
Ribonomics Approaches to Identify RBPome in Plants and Other Eukaryotes: Current Progress and Future Prospects



Abstract
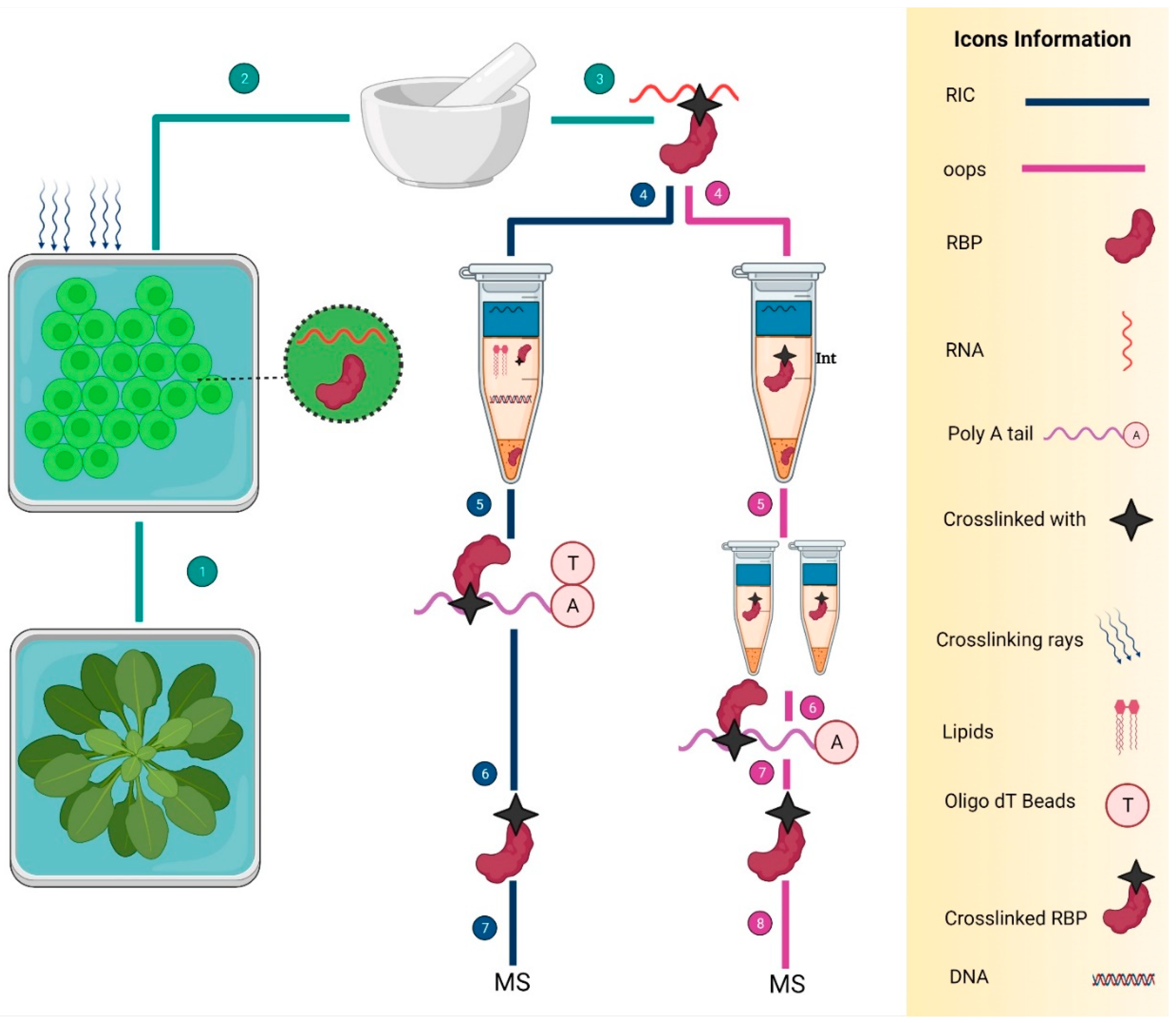
:1. Introduction
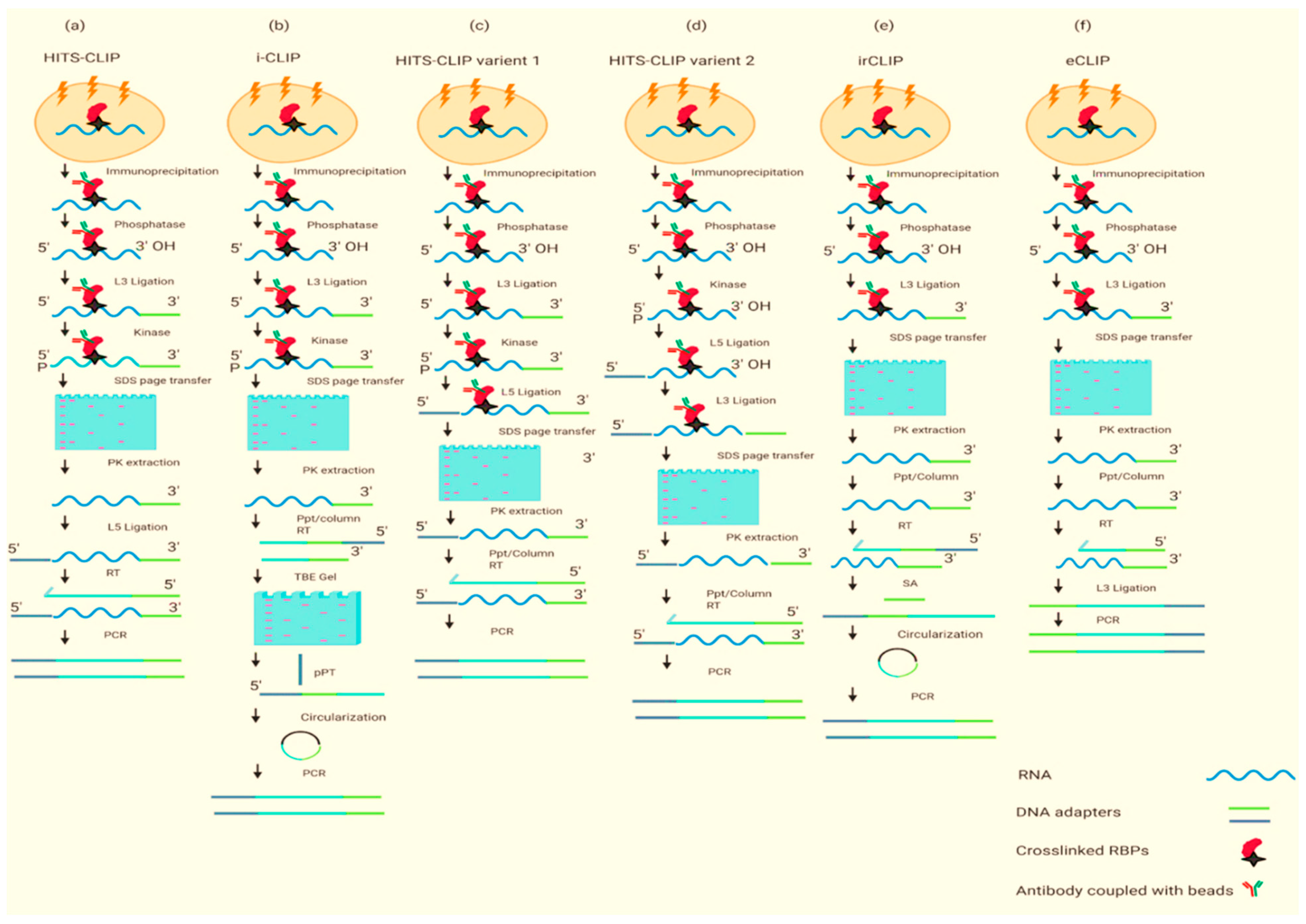
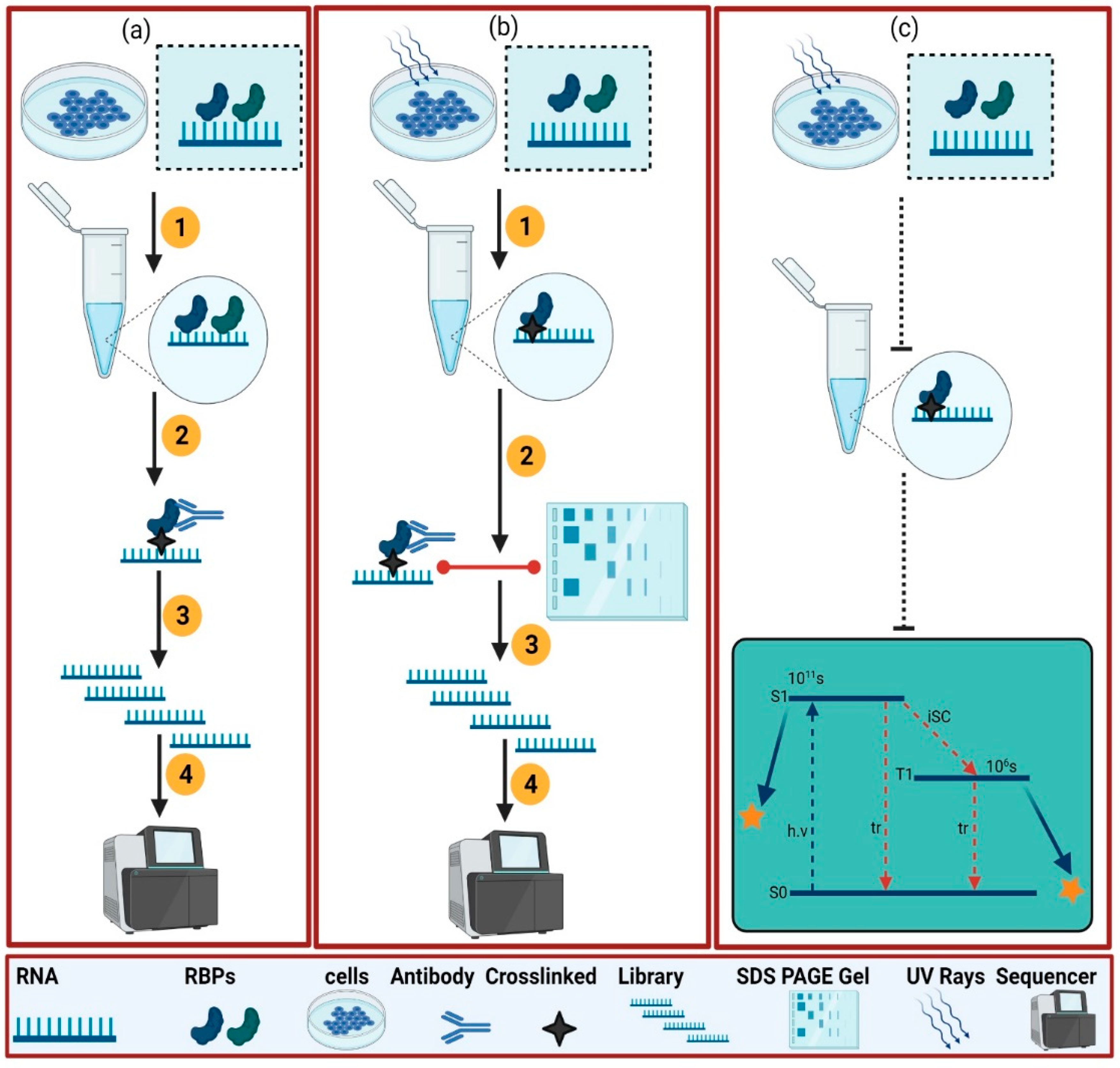
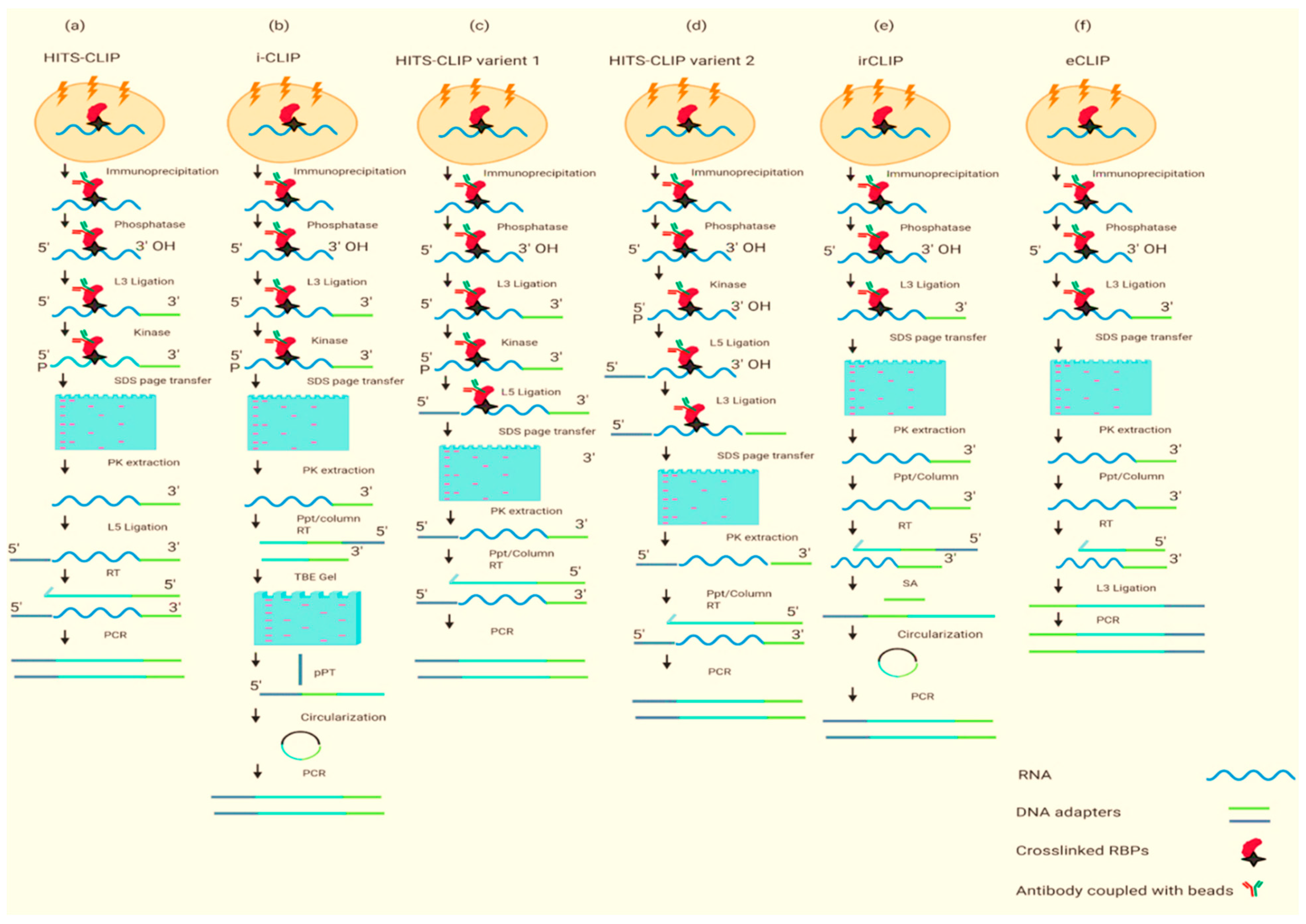
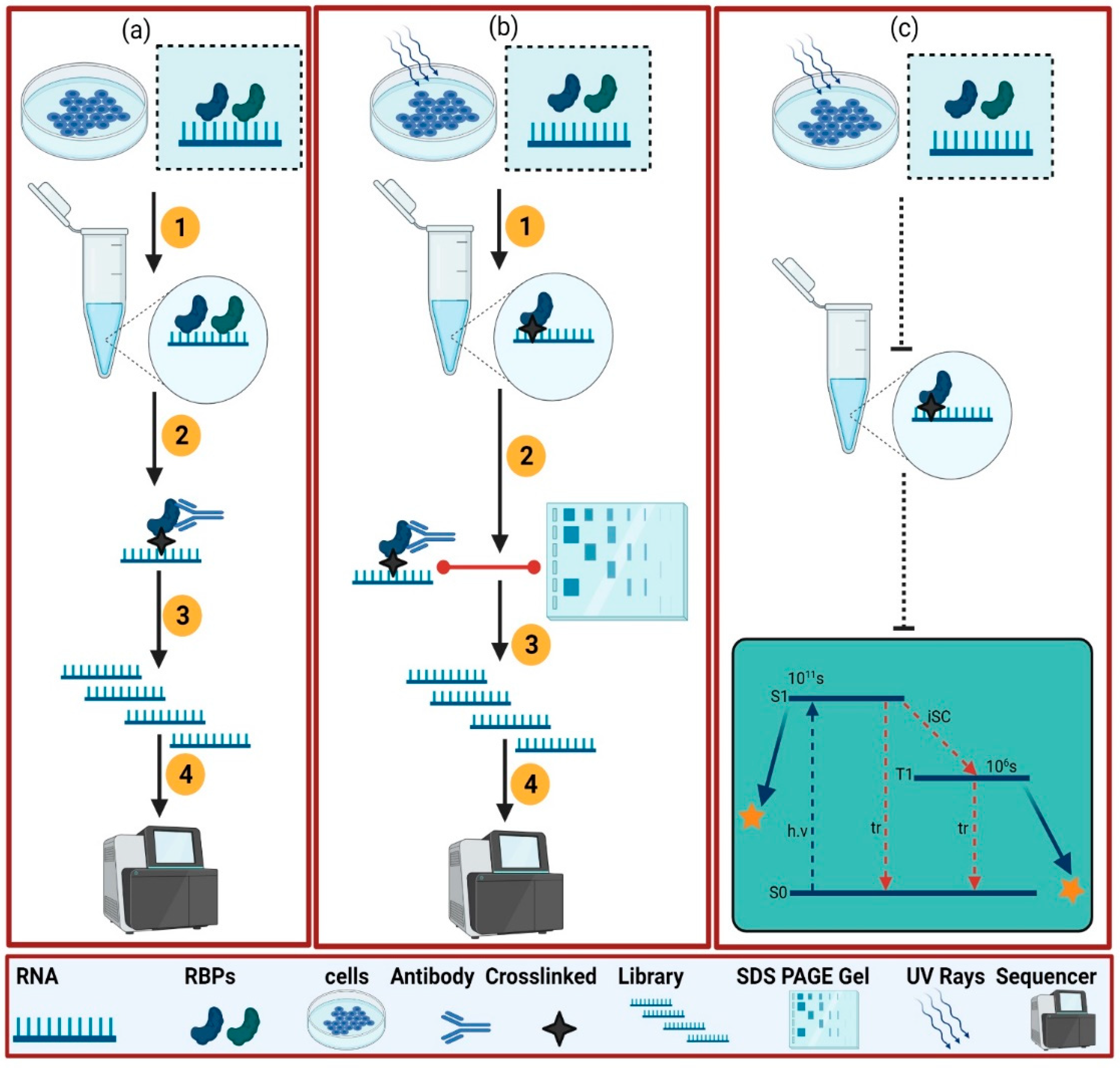
2. Experimental Approaches to Dissect the RBPome
2.1. Protein-Centric
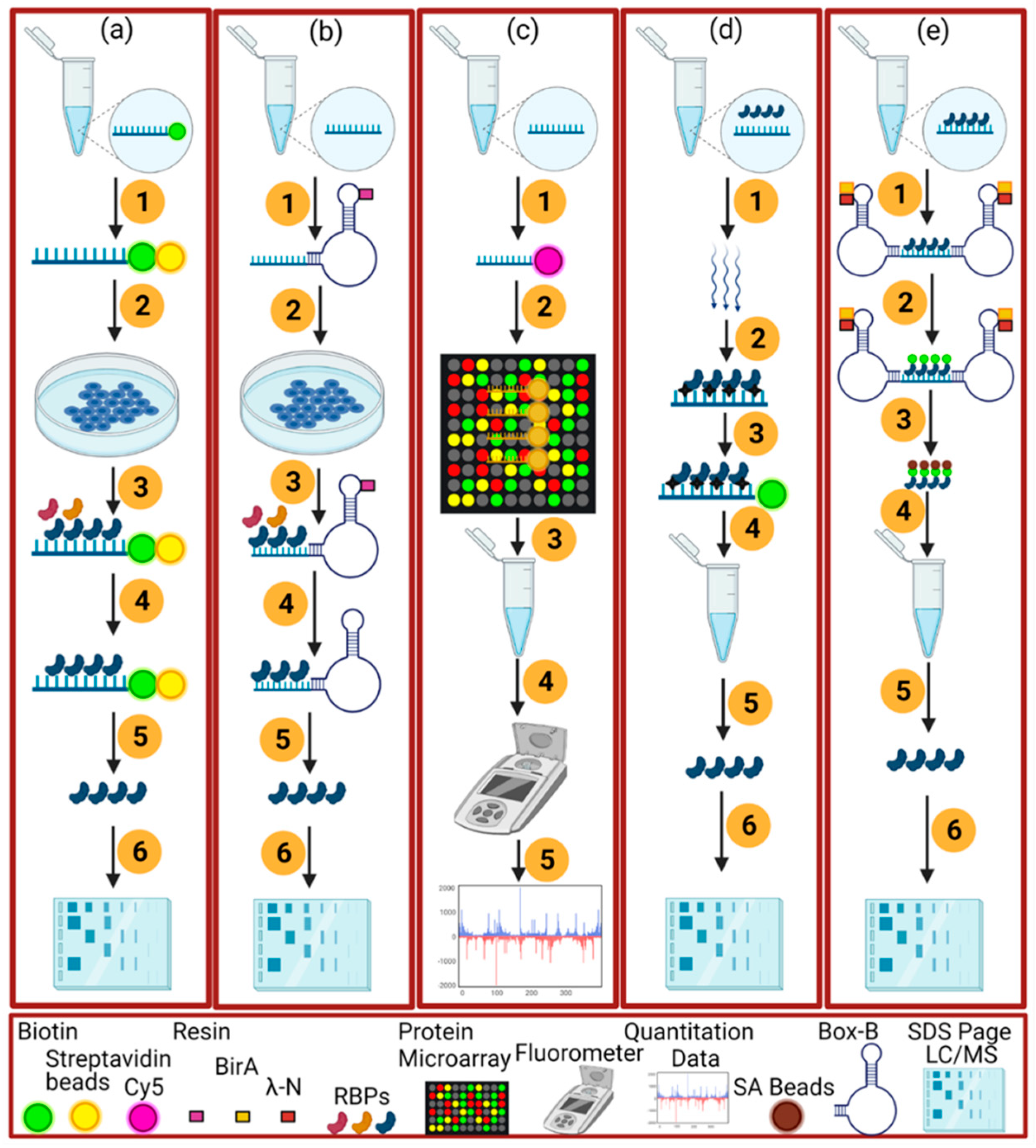
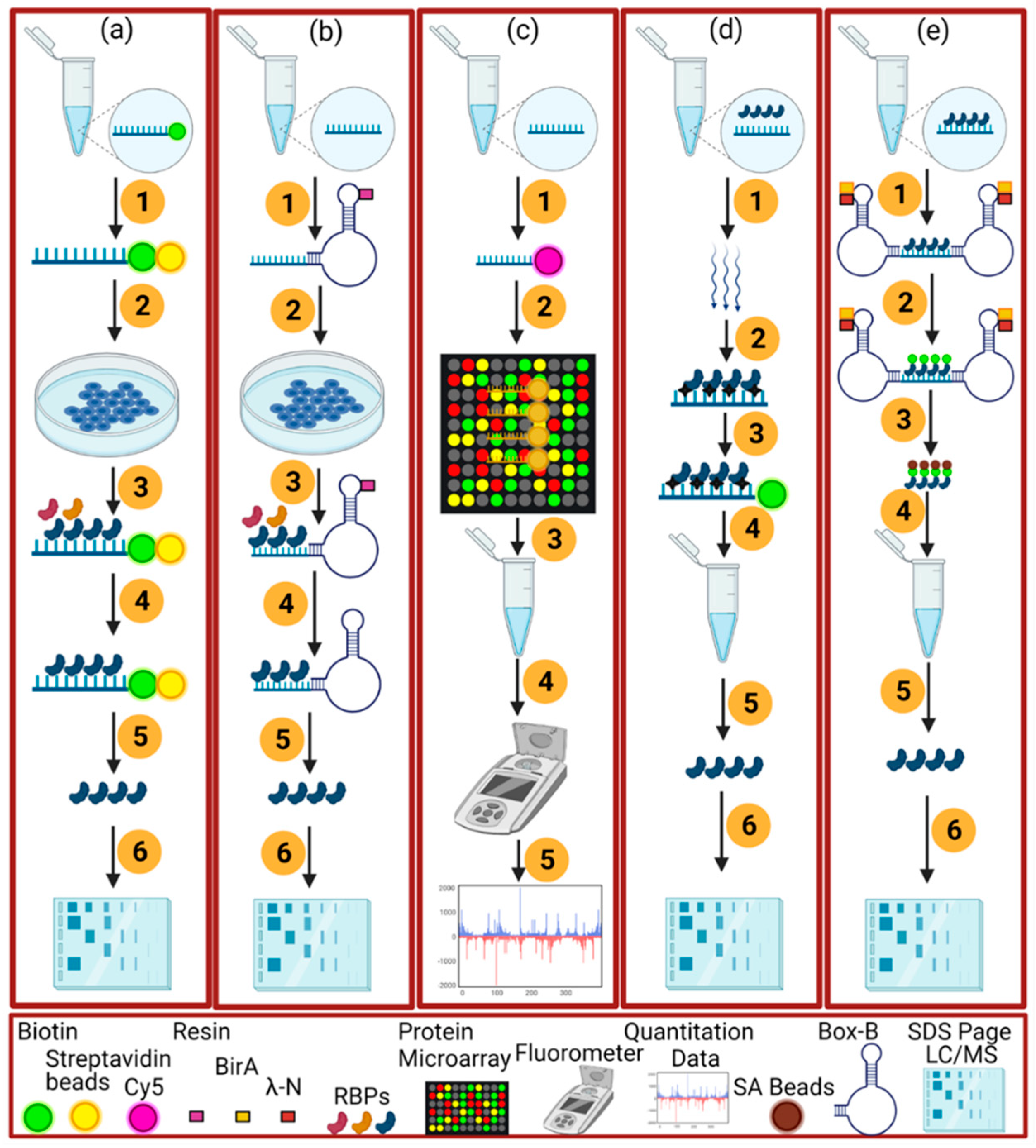
2.2. RNA-Centric Approaches

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Advantage | Disadvantage | Ref |
|---|---|---|---|
| PAR-CLIP, HITS-CLIP | Efficient, RBPs can be identified at the 3′ end of RNA, highly specific | UV cross-linking does not bind the RNA proteins well which can lead to a high number of false positives, crosslinked nucleotides are bypassed by reverse transcriptase, it is a laborious and time-consuming protocol | [24,25] |
| iCLIP | An efficient method that does not need the reverse transcription to bypass the bound nucleotides, high resolution | UV cross-linking does not bind the RNA proteins well, interactions near the 3′ end of an RNA may be unidentifiable because reverse transcriptase stops at the cross-linked nucleotide, unable to identify RNA–protein interaction near 3′ RNA end as reverse transcription stops at the crosslinked nucleotide | [26] |
| HITS-CLIP variants 1 and 2 | A short method, more reliable as both ligations are carried out by beads, genome-wide | Low efficiency of UV cross-linking, crosslinked nucleotides are bypassed by reverse transcriptase | [27] |
| eCLIP, | It does not need a circularization step which makes it unreliable, decreases requisite amplification by ~1000-fold | Low efficiency of UV cross-linking, time-consuming protocol, single-stranded DNA adapter is obligatory for the ligation to single-stranded cDNA. | [27] |
| sCLIP | Simplified, robust, permits a radiolabel-free visualization of immunoprecipitated RNA, improves the complexity of the sequencing library | Low efficiency of UV cross-linking, time consuming | [28] |
