
Drugs, Guts, Brains, but Not Rock and Roll: The Need to Consider the Role of Gut Microbiota in Contemporary Mental Health and Wellness of Emerging Adults

 , , and
, , and {kind=link}
{kind=link}
Abstract
:1. Introduction: Emerging Adulthood, Mental Health, and the Gut Microbiome
2. The Gut–Brain–Microbiota (GBM) and the Stress Response
3. Associations between GM and Brain Health
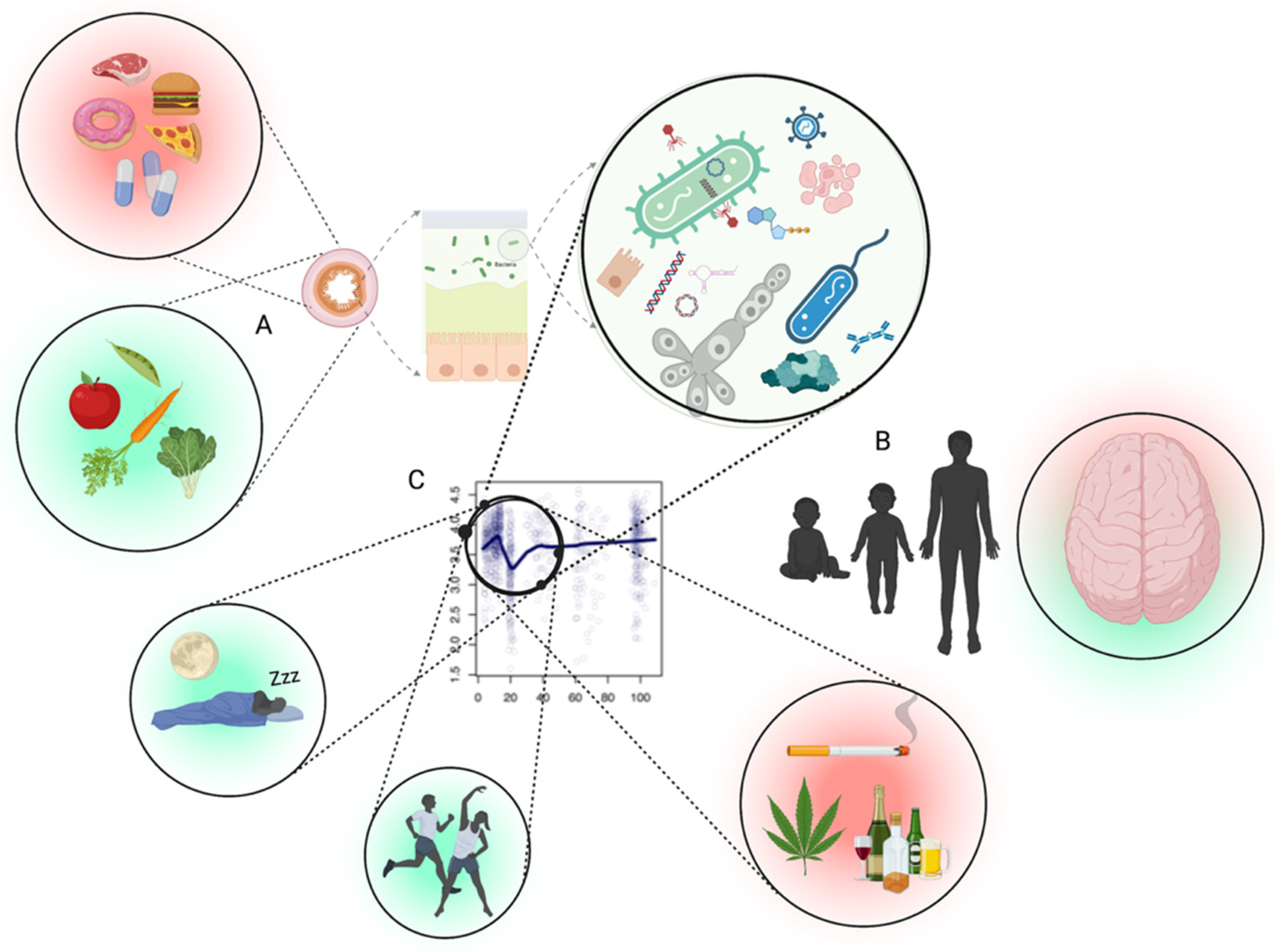
4. Lifestyle Factors Affecting GM and Mental Health: Diet, Physical Activity/Exercise, Substance Use, Sleep
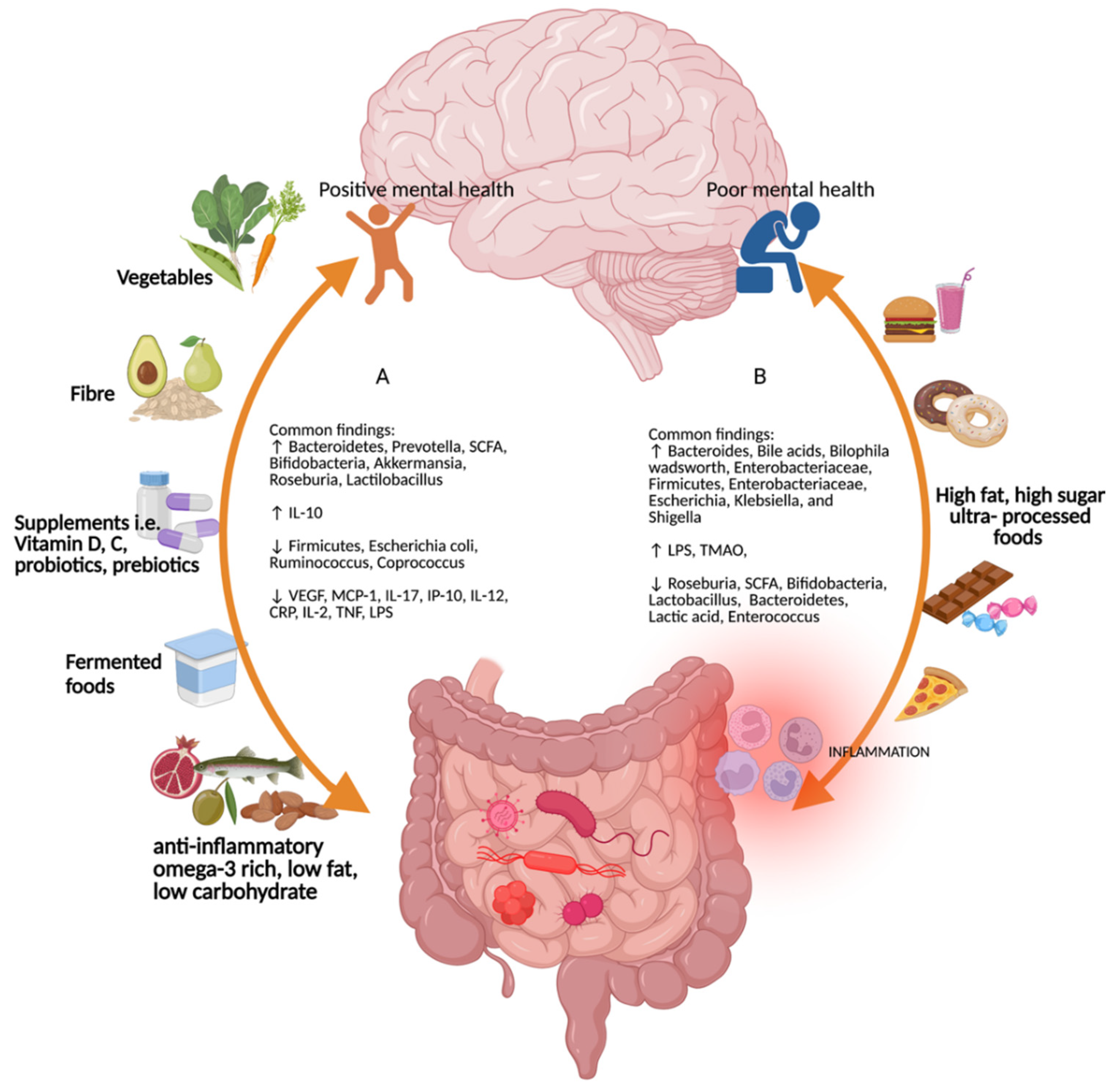
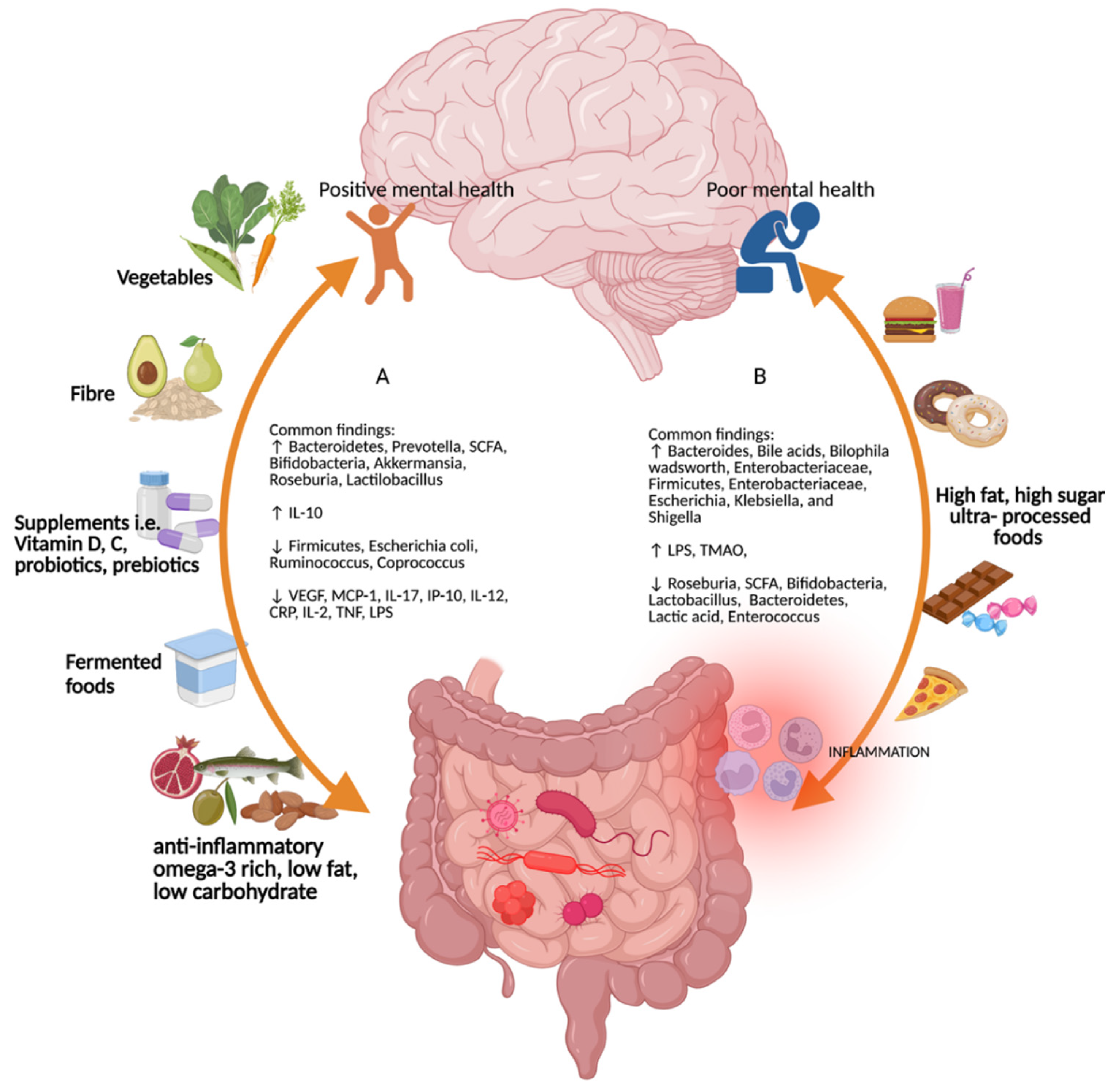
4.1. Diet
4.2. Physical Activity and Exercise
4.3. Substance Use
4.3.1. Nicotine
4.3.2. Alcohol
4.3.3. Cannabis
4.4. Sleep/Circadian Rhythm
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Romeo, R.D.; Bellani, R.; Karatsoreos, I.N.; Chhua, N.; Vernov, M.; Conrad, C.D.; McEwen, B.S. Stress history and pubertal development interact to shape hypothalamic-pituitary-adrenal axis plasticity. Endocrinology 2006, 147, 1664–1674. [Google Scholar] [CrossRef] [PubMed]
- Spear, L.P. Adolescent neurodevelopment. J. Adolesc. Health 2013, 52, S7–S13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnett, S.; Sebastian, C.; Cohen Kadosh, K.; Blakemore, S.J. The social brain in adolescence: Evidence from functional magnetic resonance imaging and behavioural studies. Neurosci. Biobehav. Rev. 2011, 35, 1654–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaworska, N.; MacQueen, G. Adolescence as a unique developmental period. J. Psychiatry Neurosci. 2015, 40, 291–293. [Google Scholar] [CrossRef]
- McVey Neufeld, K.A.; Luczynski, P.; Seira Oriach, C.; Dinan, T.G.; Cryan, J.F. What’s bugging your teen?-The microbiota and adolescent mental health. Neurosci. Biobehav. Rev. 2016, 70, 300–312. [Google Scholar] [CrossRef]
- Sudo, N. Microbiome, HPA axis and production of endocrine hormones in the gut. Adv. Exp. Med. Biol. 2014, 817, 177–194. [Google Scholar] [CrossRef]
- Cresci, G.A.; Bawden, E. Gut Microbiome: What We Do and Don’t Know. Nutr. Clin. Pract. 2015, 30, 734–746. [Google Scholar] [CrossRef] [Green Version]
- Fulkerson, J.A.; Sherwood, N.E.; Perry, C.L.; Neumark-Sztainer, D.; Story, M. Depressive symptoms and adolescent eating and health behaviors: A multifaceted view in a population-based sample. Prev. Med. 2004, 38, 865–875. [Google Scholar] [CrossRef]
- Robillard, R.; Carpenter, J.S.; Rogers, N.L.; Fares, S.; Grierson, A.B.; Hermens, D.F.; Naismith, S.L.; Mullin, S.J.; Feilds, K.L.; Glozier, N.; et al. Circadian rhythms and psychiatric profiles in young adults with unipolar depressive disorders. Transl. Psychiatry 2018, 8, 213. [Google Scholar] [CrossRef] [Green Version]
- Toyoura, M.; Miike, T.; Tajima, S.; Matsuzawa, S.; Konishi, Y. Inadequate sleep as a contributor to impaired glucose tolerance: A cross-sectional study in children, adolescents, and young adults with circadian rhythm sleep-wake disorder. Pediatr. Diabetes 2020, 21, 557–564. [Google Scholar] [CrossRef]
- Lach, G.; Fulling, C.; Bastiaanssen, T.F.S.; Fouhy, F.; Donovan, A.N.O.; Ventura-Silva, A.P.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Enduring neurobehavioral effects induced by microbiota depletion during the adolescent period. Transl. Psychiatry 2020, 10, 382. [Google Scholar] [CrossRef] [PubMed]
