
Peripheral Blood T Cells of Patients with IPAH Have a Reduced Cytokine-Producing Capacity

 ,
, 

Abstract
:1. Introduction
2. Results
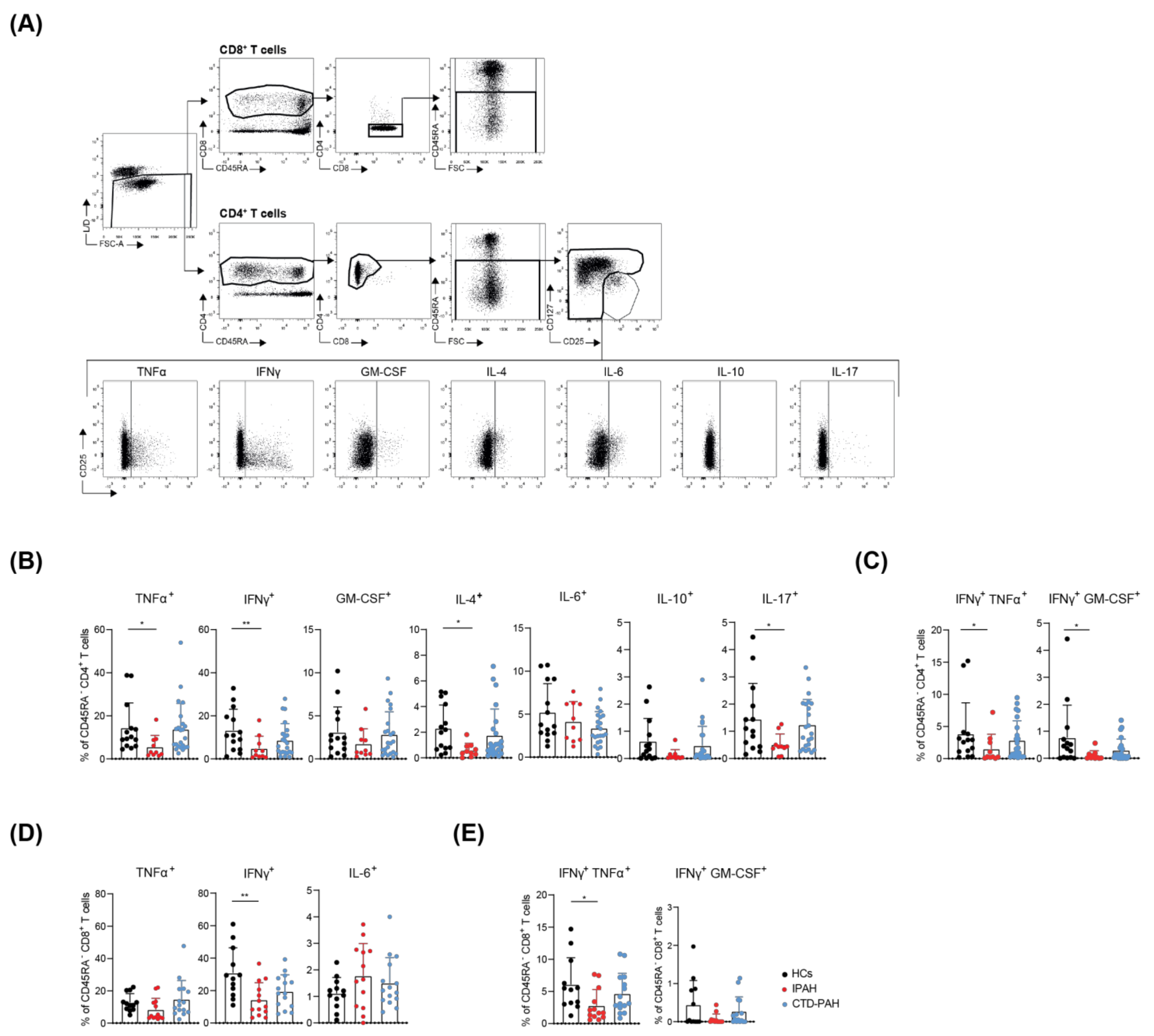
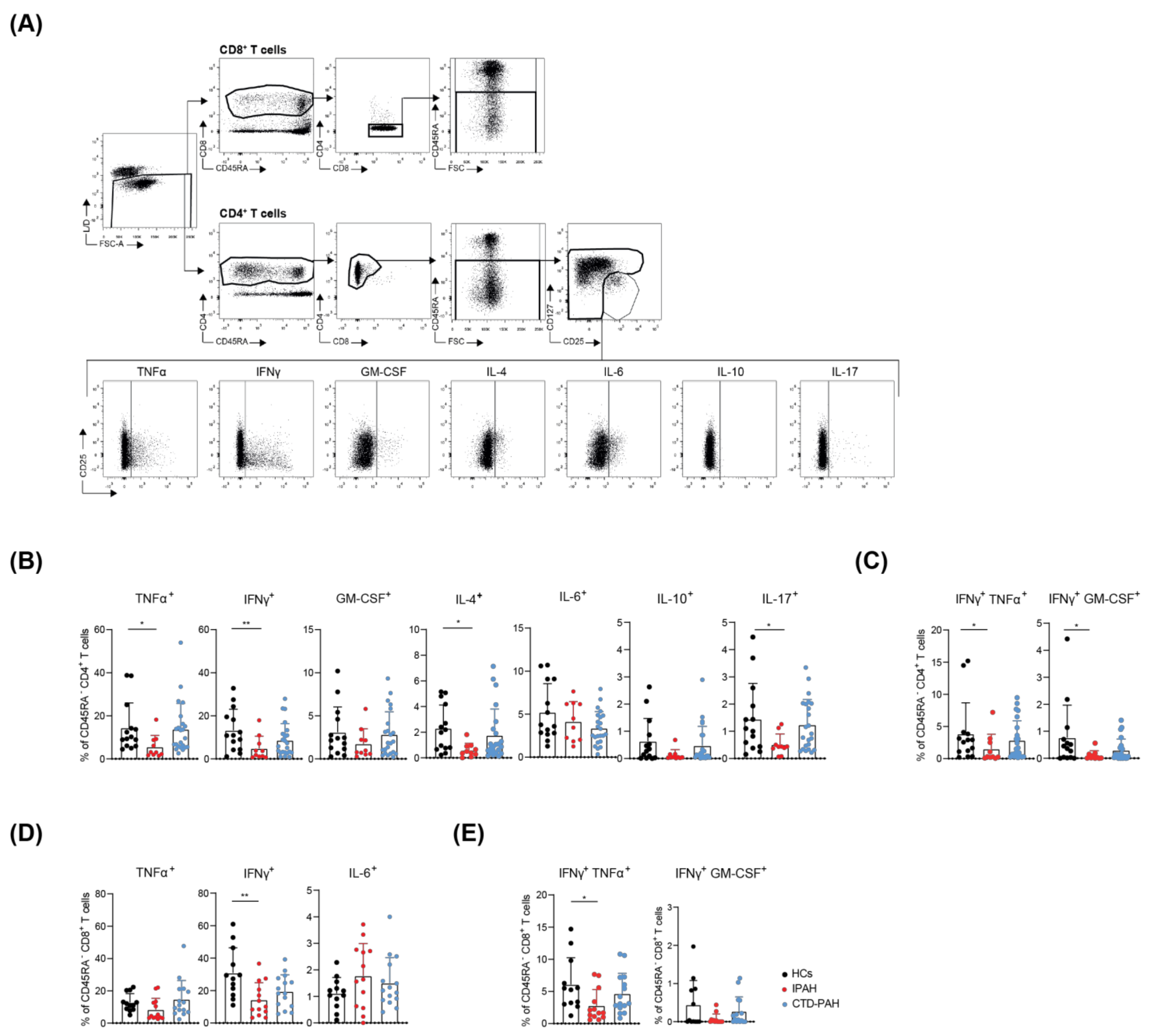
2.1. T Cells from Treatment-Naive IPAH Patients Show Reduced Cytokine-Producing Capacity
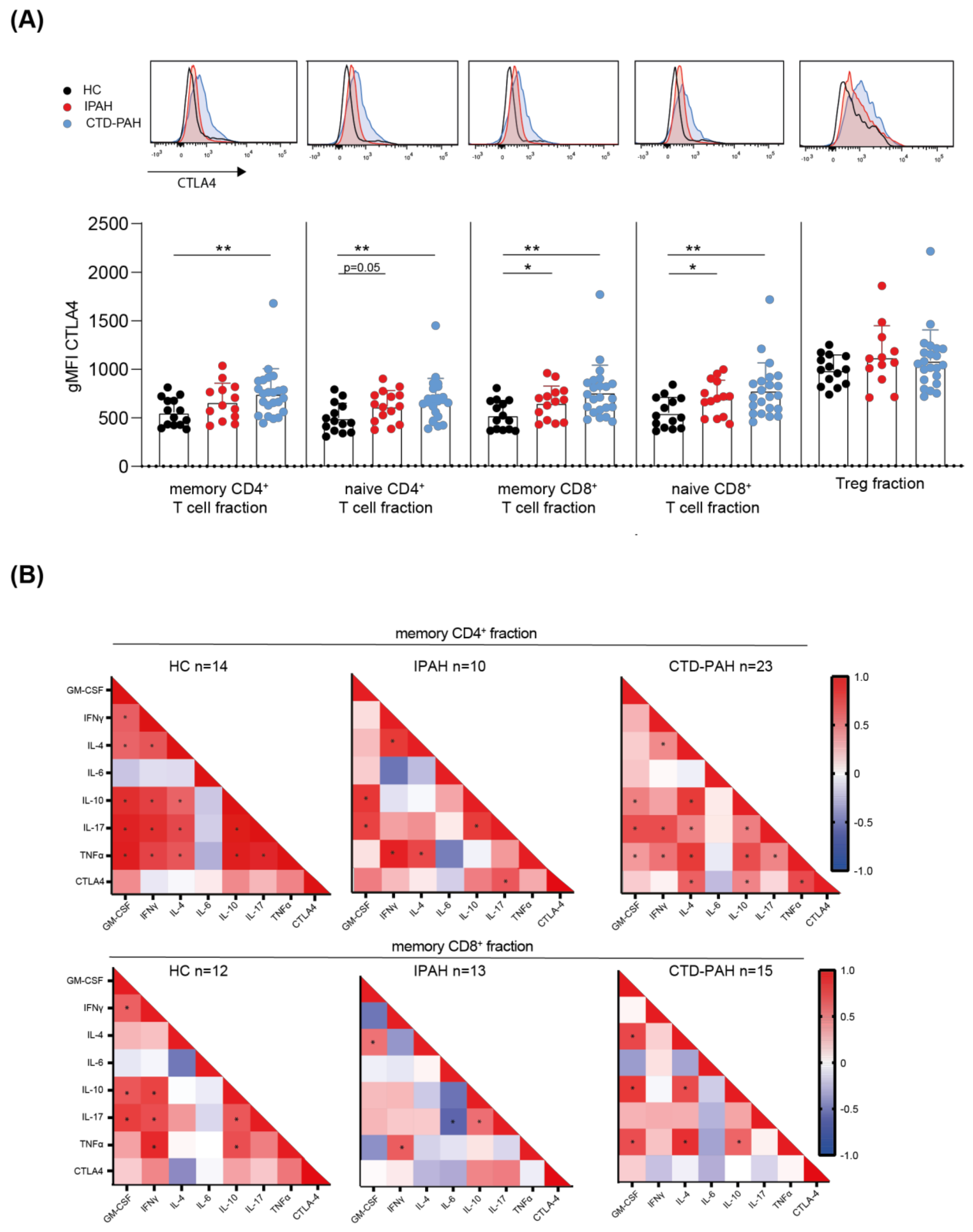
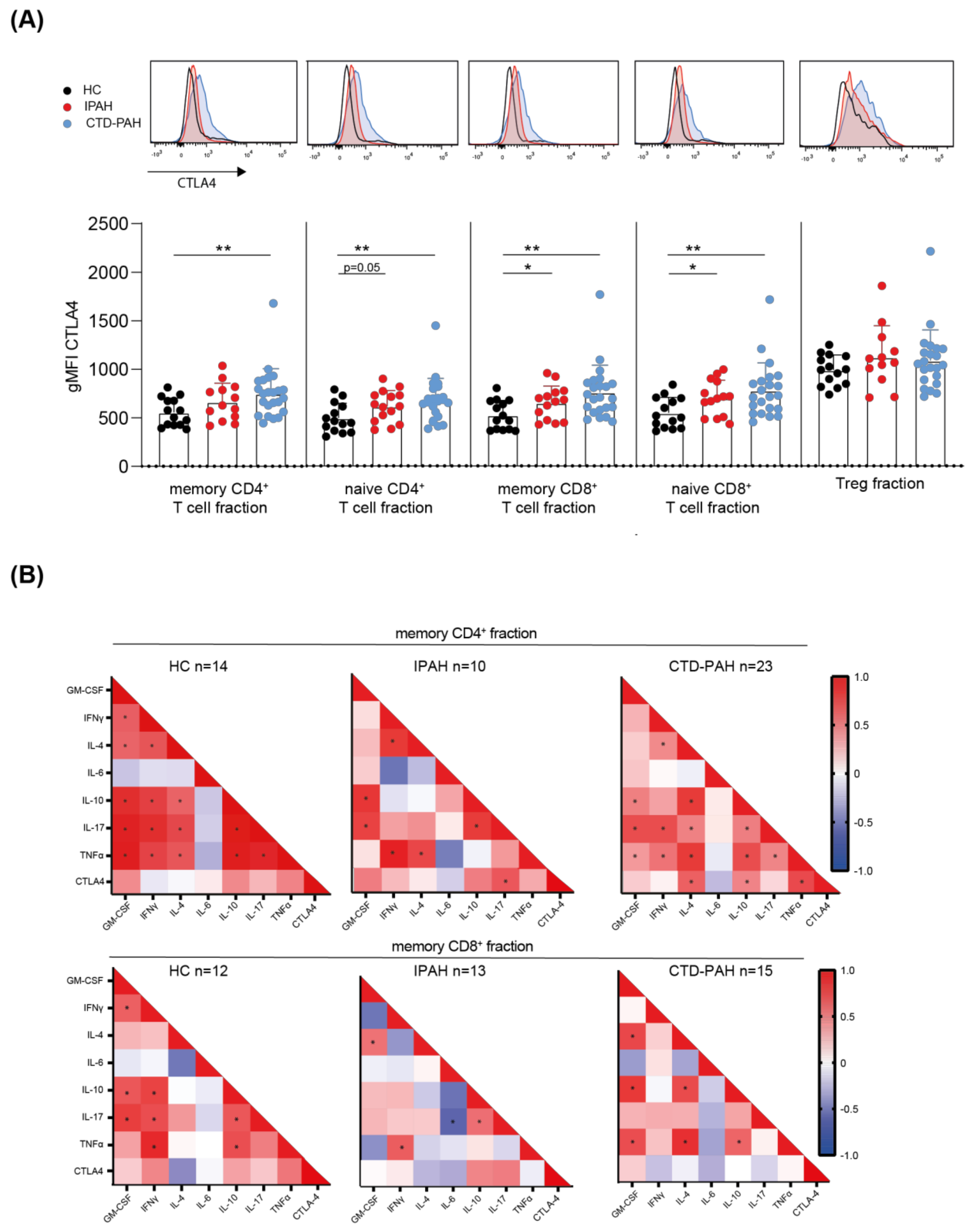
2.2. T Cells from PAH Patients Show Increased CTLA4 Expression, Correlating with Cytokine-Producing Capacity
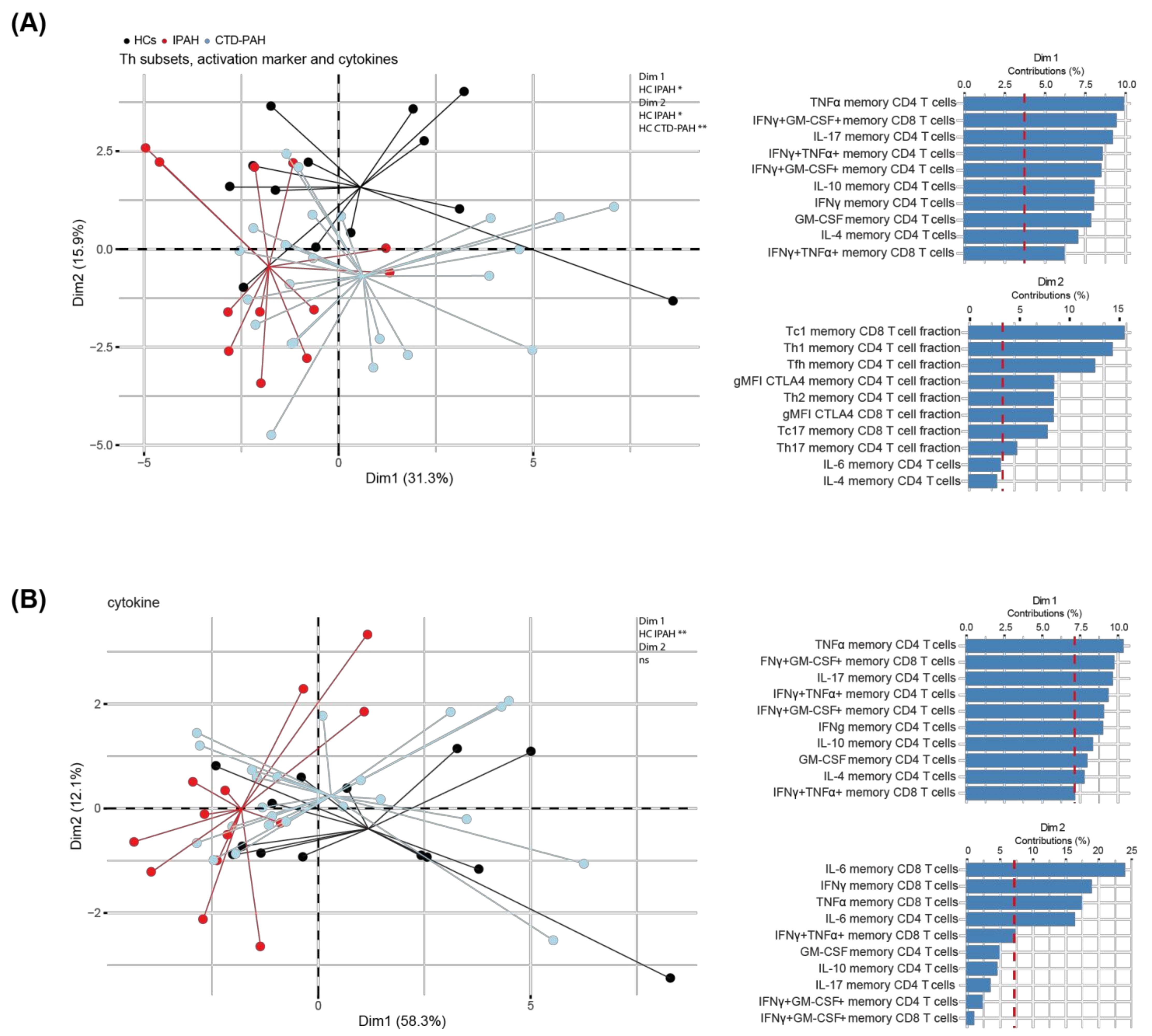
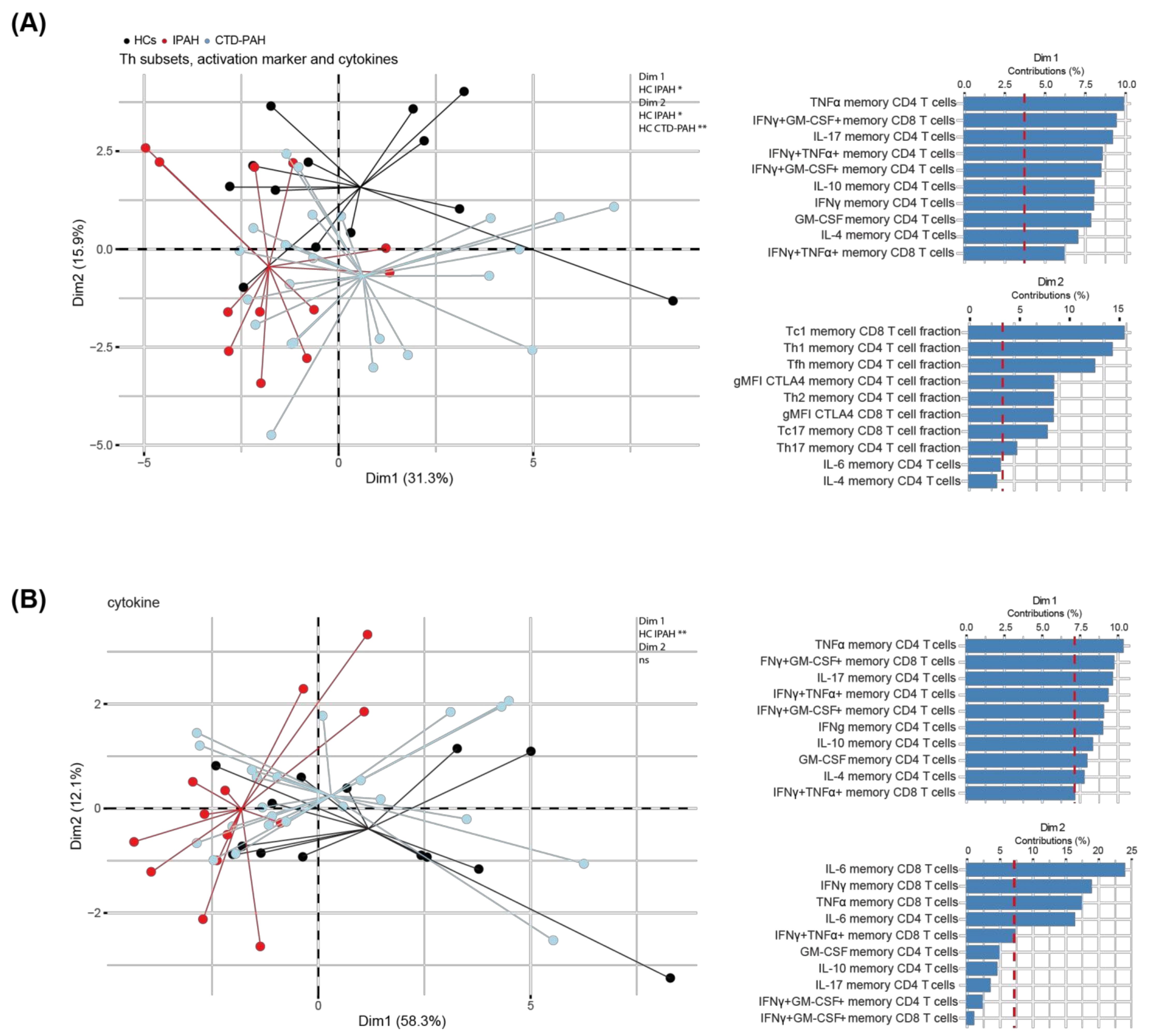
2.3. Principal Component Analysis of T Cell Cytokine Production Separates IPAH Patients from HCs
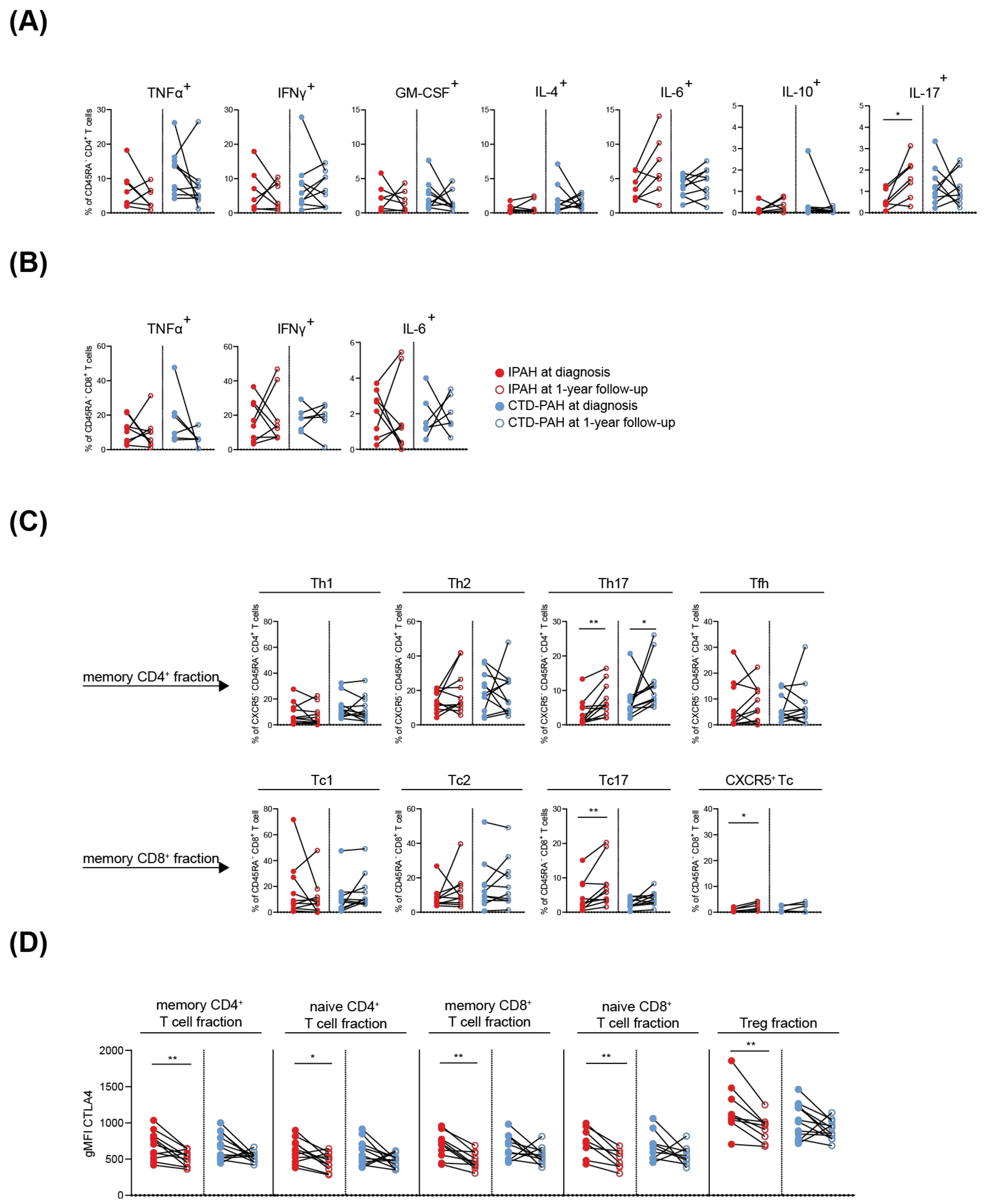
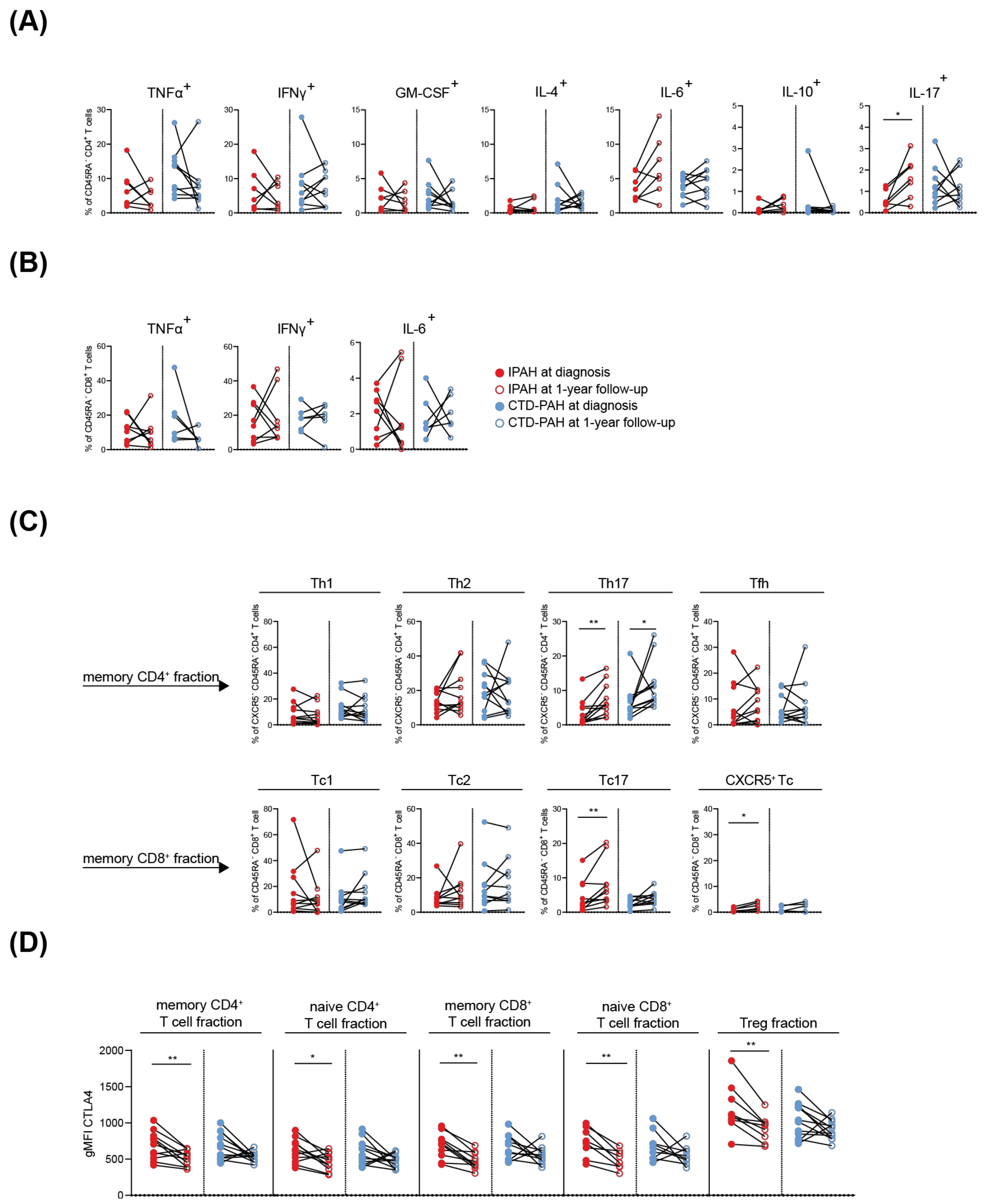
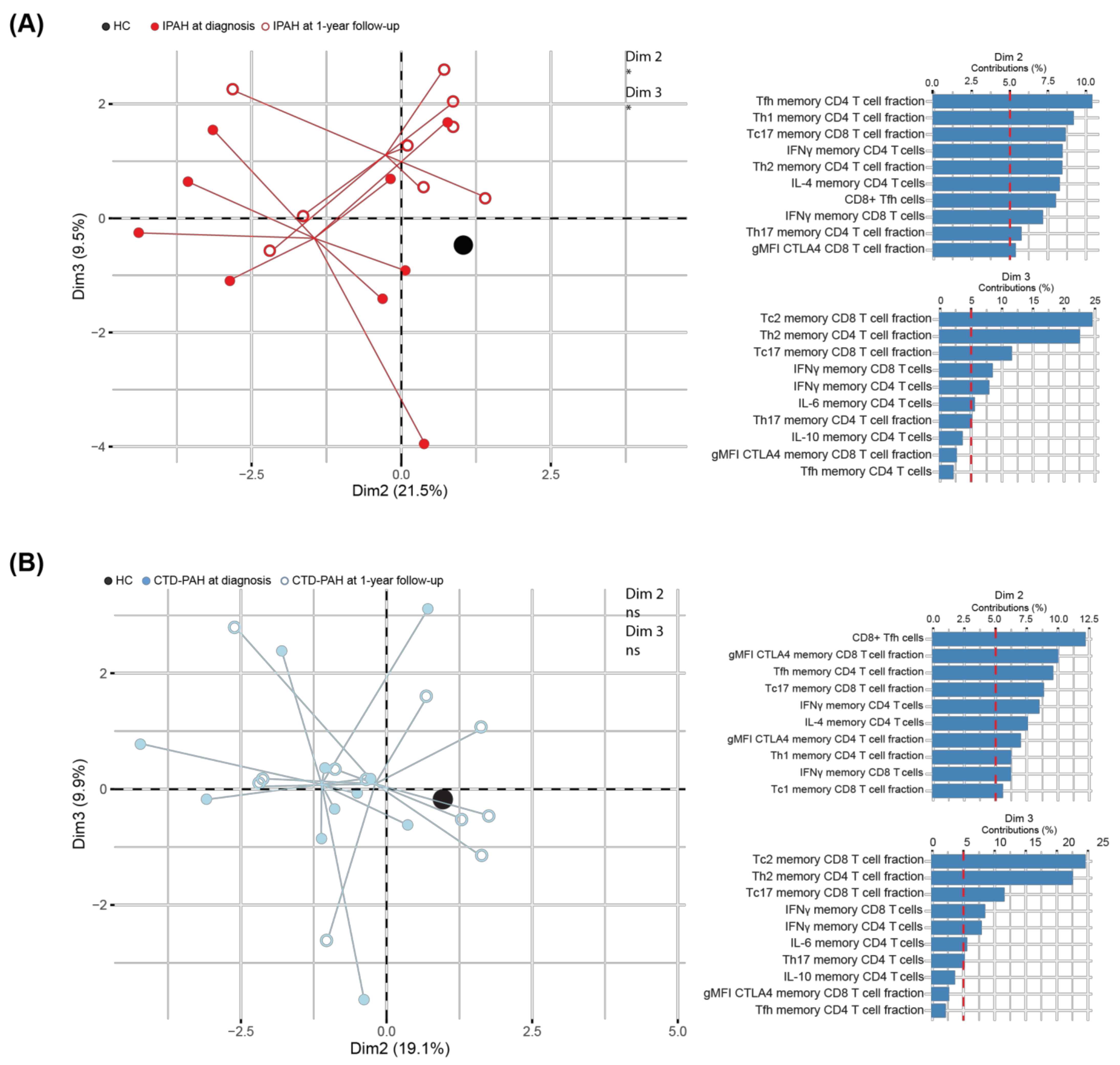
2.4. T Cell Cytokine and CTLA4 Expression Profiles in IPAH Patients Significantly Change over Time
3. Discussion
4. Materials and Methods
4.1. Subjects and Study Design
4.2. Clinical Data Collection, Follow-Up and Definition of Endpoints
4.3. Flow Cytometry
4.4. Principal Component Analysis and Statistical Evaluation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simonneau, G.; Montani, D.; Celermajer, D.; Denton, C.P.; Gatzoulis, M.A.; Krowka, M.; Williams, P.G.; Souza, R. Haemodynamic definitions and updated clinical classification of pulmonary hypertension. Eur. Respir. J. 2019, 53, 1801913. [Google Scholar] [CrossRef] [PubMed]
- Rabinovitch, M.; Guignabert, C.; Humbert, M.; Nicolls, M.R. Inflammation and Immunity in the Pathogenesis of Pulmonary Arterial Hypertension. Circ. Res. 2014, 115, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Koudstaal, T.; Boomars, K.A.; Kool, M. Pulmonary Arterial Hypertension and Chronic Thromboembolic Pulmonary Hypertension: An Immunological Perspective. J. Clin. Med. 2020, 9, 561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsh, L.M.; Jandl, K.; Grünig, G.; Foris, V.; Bashir, M.; Ghanim, B.; Klepetko, W.; Olschewski, H.; Olschewski, A.; Kwapiszewska, G. The inflammatory cell landscape in the lungs of patients with idiopathic pulmonary arterial hypertension. Eur. Respir. J. 2018, 51, 1701214. [Google Scholar] [CrossRef] [Green Version]