| irCLIP | Efficient, use of fluorescent adapter at each step to visualize crosslinked RNA | Low efficiency of UV cross-linking, time-consuming protocol, unable to identify RNA–protein interaction near 3′ RNA end as reverse transcription stops at the crosslinked nucleotide | [29] |
| GoldCLIP | Less time-consuming protocol, gel purification is not necessary | Low efficiency of UV cross-linking, expression of the fusion protein is necessary, unable to identify RNA–protein interaction near 3′ RNA end as reverse transcription stops at the crosslinked nucleotide | [30] |
| fCLIP | Use of formaldehyde instead of UV cross-linking, higher efficiency for developing the interaction between double-stranded RNA and proteins | Time-consuming protocol, formaldehyde crosslinking is not as good as UV cross-linking | [31] |
| BrdU-CLIP | Efficient for removing the “empty adapter” reads during reverse transcription because these reads can clutter HITS data | Low efficiency of UV cross-linking, time-consuming protocol, unable to identify RNA–protein interaction near 3′ RNA end as reverse transcription stops at the crosslinked nucleotide | [32] |
| TRIBE | The protein of interest is not purified for this protocol, no use of UV cross-linking, RBPs can develop interaction at any site of RNA | Not very effective in RBPome, it was not used for many studies | [33] |
| CRAC | Efficient due to the two-step affinity purification of tagged proteins in yeast, completely removes any interaction RBPs, RNAs that are not crosslinked to the protein of interest | Only works under denaturing conditions, very challenging, time-consuming, long protocol | [34] |
| RNA tagging | The protein of interest is not purified for this protocol, no use of UV cross-linking, RBPs can develop interaction at any site of RNA, easy protocol | Only studied in Saccharomyces cerevisiae, does not work well for the RBPs which are away from the 3′ RNA end | [35] |
| HyperTRIBE | Simple as a comparison to other protein-centric methods to identify the targets of the RBPs | A lot of validation steps are involved | [17] |
2.2.1. In Vitro Methods
2.2.2. In Vivo Method
3. In Silico Approaches
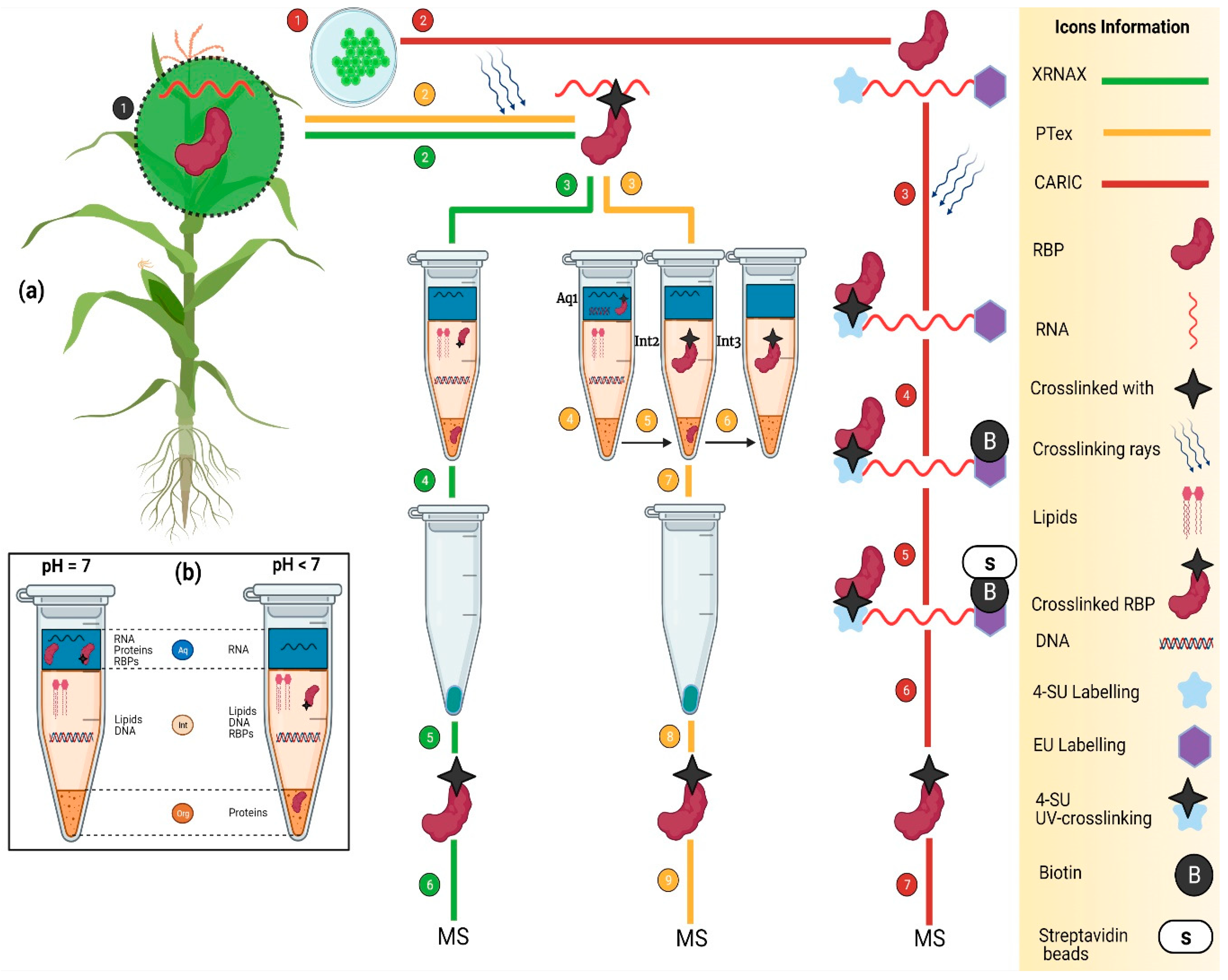
4. Transcriptome-Wide Identification of RBPs
4.1. XRNAX
4.2. PTex
4.3. CARIC
4.4. OOPS
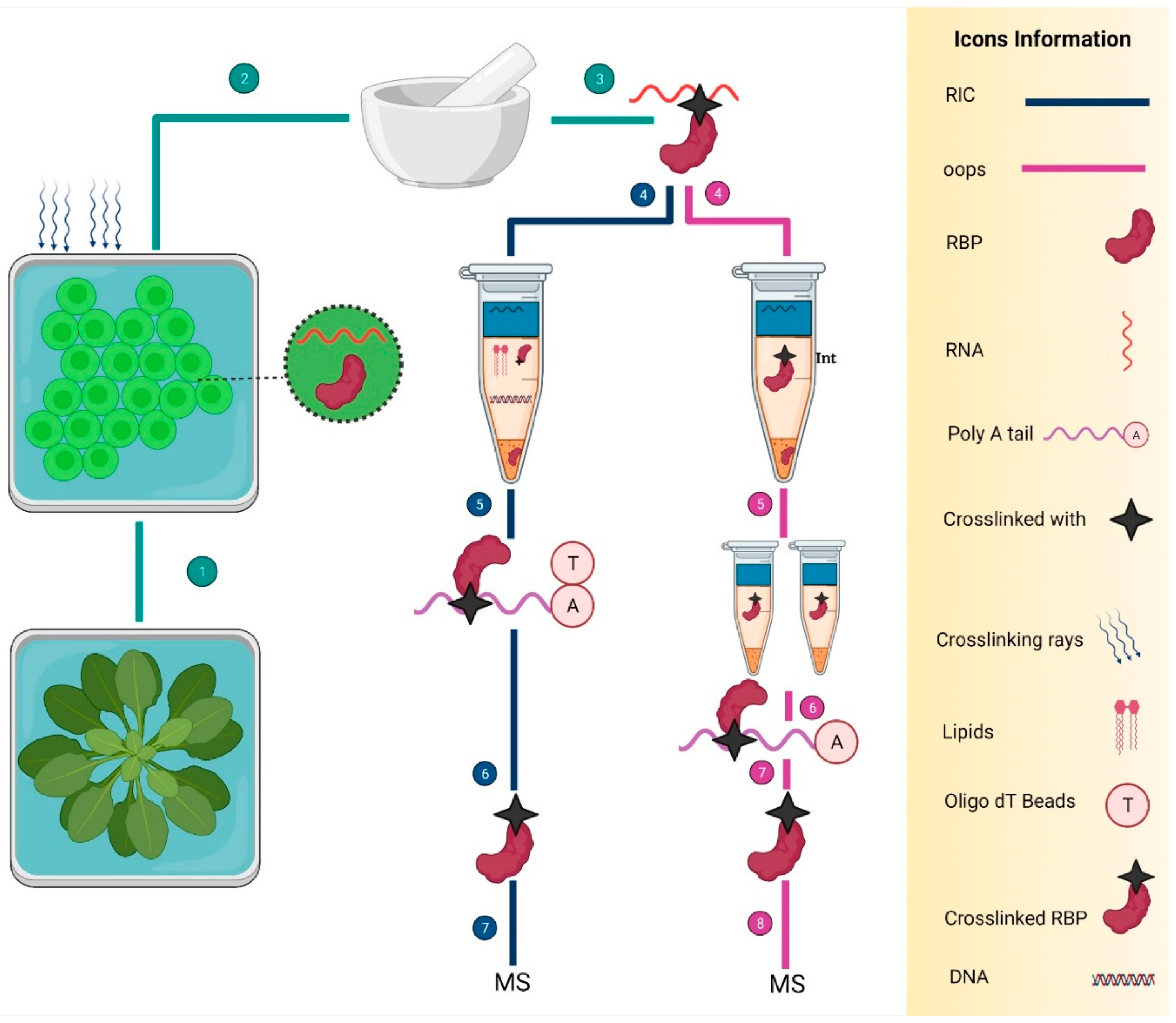
