
KOMPEITO, an Atypical Arabidopsis Rhomboid-Related Gene, Is Required for Callose Accumulation and Pollen Wall Development

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
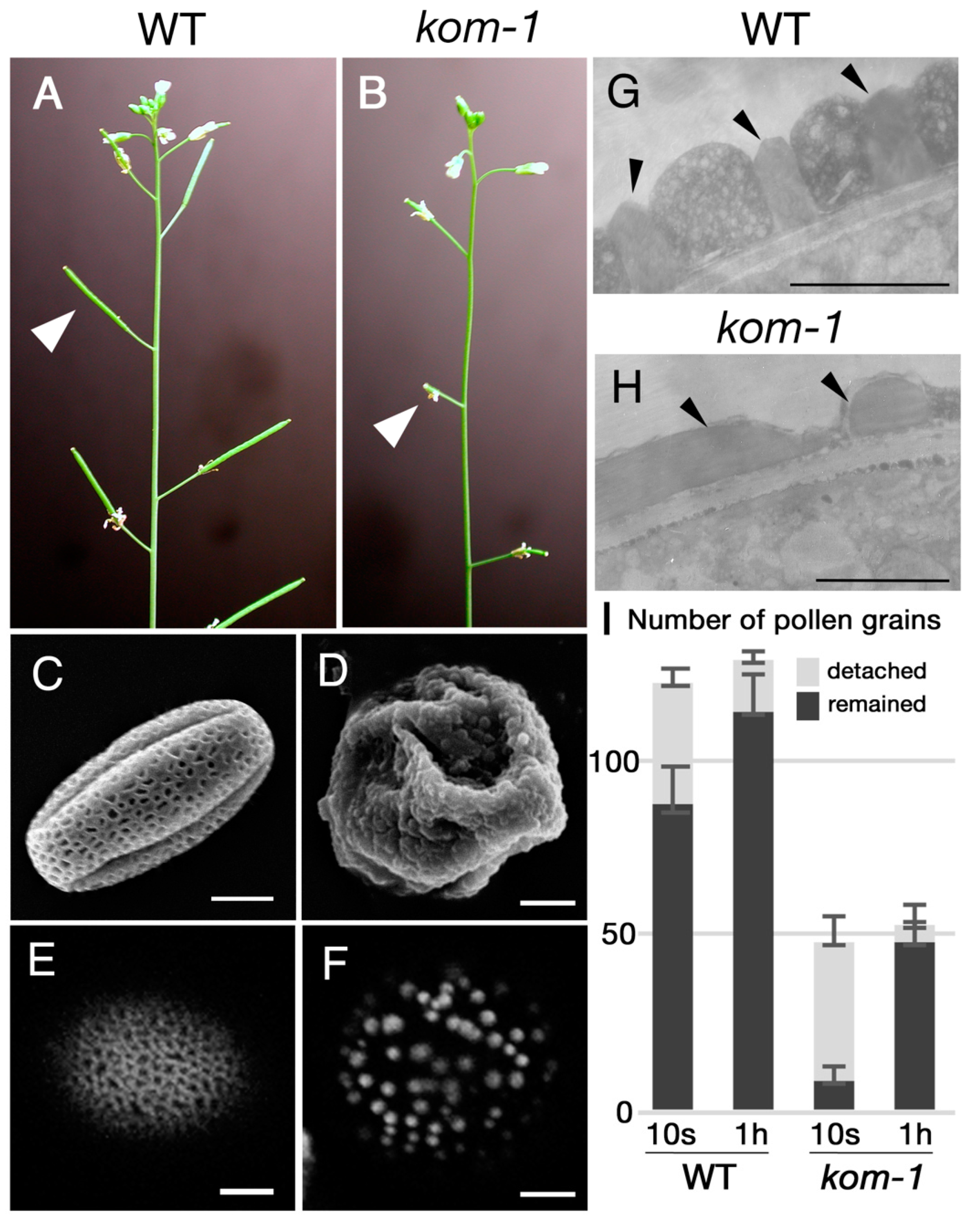
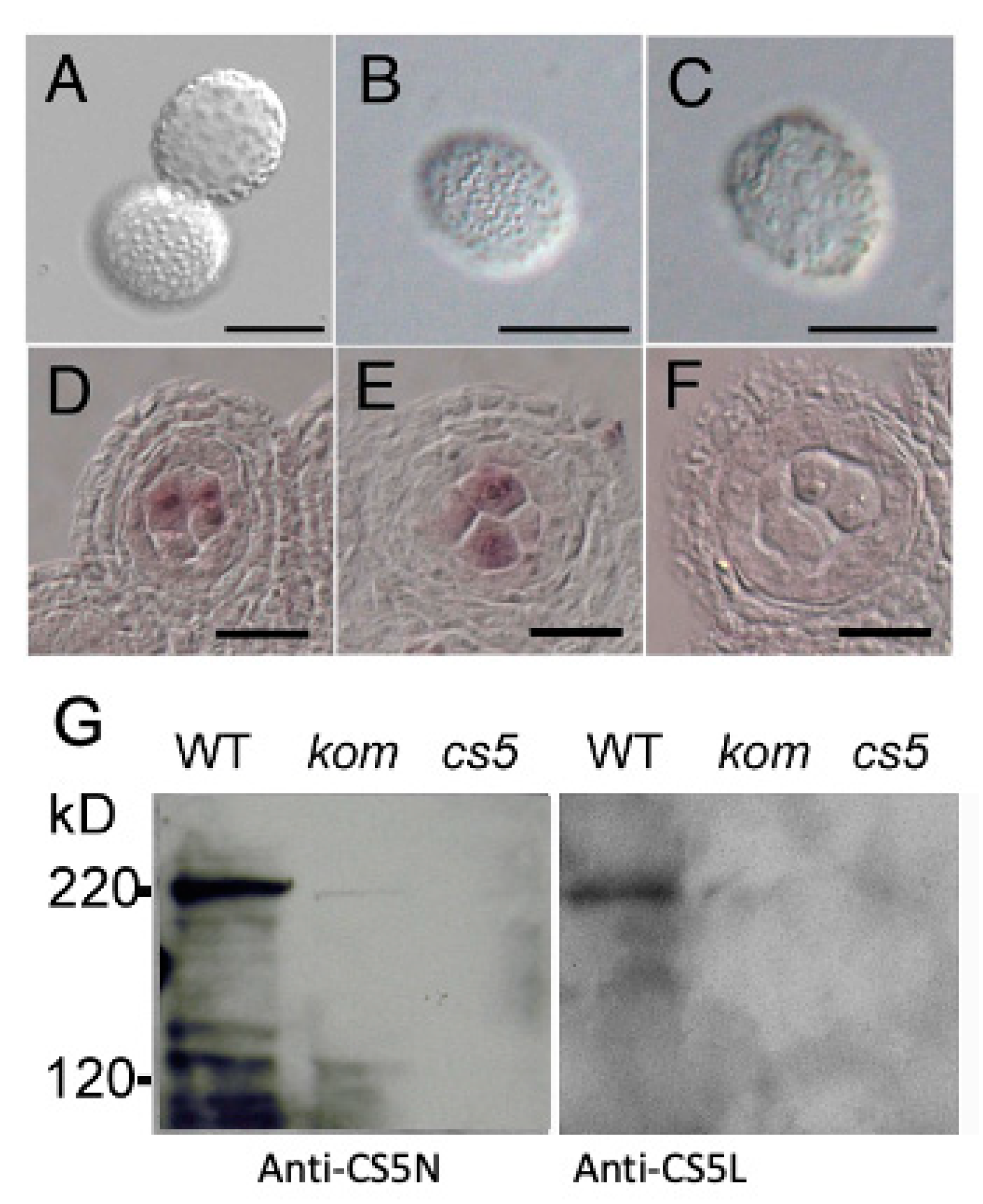
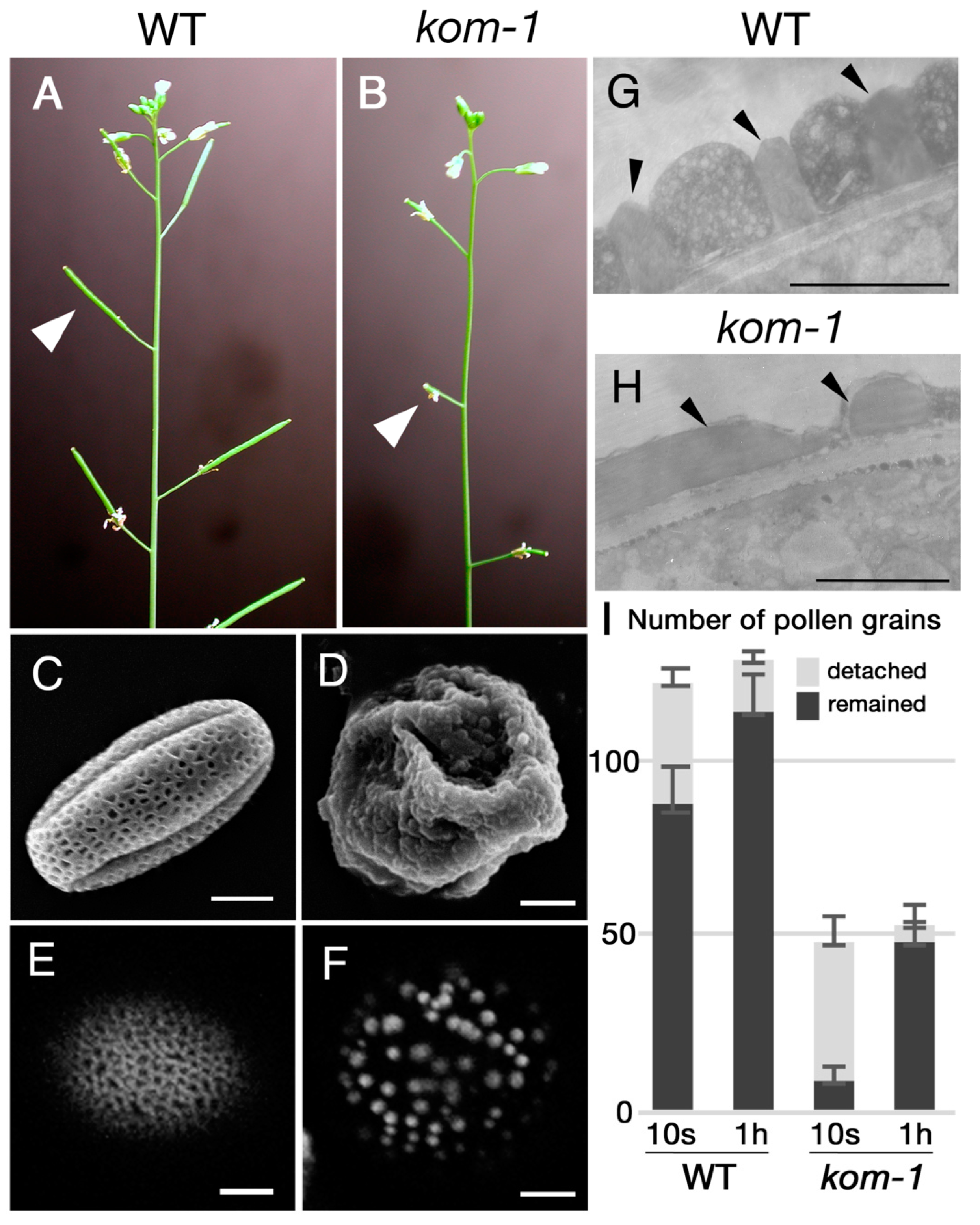
2.1. Isolation of Kompeito-1 (Kom-1), Which Was Defective in Pollen Wall and Pollen–Stigma Adhesion
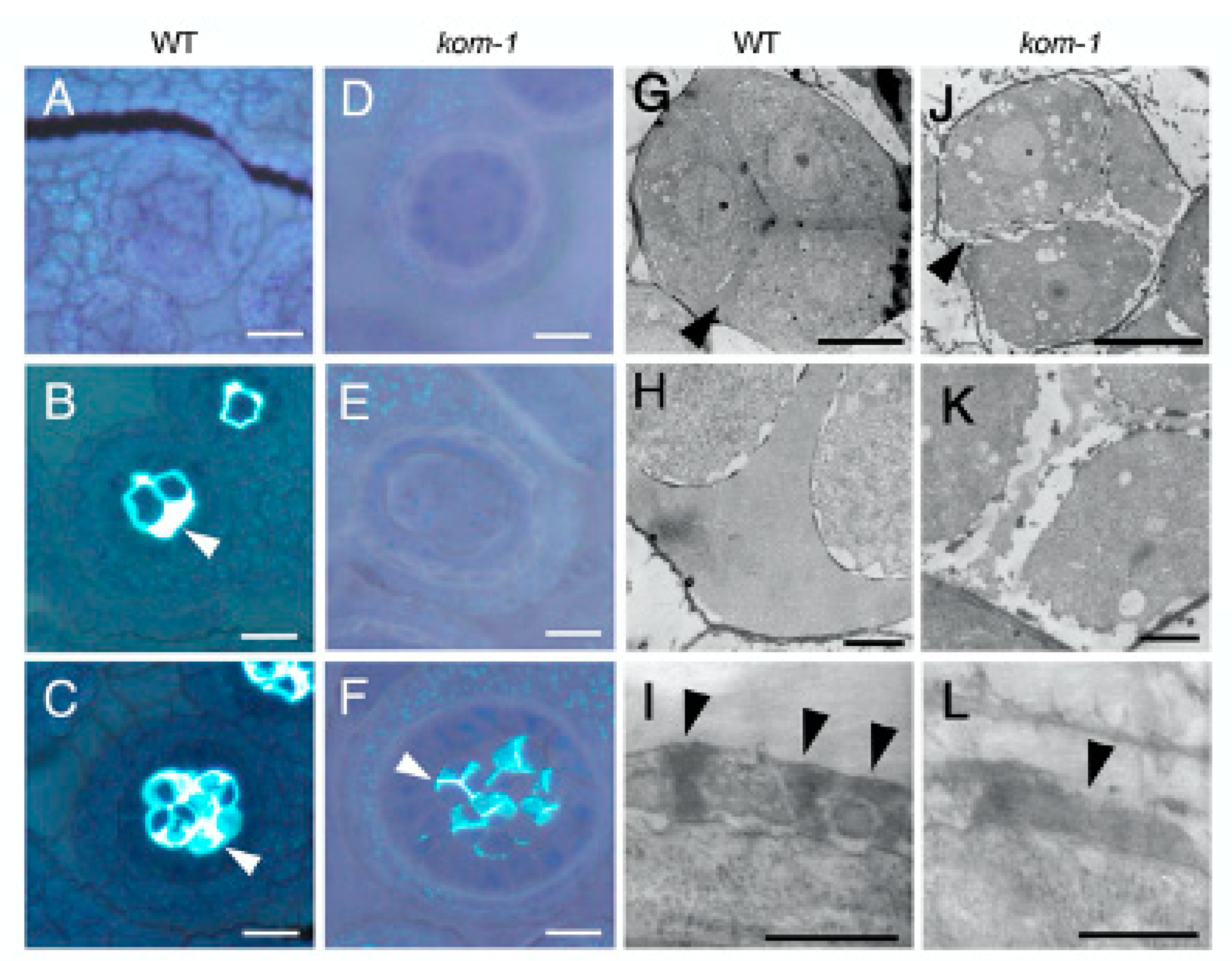
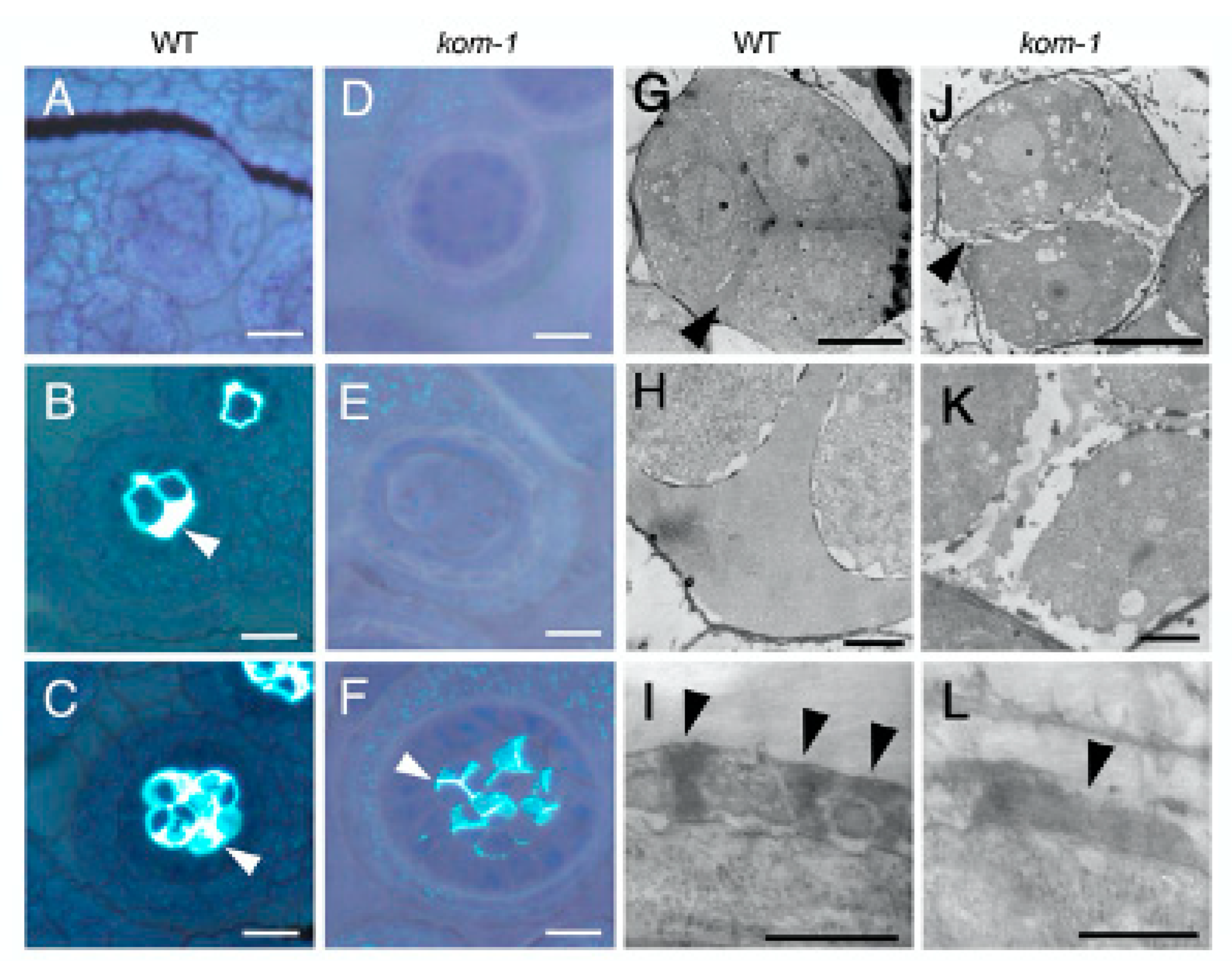
2.2. Defective Callose Wall Development in Kom-1
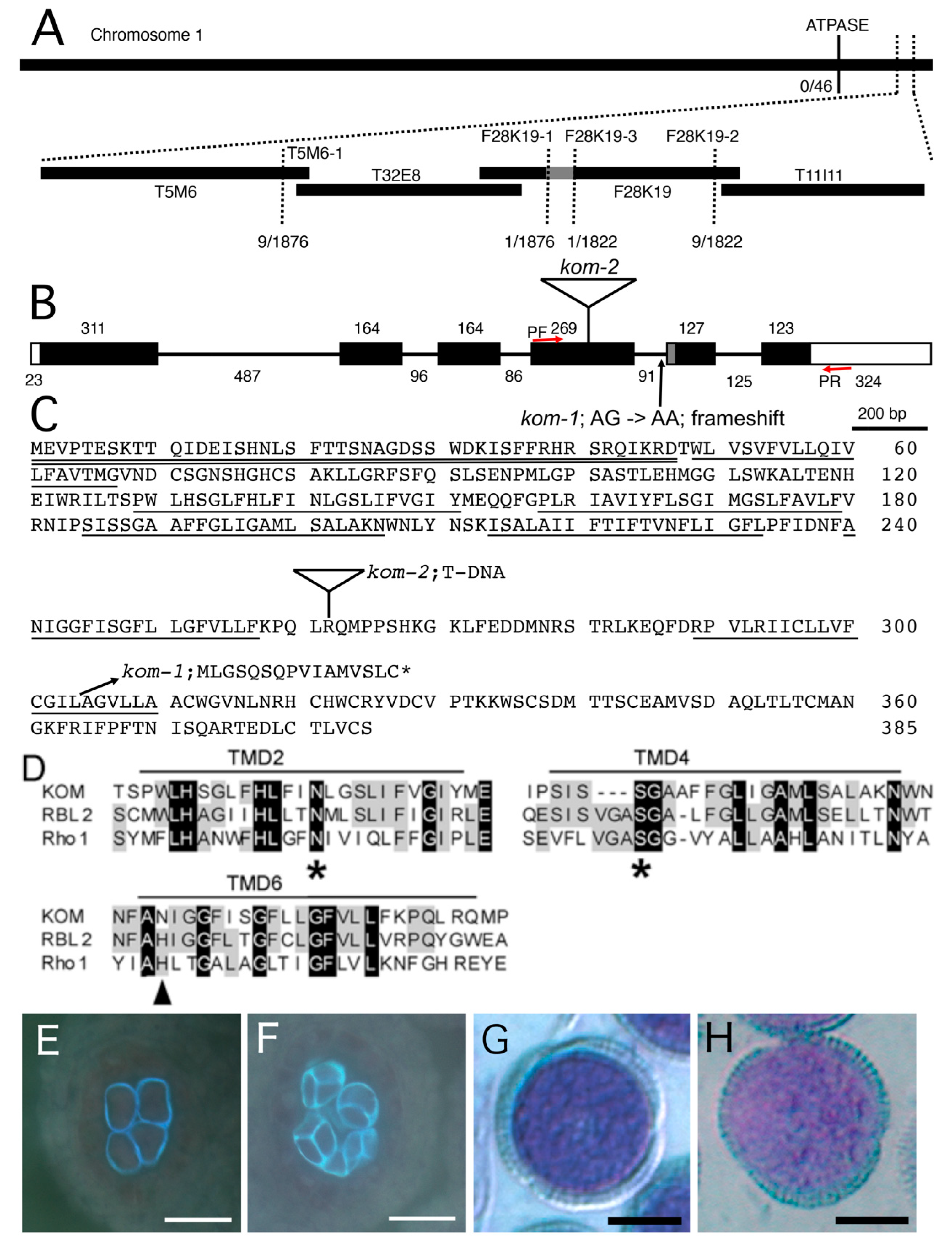
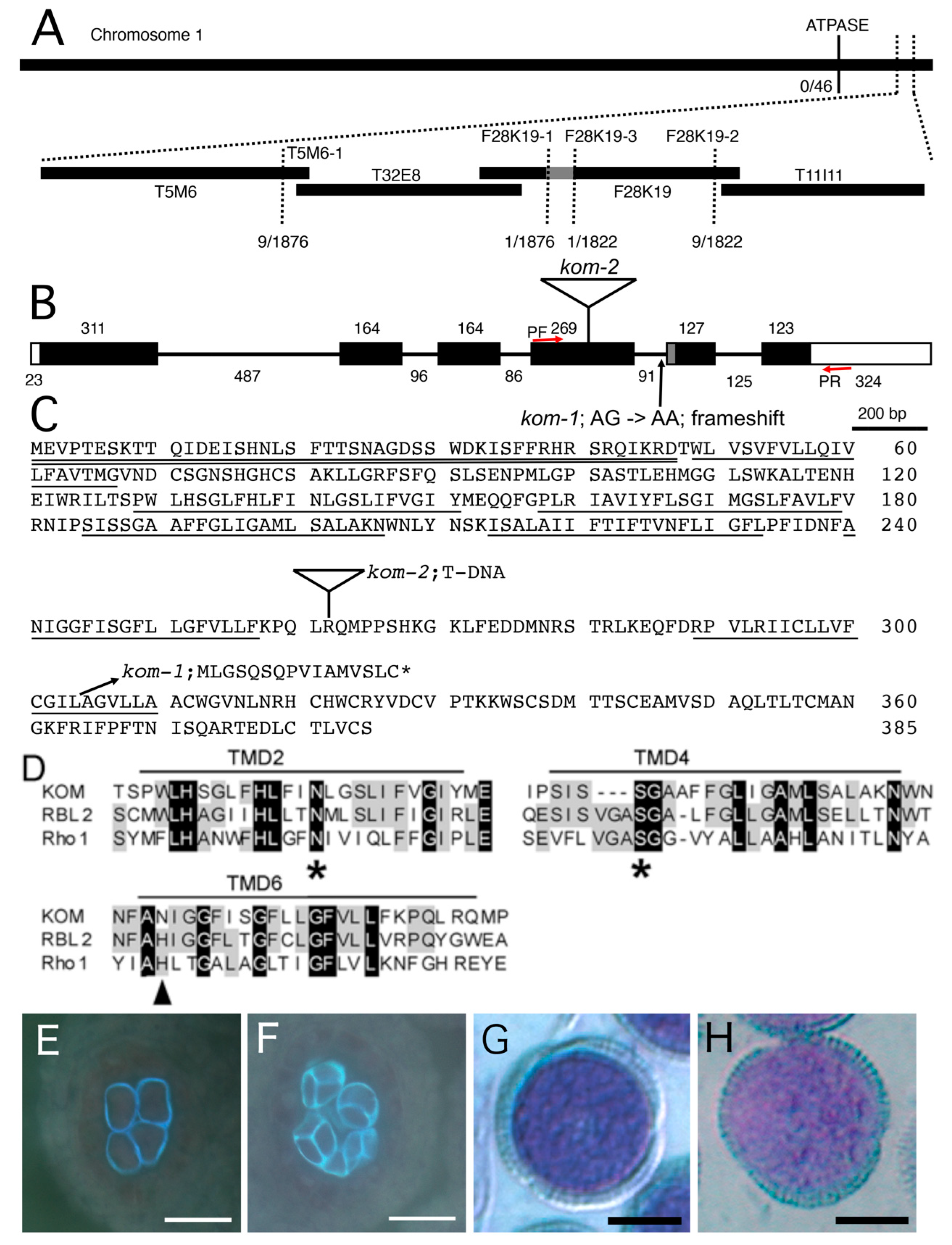
2.3. Mapping of the Kom-1 Locus and Complementation Using the KOM Genomic DNA
2.4. KOM Was Found to Be a Unique Member of the Rhomboid Family of Proteins
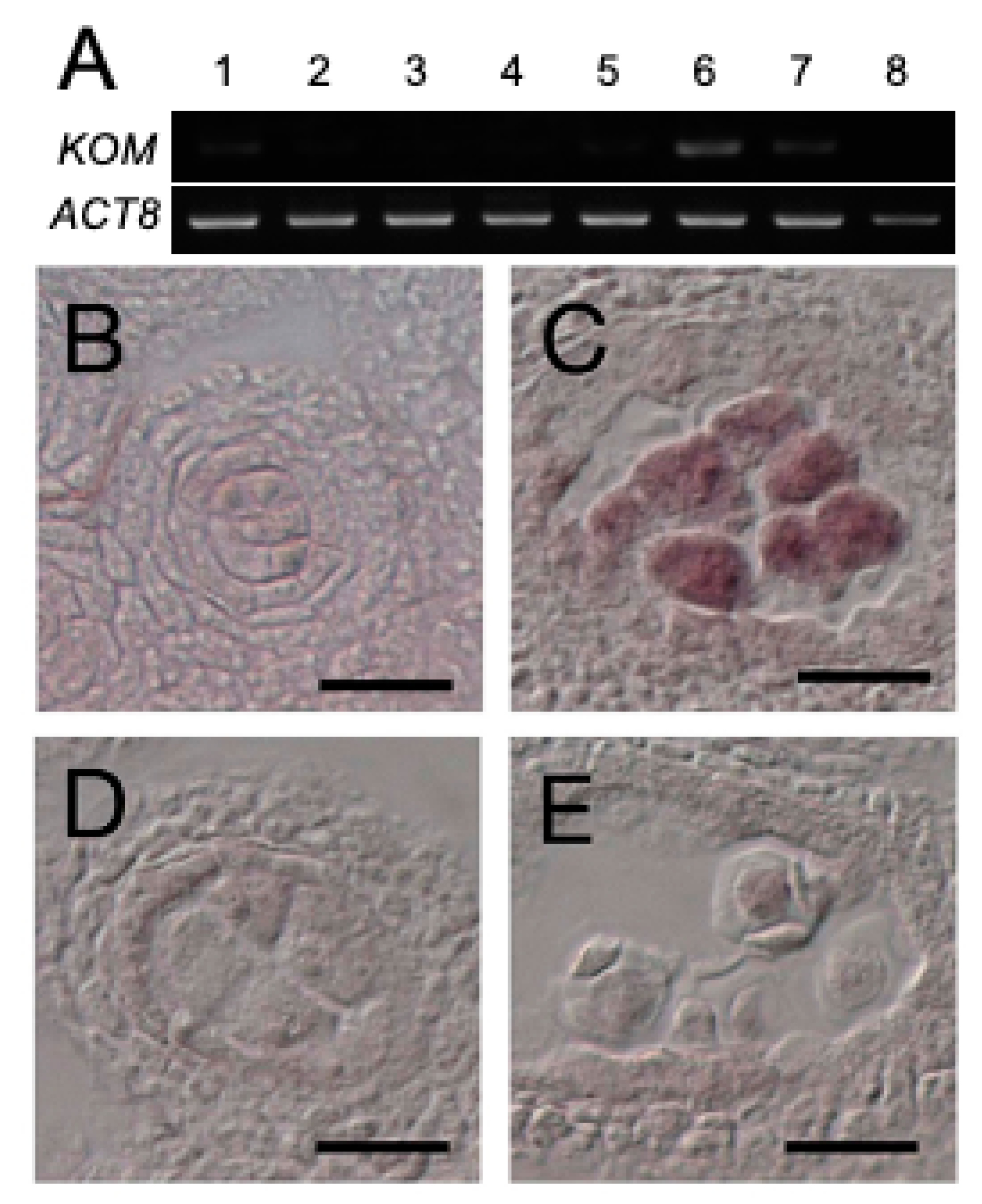
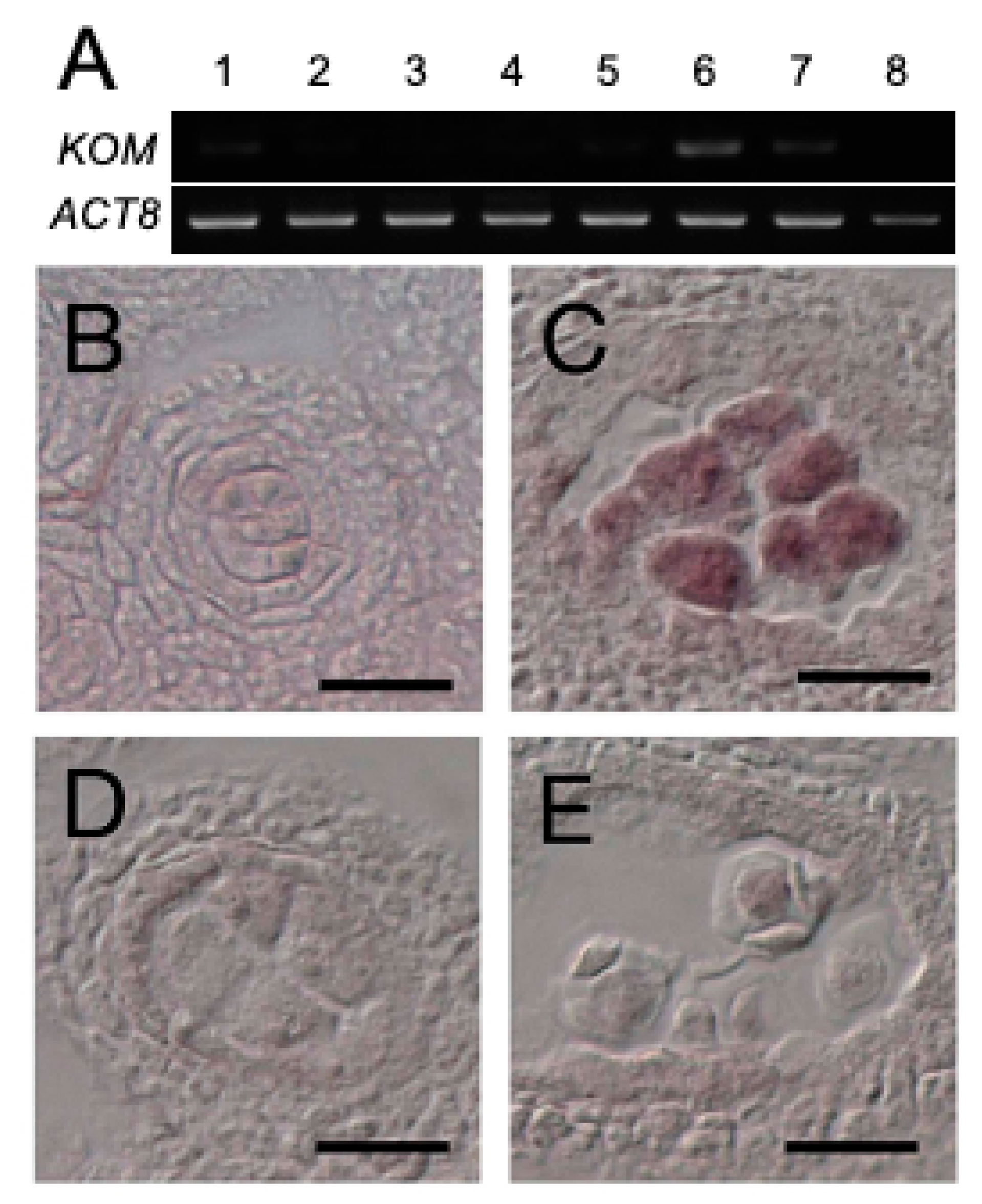
2.5. Expression of KOM-1 Was Restricted to the Meiocytes
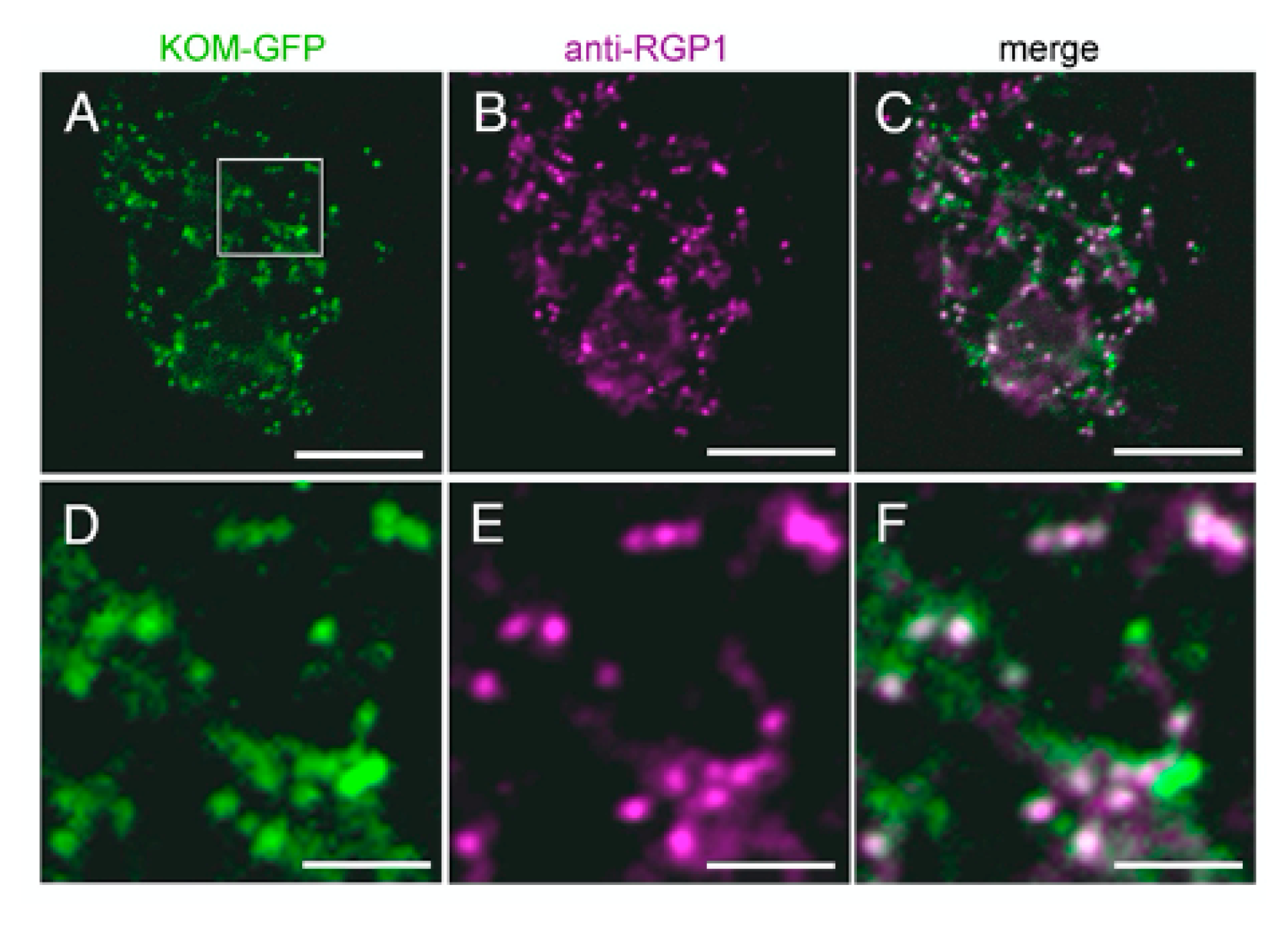
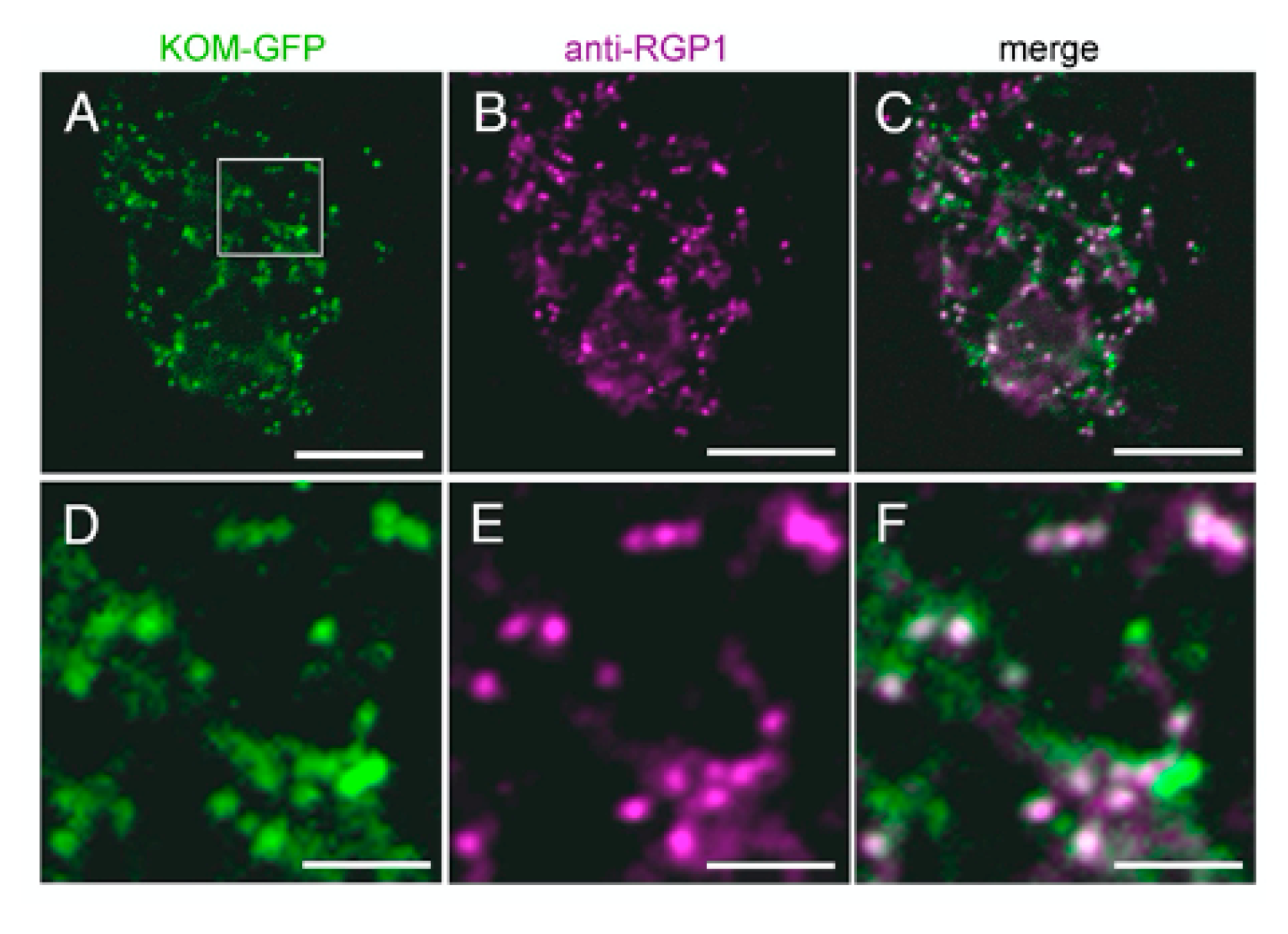
2.6. KOM Was Localized in the Golgi Apparatus
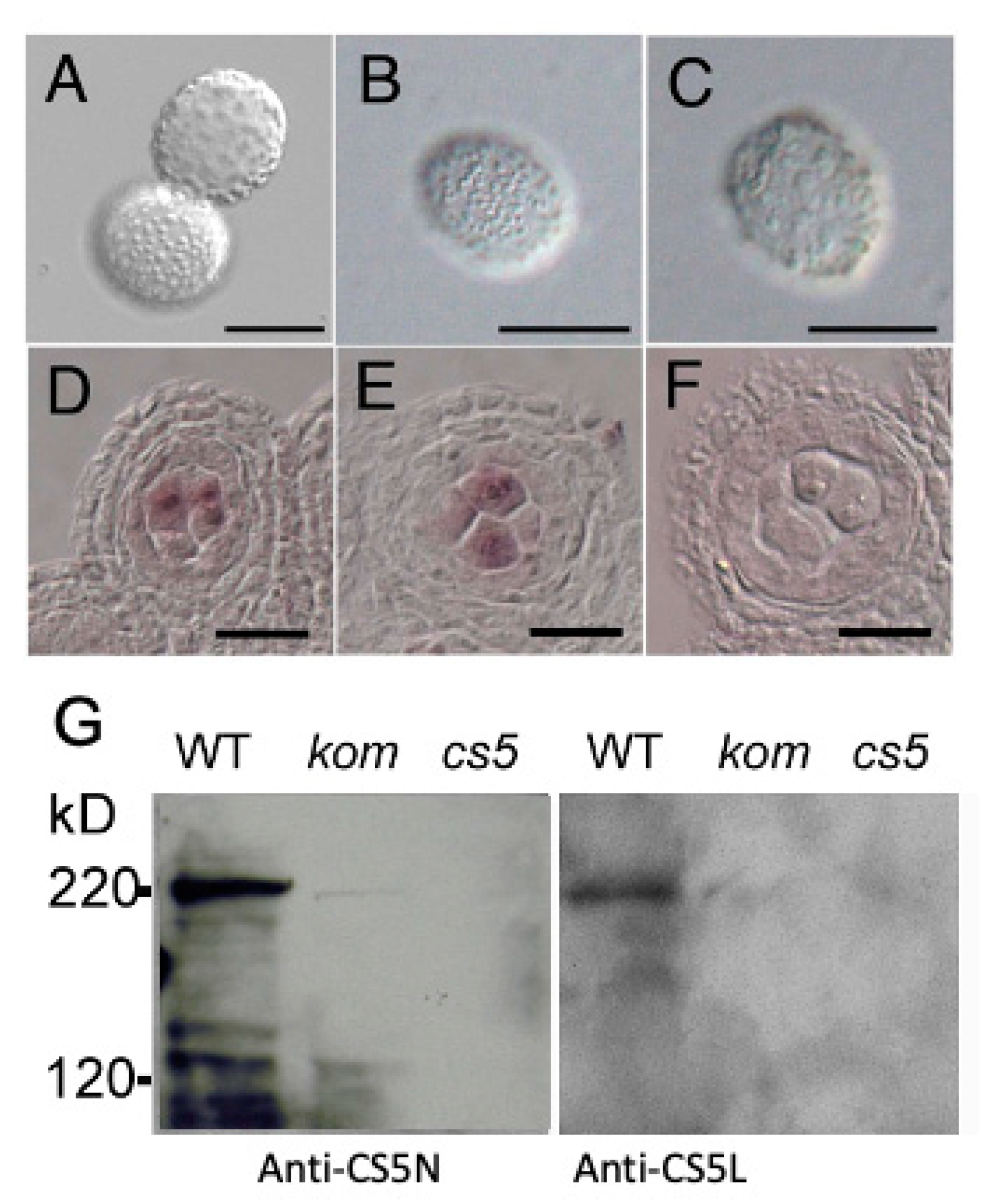
2.7. KOM Was Required for the Accumulation of CalS5 during Pollen Development
3. Discussion
3.1. Pollen Wall Patterning Was Required for Pollen–Stigma Adhesion
3.2. KOM Was a Novel Regulator of Callose and Pollen Wall Formation
3.3. KOM May Represent a New Function for Rhomboid-like Protein
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Microscopy
4.3. Pollen–Stigma Adhesion Assay
4.4. Mapping and Cloning of the KOM Gene
4.5. Complementation Test
4.6. Peptide Sequence Analysis
4.7. cDNA Cloning and RT-PCR
4.8. In Situ RNA Hybridization
4.9. Subcellular Localization Analysis
4.10. Western Blotting
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zinkl, G.M.; Zwiebel, B.I.; Grier, D.G.; Preuss, D. Pollen-stigma adhesion in Arabidopsis: A species-specific interaction mediated by lipophilic molecules in the pollen exine. Development 1999, 126, 5431–5440. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wu, Y.; Zhang, G. Formation pattern and regulatory mechanisms of pollen wall in Arabidopsis. J. Plant Physiol. 2021, 260, 153388. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Shi, J.; Yang, X. Role of Lipid Metabolism in Plant Pollen Exine Development. Subcell Biochem. 2016, 86, 315–337. [Google Scholar] [CrossRef] [PubMed]