- Bian, G.; Gloor, G.B.; Gong, A.; Jia, C.; Zhang, W.; Hu, J.; Zhang, H.; Zhang, Y.; Zhou, Z.; Zhang, J.; et al. The Gut Microbiota of Healthy Aged Chinese Is Similar to That of the Healthy Young. mSphere 2017, 2, e00327-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Li, J.; Lu, S.; Han, N.; Miao, J.; Zhang, T.; Qiang, Y.; Kong, Y.; Wang, H.; Gao, T.; et al. Gut microbiota community characteristics and disease-related microorganism pattern in a population of healthy Chinese people. Sci. Rep. 2019, 9, 1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Mental Health. Available online: https://www.who.int/health-topics/mental-health#tab=tab_1 (accessed on 1 December 2021).
- Wang, H.X.; Wang, Y.P. Gut Microbiota-brain Axis. Chin. Med. J. (Engl.) 2016, 129, 2373–2380. [Google Scholar] [CrossRef] [PubMed]
- Simpson, C.A.; Diaz-Arteche, C.; Eliby, D.; Schwartz, O.S.; Simmons, J.G.; Cowan, C.S.M. The gut microbiota in anxiety and depression—A systematic review. Clin. Psychol. Rev. 2021, 83, 101943. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; Gonzalez, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [Green Version]
- van de Wouw, M.; Boehme, M.; Lyte, J.M.; Wiley, N.; Strain, C.; O’Sullivan, O.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Short-chain fatty acids: Microbial metabolites that alleviate stress-induced brain-gut axis alterations. J. Physiol. 2018, 596, 4923–4944. [Google Scholar] [CrossRef]
- Mogilevski, T. The bi-directional role of the gut-brain axis in inflammatory and other gastrointestinal diseases. Curr. Opin. Gastroenterol. 2021, 37, 572–577. [Google Scholar] [CrossRef]
- Pellegrini, C.; Antonioli, L.; Calderone, V.; Colucci, R.; Fornai, M.; Blandizzi, C. Microbiota-gut-brain axis in health and disease: Is NLRP3 inflammasome at the crossroads of microbiota-gut-brain communications? Prog. Neurobiol. 2020, 191, 101806. [Google Scholar] [CrossRef]
- Desbonnet, L.; Clarke, G.; Traplin, A.; O’Sullivan, O.; Crispie, F.; Moloney, R.D.; Cotter, P.D.; Dinan, T.G.; Cryan, J.F. Gut microbiota depletion from early adolescence in mice: Implications for brain and behaviour. Brain Behav. Immun. 2015, 48, 165–173. [Google Scholar] [CrossRef]
- Kaur, G.; Behl, T.; Bungau, S.; Kumar, A.; Uddin, M.S.; Mehta, V.; Zengin, G.; Mathew, B.; Shah, M.A.; Arora, S. Dysregulation of the Gut-Brain Axis, Dysbiosis and Influence of Numerous Factors on Gut Microbiota Associated Parkinson’s Disease. Curr. Neuropharmacol. 2021, 19, 233–247. [Google Scholar] [CrossRef] [PubMed]
- Carlessi, A.S.; Borba, L.A.; Zugno, A.I.; Quevedo, J.; Réus, G.Z. Gut microbiota-brain axis in depression: The role of neuroinflammation. Eur. J. Neurosci. 2021, 53, 222–235. [Google Scholar] [CrossRef]
- Forslund, S.K.; Chakaroun, R.; Zimmermann-Kogadeeva, M.; Markó, L.; Aron-Wisnewsky, J.; Nielsen, T.; Moitinho-Silva, L.; Schmidt, T.S.B.; Falony, G.; Vieira-Silva, S.; et al. Combinatorial, additive and dose-dependent drug–microbiome associations. Nature 2021, 600, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Sogari, G.; Velez-Argumedo, C.; Gomez, M.I.; Mora, C. College Students and Eating Habits: A Study Using An Ecological Model for Healthy Behavior. Nutrients 2018, 10, 1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, N.; Laska, M.N.; Neumark-Sztainer, D. Food Insecurity, Diet Quality, Home Food Availability, and Health Risk Behaviors Among Emerging Adults: Findings From the EAT 2010–2018 Study. Am. J. Public Health 2020, 110, 1422–1428. [Google Scholar] [CrossRef]
- Martinez Leo, E.E.; Segura Campos, M.R. Effect of ultra-processed diet on gut microbiota and thus its role in neurodegenerative diseases. Nutrition 2020, 71, 110609. [Google Scholar] [CrossRef]