5. Identification of RBPs in Plants
6. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Castello, A.; Fischer, B.; Eichelbaum, K.; Horos, R.; Beckmann, B.M.; Strein, C.; Davey, N.E.; Humphreys, D.T.; Preiss, T.; Steinmetz, L.M.; et al. Insights into RNA Biology from an Atlas of Mammalian MRNA-Binding Proteins. Cell 2012, 149, 1393–1406. [Google Scholar] [CrossRef] [Green Version]
- Woloshen, V.; Huang, S.; Li, X. RNA-Binding Proteins in Plant Immunity. J. Pathog. 2011, 2011, 278697. [Google Scholar] [CrossRef] [Green Version]
- Lunde, B.M.; Moore, C.; Varani, G. RNA-Binding Proteins: Modular Design for Efficient Function. Nat. Rev. Mol. Cell Biol. 2007, 8, 479–490. [Google Scholar] [CrossRef] [Green Version]
- Cléry, A.; Blatter, M.; Allain, F.H.-T. RNA Recognition Motifs: Boring? Not Quite. Curr. Opin. Struct. Biol. 2008, 18, 290–298. [Google Scholar] [CrossRef]
- Valverde, R.; Edwards, L.; Regan, L. Structure and Function of KH Domains. FEBS J. 2008, 275, 2712–2726. [Google Scholar] [CrossRef]
- Linder, P.; Jankowsky, E. From Unwinding to Clamping—The DEAD Box RNA Helicase Family. Nat. Rev. Mol. Cell Biol. 2011, 12, 505–516. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Kang, H. Emerging Roles of RNA-Binding Proteins in Plant Growth, Development, and Stress Responses. Mol. Cells 2016, 39, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Stefl, R.; Skrisovska, L.; Allain, F.H.-T. RNA Sequence- and Shape-Dependent Recognition by Proteins in the Ribonucleoprotein Particle. EMBO Rep. 2005, 6, 33–38. [Google Scholar] [CrossRef]
- Weis, B.L.; Schleiff, E.; Zerges, W. Protein Targeting to Subcellular Organelles via MRNA Localization. Biochim. Biophys. Acta-Mol. Cell Res. 2013, 1833, 260–273. [Google Scholar] [CrossRef] [Green Version]
- Muleya, V.; Marondedze, C. Functional Roles of RNA-Binding Proteins in Plant Signaling. Life 2020, 10, 288. [Google Scholar] [CrossRef]
- Ramanathan, M.; Porter, D.F.; Khavari, P.A. Methods to Study RNA–Protein Interactions. Nat. Methods 2019, 16, 225–234. [Google Scholar] [CrossRef]