- Quilichini, T.D.; Grienenberger, E.; Douglas, C.J. The biosynthesis, composition and assembly of the outer pollen wall: A tough case to crack. Phytochemistry 2015, 113, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Fan, X.D. Tapetum: Regulation and role in sporopollenin biosynthesis in Arabidopsis. Plant Mol. Biol. 2013, 83, 165–175. [Google Scholar] [CrossRef]
- Ariizumi, T.; Toriyama, K. Genetic regulation of sporopollenin synthesis and pollen exine development. Annu. Rev. Plant Biol. 2011, 62, 437–460. [Google Scholar] [CrossRef]
- Blackmore, S.; Wortley, A.H.; Skvarla, J.J.; Rowley, J.R. Pollen wall development in flowering plants. New Phytol. 2007, 174, 483–498. [Google Scholar] [CrossRef]
- Liu, L.; Wang, T. Male gametophyte development in flowering plants: A story of quarantine and sacrifice. J. Plant Physiol. 2021, 258–259, 153365. [Google Scholar] [CrossRef]
- Khan, R.M.; Yu, P.; Sun, L.; Abbas, A.; Shah, L.; Xiang, X.; Wang, D.; Sohail, A.; Zhang, Y.; Liu, Q.; et al. DCET1 Controls Male Sterility Through Callose Regulation, Exine Formation, and Tapetal Programmed Cell Death in Rice. Front. Genet. 2021, 12, 790789. [Google Scholar] [CrossRef]
- Chen, X.Y.; Kim, J.Y. Callose synthesis in higher plants. Plant Signal. Behav. 2009, 4, 489–492. [Google Scholar] [CrossRef] [Green Version]
- Hong, Z.; Delauney, A.J.; Verma, D.P. A cell plate-specific callose synthase and its interaction with phragmoplastin. Plant Cell 2001, 13, 755–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enns, L.C.; Kanaoka, M.M.; Torii, K.U.; Comai, L.; Okada, K.; Cleland, R.E. Two callose synthases, GSL1 and GSL5, play an essential and redundant role in plant and pollen development and in fertility. Plant Mol. Biol. 2005, 58, 333–349. [Google Scholar] [CrossRef] [PubMed]
- Verma, D.P.; Hong, Z. Plant callose synthase complexes. Plant Mol. Biol. 2001, 47, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Abercrombie, J.M.; O’Meara, B.C.; Moffatt, A.R.; Williams, J.H. Developmental evolution of flowering plant pollen tube cell walls: Callose synthase (CalS) gene expression patterns. Evodevo 2011, 2, 14. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Hong, Z.; Sivaramakrishnan, M.; Mahfouz, M.; Verma, D.P. Callose synthase (CalS5) is required for exine formation during microgametogenesis and for pollen viability in Arabidopsis. Plant J. 2005, 42, 315–328. [Google Scholar] [CrossRef]