- Savai, R.; Pullamsetti, S.S.; Kolbe, J.; Bieniek, E.; Voswinckel, R.; Fink, L.; Scheed, A.; Ritter, C.; Dahal, B.K.; Vater, A.; et al. Immune and Inflammatory Cell Involvement in the Pathology of Idiopathic Pulmonary Arterial Hypertension. Am. J. Respir. Crit. Care Med. 2012, 186, 897–908. [Google Scholar] [CrossRef]
- Mauad, T.; Pozzan, G.; Lanças, T.; Overbeek, M.J.; Souza, R.; Jardim, C.; Dolhnikoff, M.; Mello, G.; Pires-Neto, R.C.; Bernardi, F.D.C.; et al. Immunopathological aspects of schistosomiasis-associated pulmonary arterial hypertension. J. Infect. 2014, 68, 90–98. [Google Scholar] [CrossRef]
- Gaffen, S.L.; Jain, R.; Garg, A.V.; Cua, D.J. The IL-23–IL-17 immune axis: From mechanisms to therapeutic testing. Nat. Rev. Immunol. 2014, 14, 585–600. [Google Scholar] [CrossRef]
- Stadhouders, R.; Lubberts, E.; Hendriks, R.W. A cellular and molecular view of T helper 17 cell plasticity in autoimmunity. J. Autoimmun. 2018, 87, 1–15. [Google Scholar] [CrossRef]
- Perros, F.; Dorfmüller, P.; Montani, D.; Hammad, H.; Waelput, W.; Girerd, B.; Raymond, N.; Mercier, O.; Mussot, S.; Cohen-Kaminsky, S.; et al. Pulmonary Lymphoid Neogenesis in Idiopathic Pulmonary Arterial Hypertension. Am. J. Respir. Crit. Care Med. 2012, 185, 311–321. [Google Scholar] [CrossRef]
- Hautefort, A.; Girerd, B.; Montani, D.; Cohen-Kaminsky, S.; Price, L.; Lambrecht, B.N.; Humbert, M.; Perros, F. T-Helper 17 Cell Polarization in Pulmonary Arterial Hypertension. Chest 2015, 147, 1610–1620. [Google Scholar] [CrossRef]
- Van Hamburg, J.P.; Tas, S.W. Molecular mechanisms underpinning T helper 17 cell heterogeneity and functions in rheumatoid arthritis. J. Autoimmun. 2018, 87, 69–81. [Google Scholar] [CrossRef]
