
Identification of 8-Hydroxyquinoline Derivatives That Decrease Cystathionine Beta Synthase (CBS) Activity

 , ,
, ,  ,
,  , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
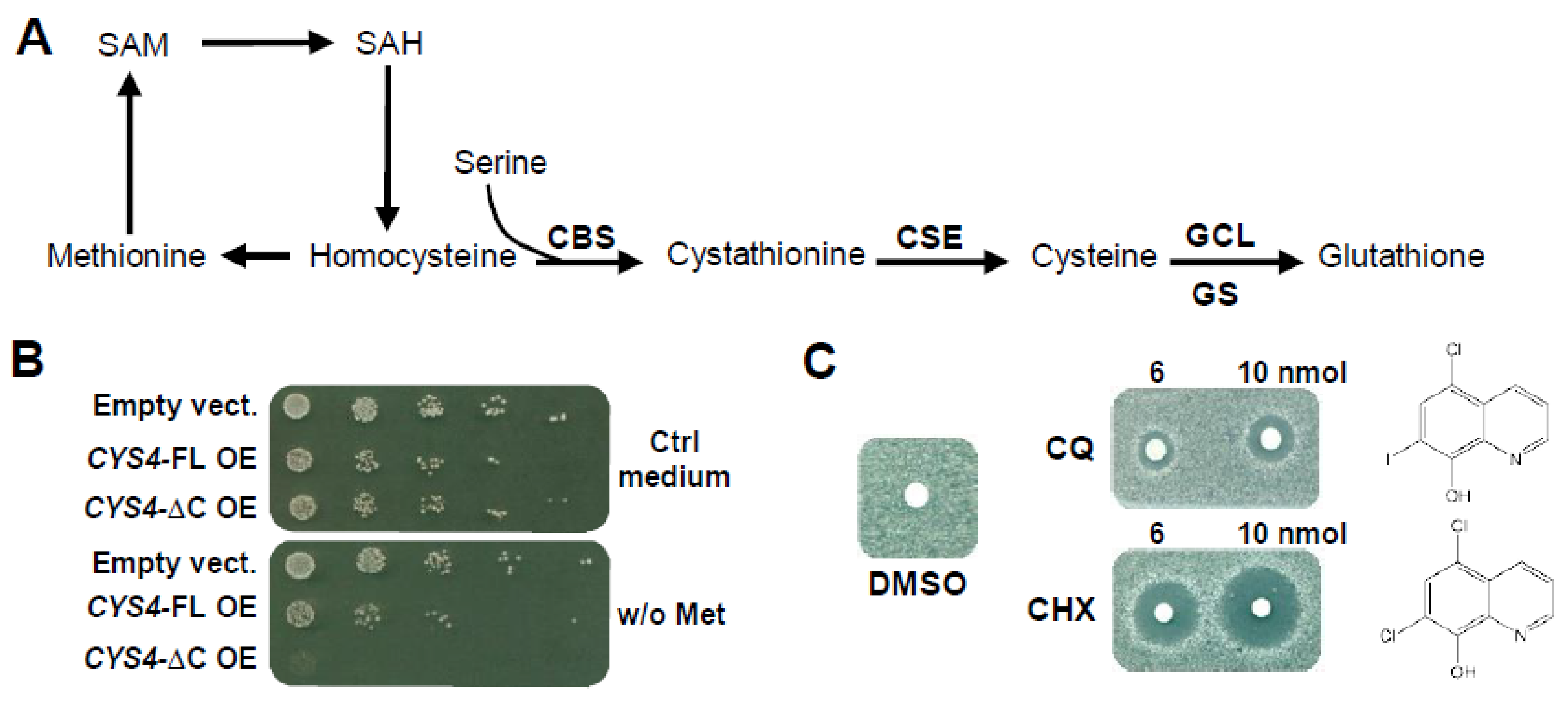
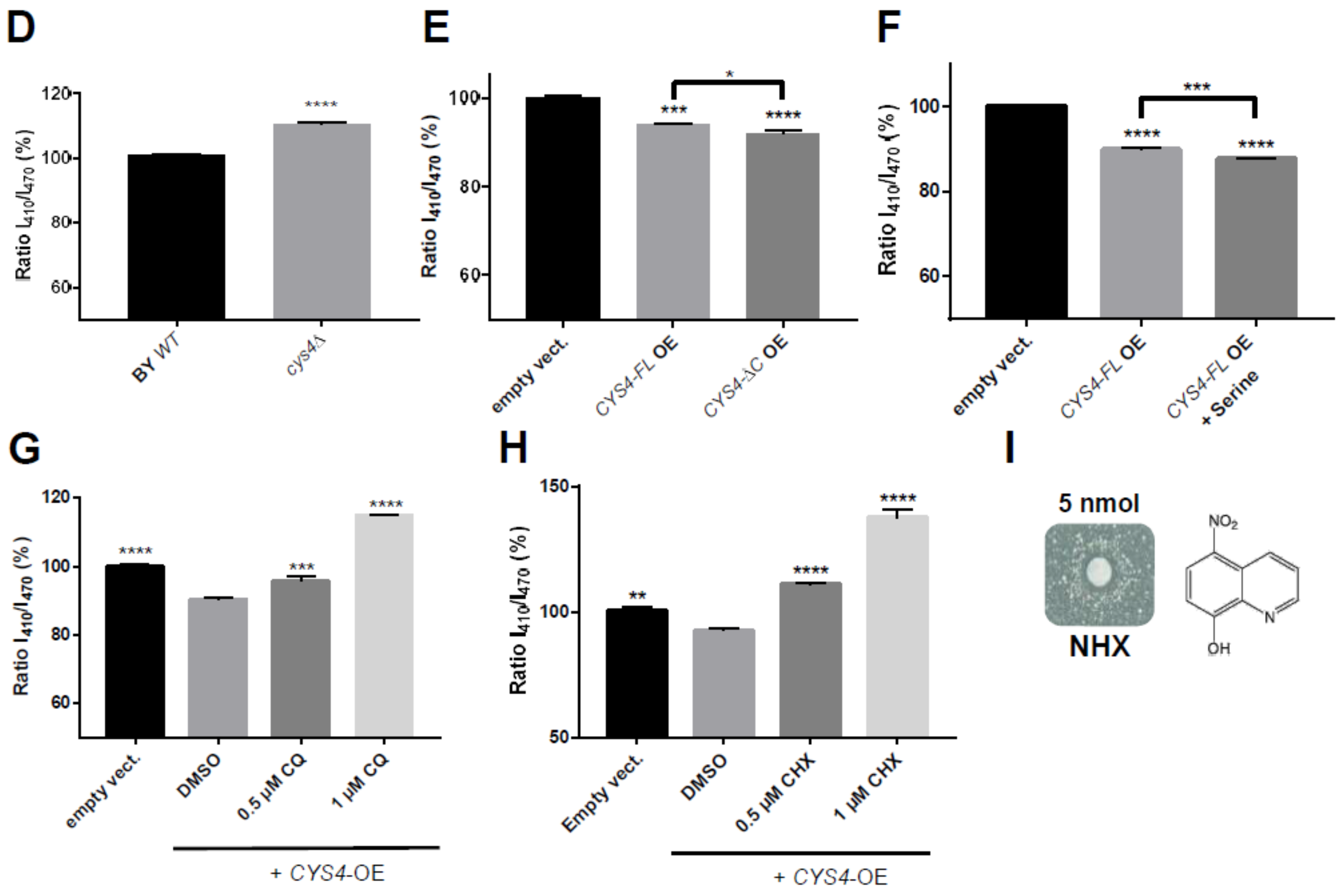
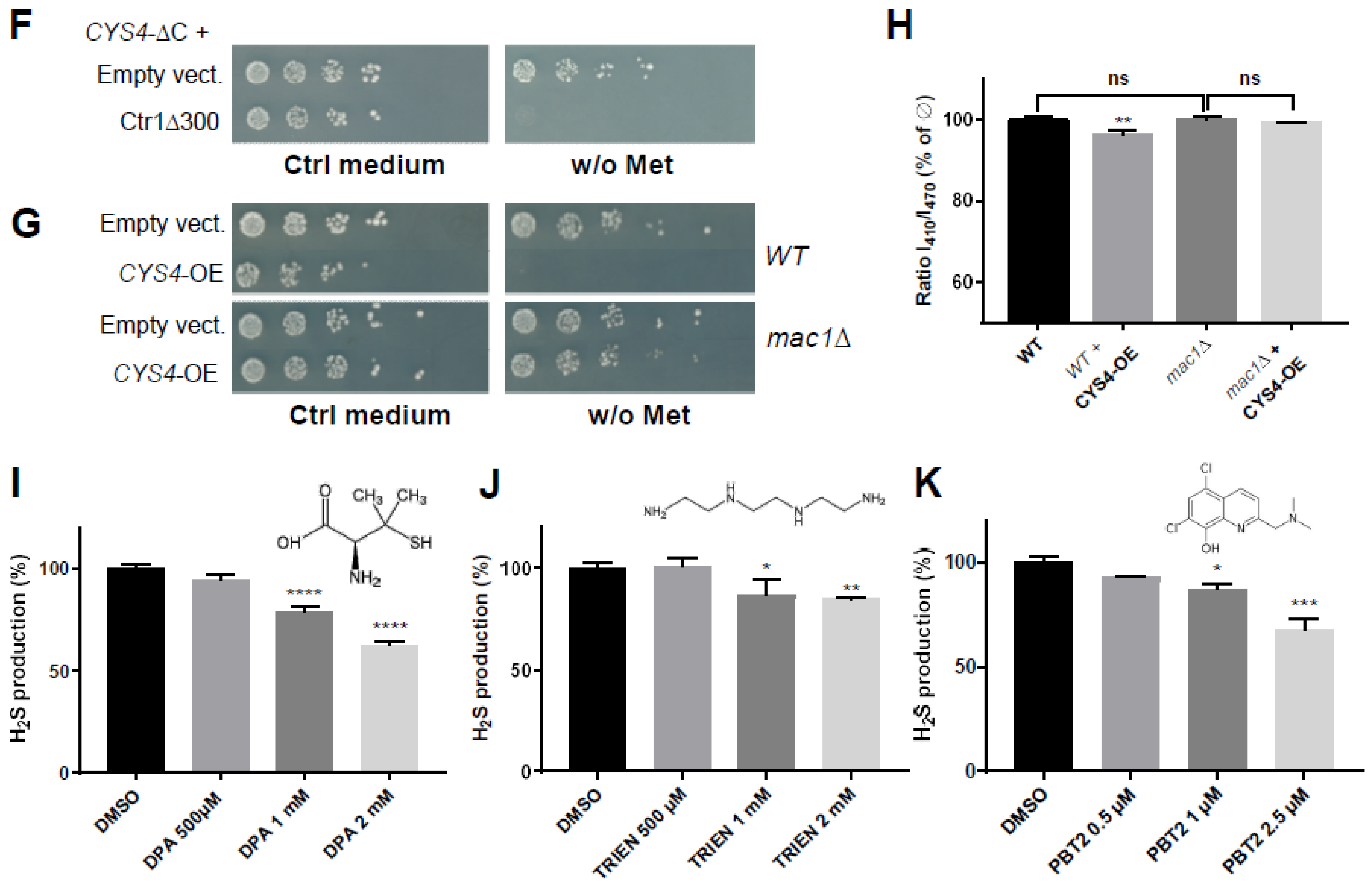
2.1. 8-Hydroxyquinoline Derivatives Suppress the Phenotypes Caused by CYS4 Overexpression
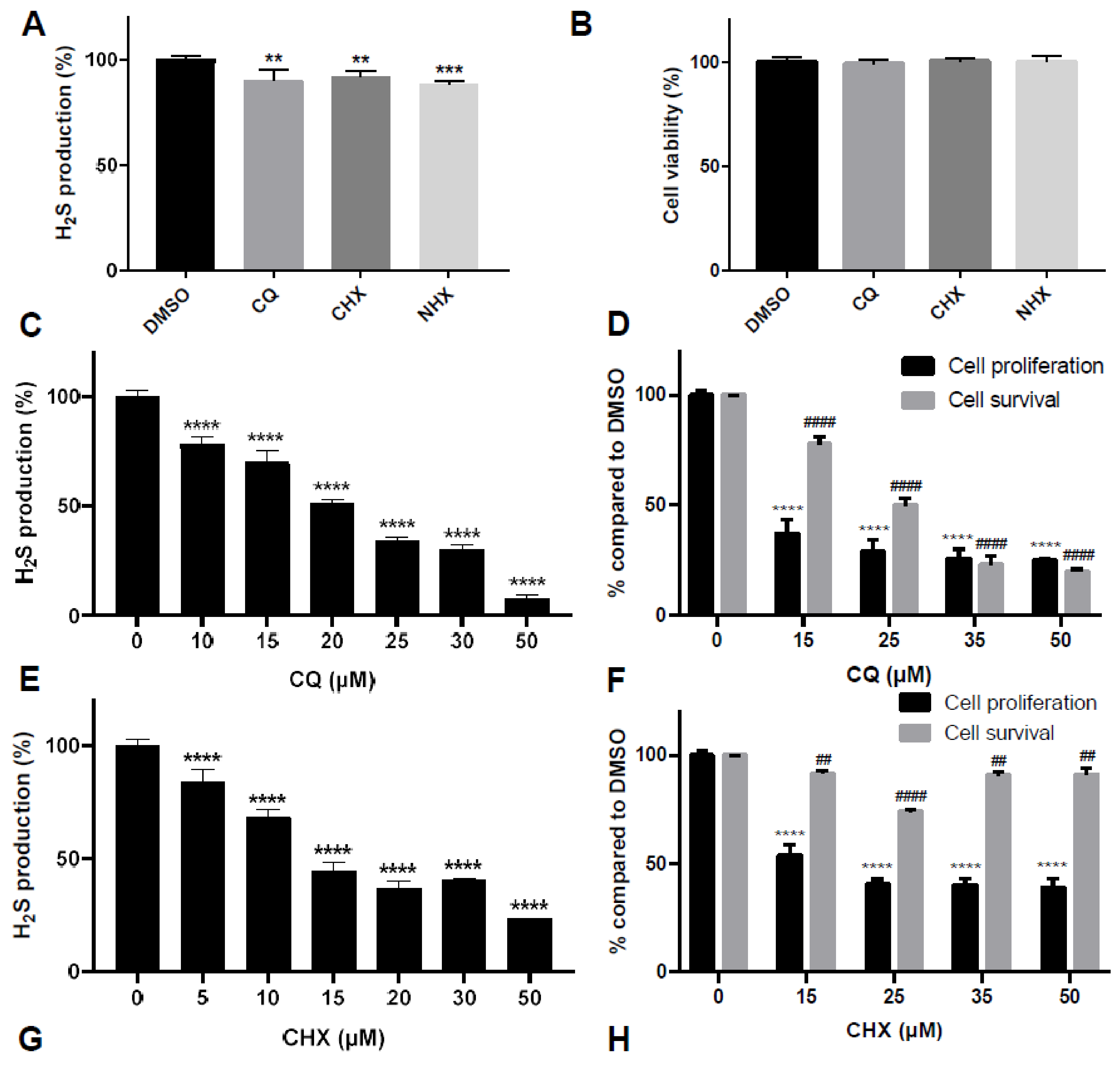
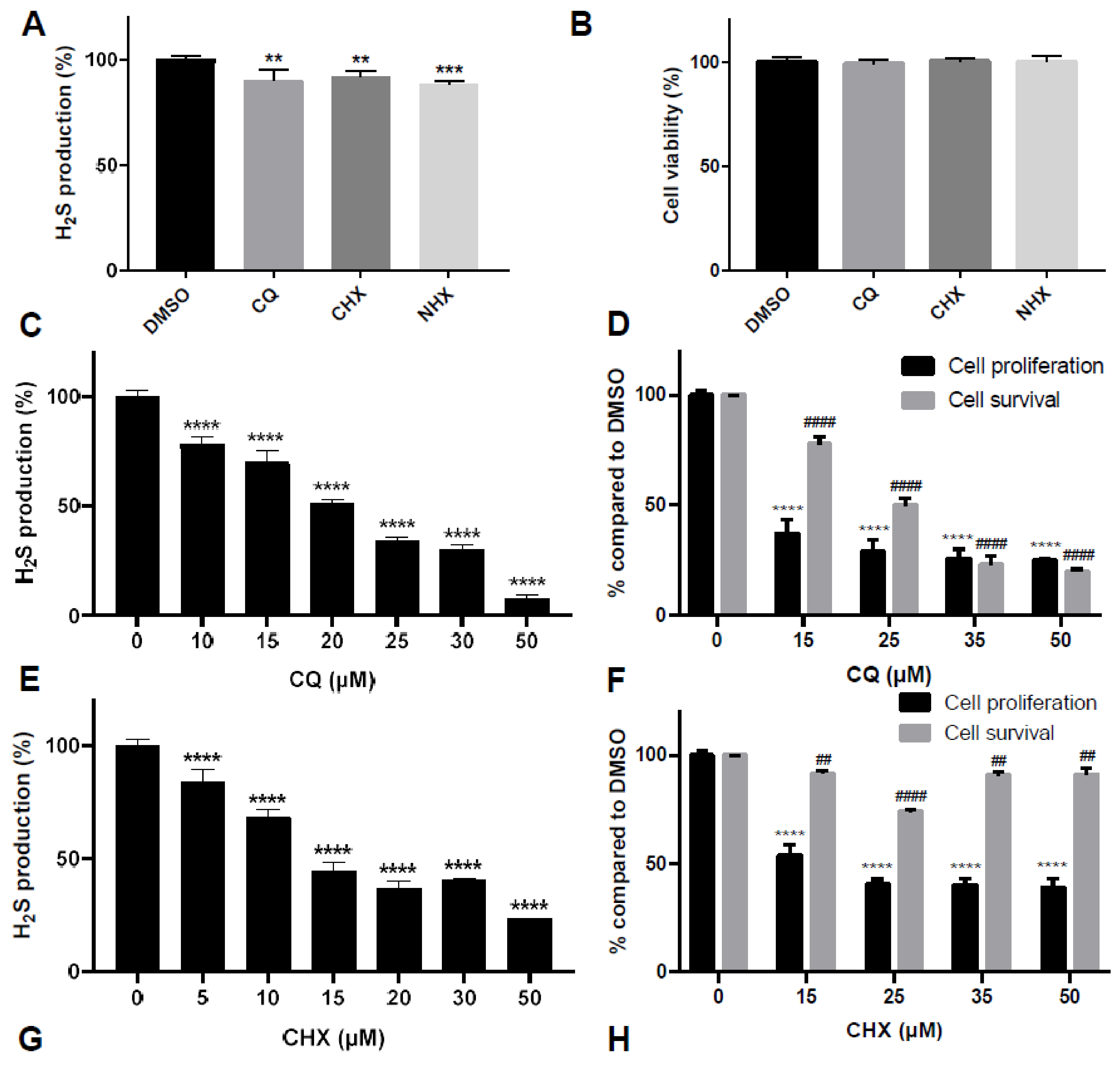
2.2. CQ, CHX and NHX Reduce Human CBS Enzymatic Activity in Liver Cells
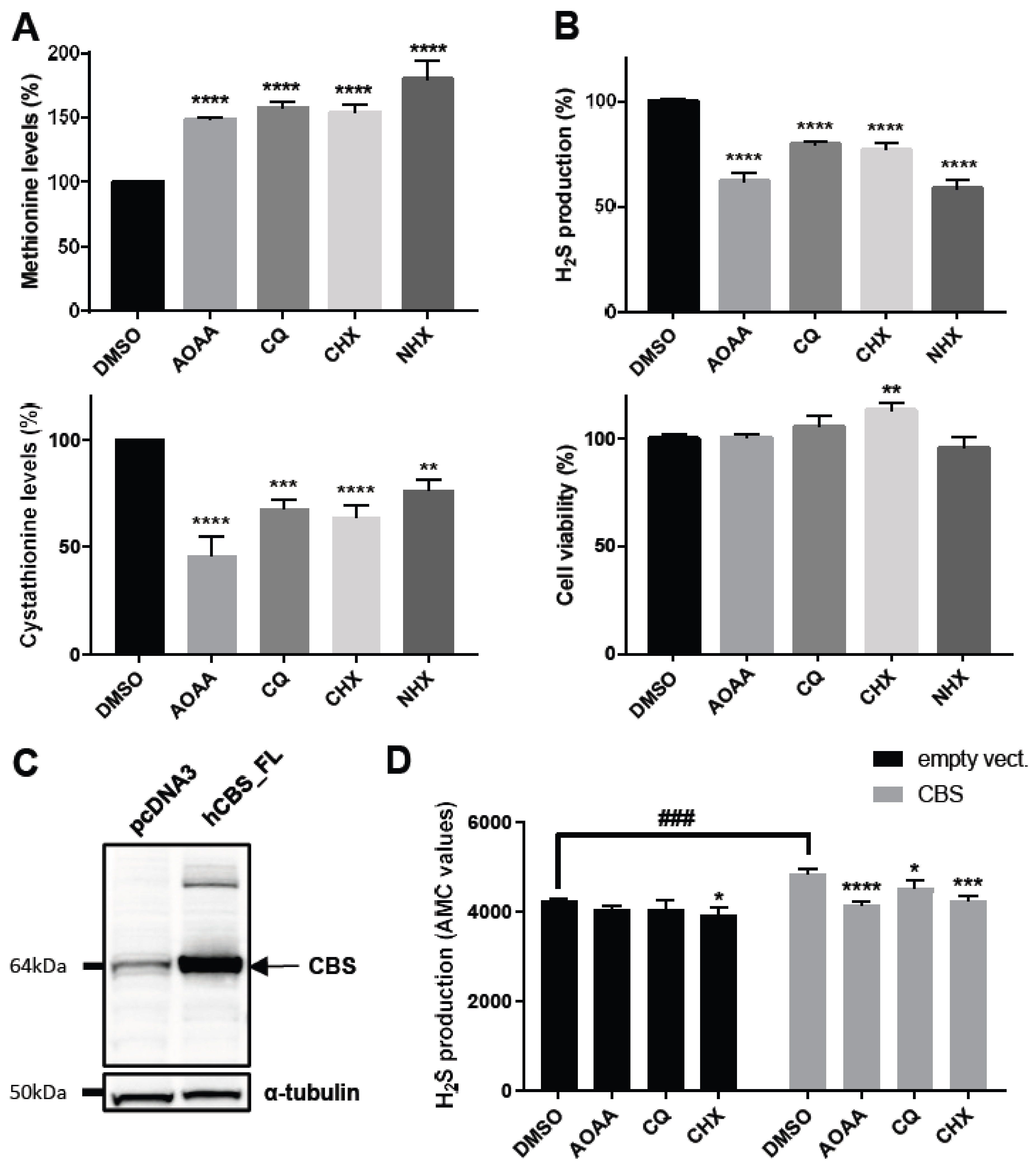
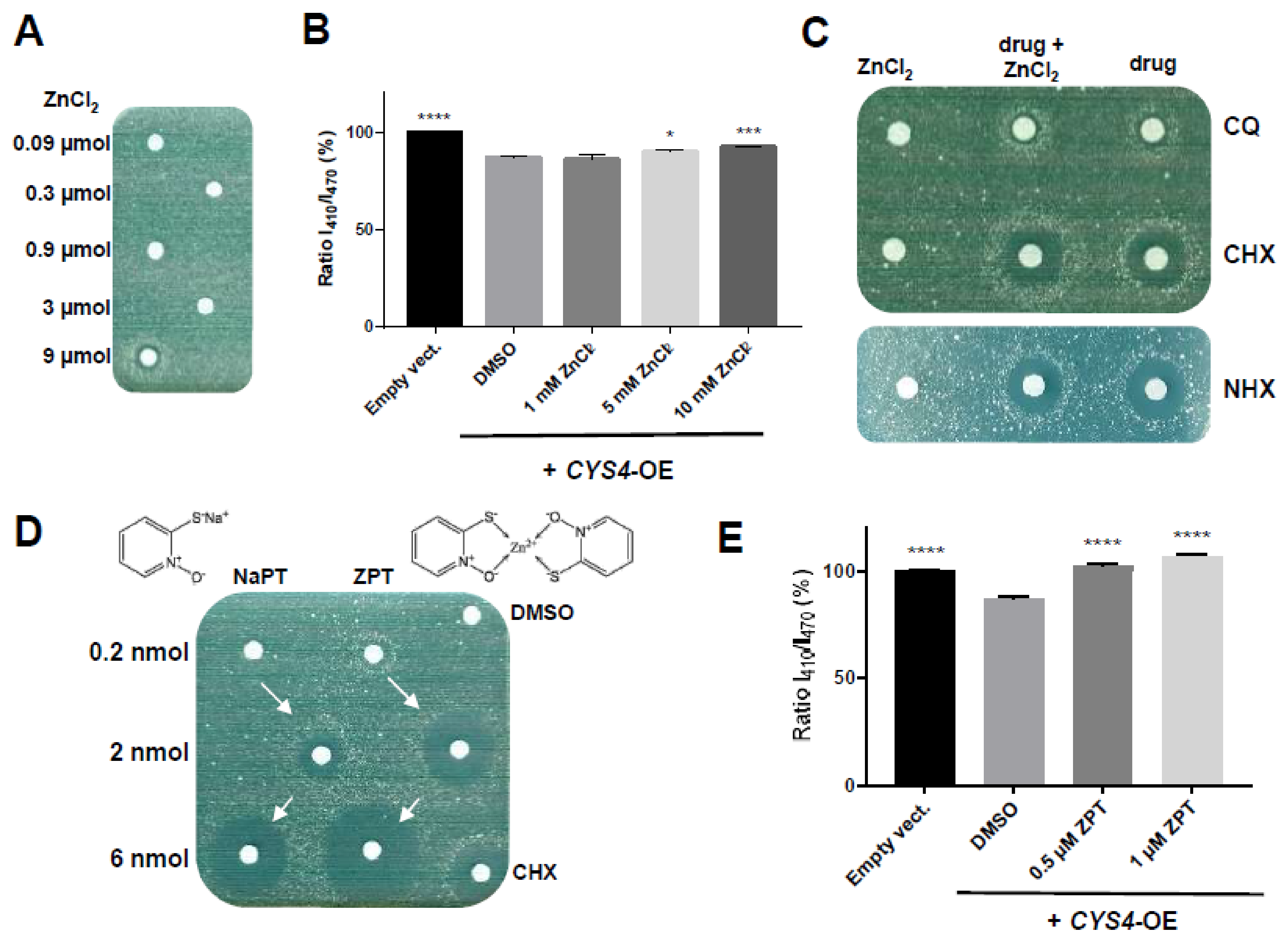
2.3. Decreasing Intracellular Copper Levels Reduces Cystathionine Beta Synthase Enzymatic Activity
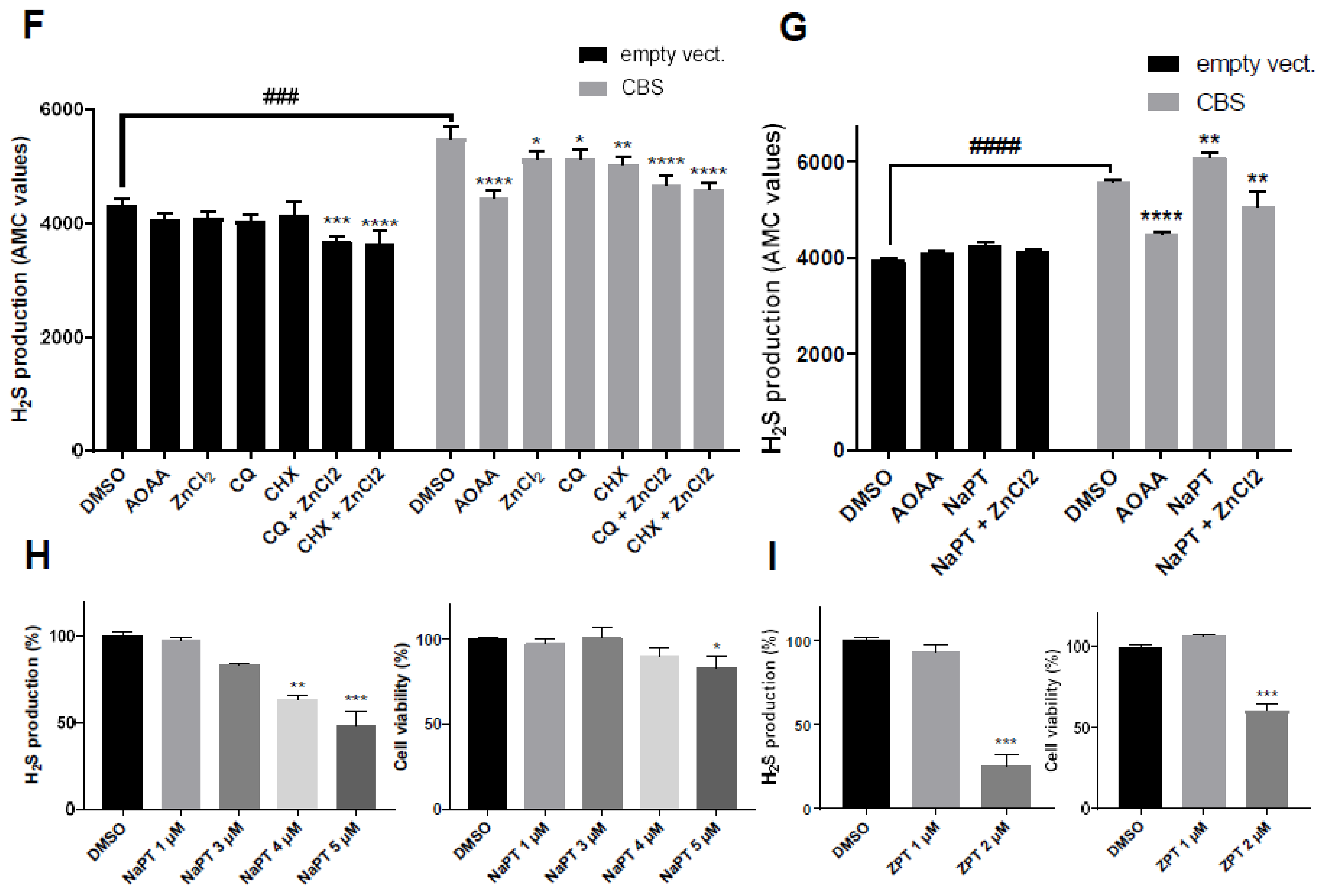
2.4. Zinc Ionophores Reduce the Effects of Cystathionine Beta Synthase Overexpression
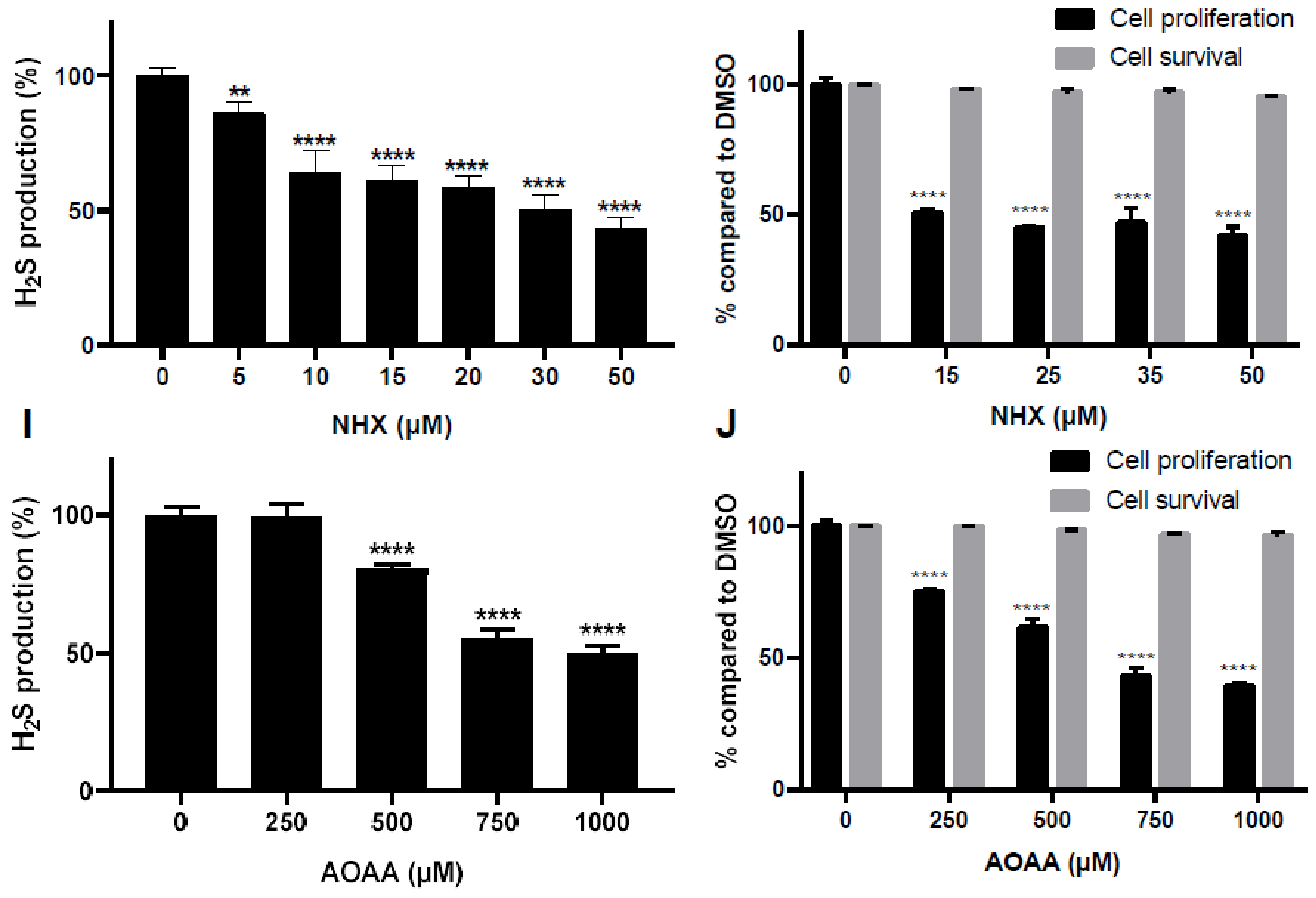
2.5. Effect of CQ, CHX and NHX on CBS Activity Inhibition in Other Human Cellular Models
3. Discussion
3.1. Identification of 8-Hydroxyquinoline Derivatives That Decrease Cystathionine Beta Synthase Enzymatic Activity
3.2. Hypotheses on the Mode of Action of CQ, CHX and NHX
3.3. CQ, CHX and NHX Efficiently Decrease CBS Activity in Other Cellular Models
4. Materials and Methods
4.1. Yeast Strains and Plasmids
4.2. Western Blot
4.3. Determination of Cytosolic pH
4.4. Cell Culture and Drug Treatment
4.5. Mass Spectrometry Quantification of Amino Acids
4.6. Measurement of H2S Production in Live Cells
4.7. Cell Viability Assessment
4.8. Measurement of H2S Production in Cell Lysates
4.9. Measurement of HCT116 Cell Proliferation and Survival
4.10. Measurement of Reactive Oxygen Species (ROS) Level in HepG2 Cells
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mosharov, E.; Cranford, M.R.; Banerjee, R. The Quantitatively Important Relationship between Homocysteine Metabolism and Glutathione Synthesis by the Transsulfuration Pathway and Its Regulation by Redox Changes. Biochemistry 2000, 39, 13005–13011. [Google Scholar] [CrossRef] [PubMed]
- Pogribna, M.; Melnyk, S.; Pogribny, I.; Chango, A.; Yi, P.; James, S.J. Homocysteine Metabolism in Children with Down Syndrome: In Vitro Modulation. Am. J. Hum. Genet. 2001, 69, 88–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraus, J.P.; Janosík, M.; Kozich, V.; Mandell, R.; Shih, V.; Sperandeo, M.P.; Sebastio, G.; de Franchis, R.; Andria, G.; Kluijtmans, L.A.; et al. Cystathionine beta-synthase mutations in homocystinuria. Hum. Mutat. 1999, 13, 362–375. [Google Scholar] [CrossRef]