- Nishikawa, S.; Zinkl, G.M.; Swanson, R.J.; Maruyama, D.; Preuss, D. Callose (beta-1,3 glucan) is essential for Arabidopsis pollen wall patterning, but not tube growth. BMC Plant Biol. 2005, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Xiong, S.X.; Zeng, Q.Y.; Hou, J.Q.; Hou, L.L.; Zhu, J.; Yang, M.; Yang, Z.N.; Lou, Y. The temporal regulation of TEK contributes to pollen wall exine patterning. PLoS Genet. 2020, 16, e1008807. [Google Scholar] [CrossRef]
- Zaveska Drabkova, L.; Honys, D. Evolutionary history of callose synthases in terrestrial plants with emphasis on proteins involved in male gametophyte development. PLoS ONE 2017, 12, e0187331. [Google Scholar] [CrossRef] [Green Version]
- Hulskamp, M.; Parekh, N.S.; Grini, P.; Schneitz, K.; Zimmermann, I.; Lolle, S.J.; Pruitt, R.E. The STUD gene is required for male-specific cytokinesis after telophase II of meiosis in Arabidopsis thaliana. Dev. Biol. 1997, 187, 114–124. [Google Scholar] [CrossRef] [Green Version]
- Spielman, M.; Preuss, D.; Li, F.L.; Browne, W.E.; Scott, R.J.; Dickinson, H.G. TETRASPORE is required for male meiotic cytokinesis in Arabidopsis thaliana. Development 1997, 124, 2645–2657. [Google Scholar] [CrossRef]
- Yang, C.Y.; Spielman, M.; Coles, J.P.; Li, Y.; Ghelani, S.; Bourdon, V.; Brown, R.C.; Lemmon, B.E.; Scott, R.J.; Dickinson, H.G. TETRASPORE encodes a kinesin required for male meiotic cytokinesis in Arabidopsis. Plant J. 2003, 34, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Ishikawa, M.; Kitamura, S.; Takahashi, Y.; Soyano, T.; Machida, C.; Machida, Y. The AtNACK1/HINKEL and STUD/TETRASPORE/AtNACK2 genes, which encode functionally redundant kinesins, are essential for cytokinesis in Arabidopsis. Genes. Cells 2004, 9, 1199–1211. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, T.; Toriyama, K.; Yoshikawa, M.; Ejiri, S.; Hinata, K. Tapetum-specific expression of the gene for an endo-beta-1,3-glucanase causes male sterility in transgenic tobacco. Plant Cell Physiol. 1995, 36, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Worrall, D.; Hird, D.L.; Hodge, R.; Paul, W.; Draper, J.; Scott, R. Premature dissolution of the microsporocyte callose wall causes male sterility in transgenic tobacco. Plant Cell 1992, 4, 759–771. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, L.; Wang, Y.; Wang, Y.C.; Wang, N.N.; Lu, R.; Wu, Y.W.; Li, X.B. Pollen-Specific Protein PSP231 Activates Callose Synthesis to Govern Male Gametogenesis and Pollen Germination. Plant Physiol. 2020, 184, 1024–1041. [Google Scholar] [CrossRef]