- Soon, E.; Holmes, A.M.; Treacy, C.M.; Doughty, N.J.; Southgate, L.; Machado, R.D.; Trembath, R.C.; Jennings, S.; Barker, L.; Nicklin, P.; et al. Elevated Levels of Inflammatory Cytokines Predict Survival in Idiopathic and Familial Pulmonary Arterial Hypertension. Circulation 2010, 122, 920–927. [Google Scholar] [CrossRef] [Green Version]
- Heukels, P.; Corneth, O.B.J.; van Uden, D.; Hulst, J.A.C.V.; Toorn, L.M.V.D.; Bosch, A.E.V.D.; Wijsenbeek, M.S.; Boomars, K.A.; Kool, M.; Hendriks, R.W. Loss of immune homeostasis in patients with idiopathic pulmonary arterial hypertension. Thorax 2021, 76, 1209–1218. [Google Scholar] [CrossRef]
- Tian, W.; Jiang, S.Y.; Jiang, X.; Tamosiuniene, R.; Kim, D.; Guan, T.; Arsalane, S.; Pasupneti, S.; Voelkel, N.F.; Tang, Q.; et al. The Role of Regulatory T Cells in Pulmonary Arterial Hypertension. Front. Immunol. 2021, 12, 684657. [Google Scholar] [CrossRef]
- Austin, E.; Rock, M.; Mosse, C.; Vnencak-Jones, C.; Yoder, S.; Robbins, I.; Loyd, J.; Meyrick, B. T lymphocyte subset abnormalities in the blood and lung in pulmonary arterial hypertension. Respir. Med. 2010, 104, 454–462. [Google Scholar] [CrossRef] [Green Version]
- Huertas, A.; Phan, C.; Bordenave, J.; Tu, L.; Thuillet, R.; Le Hiress, M.; Avouac, J.; Tamura, Y.; Allanore, Y.; Jovan, R.; et al. Regulatory T Cell Dysfunction in Idiopathic, Heritable and Connective Tissue-Associated Pulmonary Arterial Hypertension. Chest 2016, 149, 1482–1493. [Google Scholar] [CrossRef]
- Gaowa, S.; Zhou, W.; Yu, L.; Zhou, X.; Liao, K.; Yang, K.; Lu, Z.; Jiang, H.; Chen, X. Effect of Th17 and Treg Axis Disorder on Outcomes of Pulmonary Arterial Hypertension in Connective Tissue Diseases. Mediat. Inflamm. 2014, 2014, 247372. [Google Scholar] [CrossRef]
- Ulrich, S.; Nicolls, M.R.; Taraseviciene, L.; Speich, R.; Voelkel, N. Increased Regulatory and Decreased CD8+ Cytotoxic T Cells in the Blood of Patients with Idiopathic Pulmonary Arterial Hypertension. Respiration 2008, 75, 272–280. [Google Scholar] [CrossRef] [Green Version]