- Mudd, S.H.; Finkelstein, J.D.; Irreverre, F.; Laster, L. Homocystinuria: An Enzymatic Defect. Science 1964, 143, 1443–1445. [Google Scholar] [CrossRef]
- Mudd, S.H.; Skovby, F.; Levy, H.L.; Pettigrew, K.D.; Wilcken, B.; Pyeritz, R.E.; Andria, G.; Boers, G.H.; Bromberg, I.L.; Cerone, R.; et al. The natural history of homocystinuria due to cystathionine beta-synthase deficiency. Am. J. Hum. Genet. 1985, 37, 1–31. [Google Scholar]
- Kamat, P.K.; Kalani, A.; Tyagi, N. Role of Hydrogen Sulfide in Brain Synaptic Remodeling. Methods Enzymol. 2015, 555, 207–229. [Google Scholar]
- Kimura, H. Hydrogen sulfide: Its production, release and functions. Amino Acids 2011, 41, 113–121. [Google Scholar] [CrossRef]
- Chen, H.B.; Wu, W.N.; Wang, W.; Gu, X.H.; Yu, B.; Wei, B.; Yang, Y.J. Cystathionine-β-synthase-derived hydrogen sulfide is required for amygdalar long-term potentiation and cued fear memory in rats. Pharmacol. Biochem. Behav. 2017, 155, 16–23. [Google Scholar] [CrossRef]
- Szabo, C.; Coletta, C.; Chao, C.; Módis, K.; Szczesny, B.; Papapetropoulos, A.; Hellmich, M.R. Tumor-derived hydrogen sulfide, produced by cystathionine-β-synthase, stimulates bioenergetics, cell proliferation, and angiogenesis in colon cancer. Proc. Natl. Acad. Sci. USA 2013, 110, 12474–12479. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, S.; Saha, S.; Giri, K.; Lanza, I.R.; Nair, K.S.; Jennings, N.B.; Rodriguez-Aguayo, C.; Lopez-Berestein, G.; Basal, E.; Weaver, L.; et al. Cystathionine beta-synthase (CBS) contributes to advanced ovarian cancer progression and drug resistance. PLoS ONE 2013, 8, e79167. [Google Scholar] [CrossRef]
- Sen, S.; Kawahara, B.; Gupta, D.; Tsai, R.; Khachatryan, M.; Roy-Chowdhuri, S.; Bose, S.; Yoon, A.; Faull, K.; Farias-Eisner, R.; et al. Role of cystathionine β-synthase in human breast Cancer. Free Radic. Biol. Med. 2015, 86, 228–238. [Google Scholar] [CrossRef]
- Kamoun, P.; Belardinelli, M.C.; Chabli, A.; Lallouchi, K.; Chadefaux-Vekemans, B. Endogenous hydrogen sulfide overproduction in Down syndrome. Am. J. Med. Genet. 2003, 116A, 310–311. [Google Scholar] [CrossRef]