- Cui, X.; Shin, H.; Song, C.; Laosinchai, W.; Amano, Y.; Brown, R.M., Jr. A putative plant homolog of the yeast beta-1,3-glucan synthase subunit FKS1 from cotton (Gossypium hirsutum L.) fibers. Planta 2001, 213, 223–230. [Google Scholar] [CrossRef]
- Nakashima, J.; Laosinchai, W.; Cui, X.J.; Brown, R.M. New insight into the mechanism of cellulose and callose biosynthesis: Proteases may regulate callose biosynthesis upon wounding. Cellulose 2003, 10, 369–389. [Google Scholar] [CrossRef]
- Huang, X.Y.; Niu, J.; Sun, M.X.; Zhu, J.; Gao, J.F.; Yang, J.; Zhou, Q.; Yang, Z.N. CYCLIN-DEPENDENT KINASE G1 is associated with the spliceosome to regulate CALLOSE SYNTHASE5 splicing and pollen wall formation in Arabidopsis. Plant Cell 2013, 25, 637–648. [Google Scholar] [CrossRef] [Green Version]
- Peirson, B.N.; Owen, H.A.; Feldmann, K.A.; Makaroff, C.A. Characterization of three male-sterile mutants of Arabidopsis thaliana exhibiting alterations in meiosis. Sex. Plant Reprod. 1996, 9, 1–16. [Google Scholar] [CrossRef]
- Koonin, E.V.; Makarova, K.S.; Rogozin, I.B.; Davidovic, L.; Letellier, M.C.; Pellegrini, L. The Rhomboids: A nearly ubiquitous family of intramembrane serine proteases that probably evolved by multiple ancient horizontal gene transfers. Genome Biol. 2003, 4, R19. [Google Scholar] [CrossRef] [Green Version]
- Kanaoka, M.M.; Urban, S.; Freeman, M.; Okada, K. An Arabidopsis Rhomboid homolog is an intramembrane protease in plants. FEBS Lett. 2005, 579, 5723–5728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, L.P.; Sowdhamini, R. Cross genome comparisons of serine proteases in Arabidopsis and rice. BMC Genom. 2006, 7, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidorn-Czarna, M.; Maziak, A.; Janska, H. Protein Processing in Plant Mitochondria Compared to Yeast and Mammals. Front Plant Sci. 2022, 13, 824080. [Google Scholar] [CrossRef] [PubMed]
- Kandel, R.R.; Neal, S.E. The role of Rhomboid superfamily members in protein homeostasis: Mechanistic insight and physiological implications. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118793. [Google Scholar] [CrossRef]