- Blum, L.K.; Cao, R.R.; Sweatt, A.J.; Bill, M.; Lahey, L.J.; Hsi, A.C.; Lee, C.S.; Kongpachith, S.; Ju, C.-H.; Mao, R.; et al. Circulating plasmablasts are elevated and produce pathogenic anti-endothelial cell autoantibodies in idiopathic pulmonary arterial hypertension. Eur. J. Immunol. 2018, 48, 874–884. [Google Scholar] [CrossRef]
- Rich, S.; Kieras, K.; Hart, K.; Groves, B.M.; Stobo, J.D.; Brundage, B.H. Antinuclear antibodies in primary pulmonary hypertension. J. Am. Coll. Cardiol. 1986, 8, 1307–1311. [Google Scholar] [CrossRef] [Green Version]
- Arends, S.J.; Damoiseaux, J.; Duijvestijn, A.; Debrus-Palmans, L.; Boomars, K.; Broers, B.; Tervaert, J.W.C.; van Paassen, P. Prevalence of anti-endothelial cell antibodies in idiopathic pulmonary arterial hypertension. Eur. Respir. J. 2010, 35, 923–925. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Yan, H.; Zhu, W.; Cui, Y.; Chen, J.; Wang, X.; Li, S.; Zhu, J. Impairment of Monocyte-derived Dendritic Cells in Idiopathic Pulmonary Arterial Hypertension. J. Clin. Immunol. 2009, 29, 705–713. [Google Scholar] [CrossRef]
- Van Uden, D.; Koudstaal, T.; van Hulst, J.; Bergen, I.; Gootjes, C.; Morrell, N.; van Loo, G.; von der Thüsen, J.; Bosch, T.V.D.; Ghigna, M.-R.; et al. Central Role of Dendritic Cells in Pulmonary Arterial Hypertension in Human and Mice. Int. J. Mol. Sci. 2021, 22, 1756. [Google Scholar] [CrossRef]
- Koudstaal, T.; van Hulst, J.A.C.; Das, T.; Neys, S.F.H.; Merkus, D.; Bergen, I.M.; de Raaf, M.A.; Bogaard, H.J.; Boon, L.; van Loo, G.; et al. DNGR1-Cre–mediated Deletion of Tnfaip3/A20 in Conventional Dendritic Cells Induces Pulmonary Hypertension in Mice. Am. J. Respir. Cell Mol. Biol. 2020, 63, 665–680. [Google Scholar] [CrossRef]
- Shu, T.; Xing, Y.; Wang, J. Autoimmunity in Pulmonary Arterial Hypertension: Evidence for Local Immunoglobulin Production. Front. Cardiovasc. Med. 2021, 8, 680109. [Google Scholar] [CrossRef]