- Moreira, A.R.O.; Batista, R.F.L.; Ladeira, L.L.C.; Thomaz, E.; Alves, C.M.C.; Saraiva, M.C.; Silva, A.A.M.; Brondani, M.A.; Ribeiro, C.C.C. Higher sugar intake is associated with periodontal disease in adolescents. Clin. Oral Investig. 2021, 25, 983–991. [Google Scholar] [CrossRef]
- Chopra, C.; Mandalika, S.; Kinger, N. Does diet play a role in the prevention and management of depression among adolescents? A narrative review. Nutr. Health 2021, 27, 243–263. [Google Scholar] [CrossRef]
- Ruiz, L.D.; Zuelch, M.L.; Dimitratos, S.M.; Scherr, R.E. Adolescent Obesity: Diet Quality, Psychosocial Health, and Cardiometabolic Risk Factors. Nutrients 2019, 12, 43. [Google Scholar] [CrossRef] [Green Version]
- Davis, E.C.; Dinsmoor, A.M.; Wang, M.; Donovan, S.M. Microbiome Composition in Pediatric Populations from Birth to Adolescence: Impact of Diet and Prebiotic and Probiotic Interventions. Dig. Dis. Sci. 2020, 65, 706–722. [Google Scholar] [CrossRef] [Green Version]
- Marx, W.; Lane, M.; Hockey, M.; Aslam, H.; Berk, M.; Walder, K.; Borsini, A.; Firth, J.; Pariante, C.M.; Berding, K.; et al. Diet and depression: Exploring the biological mechanisms of action. Mol. Psychiatry 2021, 26, 134–150. [Google Scholar] [CrossRef] [PubMed]
- Dash, S.; Clarke, G.; Berk, M.; Jacka, F.N. The gut microbiome and diet in psychiatry: Focus on depression. Curr. Opin. Psychiatry 2015, 28, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Illiano, P.; Brambilla, R.; Parolini, C. The mutual interplay of gut microbiota, diet and human disease. FEBS J. 2020, 287, 833–855. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mantrana, I.; Selma-Royo, M.; Alcantara, C.; Collado, M.C. Shifts on Gut Microbiota Associated to Mediterranean Diet Adherence and Specific Dietary Intakes on General Adult Population. Front. Microbiol. 2018, 9, 890. [Google Scholar] [CrossRef]
- Lima-Ojeda, J.M.; Rupprecht, R.; Baghai, T.C. “I Am I and My Bacterial Circumstances”: Linking Gut Microbiome, Neurodevelopment, and Depression. Front. Psychiatry 2017, 8, 153. [Google Scholar] [CrossRef] [Green Version]
- Seitz, J.; Dahmen, B.; Keller, L.; Herpertz-Dahlmann, B. Gut Feelings: How Microbiota Might Impact the Development and Course of Anorexia Nervosa. Nutrients 2020, 12, 3295. [Google Scholar] [CrossRef]
- Hata, T.; Miyata, N.; Takakura, S.; Yoshihara, K.; Asano, Y.; Kimura-Todani, T.; Yamashita, M.; Zhang, X.T.; Watanabe, N.; Mikami, K.; et al. The Gut Microbiome Derived From Anorexia Nervosa Patients Impairs Weight Gain and Behavioral Performance in Female Mice. Endocrinology 2019, 160, 2441–2452. [Google Scholar] [CrossRef]
- Leong, K.S.W.; Jayasinghe, T.N.; Wilson, B.C.; Derraik, J.G.B.; Albert, B.B.; Chiavaroli, V.; Svirskis, D.M.; Beck, K.L.; Conlon, C.A.; Jiang, Y.; et al. Effects of Fecal Microbiome Transfer in Adolescents With Obesity: The Gut Bugs Randomized Controlled Trial. JAMA Netw. Open 2020, 3, e2030415. [Google Scholar] [CrossRef]
- Borsini, A.; Alboni, S.; Horowitz, M.A.; Tojo, L.M.; Cannazza, G.; Su, K.P.; Pariante, C.M.; Zunszain, P.A. Rescue of IL-1beta-induced reduction of human neurogenesis by omega-3 fatty acids and antidepressants. Brain Behav. Immun. 2017, 65, 230–238. [Google Scholar] [CrossRef] [Green Version]