- Panda, A.C.; Martindale, J.L.; Gorospe, M. Affinity Pulldown of Biotinylated RNA for Detection of Protein-RNA Complexes. Bio-Protocol 2016, 6, e2062. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.; Han, M.; Meng, L.; Chen, X. Transcriptome-Wide Discovery of Coding and Noncoding RNA-Binding Proteins. Proc. Natl. Acad. Sci. USA 2018, 115, E3879–E3887. [Google Scholar] [CrossRef] [Green Version]
- McHugh, C.A.; Russell, P.; Guttman, M. Methods for Comprehensive Experimental Identification of RNA-Protein Interactions. Genome Biol. 2014, 15, 203. [Google Scholar] [CrossRef] [Green Version]
- Kishore, S.; Jaskiewicz, L.; Burger, L.; Hausser, J.; Khorshid, M.; Zavolan, M. A Quantitative Analysis of CLIP Methods for Identifying Binding Sites of RNA-Binding Proteins. Nat. Methods 2011, 8, 559–564. [Google Scholar] [CrossRef]
- Darnell, R.B. HITS-CLIP: Panoramic Views of Protein-RNA Regulation in Living Cells. Wiley Interdiscip. Rev. RNA 2010, 1, 266–286. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.-L.; Hsieh, H.-Y.; Tu, S.-L. A New Method to Identify Global Targets of RNA-Binding Proteins in Plants. bioRxiv 2021, 2021.06.11.448000. [Google Scholar] [CrossRef]
- Urdaneta, E.C.; Beckmann, B.M. Fast and Unbiased Purification of RNA-Protein Complexes after UV Cross-Lin. Methods 2020, 178, 72–82. [Google Scholar] [CrossRef]
- Panhale, A.; Richter, F.M.; Ramírez, F.; Shvedunova, M.; Manke, T.; Mittler, G.; Akhtar, A. CAPRI Enables Comparison of Evolutionarily Conserved RNA Interacting Regions. Nat. Commun. 2019, 10, 2682. [Google Scholar] [CrossRef]
- Gehring, N.H.; Wahle, E.; Fischer, U. Deciphering the MRNP Code: RNA-Bound Determinants of Post-Transcriptional Gene Regulation. Trends Biochem. Sci. 2017, 42, 369–382. [Google Scholar] [CrossRef]
- van Nues, R.; Schweikert, G.; de Leau, E.; Selega, A.; Langford, A.; Franklin, R.; Iosub, I.; Wadsworth, P.; Sanguinetti, G.; Granneman, S. Kinetic CRAC Uncovers a Role for Nab3 in Determining Gene Expression Profiles during Stress. Nat. Commun. 2017, 8, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bach-Pages, M.; Homma, F.; Kourelis, J.; Kaschani, F.; Mohammed, S.; Kaiser, M.; van der Hoorn, R.A.L.; Castello, A.; Preston, G.M. Discovering the RNA-Binding Proteome of Plant Leaves with an Improved RNA Interactome Capture Method. Biomol. 2020, 10, 661. [Google Scholar] [CrossRef] [PubMed]