- Villani, A.C.; Satija, R.; Reynolds, G.; Sarkizova, S.; Shekhar, K.; Fletcher, J.; Griesbeck, M.; Butler, A.; Zheng, S.; Lazo, S.; et al. Single-cell RNA-seq reveals new types of human blood dendritic cells, monocytes, and progenitors. Science 2017, 356, eaah4573. [Google Scholar] [CrossRef] [Green Version]
- Van Uden, D.; Boomars, K.; Kool, M. Dendritic Cell Subsets and Effector Function in Idiopathic and Connective Tissue Disease-Associated Pulmonary Arterial Hypertension. Front. Immunol. 2019, 10, 11. [Google Scholar] [CrossRef]
- Chen, L.; Flies, D.B. Molecular mechanisms of T cell co-stimulation and co-inhibition. Nat. Rev. Immunol. 2013, 13, 227–242. [Google Scholar] [CrossRef]
- Koudstaal, T.; van Uden, D.; van Hulst, J.A.C.; Heukels, P.; Bergen, I.M.; Geenen, L.W.; Baggen, V.J.M.; Bosch, A.E.V.D.; Toorn, L.M.V.D.; Chandoesing, P.P.; et al. Plasma markers in pulmonary hypertension subgroups correlate with patient survival. Respir. Res. 2021, 22, 137. [Google Scholar] [CrossRef]
- Paats, M.S.; Bergen, I.M.; Hoogsteden, H.C.; van der Eerden, M.M.; Hendriks, R.W. Systemic CD4+ and CD8+ T-cell cytokine profiles correlate with GOLD stage in stable COPD. Eur. Respir. J. 2011, 40, 330–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhodes, C.J.; Batai, K.; Bleda, M.; Haimel, M.; Southgate, L.; Germain, M.; Pauciulo, M.W.; Hadinnapola, C.; Aman, J.; Girerd, B.; et al. Genetic determinants of risk in pulmonary arterial hypertension: International genome-wide association studies and meta-analysis. Lancet Respir. Med. 2018, 7, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.; Yu, P.B. In Search of the Second Hit in Pulmonary Arterial Hypertension. Circ. Res. 2019, 124, 6–8. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Theiss, A.L.; Venuprasad, K. RORγt protein modifications and IL-17-mediated inflammation. Trends Immunol. 2021, 42, 1037–1050. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Flores, C.; Castro-Escamilla, O.; Ortega-Rocha, E.M.; Maldonado-García, C.; Cruz, F.J.-S.; Pérez-Montesinos, G.; Lemini-López, A.; Bonifaz, L.C. Association of Pathogenic Th17 Cells with the Disease Severity and Its Potential Implication for Biological Treatment Selection in Psoriasis Patients. Mediat. Inflamm. 2020, 2020, 8065147. [Google Scholar] [CrossRef]