- Kuhnle, N.; Dederer, V.; Lemberg, M.K. Intramembrane proteolysis at a glance: From signalling to protein degradation. J. Cell Sci. 2019, 132, jcs217745. [Google Scholar] [CrossRef] [Green Version]
- Ticha, A.; Collis, B.; Strisovsky, K. The Rhomboid Superfamily: Structural Mechanisms and Chemical Biology Opportunities. Trends Biochem. Sci. 2018, 43, 726–739. [Google Scholar] [CrossRef]
- Urban, S. A guide to the Rhomboid protein superfamily in development and disease. Semin. Cell Dev. Biol. 2016, 60, 1–4. [Google Scholar] [CrossRef]
- Urban, S.; Schlieper, D.; Freeman, M. Conservation of intramembrane proteolytic activity and substrate specificity in prokaryotic and eukaryotic Rhomboids. Curr. Biol. 2002, 12, 1507–1512. [Google Scholar] [CrossRef] [Green Version]
- Freeman, M. The Rhomboid-like superfamily: Molecular mechanisms and biological roles. Annu. Rev. Cell Dev. Biol. 2014, 30, 235–254. [Google Scholar] [CrossRef]
- McQuibban, G.A.; Saurya, S.; Freeman, M. Mitochondrial membrane remodelling regulated by a conserved Rhomboid protease. Nature 2003, 423, 537–541. [Google Scholar] [CrossRef]
- Lavell, A.; Froehlich, J.E.; Baylis, O.; Rotondo, A.D.; Benning, C. A predicted plastid Rhomboid protease affects phosphatidic acid metabolism in Arabidopsis thaliana. Plant J. 2019, 99, 978–987. [Google Scholar] [CrossRef] [PubMed]
- Adamiec, M.; Ciesielska, M.; Zalas, P.; Lucinski, R. Arabidopsis thaliana intramembrane proteases. Acta Physiol. Plant 2017, 39, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Dhugga, K.S.; Tiwari, S.C.; Ray, P.M. A reversibly glycosylated polypeptide (RGP1) possibly involved in plant cell wall synthesis: Purification, gene cloning, and trans-Golgi localization. Proc. Natl. Acad. Sci. USA 1997, 94, 7679–7684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urban, S.; Freeman, M. Intramembrane proteolysis controls diverse signalling pathways throughout evolution. Curr. Opin. Genet. Dev. 2002, 12, 512–518. [Google Scholar] [CrossRef]
- Urban, S.; Freeman, M. Substrate specificity of Rhomboid intramembrane proteases is governed by helix-breaking residues in the substrate transmembrane domain. Mol. Cell 2003, 11, 1425–1434. [Google Scholar] [CrossRef]