- Maréchal, D.; Brault, V.; Leon, A.; Martin, D.; Lopes Pereira, P.; Loaëc, N.; Birling, M.C.; Friocourt, G.; Blondel, M.; Hérault, Y. Cbs overdosage is necessary and sufficient to induce cognitive phenotypes in mouse models of Down syndrome and interacts genetically with Dyrk1a. Hum. Mol. Genet. 2019, 28, 1561–1577. [Google Scholar] [CrossRef] [Green Version]
- Szabo, C. The re-emerging pathophysiological role of the cystathionine-β-synthase-hydrogen sulfide system in Down syndrome. FEBS J. 2020, 287, 3150–3160. [Google Scholar] [CrossRef] [Green Version]
- Asimakopoulou, A.; Panopoulos, P.; Chasapis, C.T.; Coletta, C.; Zhou, Z.; Cirino, G.; Giannis, A.; Szabo, C.; Spyroulias, G.A.; Papapetropoulos, A. Selectivity of commonly used pharmacological inhibitors for cystathionine β synthase (CBS) and cystathionine γ lyase (CSE): Characterization of CSE and CBS inhibitors. Br. J. Pharmacol. 2013, 169, 922–932. [Google Scholar] [CrossRef] [Green Version]
- Thorson, M.K.; Majtan, T.; Kraus, J.P.; Barrios, A.M. Identification of Cystathionine β-Synthase Inhibitors Using a Hydrogen Sulfide Selective Probe. Angew. Chem. Int. Ed. 2013, 52, 4641–4644. [Google Scholar] [CrossRef]
- Thorson, M.K.; Van Wagoner, R.M.; Harper, M.K.; Ireland, C.M.; Majtan, T.; Kraus, J.P.; Barrios, A.M. Marine natural products as inhibitors of cystathionine beta-synthase activity. Bioorg. Med. Chem. Lett. 2015, 25, 1064–1066. [Google Scholar] [CrossRef]
- Zhou, Y.; Yu, J.; Lei, X.; Wu, J.; Niu, Q.; Zhang, Y.; Liu, H.; Christen, P.; Gehring, H.; Wu, F. High-throughput tandem-microwell assay identifies inhibitors of the hydrogen sulfide signaling pathway. Chem. Commun. 2013, 49, 11782. [Google Scholar] [CrossRef]
- Druzhyna, N.; Szczesny, B.; Olah, G.; Módis, K.; Asimakopoulou, A.; Pavlidou, A.; Szoleczky, P.; Gerö, D.; Yanagi, K.; Törö, G.; et al. Screening of a composite library of clinically used drugs and well-characterized pharmacological compounds for cystathionine β-synthase inhibition identifies benserazide as a drug potentially suitable for repurposing for the experimental therapy of colon cancer. Pharmacol. Res. 2016, 113, 18–37. [Google Scholar]
- Shan, X.; Kruger, W.D. Correction of disease-causing CBS mutations in yeast. Nat. Genet. 1998, 19, 91–93. [Google Scholar] [CrossRef]
- Oluwatosin, Y.E.; Kane, P.M. Mutations in the CYS4 Gene Provide Evidence for Regulation of the Yeast Vacuolar H+-ATPase by Oxidation and Reduction In Vivo. J. Biol. Chem. 1997, 272, 28149–28157. [Google Scholar] [CrossRef] [Green Version]