- Marchese, D.; de Groot, N.S.; Lorenzo Gotor, N.; Livi, C.M.; Tartaglia, G.G. Advances in the Characterization of RNA-Binding Proteins. Wiley Interdiscip. Rev. RNA 2016, 7, 793–810. [Google Scholar] [CrossRef] [PubMed]
- Licatalosi, D.D.; Mele, A.; Fak, J.J.; Ule, J.; Kayikci, M.; Chi, S.W.; Clark, T.A.; Schweitzer, A.C.; Blume, J.E.; Wang, X.; et al. HITS-CLIP Yields Genome-Wide Insights into Brain Alternative RNA Processing. Nature 2008, 456, 464–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafner, M.; Landthaler, M.; Burger, L.; Khorshid, M.; Hausser, J.; Berninger, P.; Rothballer, A.; Ascano, M., Jr.; Jungkamp, A.-C.; Munschauer, M.; et al. Transcriptome-Wide Identification of RNA-Binding Protein and MicroRNA Target Sites by PAR-CLIP. Cell 2010, 141, 129–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konig, J.; Zarnack, K.; Rot, G.; Curk, T.; Kayikci, M.; Zupan, B.; Turner, D.J.; Luscombe, N.M.; Ule, J. ICLIP–Transcriptome-Wide Mapping of Protein-RNA Interactions with Individual Nucleotide Resolution. J. Vis. Exp. 2011, e2638. [Google Scholar] [CrossRef] [Green Version]
- Van Nostrand, E.L.; Pratt, G.A.; Shishkin, A.A.; Gelboin-Burkhart, C.; Fang, M.Y.; Sundararaman, B.; Blue, S.M.; Nguyen, T.B.; Surka, C.; Elkins, K.; et al. Robust Transcriptome-Wide Discovery of RNA-Binding Protein Binding Sites with Enhanced CLIP (ECLIP). Nat. Methods 2016, 13, 508–514. [Google Scholar] [CrossRef]
- Kargapolova, Y.; Levin, M.; Lackner, K.; Danckwardt, S. SCLIP-an Integrated Platform to Study RNA-Protein Interactomes in Biomedical Research: Identification of CSTF2tau in Alternative Processing of Small Nuclear RNAs. Nucleic Acids Res. 2017, 45, 6074–6086. [Google Scholar] [CrossRef]
- Zarnegar, B.J.; Flynn, R.A.; Shen, Y.; Do, B.T.; Chang, H.Y.; Khavari, P.A. IrCLIP Platform for Efficient Characterization of Protein-RNA Interactions. Nat. Methods 2016, 13, 489–492. [Google Scholar] [CrossRef]
- Gu, J.; Wang, M.; Yang, Y.; Qiu, D.; Zhang, Y.; Ma, J.; Zhou, Y.; Hannon, G.J.; Yu, Y. GoldCLIP: Gel-Omitted Ligation-Dependent CLIP. Genom. Proteom. Bioinform. 2018, 16, 136–143. [Google Scholar] [CrossRef]