- Yoo, H.H.B.; Marin, F.L. Treating Inflammation Associated with Pulmonary Hypertension: An Overview of the Literature. Int. J. Gen. Med. 2022, 15, 1075–1083. [Google Scholar] [CrossRef]
- GeurtsvanKessel, C.H.; Willart, M.A.; Bergen, I.M.; Van Rijt, L.S.; Muskens, F.; Elewaut, D.; Osterhaus, A.; Hendriks, R.; Rimmelzwaan, G.F.; Lambrecht, B.N. Dendritic cells are crucial for maintenance of tertiary lymphoid structures in the lung of influenza virus–infected mice. J. Exp. Med. 2009, 206, 2339–2349. [Google Scholar] [CrossRef]
- Collin, M.; Bigley, V. Human dendritic cell subsets: An update. Immunology 2018, 154, 3–20. [Google Scholar] [CrossRef]
- Hu, Y.; Chi, L.; Kuebler, W.M.; Goldenberg, N.M. Perivascular Inflammation in Pulmonary Arterial Hypertension. Cells 2020, 9, 2338. [Google Scholar] [CrossRef]
- Galié, N.; Humbert, M.; Vachiéry, J.-L.; Gibbs, S.; Lang, I.; Torbicki, A.; Simonneau, G.; Peacock, A.; Noordegraaf, A.V.; Beghetti, M.; et al. 2015 ESC/ERS Guidelines for the diagnosis and treatment of pulmonary hypertension. Eur. Respir. J. 2015, 46, 903–975. [Google Scholar] [CrossRef]
- Geenen, L.W.; Baggen, V.J.M.; Koudstaal, T.; Boomars, K.A.; Eindhoven, J.A.; Boersma, E.; Roos-Hesselink, J.W.; van den Bosch, A.E. The prognostic value of various biomarkers in adults with pulmonary hypertension; a multi-biomarker approach. Am. Heart J. 2019, 208, 91–99. [Google Scholar] [CrossRef]
- Van Uden, D.; Koudstaal, T.; van Hulst, J.A.C.; van den Bosch, T.P.P.; Vink, M.; Bergen, I.M.; Lila, K.A.; van den Bosch, A.E.; Bresser, P.; Kool, M.; et al. Evidence for a Role of CCR6+ T Cells in Chronic Thromboembolic Pulmonary Hypertension. Front. Immunol. 2022, 13, 861450. [Google Scholar] [CrossRef]