- Whiteman, M.; Le Trionnaire, S.; Chopra, M.; Fox, B.; Whatmore, J. Emerging role of hydrogen sulfide in health and disease: Critical appraisal of biomarkers and pharmacological tools. Clin. Sci. 2011, 121, 459–488. [Google Scholar] [CrossRef]
- Hellmich, M.R.; Chao, C.; Módis, K.; Ding, Y.; Zatarain, J.R.; Thanki, K.; Maskey, M.; Druzhyna, N.; Untereiner, A.A.; Ahmad, A.; et al. Efficacy of Novel Aminooxyacetic Acid Prodrugs in Colon Cancer Models: Towards Clinical Translation of the Cystathionine β-Synthase Inhibition Concept. Biomolecules 2021, 11, 1073. [Google Scholar] [CrossRef]
- El Sakka, N.; Gould, I. Role of old antimicrobial agents in the management of urinary tract infection. Expert Rev. Clin. Pharmacol. 2016, 9, 1047–1056. [Google Scholar] [CrossRef]
- Ritchie, C.W.; Bush, A.I.; Masters, C. Metal-protein attenuating compounds and Alzheimer’s disease. Expert Opin. Investig. Drugs 2004, 13, 1585–1592. [Google Scholar] [CrossRef]
- Nguyen, T.; Hamby, A.; Massa, S.M. Clioquinol down-regulates mutant huntingtin expression in vitro and mitigates pathology in a Huntington’s disease mouse model. Proc. Natl. Acad. Sci. USA 2005, 102, 11840–11845. [Google Scholar] [CrossRef] [Green Version]
- Cherny, R.A.; Legg, J.T.; McLean, C.A.; Fairlie, D.P.; Huang, X.; Atwood, C.S.; Beyreuther, K.; Tanzi, R.E.; Masters, C.L.; Bush, A.I. Aqueous Dissolution of Alzheimer’s Disease Aβ Amyloid Deposits by Biometal Depletion. J. Biol. Chem. 1999, 274, 23223–23228. [Google Scholar] [CrossRef] [Green Version]
- Grossi, C.; Francese, S.; Casini, A.; Rosi, M.C.; Luccarini, I.; Fiorentini, A.; Gabbiani, C.; Messori, L.; Moneti, G.; Casamenti, F. Clioquinol Decreases Amyloid-β Burden and Reduces Working Memory Impairment in a Transgenic Mouse Model of Alzheimer’s Disease. J. Alzheimer’s Dis. 2009, 17, 423–440. [Google Scholar] [CrossRef]
- Ding, W.Q.; Lind, S.E. Metal ionophores—An emerging class of anticancer drugs. IUBMB Life 2009, 61, 1013–1018. [Google Scholar] [CrossRef]
- Bar-Or, D.; Rael, L.; Thomas, G.; Kraus, J. Inhibitory Effect of Copper on Cystathionine beta-Synthase Activity: Protective Effect of an Analog of the Human Albumin N-Terminus. Protein Pept. Lett. 2005, 12, 271–273. [Google Scholar] [CrossRef]
- Zuhra, K.; Panagaki, T.; Randi, E.B.; Augsburger, F.; Blondel, M.; Friocourt, G.; Herault, Y.; Szabo, C. Mechanism of cystathionine-β-synthase inhibition by disulfiram: The role of bis(N,N-diethyldithiocarbamate)-copper(II). Biochem. Pharmacol. 2020, 182, 114267. [Google Scholar] [CrossRef] [PubMed]