- Kanoski, S.E.; Davidson, T.L. Western diet consumption and cognitive impairment: Links to hippocampal dysfunction and obesity. Physiol. Behav. 2011, 103, 59–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapia-Serrano, M.A.; Esteban-Cornejo, I.; Rodriguez-Ayllon, M.; Vaquero-Solis, M.; Sanchez-Oliva, D.; Sanchez-Miguel, P.A. Adherence to the Mediterranean diet and academic performance in adolescents: Does BMI status moderate this association? Clin. Nutr. 2021, 40, 4465–4472. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Lof, M.; Chen, R.; Hultman, C.M.; Fang, F.; Sandin, S. Mediterranean diet and depression: A population-based cohort study. Int. J. Behav. Nutr. Phys. Act. 2021, 18, 153. [Google Scholar] [CrossRef] [PubMed]
- Francis, H.M.; Stevenson, R.J.; Chambers, J.R.; Gupta, D.; Newey, B.; Lim, C.K. A brief diet intervention can reduce symptoms of depression in young adults—A randomised controlled trial. PLoS ONE 2019, 14, e0222768. [Google Scholar] [CrossRef] [PubMed]
- Corb Aron, R.A.; Abid, A.; Vesa, C.M.; Nechifor, A.C.; Behl, T.; Ghitea, T.C.; Munteanu, M.A.; Fratila, O.; Andronie-Cioara, F.L.; Toma, M.M.; et al. Recognizing the Benefits of Pre-/Probiotics in Metabolic Syndrome and Type 2 Diabetes Mellitus Considering the Influence of Akkermansia muciniphila as a Key Gut Bacterium. Microorganisms 2021, 9, 618. [Google Scholar] [CrossRef]
- Sharma, R.; Gupta, D.; Mehrotra, R.; Mago, P. Psychobiotics: The Next-Generation Probiotics for the Brain. Curr. Microbiol. 2021, 78, 449–463. [Google Scholar] [CrossRef]
- Tindall, L.; Mikocka-Walus, A.; McMillan, D.; Wright, B.; Hewitt, C.; Gascoyne, S. Is behavioural activation effective in the treatment of depression in young people? A systematic review and meta-analysis. Psychol. Psychother. 2017, 90, 770–796. [Google Scholar] [CrossRef] [Green Version]
- Beam, A.; Clinger, E.; Hao, L. Effect of Diet and Dietary Components on the Composition of the Gut Microbiota. Nutrients 2021, 13, 2795. [Google Scholar] [CrossRef]
- Dimidi, E.; Cox, S.R.; Rossi, M.; Whelan, K. Fermented Foods: Definitions and Characteristics, Impact on the Gut Microbiota and Effects on Gastrointestinal Health and Disease. Nutrients 2019, 11, 1806. [Google Scholar] [CrossRef] [Green Version]
- So, D.; Whelan, K.; Rossi, M.; Morrison, M.; Holtmann, G.; Kelly, J.T.; Shanahan, E.R.; Staudacher, H.M.; Campbell, K.L. Dietary fiber intervention on gut microbiota composition in healthy adults: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2018, 107, 965–983. [Google Scholar] [CrossRef] [Green Version]
- Barber, T.M.; Valsamakis, G.; Mastorakos, G.; Hanson, P.; Kyrou, I.; Randeva, H.S.; Weickert, M.O. Dietary Influences on the Microbiota-Gut-Brain Axis. Int. J. Mol. Sci. 2021, 22, 3502. [Google Scholar] [CrossRef] [PubMed]
- Schuch, F.B.; Deslandes, A.C.; Stubbs, B.; Gosmann, N.P.; Silva, C.T.; Fleck, M.P. Neurobiological effects of exercise on major depressive disorder: A systematic review. Neurosci. Biobehav. Rev. 2016, 61, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Stranahan, A.M.; Lee, K.; Mattson, M.P. Central mechanisms of HPA axis regulation by voluntary exercise. Neuromol. Med. 2008, 10, 118–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, I.; Leblanc, A.G. Systematic review of the health benefits of physical activity and fitness in school-aged children and youth. Int. J. Behav. Nutr. Phys. Act. 2010, 7, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, R.V.; Murthy, V.L.; Colangelo, L.A.; Reis, J.; Venkatesh, B.A.; Sharma, R.; Abbasi, S.A.; Goff, D.C., Jr.; Carr, J.J.; Rana, J.S.; et al. Association of Fitness in Young Adulthood With Survival and Cardiovascular Risk: The Coronary Artery Risk Development in Young Adults (CARDIA) Study. JAMA Intern. Med. 2016, 176, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Inoue, R.; Tsukahara, T.; Ushida, K.; Chiji, H.; Matsubara, N.; Hara, H. Voluntary running exercise alters microbiota composition and increases n-butyrate concentration in the rat cecum. Biosci. Biotechnol. Biochem. 2008, 72, 572–576. [Google Scholar] [CrossRef]
