wound J. 2019, 19, 1416–1425. [Google Scholar] [CrossRef] [PubMed]
- Behm, B.; Babilas, P.; Landthaler, M.; Schreml, C. Cytokines, chemokines and growth factors in wound healing. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 812–820. [Google Scholar] [CrossRef]
- Rezaii, M.; Oryan, S.; Javeri, A. Curcumin nanoparticles incorporated collagen-chitosan csaffold promotes cutaneous wound healing through regulation of TGF-ß1/Smad7 gene expression. Mater. Sci. Eng. C 2019, 98, 347–357. [Google Scholar] [CrossRef]
- Werner, S.; Grose, R. Regulation of wound healing by growth factors and cytokines. Physiol. Rev. 2003, 83, 835–870. [Google Scholar] [CrossRef]
- Chanda, W.; Joseph, T.P.; Guo, X.F.; Wang, W.D.; Liu, M.; Vuai, M.S.; Zhong, M.T. Effectiveness of omega-3 polyunsaturated fatty acids against microbial pathogens. J. Zhejiang Univ.-SCIENCE B 2018, 19, 253–262. [Google Scholar] [CrossRef]
- Casillas-Vargas, G.; Ocasio-Malavé, C.; Medina, S.; Morales-Guzmán, C.; Del Valle, R.G.; Carballeira, N.M.; Sanabria-Ríos, D.J. Antibacterial fatty acids: An update of possible mechanisms of action and implications in the development of the next-generation of antibacterial agents. Prog. Lipid Res. 2021, 82, 101093. [Google Scholar] [CrossRef]
- Azzazy, H.M.E.-S.; Fahmy, S.A.; Mahdi, N.K.; Meselhy, M.R.; Bakowsky, U. Chitosan-coated PLGA nanoparticles loaded with Peganum harmala alkaloids with promising antibacterial and wound healing activities. Nanomaterials 2021, 11, 2438. [Google Scholar] [CrossRef]
- Hasan, N.; Cao, J.; Lee, J.; Hlaing, S.P.; Oshi, M.A.; Naeem, M.; Ki, M.-H.; Lee, B.L.; Jung, Y.; Yoo, J.-W. Bacteria-targeted clindamycin loaded polymeric nanoparticles: Effect of surface charge on nanoparticle adhesion to MRS, antibacterial activity, and wound healing. Pharmaceutics 2019, 11, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komprda, T.; Popelková, V.; Košarišťanová, L.; Šmídová, V. Poly(lactic-co-glycolic) Acid Nanoparticles as Delivery System of Fish Oil for Wound Healing. Acta Vet. Brno 2022, 91. in print. [Google Scholar]
- Popelková, V.; Košarišťanová, L.; Komprda, T.; Vymazalová, P.; Fialová, T.; Švec, P.; Šmídová, V.; Bytešníková, Z.; Astete, C.E.; Sabliov, C.; et al. Poly(lactic-co-glycolic) Acid Nanoparticles as Feasible Delivery System of Antimicrobial Mupirocin for Possible Wound Healing. RSC Adv. 2022. submitted. [Google Scholar]
- Dorazilová, J.; Muchová, J.; Šmerková, K.; Kočiová, S.; Diviš, P.; Kopel, P.; Veselý, R.; Pavliňáková, V.; Adam, V.; Vojtová, L. Synergistic effect of chitosan and selenium nanoparticles on biodegradation and antibacterial properties of collagenous scaffolds designed for infected burn wounds. Nanomaterials 2020, 10, 1971. [Google Scholar] [CrossRef] [PubMed]
- Suresh, K.P. An overview of randomization techniques: An unbiased assessment of outcome in clinical research. J. Hum. Reprod. Sci. 2011, 4, 8–11. [Google Scholar] [CrossRef]
- Maia-Figueiró, T.L.; Odashiro, A.N.; De Menezes, G.P.; Coelho, L.R.; Breda, I.; De Souza, B.A.; Figueiró-Filho, E.A. Semi-Quantitative Histological Analysis of the Effect of Intense Pulsed Light (IPL) and Carbon Dioxide (CO2) Intradermic injection on fibroblast and collagen proliferation in the skin of Wistar rats. J. Cosmet. Dermatol. Sci. Appl. 2012, 2, 164–173. [Google Scholar]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- von der Hardt, K.; Kandler, M.A.; Fink, L.; Schoof, E.; Dötsch, J.; Brandenstein, O.; Bohle, R.M.; Rascher, W. High frequency oscillatory ventilation suppresses inflammatory response in lung tissue and microdissected alveolar macrophages in surfactant depleted piglets. Pediatr. Res. 2004, 55, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Ondrackova, P.; Leva, L.; Kucerova, Z.; Vicenova, M.; Mensikova, M.; Faldyna, M. Distribution