- Schuller, A.; Auffermann, G.; Zoschke, K.; Schmidt, U.; Ostermann, K.; Rödel, G. Overexpression of ctr1Δ300, a high-affinity copper transporter with deletion of the cytosolic C-terminus in Saccharomyces cerevisiae under excess copper, leads to disruption of transition metal homeostasis and transcriptional remodelling of cellular processes. Yeast 2013, 30, 201–218. [Google Scholar] [PubMed]
- Jungmann, J.; Reins, H.A.; Lee, J.; Romeo, A.; Hassett, R.; Kosman, D.; Jentsch, S. MAC1, a nuclear regulatory protein related to Cu-dependent transcription factors is involved in Cu/Fe utilization and stress resistance in yeast. EMBO J. 1993, 12, 5051–5056. [Google Scholar] [CrossRef] [PubMed]
- Adlard, P.A.; Cherny, R.A.; Finkelstein, D.I.; Gautier, E.; Robb, E.; Cortes, M.; Volitakis, I.; Liu, X.; Smith, J.P.; Perez, K.; et al. Rapid Restoration of Cognition in Alzheimer’s Transgenic Mice with 8-Hydroxy Quinoline Analogs Is Associated with Decreased Interstitial Aβ. Neuron 2008, 59, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Faux, N.G.; Ritchie, C.W.; Gunn, A.; Rembach, A.; Tsatsanis, A.; Bedo, J.; Harrison, J.; Lannfelt, L.; Blennow, K.; Zetterberg, H.; et al. PBT2 Rapidly Improves Cognition in Alzheimer’s Disease: Additional Phase II Analyses. J. Alzheimer’s Dis. 2010, 20, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Colvin, R.A.; Bush, A.I.; Volitakis, I.; Fontaine, C.P.; Thomas, D.; Kikuchi, K.; Holmes, W.R. Insights into Zn2+ homeostasis in neurons from experimental and modeling studies. Am. J. Physiol. Cell Physiol. 2008, 294, C726–C742. [Google Scholar] [CrossRef]
- Ding, W.Q.; Liu, B.; Vaught, J.L.; Yamauchi, H.; Lind, S.E. Anticancer Activity of the Antibiotic Clioquinol. Cancer Res. 2005, 65, 3389–3395. [Google Scholar] [CrossRef] [Green Version]
- Ding, W.Q.; Yu, H.J.; Lind, S.E. Zinc-binding compounds induce cancer cell death via distinct modes of action. Cancer Lett. 2008, 271, 251–259. [Google Scholar] [CrossRef]
- Park, M.H.; Lee, S.J.; Byun, H.R.; Kim, Y.; Oh, Y.J.; Koh, J.Y.; Hwang, J.J. Clioquinol induces autophagy in cultured astrocytes and neurons by acting as a zinc ionophore. Neurobiol. Dis. 2011, 42, 242–251. [Google Scholar] [CrossRef]