- Lensu, S.; Pekkala, S. Gut Microbiota, Microbial Metabolites and Human Physical Performance. Metabolites 2021, 11, 716. [Google Scholar] [CrossRef]
- Scheiman, J.; Luber, J.M.; Chavkin, T.A.; MacDonald, T.; Tung, A.; Pham, L.D.; Wibowo, M.C.; Wurth, R.C.; Punthambaker, S.; Tierney, B.T.; et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nat. Med. 2019, 25, 1104–1109. [Google Scholar] [CrossRef]
- Clark, A.; Mach, N. Exercise-induced stress behavior, gut-microbiota-brain axis and diet: A systematic review for athletes. J. Int. Soc. Sports Nutr. 2016, 13, 43. [Google Scholar] [CrossRef] [Green Version]
- Bressa, C.; Bailen-Andrino, M.; Perez-Santiago, J.; Gonzalez-Soltero, R.; Perez, M.; Montalvo-Lominchar, M.G.; Mate-Munoz, J.L.; Dominguez, R.; Moreno, D.; Larrosa, M. Differences in gut microbiota profile between women with active lifestyle and sedentary women. PLoS ONE 2017, 12, e0171352. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.M.; Mailing, L.J.; Niemiro, G.M.; Moore, R.; Cook, M.D.; White, B.A.; Holscher, H.D.; Woods, J.A. Exercise Alters Gut Microbiota Composition and Function in Lean and Obese Humans. Med. Sci. Sports Exerc. 2018, 50, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Kim, D.; Kang, H. Exercise Preconditioning Attenuates the Response to Experimental Colitis and Modifies Composition of Gut Microbiota in Wild-Type Mice. Life 2020, 10, 200. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.; Fernandez-Sanjurjo, M.; Iglesias-Gutierrez, E.; Martinez-Camblor, P.; Villar, C.J.; Tomas-Zapico, C.; Fernandez-Garcia, B.; Lombo, F. Resistance and Endurance Exercise Training Induce Differential Changes in Gut Microbiota Composition in Murine Models. Front. Physiol. 2021, 12, 748854. [Google Scholar] [CrossRef] [PubMed]
- Morishima, S.; Aoi, W.; Kawamura, A.; Kawase, T.; Takagi, T.; Naito, Y.; Tsukahara, T.; Inoue, R. Intensive, prolonged exercise seemingly causes gut dysbiosis in female endurance runners. J. Clin. Biochem. Nutr. 2021, 68, 253–258. [Google Scholar] [CrossRef]
- Mika, A.; Van Treuren, W.; Gonzalez, A.; Herrera, J.J.; Knight, R.; Fleshner, M. Exercise is More Effective at Altering Gut Microbial Composition and Producing Stable Changes in Lean Mass in Juvenile versus Adult Male F344 Rats. PLoS ONE 2015, 10, e0125889. [Google Scholar] [CrossRef]
- Wang, R.; Cai, Y.; Li, J.; Yau, S.Y.; Lu, W.; Stubbs, B.; Su, K.P.; Xu, G.; So, K.F.; Lin, K.; et al. Effects of aerobic exercise on gut microbiota in adolescents with subthreshold mood syndromes and healthy adolescents: A 12-week, randomized controlled trial. J. Affect. Disord. 2021, 293, 363–372. [Google Scholar] [CrossRef]
- Ticinesi, A.; Lauretani, F.; Tana, C.; Nouvenne, A.; Ridolo, E.; Meschi, T. Exercise and immune system as modulators of intestinal microbiome: Implications for the gut-muscle axis hypothesis. Exerc. Immunol. Rev. 2019, 25, 84–95. [Google Scholar]
- Otte, J.A.; Oostveen, E.; Geelkerken, R.H.; Groeneveld, A.B.; Kolkman, J.J. Exercise induces gastric ischemia in healthy volunteers: A tonometry study. J. Appl. Physiol. (1985) 2001, 91, 866–871. [Google Scholar] [CrossRef]