- Van der Ploeg, E.K.; Golebski, K.; van Nimwegen, M.; Fergusson, J.R.; Heesters, B.A.; Martinez-Gonzalez, I.; Kradolfer, C.M.A.; van Tol, S.; Scicluna, B.P.; de Bruijn, M.J.W.; et al. Steroid-resistant human inflammatory ILC2s are marked by CD45RO and elevated in type 2 respiratory diseases. Sci. Immunol. 2021, 6, eabd3489. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Tesmer, L.A.; Lundy, S.K.; Sarkar, S.; Fox, D.A. Th17 cells in human disease. Immunol. Rev. 2008, 223, 87–113. [Google Scholar] [CrossRef]
- Rakebrandt, N.; Littringer, K.; Joller, N. Regulatory T cells: Balancing protection versus pathology. Swiss Med. Wkly 2016, 146, w14343. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PAH—BASELINE | PAH—1Y FOLLOW-UP | |||||
|---|---|---|---|---|---|---|
| IPAH (n = 15) | CTD-PAH (n = 24) | p Value | IPAH (n = 11) | CTD-PAH (n = 12) | p Value | |
| Baseline clinical characteristics | ||||||
| Gender, female (%) | 13 (87%) | 20 (83%) | 10 (91%) | 11 (92%) | ||
| Age, y | 55.6 ± 16.7 | 64.6 ± 11.2 | 0.15 | 60.8 ± 14.8 | 65.3 ± 12.2 | 0.59 |
| BMI, kg/m2 | 28.4 ± 7.4 | 27.6 ± 5.4 | 0.86 | 29.1 ± 8.4 | 28.6 ± 6.1 | 0.96 |
| NYHA class 3–4, n (%) | 11 (73%) | 14 (58%) | 8 (73%) | 7 (58%) | ||
| 6MWT, m | 332 ± 126 | 299 ± 138 | 0.70 | 298 ± 121 | 334 ± 131 | 0.40 |
| NT-pro BNP, pmol/L | 242 ± 272 | 541 ± 1056 | 0.83 | 281 ± 308 | 301 ± 502 | 0.57 |
| Underlying CTD | ||||||
| SSc, n (%) | 20/24 (83%) | |||||
| SLE, n (%) | 4/24 (17%) | |||||
| Baseline right heart catheterization | ||||||
| mPAP, mmHg | 55.5 ± 15.2 | 44.0 ± 13.0 | 0.02 | 51.1 ± 12.7 | 41.5 ± 12.2 | 0.08 |
| mRAP, mmHg | 12.4 ± 6.3 | 9.5 ± 5.2 | 0.16 | 11.3 ± 6.5 | 8.3 ± 5.0 | 0.22 |
| Capillary wedge pressure, mmHg | 9.5 ± 4.8 | 13.5 ± 7.8 | 0.11 | 9.8 ± 5.3 | 13.6 ± 10 | 0.44 |
| PVR, wood units | 10.2 ± 3.1 | 6.2 ± 3.4 | 0.002 | 9.2 ± 3.0 | 5.7 ± 3.4 | 0.01 |
| PH-Medication | ||||||
| At baseline, n (%) | 0/15 (0%) | 0/24 (0%) | ||||
| At 1-year follow-up | ||||||