- Jiang, H.; Taggart, J.E.; Zhang, X.; Benbrook, D.M.; Lind, S.E.; Ding, W.Q. Nitroxoline (8-hydroxy-5-nitroquinoline) is more a potent anti-cancer agent than clioquinol (5-chloro-7-iodo-8-quinoline). Cancer Lett. 2011, 312, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Sanokawa-Akakura, R.; Akakura, S.; Tabibzadeh, S. Replicative Senescence in Human Fibroblasts Is Delayed by Hydrogen Sulfide in a NAMPT/SIRT1 Dependent Manner. PLoS ONE 2016, 11, e0164710. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.; Wang, M.; Ju, L.; Wang, C.; Zhu, Y. Hydrogen sulfide induces human colon cancer cell proliferation: Role of Akt, ERK and p21. Cell. Biol. Int. 2010, 34, 565–572. [Google Scholar] [CrossRef]
- Yue, T.; Zuo, S.; Bu, D.; Zhu, J.; Chen, S.; Ma, Y.; Ma, J.; Guo, S.; Wen, L.; Zhang, X.; et al. Aminooxyacetic acid (AOAA) sensitizes colon cancer cells to oxaliplatin via exaggerating apoptosis induced by ROS. J. Cancer 2020, 11, 1828–1838. [Google Scholar] [CrossRef]
- Kashiwamata, S.; Greenberg, D.M. Studies on cystathionine synthase of rat liver properties of the highly purified enzyme. Biochem. Biophys. Acta BBA Enzymol. 1970, 212, 488–500. [Google Scholar] [CrossRef]
- Evans, J.C.; Huddler, D.P.; Jiracek, J.; Castro, C.; Millian, N.S.; Garrow, T.A.; Ludwig, M.L. Betaine-Homocysteine Methyltransferase. Structure 2002, 10, 1159–1171. [Google Scholar] [CrossRef] [Green Version]
- Millian, N.S.; Garrow, T.A. Human Betaine–Homocysteine Methyltransferase Is a Zinc Metalloenzyme. Arch. Biochem. Biophys. 1998, 356, 93–98. [Google Scholar] [CrossRef]
- Bremner, I.; Beattie, J.H. Copper and zinc metabolism in health and disease: Speciation and interactions. Proc. Nutr. Soc. 1995, 54, 489–499. [Google Scholar] [CrossRef] [Green Version]
- Niu, W.; Wang, J.; Qian, J.; Wang, M.; Wu, P.; Chen, F.; Yan, S. Allosteric control of human cystathionine β-synthase activity by a redox active disulfide bond. J. Biol. Chem. 2018, 293, 2523–2533. [Google Scholar] [CrossRef] [Green Version]
- Tamura, T.; Hong, K.H.; Mizuno, Y.; Johnston, K.E.; Keen, C.L. Folate and homocysteine metabolism in copper-deficient rats. Biochem. Biophys. Acta BBA Gen. Subj. 1999, 1427, 351–356. [Google Scholar] [CrossRef]
- Vitvitsky, V.; Mosharov, E.; Tritt, M.; Ataullakhanov, F.; Banerjee, R. Redox regulation of homocysteine-dependent glutathione synthesis. Redox Rep. 2013, 8, 57–63. [Google Scholar] [CrossRef]