- van Wijck, K.; Lenaerts, K.; van Loon, L.J.; Peters, W.H.; Buurman, W.A.; Dejong, C.H. Exercise-induced splanchnic hypoperfusion results in gut dysfunction in healthy men. PLoS ONE 2011, 6, e22366. [Google Scholar] [CrossRef] [Green Version]
- Mailing, L.J.; Allen, J.M.; Buford, T.W.; Fields, C.J.; Woods, J.A. Exercise and the Gut Microbiome: A Review of the Evidence, Potential Mechanisms, and Implications for Human Health. Exerc. Sport Sci. Rev. 2019, 47, 75–85. [Google Scholar] [CrossRef]
- Leyton, M.; Stewart, S. (Eds.) Substance Abuse in Canada: Childhood and Adoelscent Pathwaya to Substance Use Disorders; Canadian Centre on Substance Abuse: Ottawa, ON, Canada, 2014.
- World Health Organization. Adolescent and Young Adult Health. Available online: https://www.who.int/news-room/fact-sheets/detail/adolescents-health-risks-and-solutions (accessed on 2 March 2022).
- SAMHSA. Mental Illness and Substance Use in Young Adults. Available online: https://www.samhsa.gov/young-adults (accessed on 6 April 2022).
- Centre for Disease Control and Prevention (CDC). Youth and Tobacco Use. Available online: https://www.cdc.gov/tobacco/data_statistics/fact_sheets/youth_data/tobacco_use/index.htm (accessed on 6 April 2022).
- Gentzke, A.S.; Wang, T.W.; Cornelius, M.; Park-Lee, E.; Ren, C.; Sawdey, M.D.; Cullen, K.A.; Loretan, C.; Jamal, A.; Homa, D.M. Tobacco Product Use among Middle and High School Students-United States, 2021; U.S. Department of Health and Human Services: Washington, DC, USA, 2022; pp. 1811–1888.
- Eurobarometer, S. Attitudes of Europeans towards Tobacco and Electronic Cigarettes; European Union: Brussels, Belgium, 2020. [Google Scholar]
- Ehlinger, D.G.; Bergstrom, H.C.; Burke, J.C.; Fernandez, G.M.; McDonald, C.G.; Smith, R.F. Adolescent nicotine-induced dendrite remodeling in the nucleus accumbens is rapid, persistent, and D1-dopamine receptor dependent. Brain Struct. Funct. 2016, 221, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.E.; Lust, K.; Fridberg, D.J.; King, A.C.; Chamberlain, S.R. E-cigarette use (vaping) is associated with illicit drug use, mental health problems, and impulsivity in university students. Ann. Clin. Psychiatry 2019, 31, 27–35. [Google Scholar] [PubMed]
- Ruszkiewicz, J.A.; Zhang, Z.; Goncalves, F.M.; Tizabi, Y.; Zelikoff, J.T.; Aschner, M. Neurotoxicity of e-cigarettes. Food Chem. Toxicol. 2020, 138, 111245. [Google Scholar] [CrossRef]
- Gui, X.; Yang, Z.; Li, M.D. Effect of Cigarette Smoke on Gut Microbiota: State of Knowledge. Front. Physiol. 2021, 12, 673341. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.; McLellan, R.; Wellmeyer, E.; Matalon, F.; George, O. Drugs and Bugs: The Gut-Brain Axis and Substance Use Disorders. J. Neuroimmune Pharmacol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Tomoda, K.; Kubo, K.; Asahara, T.; Andoh, A.; Nomoto, K.; Nishii, Y.; Yamamoto, Y.; Yoshikawa, M.; Kimura, H. Cigarette smoke decreases organic acids levels and population of bifidobacterium in the caecum of rats. J. Toxicol. Sci. 2011, 36, 261–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanahan, E.R.; Shah, A.; Koloski, N.; Walker, M.M.; Talley, N.J.; Morrison, M.; Holtmann, G.J. Influence of cigarette smoking on the human duodenal mucosa-associated microbiota. Microbiome 2018, 6, 150. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Underage Drinking. Available online: https://www.cdc.gov/alcohol/fact-sheets/underage-drinking.htm (accessed on 2 March 2022).