- Urban, S.; Lee, J.R.; Freeman, M. Drosophila Rhomboid-1 defines a family of putative intramembrane serine proteases. Cell 2001, 107, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Luu, D.T.; Heizmann, P.; Dumas, C. Pollen-Stigma Adhesion in Kale Is Not Dependent on the Self-(In)Compatibility Genotype. Plant Physiol. 1997, 115, 1221–1230. [Google Scholar] [CrossRef] [Green Version]
- Stead, A.D.; Roberts, I.N.; Dickinson, H.G. Pollen-stigma interaction in Brassica oleracea: The role of stigmatic proteins in pollen grain adhesion. J. Cell Sci. 1980, 42, 417–423. [Google Scholar] [CrossRef]
- Ariizumi, T.; Hatakeyama, K.; Hinata, K.; Inatsugi, R.; Nishida, I.; Sato, S.; Kato, T.; Tabata, S.; Toriyama, K. Disruption of the novel plant protein NEF1 affects lipid accumulation in the plastids of the tapetum and exine formation of pollen, resulting in male sterility in Arabidopsis thaliana. Plant J. 2004, 39, 170–181. [Google Scholar] [CrossRef]
- Ariizumi, T.; Hatakeyama, K.; Hinata, K.; Sato, S.; Kato, T.; Tabata, S.; Toriyama, K. A novel male-sterile mutant of Arabidopsis thaliana, faceless pollen-1, produces pollen with a smooth surface and an acetolysis-sensitive exine. Plant Mol. Biol. 2003, 53, 107–116. [Google Scholar] [CrossRef]
- Paxson-Sowders, D.M.; Dodrill, C.H.; Owen, H.A.; Makaroff, C.A. DEX1, a novel plant protein, is required for exine pattern formation during pollen development in Arabidopsis. Plant Physiol. 2001, 127, 1739–1749. [Google Scholar] [CrossRef] [PubMed]
- Aarts, M.G.; Hodge, R.; Kalantidis, K.; Florack, D.; Wilson, Z.A.; Mulligan, B.J.; Stiekema, W.J.; Scott, R.; Pereira, A. The Arabidopsis MALE STERILITY 2 protein shares similarity with reductases in elongation/condensation complexes. Plant J. 1997, 12, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Freeman, M. Proteolysis within the membrane: Rhomboids revealed. Nat. Rev. Mol. Cell Biol. 2004, 5, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Urban, S. Rhomboid proteins: Conserved membrane proteases with divergent biological functions. Genes. Dev. 2006, 20, 3054–3068. [Google Scholar] [CrossRef] [Green Version]
- Baker, R.P.; Wijetilaka, R.; Urban, S. Two Plasmodium Rhomboid proteases preferentially cleave different adhesins implicated in all invasive stages of malaria. PLoS Pathog. 2006, 2, e113. [Google Scholar] [CrossRef] [Green Version]