| No PH-medication | 0/11 (0%) | 0/12 (0%) | ||||
| Mono therapy (ERA), n (%) | 1/11 (9%) 1 | |||||
| Mono therapy (PDE5), n (%) | 1/12 (18%) 2 | |||||
| Duo therapy (PDE5 + ERA), n (%) | 6/11 (55%) | 9/12 (73%) | ||||
| Triple therapy (PDE5 + ERA + PRC), n (%) | 4/11 (36%) | 2/12 (9%) | ||||
| Immunomodulatory drugs | ||||||
| At baseline, n (%) | 0/15 (0%) | 3/24 (13%) | ||||
| At 1-year follow-up, n (%) | 0/11 (0%) | 3/12 (25%) | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Uden, D.; Koudstaal, T.; van Hulst, J.A.C.; Vink, M.; van Nimwegen, M.; van den Toorn, L.M.; Chandoesing, P.P.; van den Bosch, A.E.; Kool, M.; Hendriks, R.W.; et al. Peripheral Blood T Cells of Patients with IPAH Have a Reduced Cytokine-Producing Capacity. Int. J. Mol. Sci. 2022, 23, 6508. https://doi.org/10.3390/ijms23126508
van Uden D, Koudstaal T, van Hulst JAC, Vink M, van Nimwegen M, van den Toorn LM, Chandoesing PP, van den Bosch AE, Kool M, Hendriks RW, et al. Peripheral Blood T Cells of Patients with IPAH Have a Reduced Cytokine-Producing Capacity. International Journal of Molecular Sciences. 2022; 23(12):6508. https://doi.org/10.3390/ijms23126508
Chicago/Turabian Stylevan Uden, Denise, Thomas Koudstaal, Jennifer A. C. van Hulst, Madelief Vink, Menno van Nimwegen, Leon M. van den Toorn, Prewesh P. Chandoesing, Annemien E. van den Bosch, Mirjam Kool, Rudi W. Hendriks, and et al. 2022. "Peripheral Blood T Cells of Patients with IPAH Have a Reduced Cytokine-Producing Capacity" International Journal of Molecular Sciences 23, no. 12: 6508. https://doi.org/10.3390/ijms23126508
APA Stylevan Uden, D., Koudstaal, T., van Hulst, J. A. C., Vink, M., van Nimwegen, M., van den Toorn, L. M., Chandoesing, P. P., van den Bosch, A. E., Kool, M., Hendriks, R. W., & Boomars, K. A. (2022). Peripheral Blood T Cells of Patients with IPAH Have a Reduced Cytokine-Producing Capacity. International Journal of Molecular Sciences, 23(12), 6508. https://doi.org/10.3390/ijms23126508