of porcine monocytes in different lymphoid tissues and the lungs during experimental Actinobacillus pleuropneumoniae infection and the role of chemokines. Vet. Res. 2013, 44, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nygard, A.B.; Jørgensen, C.B.; Cirera, S.; Fredholm, M. Selection of reference genes for gene expression studies in pig tissues using SYBR green qPCR. BMC Mol. Biol. 2007, 8, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brakstad, O.G.; Aasbakk, K.; Maeland, J.A. Detection of Staphylococcus aureus by polymerase chain reaction amplification of the nuc gene. J. Clin. Microbiol. 1992, 30, 1654–1660. [Google Scholar] [CrossRef] [Green Version]
- Candrian, U.; Furrer, B.; Höfelein, C.; Meyer, R.; Jermini, M.; Lüthy, J. Detection of Escherichia coli and identification of enterotoxigenic strains by primer-directed enzymatic amplification of specific DNA sequences. Int. J. Food Microbiol. 1991, 12, 339–351. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Days after Excision | |||
|---|---|---|---|---|
| 3 | 7 | 14 | 21 | |
| NP 1 | 0 | – | – | – |
| FO 2 | 0 | – | – | 0 |
| MUP 3 | 0 | – | 0 | 0 |
| NP/FO 4 | 0 | 0 | – | 0 |
| NP/MUP 5 | 0 | – | – | 0 |
| Gene | 5′-Forward Primer-3′ 5′-Reverse Primer-3′ | Primer Origin Amplicon Length 1/Eff 2 |
|---|---|---|
| COX2 | CTTAAACAGGAGCACCCGGAATCACAATCTTAATCGTTTCTCCTATCAG | designed in this study 87/2.053 |
| EGF |
AGCTATGCCTGCAACTGTGTTT ACCATTTCAAGTCTCTGTGCTGAC | designed in this study 67/1.945 |
| COL1A1 | ACGCCATCAAAGTCTTCTGCAAC TTGGGGTTCTTGCTGATGTACCA | designed in this study 103/2.051 |
| COL3A1 | GACGAGATGGAAACCCTGGATCAAGGAGAGCCATTTTCACCACGAT | designed in this study 89/2.040 |
| TGFb1 | TACGCCAAGGAGGTCACCC CAGCTCTGCCCGAGAGAGC | von der Hardt et al. [70] 156/2.007 |
| VEGFA | TAGAGCGAGGCAAGAAAATCCCT CAGGAACATTTACACGTCTGCGG | designed in this study 90/2.089 |
| CCL5 (RANTES) |
ACCACACCCTGCTGTTTTTC GGCGGTTCTTTCTGGTGATA | Ondrackova et al. [71] 124/2.014 |
| CCR5 |
TGGTCAGAGGAGCTGAGACA AGAAGGGACTCGTCGTTTGA | Ondrackova et al. [70] 86/2.084 |
| TBP1 |
AACAGTTCAGTAGTTATGAGCCAGA AGATGTTCTCAAACGCTTCG | Nygard et al. [72] 153/1.938 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komprda, T.; Sládek, Z.; Vícenová, M.; Simonová, J.; Franke, G.; Lipový, B.; Matejovičová, M.; Kacvinská, K.; Sabliov, C.; Astete, C.E.; et al. Effect of Polymeric Nanoparticles with Entrapped Fish Oil or Mupirocin on Skin Wound Healing Using a Porcine Model. Int. J. Mol. Sci. 2022, 23, 7663. https://doi.org/10.3390/ijms23147663
Komprda T, Sládek Z, Vícenová M, Simonová J, Franke G, Lipový B, Matejovičová M, Kacvinská K, Sabliov C, Astete CE, et al. Effect of Polymeric Nanoparticles with Entrapped Fish Oil or Mupirocin on Skin Wound Healing Using a Porcine Model. International Journal of Molecular Sciences. 2022; 23(14):7663. https://doi.org/10.3390/ijms23147663
Chicago/Turabian StyleKomprda, Tomáš, Zbyšek Sládek, Monika Vícenová, Jana Simonová, Gabriela Franke, Břetislav Lipový, Milena Matejovičová, Katarína Kacvinská, Cristina Sabliov, Carlos E. Astete, and et al. 2022. "Effect of Polymeric Nanoparticles with Entrapped Fish Oil or Mupirocin on Skin Wound Healing Using a Porcine Model" International Journal of Molecular Sciences 23, no. 14: 7663. https://doi.org/10.3390/ijms23147663
APA StyleKomprda, T., Sládek, Z., Vícenová, M., Simonová, J., Franke, G., Lipový, B., Matejovičová, M., Kacvinská, K., Sabliov, C., Astete, C. E., Levá, L., Popelková, V., Bátik, A., & Vojtová, L. (2022). Effect of Polymeric Nanoparticles with Entrapped Fish Oil or Mupirocin on Skin Wound Healing Using a Porcine Model. International Journal of Molecular Sciences, 23(14), 7663. https://doi.org/10.3390/ijms23147663