- Banerjee, R.; Zou, C. Redox regulation and reaction mechanism of human cystathionine-β-synthase: A PLP-dependent hemesensor protein. Arch. Biochem. Biophys. 2005, 433, 144–156. [Google Scholar] [CrossRef]
- Todorich, B.M.; Connor, J.R. Redox Metals in Alzheimer’s Disease. Ann. N. Y. Acad. Sci. 2004, 1012, 171–178. [Google Scholar] [CrossRef]
- Kamoun, P.P. Mental retardation in Down syndrome: Two ways to treat. Med. Hypotheses 2019, 131, 109289. [Google Scholar] [CrossRef]
- Li, C.; Wang, J.; Zhou, B. The Metal Chelating and Chaperoning Effects of Clioquinol: Insights from Yeast Studies. J. Alzheimer’s Dis. 2010, 21, 1249–1262. [Google Scholar] [CrossRef] [Green Version]
- Lima, A.S.; Cardoso, B.R.; Cozzolino, S.F. Nutritional Status of Zinc in Children with Down Syndrome. Biol. Trace Elem. Res. 2010, 133, 20–28. [Google Scholar] [CrossRef]
- Longtine, M.S.; McKenzie, A., III; Demarini, D.J.; Shah, N.G.; Wach, A.; Brachat, A.; Philippsen, P.; Pringle, J.R. Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast 1998, 14, 953–961. [Google Scholar] [CrossRef]
- Miesenböck, G.; De Angelis, D.A.; Rothman, J.E. Visualizing secretion and synaptic transmission with pH-sensitive green fluorescent proteins. Nature 1998, 394, 192–195. [Google Scholar] [CrossRef]
- Dechant, R.; Binda, M.; Lee, S.S.; Pelet, S.; Winderickx, J.; Peter, M. Cytosolic pH is a second messenger for glucose and regulates the PKA pathway through V-ATPase. EMBO J. 2010, 29, 2515–2526. [Google Scholar] [CrossRef] [Green Version]
- Brett, C.L.; Tukaye, D.N.; Mukherjee, S.; Rao, R. The yeast endosomal Na+K+/H+ exchanger Nhx1 regulates cellular pH to control vesicle trafficking. Mol. Biol. Cell 2005, 16, 1396–1405. [Google Scholar] [CrossRef] [Green Version]
- Baggett, J.J.; Shaw, J.D.; Sciambi, C.J.; Watson, H.A.; Wendland, B. Fluorescent labeling of yeast. In Current Protocols in Cell Biology; Wiley: Hoboken, NJ, USA, 2003; Chapter 4, Unit 4.13. [Google Scholar]
- Sambade, M.; Alba, M.; Smardon, A.M.; West, R.W.; Kane, P.M. A genomic screen for yeast vacuolar membrane ATPase mutants. Genetics 2005, 170, 1539–1551. [Google Scholar] [CrossRef] [Green Version]