- Kim, B.; Kim, V.N. FCLIP-Seq for Transcriptomic Footprinting of DsRNA-Binding Proteins: Lessons from DROSHA. Methods 2019, 152, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Weyn-Vanhentenryck, S.M.; Mele, A.; Yan, Q.; Sun, S.; Farny, N.; Zhang, Z.; Xue, C.; Herre, M.; Silver, P.A.; Zhang, M.Q.; et al. HITS-CLIP and Integrative Modeling Define the Rbfox Splicing-Regulatory Network Linked to Brain Development and Autism. Cell Rep. 2014, 6, 1139–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMahon, A.C.; Rahman, R.; Jin, H.; Shen, J.L.; Fieldsend, A.; Luo, W.; Rosbash, M. TRIBE: Hijacking an RNA-Editing Enzyme to Identify Cell-Specific Targets of RNA-Binding Proteins. Cell 2016, 165, 742–753. [Google Scholar] [CrossRef] [Green Version]
- Granneman, S.; Kudla, G.; Petfalski, E.; Tollervey, D. Identification of Protein Binding Sites on U3 SnoRNA and Pre-RRNA by UV Cross-Linking and High-Throughput Analysis of CDNAs. Proc. Natl. Acad. Sci. USA 2009, 106, 9613–9618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapointe, C.P.; Wilinski, D.; Saunders, H.A.J.; Wickens, M. Protein-RNA Networks Revealed through Covalent RNA Marks. Nat. Methods 2015, 12, 1163–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.I.; Birendra, K.C.; Zhu, W.; Motamedchaboki, K.; Doye, V.; Roux, K.J. Probing Nuclear Pore Complex Architecture with Proximity-Dependent Biotinylation. Proc. Natl. Acad. Sci. USA 2014, 111, E2453–E2461. [Google Scholar] [CrossRef] [Green Version]
- Lewinski, M.; Köster, T. Systems Approaches to Map In Vivo RNA–Protein Interactions in Arabidopsis Thaliana BT—Systems Biology. In System Biology; Rajewsky, N., Jurga, S., Barciszewski, J., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 77–95. ISBN 978-3-319-92967-5. [Google Scholar]
- Kaewsapsak, P.; Shechner, D.M.; Mallard, W.; Rinn, J.L.; Ting, A.Y. Live-Cell Mapping of Organelle-Associated RNAs via Proximity Biotinylation Combined with Protein-RNA Crosslinking. Elife 2017, 6, e29224. [Google Scholar] [CrossRef]
- Faoro, C.; Ataide, S.F. Ribonomic Approaches to Study the RNA-Binding Proteome. FEBS Lett. 2014, 588, 3649–3664. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Cho, S.; Moon, H.; Loh, T.J.; Jang, H.N.; Shen, H. Detecting RNA-Protein Interaction Using End-Labeled Biotinylated RNA Oligonucleotides and Immunoblotting. In Methods in Molecular Biology; Humana: New York, NY, USA, 2016; Volume 1421, pp. 35–44. [Google Scholar]