- Brossier, F.; Jewett, T.J.; Sibley, L.D.; Urban, S. A spatially localized Rhomboid protease cleaves cell surface adhesins essential for invasion by Toxoplasma. Proc. Natl. Acad. Sci. USA 2005, 102, 4146–4151. [Google Scholar] [CrossRef] [Green Version]
- Burzenski, L.M.; Low, B.E.; Kohar, V.; Shultz, L.D.; Wiles, M.V.; Hosur, V. Inactive Rhomboid proteins RHBDF1 and RHBDF2 (iRhoms): A decade of research in murine models. Mamm. Genome 2021, 32, 415–426. [Google Scholar] [CrossRef]
- Chao-Chu, J.; Murtough, S.; Zaman, N.; Pennington, D.J.; Blaydon, D.C.; Kelsell, D.P. iRHOM2: A Regulator of Palmoplantar Biology, Inflammation, and Viral Susceptibility. J. Investig. Dermatol. 2021, 141, 722–726. [Google Scholar] [CrossRef]
- Al-Salihi, M.A.; Lang, P.A. iRhom2: An Emerging Adaptor Regulating Immunity and Disease. Int. J. Mol. Sci. 2020, 21, 6570. [Google Scholar] [CrossRef]
- Geesala, R.; Issuree, P.D.; Maretzky, T. Novel functions of inactive Rhomboid proteins in immunity and disease. J. Leukoc. Biol. 2019, 106, 823–835. [Google Scholar] [CrossRef]
- Dulloo, I.; Muliyil, S.; Freeman, M. The molecular, cellular and pathophysiological roles of iRhom pseudoproteases. Open Biol. 2019, 9, 190003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, T.; Guichard, A.; Castro, C.P.; Xiao, Y.; Rizen, M.; Zhang, H.Z.; Hu, D.; Bang, A.; Helms, J.; Bier, E.; et al. Characterization of a human Rhomboid homolog, p100hRho/RHBDF1, which interacts with TGF-alpha family ligands. Dev. Dyn. 2005, 233, 1315–1331. [Google Scholar] [CrossRef] [PubMed]
- Hajdukiewicz, P.; Svab, Z.; Maliga, P. The small, versatile pPZP family of Agrobacterium binary vectors for plant transformation. Plant Mol. Biol. 1994, 25, 989–994. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, N.; Okada, K. A homeobox gene, PRESSED FLOWER, regulates lateral axis-dependent development of Arabidopsis flowers. Genes. Dev. 2001, 15, 3355–3364. [Google Scholar] [CrossRef] [Green Version]
- Ueda, M.; Matsui, K.; Ishiguro, S.; Sano, R.; Wada, T.; Paponov, I.; Palme, K.; Okada, K. The HALTED ROOT gene encoding the 26S proteasome subunit RPT2a is essential for the maintenance of Arabidopsis meristems. Development 2004, 131, 2101–2111. [Google Scholar] [CrossRef] [Green Version]
- Mathur, J.; Szabados, L.; Schaefer, S.; Grunenberg, B.; Lossow, A.; Jonas-Straube, E.; Schell, J.; Koncz, C.; Koncz-Kalman, Z. Gene identification with sequenced T-DNA tags generated by transformation of Arabidopsis cell suspension. Plant J. 1998, 13, 707–716. [Google Scholar] [CrossRef] [Green Version]