- Inchley, J.C.D.; Budisavljevic, S.; Torsheim, T.; Jåstad, A.; Cosma, A.; Kelly, C.; Arnarsson, Á.M. Spotlight on Adolescent Health and Well-Being; World Health Organization: Copenhagen, Denmark, 2020. [Google Scholar]
- Government of Canada. Alcohol use among Canadians. Available online: https://health-infobase.canada.ca/alcohol/ctads/#shr-pg0 (accessed on 2 March 2022).
- Pilowsky, D.J.; Keyes, K.M.; Hasin, D.S. Adverse childhood events and lifetime alcohol dependence. Am. J. Public Health 2009, 99, 258–263. [Google Scholar] [CrossRef]
- Ports, K.A.; Holman, D.M.; Guinn, A.S.; Pampati, S.; Dyer, K.E.; Merrick, M.T.; Lunsford, N.B.; Metzler, M. Adverse Childhood Experiences and the Presence of Cancer Risk Factors in Adulthood: A Scoping Review of the Literature From 2005 to 2015. J. Pediatr. Nurs. 2019, 44, 81–96. [Google Scholar] [CrossRef]
- Kirsch, D.E.; Lippard, E.T.C. Early life stress and substance use disorders: The critical role of adolescent substance use. Pharmacol. Biochem. Behav. 2022, 215, 173360. [Google Scholar] [CrossRef]
- Lees, B.; Meredith, L.R.; Kirkland, A.E.; Bryant, B.E.; Squeglia, L.M. Effect of alcohol use on the adolescent brain and behavior. Pharmacol. Biochem. Behav. 2020, 192, 172906. [Google Scholar] [CrossRef] [PubMed]
- Government of Canada. Canadian Cannabis Survey 2021: Summary. Available online: https://www.canada.ca/en/health-canada/services/drugs-medication/cannabis/research-data/canadian-cannabis-survey-2021-summary.html (accessed on 2 March 2022).
- European Monitoring Centre for Drugs and Drug Addiction. European Drug Report Trends and Developments 2021; Publications Office of the European Union: Luxembourg, 2021. [Google Scholar]
- Jones, C.M.; Clayton, H.B.; Deputy, N.P.; Roehler, D.R.; Ko, J.Y.; Esser, M.B.; Brookmeyer, K.A.; Hertz, M.F. Prescription Opioid Misuse and Use of Alcohol and Other Substances Among High School Students—Youth Risk Behavior Survey, United States, 2019. MMWR Suppl. 2020, 69, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Medina, K.L.; McQueeny, T.; Nagel, B.J.; Hanson, K.L.; Schweinsburg, A.D.; Tapert, S.F. Prefrontal cortex volumes in adolescents with alcohol use disorders: Unique gender effects. Alcohol. Clin. Exp. Res. 2008, 32, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Jacobus, J.; Squeglia, L.M.; Meruelo, A.D.; Castro, N.; Brumback, T.; Giedd, J.N.; Tapert, S.F. Cortical thickness in adolescent marijuana and alcohol users: A three-year prospective study from adolescence to young adulthood. Dev. Cogn. Neurosci. 2015, 16, 101–109. [Google Scholar] [CrossRef] [Green Version]
- DiPatrizio, N.V. Endocannabinoids in the Gut. Cannabis Cannabinoid Res. 2016, 1, 67–77. [Google Scholar] [CrossRef]
- Muccioli, G.G.; Naslain, D.; Backhed, F.; Reigstad, C.S.; Lambert, D.M.; Delzenne, N.M.; Cani, P.D. The endocannabinoid system links gut microbiota to adipogenesis. Mol. Syst. Biol. 2010, 6, 392. [Google Scholar] [CrossRef]
- Panee, J.; Gerschenson, M.; Chang, L. Associations Between Microbiota, Mitochondrial Function, and Cognition in Chronic Marijuana Users. J. Neuroimmune Pharmacol. 2018, 13, 113–122. [Google Scholar] [CrossRef]
- National Institute of General Medical Sciences. Circadian Rhythms. National Institues of Health; 2020. Available online: https://www.nigms.nih.gov/education/fact-sheets/Pages/circadian-rhythms.aspx (accessed on 2 March 2022).
- Teichman, E.M.; O’Riordan, K.J.; Gahan, C.G.M.; Dinan, T.G.; Cryan, J.F. When Rhythms Meet the Blues: Circadian Interactions with the Microbiota-Gut-Brain Axis. Cell Metab. 2020, 31, 448–471. [Google Scholar] [CrossRef]
- Walker, W.H., 2nd; Walton, J.C.; DeVries, A.C.; Nelson, R.J. Circadian rhythm disruption and mental health. Transl. Psychiatry 2020, 10, 28. [Google Scholar] [CrossRef] [Green Version]
- Galima, S.V.; Vogel, S.R.; Kowalski, A.W. Seasonal Affective Disorder: Common Questions and Answers. Am. Fam. Physician 2020, 102, 668–672. [Google Scholar]
- Crowley, S.J.; Acebo, C.; Carskadon, M.A. Sleep, circadian rhythms, and delayed phase in adolescence. Sleep Med. 2007, 8, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Pesonen, A.K.; Martikainen, S.; Kajantie, E.; Heinonen, K.; Wehkalampi, K.; Lahti, J.; Strandberg, T.; Raikkonen, K. The associations between adolescent sleep, diurnal cortisol patterns and cortisol reactivity to dexamethasone suppression test. Psychoneuroendocrinology 2014, 49, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Hagenauer, M.H.; Perryman, J.I.; Lee, T.M.; Carskadon, M.A. Adolescent changes in the homeostatic and circadian regulation of sleep. Dev. Neurosci. 2009, 31, 276–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalsbeek, A.; van der Spek, R.; Lei, J.; Endert, E.; Buijs, R.M.; Fliers, E. Circadian rhythms in the hypothalamo-pituitary-adrenal (HPA) axis. Mol. Cell. Endocrinol. 2012, 349, 20–29. [Google Scholar] [CrossRef]
- Wu, G.; Tang, W.; He, Y.; Hu, J.; Gong, S.; He, Z.; Wei, G.; Lv, L.; Jiang, Y.; Zhou, H.; et al. Light exposure influences the diurnal oscillation of gut microbiota in mice. Biochem. Biophys. Res. Commun. 2018, 501, 16–23. [Google Scholar] [CrossRef]