- Walker, S.C.; Good, P.D.; Gipson, T.A.; Engelke, D.R. The Dual Use of RNA Aptamer Sequences for Affinity Purification and Localization Studies of RNAs and RNA-Protein Complexes. Methods Mol. Biol. 2011, 714, 423–444. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.Y.; Haurwitz, R.E.; Apffel, A.; Zhou, K.; Smart, B.; Wenger, C.D.; Laderman, S.; Bruhn, L.; Doudna, J.A. RNA–Protein Analysis Using a Conditional CRISPR Nuclease. Proc. Natl. Acad. Sci. USA 2013, 110, 5416–5421. [Google Scholar] [CrossRef] [Green Version]
- Poria, D.K.; Ray, P.S. RNA-Protein UV-Crosslinking Assay. Bio-Protocol 2017, 7, e2193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walton, S.P.; Stephanopoulos, G.N.; Yarmush, M.L.; Roth, C.M. Thermodynamic and Kinetic Characterization of Antisense Oligodeoxynucleotide Binding to a Structured MRNA. Biophys. J. 2002, 82, 366–377. [Google Scholar] [CrossRef] [Green Version]
- Gräwe, C.; Stelloo, S.; van Hout, F.A.H.; Vermeulen, M. RNA-Centric Methods: Toward the Interactome of Specific RNA Transcripts. Trends Biotechnol. 2021, 39, 890–900. [Google Scholar] [CrossRef] [PubMed]
- Dasti, A.; Cid-Samper, F.; Bechara, E.; Tartaglia, G.G. RNA-Centric Approaches to Study RNA-Protein Interactions in Vitro and in Silico. Methods 2020, 178, 11–18. [Google Scholar] [CrossRef]
- Sutherland, B.W.; Toews, J.; Kast, J. Utility of Formaldehyde Cross-Linking and Mass Spectrometry in the Study of Protein–Protein Interactions. J. Mass Spectrom. 2008, 43, 699–715. [Google Scholar] [CrossRef]
- Li, X.; Song, J.; Yi, C. Genome-Wide Mapping of Cellular Protein-RNA Interactions Enabled by Chemical Crosslinking. Genom. Proteom. Bioinform. 2014, 12, 72–78. [Google Scholar] [CrossRef] [Green Version]
- Marondedze, C. The Increasing Diversity and Complexity of the RNA-Binding Protein Repertoire in Plants. Proc. R. Soc. B 2020, 287, 20201397. [Google Scholar] [CrossRef]