- Uemura, T.; Ueda, T.; Ohniwa, R.L.; Nakano, A.; Takeyasu, K.; Sato, M.H. Systematic analysis of SNARE molecules in Arabidopsis: Dissection of the post-Golgi network in plant cells. Cell Struct. Funct. 2004, 29, 49–65. [Google Scholar] [CrossRef] [Green Version]
- Mitsuda, N.; Enami, K.; Nakata, M.; Takeyasu, K.; Sato, M.H. Novel type Arabidopsis thaliana H+-PPase is localized to the Golgi apparatus. FEBS Lett. 2001, 488, 29–33. [Google Scholar] [CrossRef] [Green Version]
- Shimada, T.; Watanabe, E.; Tamura, K.; Hayashi, Y.; Nishimura, M.; Hara-Nishimura, I. A vacuolar sorting receptor PV72 on the membrane of vesicles that accumulate precursors of seed storage proteins (PAC vesicles). Plant Cell Physiol. 2002, 43, 1086–1095. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.R.; Urban, S.; Garvey, C.F.; Freeman, M. Regulated intracellular ligand transport and proteolysis control EGF signal activation in Drosophila. Cell 2001, 107, 161–171. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanaoka, M.M.; Shimizu, K.K.; Xie, B.; Urban, S.; Freeman, M.; Hong, Z.; Okada, K. KOMPEITO, an Atypical Arabidopsis Rhomboid-Related Gene, Is Required for Callose Accumulation and Pollen Wall Development. Int. J. Mol. Sci. 2022, 23, 5959. https://doi.org/10.3390/ijms23115959
Kanaoka MM, Shimizu KK, Xie B, Urban S, Freeman M, Hong Z, Okada K. KOMPEITO, an Atypical Arabidopsis Rhomboid-Related Gene, Is Required for Callose Accumulation and Pollen Wall Development. International Journal of Molecular Sciences. 2022; 23(11):5959. https://doi.org/10.3390/ijms23115959
Chicago/Turabian StyleKanaoka, Masahiro M., Kentaro K. Shimizu, Bo Xie, Sinisa Urban, Matthew Freeman, Zonglie Hong, and Kiyotaka Okada. 2022. "KOMPEITO, an Atypical Arabidopsis Rhomboid-Related Gene, Is Required for Callose Accumulation and Pollen Wall Development" International Journal of Molecular Sciences 23, no. 11: 5959. https://doi.org/10.3390/ijms23115959
APA StyleKanaoka, M. M., Shimizu, K. K., Xie, B., Urban, S., Freeman, M., Hong, Z., & Okada, K. (2022). KOMPEITO, an Atypical Arabidopsis Rhomboid-Related Gene, Is Required for Callose Accumulation and Pollen Wall Development. International Journal of Molecular Sciences, 23(11), 5959. https://doi.org/10.3390/ijms23115959