- Mendes, M.I.; Colaço, H.G.; Smith, D.E.; Ramos, R.J.; Pop, A.; van Dooren, S.J.; Tavares de Almeida, I.; Kluijtmans, L.A.; Janssen, M.C.; Rivera, I.; et al. Reduced response of Cystathionine Beta-Synthase (CBS) to S-Adenosylmethionine (SAM): Identification and functional analysis of CBS gene mutations in Homocystinuria patients. J. Inherit. Metab. Dis. 2014, 37, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Dhanasekaran, A.R.; Gardiner, K.J. Mouse models of Down syndrome: Gene content and consequences. Mamm. Genome 2016, 27, 538–555. [Google Scholar] [CrossRef] [PubMed]
- Majtan, T.; Pey, A.L.; Fernández, R.; Fernández, J.A.; Martínez-Cruz, L.A.; Kraus, J.P. Domain Organization, Catalysis and Regulation of Eukaryotic Cystathionine Beta-Synthases. PLoS ONE 2014, 9, e105290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhaoui, M.; Auchère, F.; Blaiseau, P.L.; Lesuisse, E.; Landoulsi, A.; Camadro, J.M.; Haguenauer-Tsapis, R.; Belgareh-Touzé, N. Gex1 is a yeast glutathione exchanger that interferes with pH and redox homeostasis. Mol. Biol. Cell 2011, 22, 2054–2067. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conan, P.; Léon, A.; Gourdel, M.; Rollet, C.; Chaïr, L.; Caroff, N.; Le Goux, N.; Le Jossic-Corcos, C.; Sinane, M.; Gentile, L.; et al. Identification of 8-Hydroxyquinoline Derivatives That Decrease Cystathionine Beta Synthase (CBS) Activity. Int. J. Mol. Sci. 2022, 23, 6769. https://doi.org/10.3390/ijms23126769
Conan P, Léon A, Gourdel M, Rollet C, Chaïr L, Caroff N, Le Goux N, Le Jossic-Corcos C, Sinane M, Gentile L, et al. Identification of 8-Hydroxyquinoline Derivatives That Decrease Cystathionine Beta Synthase (CBS) Activity. International Journal of Molecular Sciences. 2022; 23(12):6769. https://doi.org/10.3390/ijms23126769
Chicago/Turabian StyleConan, Pierre, Alice Léon, Mathilde Gourdel, Claire Rollet, Loubna Chaïr, Noéline Caroff, Nelig Le Goux, Catherine Le Jossic-Corcos, Maha Sinane, Lucile Gentile, and et al. 2022. "Identification of 8-Hydroxyquinoline Derivatives That Decrease Cystathionine Beta Synthase (CBS) Activity" International Journal of Molecular Sciences 23, no. 12: 6769. https://doi.org/10.3390/ijms23126769
APA StyleConan, P., Léon, A., Gourdel, M., Rollet, C., Chaïr, L., Caroff, N., Le Goux, N., Le Jossic-Corcos, C., Sinane, M., Gentile, L., Maillebouis, L., Loaëc, N., Martin, J., Vilaire, M., Corcos, L., Mignen, O., Croyal, M., Voisset, C., Bihel, F., & Friocourt, G. (2022). Identification of 8-Hydroxyquinoline Derivatives That Decrease Cystathionine Beta Synthase (CBS) Activity. International Journal of Molecular Sciences, 23(12), 6769. https://doi.org/10.3390/ijms23126769