- Bowers, S.J.; Vargas, F.; Gonzalez, A.; He, S.; Jiang, P.; Dorrestein, P.C.; Knight, R.; Wright, K.P., Jr.; Lowry, C.A.; Fleshner, M.; et al. Repeated sleep disruption in mice leads to persistent shifts in the fecal microbiome and metabolome. PLoS ONE 2020, 15, e0229001. [Google Scholar] [CrossRef] [Green Version]
- Kinnucan, J.A.; Rubin, D.T.; Ali, T. Sleep and inflammatory bowel disease: Exploring the relationship between sleep disturbances and inflammation. Gastroenterol. Hepatol. 2013, 9, 718–727. [Google Scholar]
- Wagner-Skacel, J.; Dalkner, N.; Moerkl, S.; Kreuzer, K.; Farzi, A.; Lackner, S.; Painold, A.; Reininghaus, E.Z.; Butler, M.I.; Bengesser, S. Sleep and Microbiome in Psychiatric Diseases. Nutrients 2020, 12, 2198. [Google Scholar] [CrossRef]
- Berendsen, M.; Boss, M.; Smits, M.; Pot, G.K. Chrono-Nutrition and Diet Quality in Adolescents with Delayed Sleep-Wake Phase Disorder. Nutrients 2020, 12, 539. [Google Scholar] [CrossRef] [Green Version]
- Leeuwendaal, N.K.; Stanton, C.; O’Toole, P.W.; Beresford, T.P. Fermented Foods, Health and the Gut Microbiome. Nutrients 2022, 14, 1527. [Google Scholar] [CrossRef]
- Mahoney, J.J., 3rd; Hanlon, C.A.; Marshalek, P.J.; Rezai, A.R.; Krinke, L. Transcranial magnetic stimulation, deep brain stimulation, and other forms of neuromodulation for substance use disorders: Review of modalities and implications for treatment. J. Neurol. Sci. 2020, 418, 117149. [Google Scholar] [CrossRef] [PubMed]
- Breeksema, J.J.; Koolen, M.H.B.; Somers, M.; Schoevers, R.A. Treatment with psilocybin: Applications for patients with psychiatric disorders. Ned. Tijdschr Geneeskd. 2021, 165, D4842. [Google Scholar] [PubMed]
- Cosgrove, L.; Morrill, Z.; Yusif, M.; Vaswani, A.; Cathcart, S.; Troeger, R.; Karter, J.M. Drivers of and Solutions for the Overuse of Antidepressant Medication in Pediatric Populations. Front. Psychiatry 2020, 11, 17. [Google Scholar] [CrossRef] [PubMed]
- Lampela, P.; Tanskanen, A.; Lahteenvuo, M.; Tiihonen, J.; Taipale, H. Switches and early discontinuations of antidepressant medication in young adults with depression. J. Affect. Disord. 2021, 295, 1474–1481. [Google Scholar] [CrossRef]
- Alosaimi, F.D.; Abalhassan, M.; Alhaddad, B.; Alzain, N.; Fallata, E.; Alhabbad, A.; Alassiry, M.Z. Prevalence of metabolic syndrome and its components among patients with various psychiatric diagnoses and treatments: A cross-sectional study. Gen. Hosp. Psychiatry 2017, 45, 62–69. [Google Scholar] [CrossRef]
- Li, C.; Birmaher, B.; Rooks, B.; Gill, M.K.; Hower, H.; Axelson, D.A.; Dickstein, D.P.; Goldstein, T.R.; Liao, F.; Yen, S.; et al. High Prevalence of Metabolic Syndrome Among Adolescents and Young Adults With Bipolar Disorder. J. Clin. Psychiatry 2019, 80, 11793. [Google Scholar] [CrossRef]
- Crookes, D.M.; Demmer, R.T.; Keyes, K.M.; Koenen, K.C.; Suglia, S.F. Depressive Symptoms, Antidepressant Use, and Hypertension in Young Adulthood. Epidemiology 2018, 29, 547–555. [Google Scholar] [CrossRef]
- Macedo, D.; Filho, A.; Soares de Sousa, C.N.; Quevedo, J.; Barichello, T.; Junior, H.V.N.; Freitas de Lucena, D. Antidepressants, antimicrobials or both? Gut microbiota dysbiosis in depression and possible implications of the antimicrobial effects of antidepressant drugs for antidepressant effectiveness. J. Affect. Disord. 2017, 208, 22–32. [Google Scholar] [CrossRef]
- Cussotto, S.; Clarke, G.; Dinan, T.G.; Cryan, J.F. Psychotropics and the Microbiome: A Chamber of Secrets. Psychopharmacology 2019, 236, 1411–1432. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.E.; Walton, D.; O’Connor, C.P.; Wammes, M.; Burton, J.P.; Osuch, E.A. Drugs, Guts, Brains, but Not Rock and Roll: The Need to Consider the Role of Gut Microbiota in Contemporary Mental Health and Wellness of Emerging Adults. Int. J. Mol. Sci. 2022, 23, 6643. https://doi.org/10.3390/ijms23126643
Lee JE, Walton D, O’Connor CP, Wammes M, Burton JP, Osuch EA. Drugs, Guts, Brains, but Not Rock and Roll: The Need to Consider the Role of Gut Microbiota in Contemporary Mental Health and Wellness of Emerging Adults. International Journal of Molecular Sciences. 2022; 23(12):6643. https://doi.org/10.3390/ijms23126643
Chicago/Turabian StyleLee, Ju Eun, David Walton, Colleen P. O’Connor, Michael Wammes, Jeremy P. Burton, and Elizabeth A. Osuch. 2022. "Drugs, Guts, Brains, but Not Rock and Roll: The Need to Consider the Role of Gut Microbiota in Contemporary Mental Health and Wellness of Emerging Adults" International Journal of Molecular Sciences 23, no. 12: 6643. https://doi.org/10.3390/ijms23126643
APA StyleLee, J. E., Walton, D., O’Connor, C. P., Wammes, M., Burton, J. P., & Osuch, E. A. (2022). Drugs, Guts, Brains, but Not Rock and Roll: The Need to Consider the Role of Gut Microbiota in Contemporary Mental Health and Wellness of Emerging Adults. International Journal of Molecular Sciences, 23(12), 6643. https://doi.org/10.3390/ijms23126643