- Roux, K.J.; Kim, D.I.; Raida, M.; Burke, B. A Promiscuous Biotin Ligase Fusion Protein Identifies Proximal and Interacting Proteins in Mammalian Cells. J. Cell Biol. 2012, 196, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Trendel, J.; Schwarzl, T.; Horos, R.; Prakash, A.; Bateman, A.; Hentze, M.W.; Krijgsveld, J. The Human RNA-Binding Proteome and Its Dynamics during Translational Arrest. Cell 2019, 176, 391–403.e19. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, E.A.; Frey, B.L.; Smith, L.M.; Auble, D.T. Formaldehyde Crosslinking: A Tool for the Study of Chromatin Complexes. J. Biol. Chem. 2015, 290, 26404–26411. [Google Scholar] [CrossRef] [Green Version]
- Chu, C.; Qu, K.; Zhong, F.L.; Artandi, S.E.; Chang, H.Y. Genomic Maps of Long Noncoding RNA Occupancy Reveal Principles of RNA-Chromatin Interactions. Mol. Cell 2011, 44, 667–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McHugh, C.A.; Chen, C.-K.; Chow, A.; Surka, C.F.; Tran, C.; McDonel, P.; Pandya-Jones, A.; Blanco, M.; Burghard, C.; Moradian, A.; et al. The Xist LncRNA Interacts Directly with SHARP to Silence Transcription through HDAC3. Nature 2015, 521, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, M.; Majzoub, K.; Rao, D.S.; Neela, P.H.; Zarnegar, B.J.; Mondal, S.; Roth, J.G.; Gai, H.; Kovalski, J.R.; Siprashvili, Z.; et al. RNA-Protein Interaction Detection in Living Cells. Nat. Methods 2018, 15, 207–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Boonen, K.; Ferrari, P.; Schoofs, L.; Janssens, E.; van Noort, V.; Rolland, F.; Geuten, K. UV Crosslinked MRNA-Binding Proteins Captured from Leaf Mesophyll Protoplasts. Plant Methods 2016, 12, 42. [Google Scholar] [CrossRef] [Green Version]
- Köster, T.; Reichel, M.; Staiger, D. CLIP and RNA Interactome Studies to Unravel Genome-Wide RNA-Protein Interactions in Vivo in Arabidopsis Thaliana. Methods 2020, 178, 63–71. [Google Scholar] [CrossRef]
- Köster, T.; Marondedze, C.; Meyer, K.; Staiger, D. RNA-Binding Proteins Revisited; The Emerging Arabidopsis MRNA Interactome. Trends Plant Sci. 2017, 22, 512–526. [Google Scholar] [CrossRef]
- Burjoski, V.; Reddy, A.S.N. The Landscape of RNA-Protein Interactions in Plants: Approaches and Current Status. Int. J. Mol. Sci. 2021, 22, 2845. [Google Scholar] [CrossRef]
- Marondedze, C.; Thomas, L.; Gehring, C.; Lilley, K.S. Changes in the Arabidopsis RNA-Binding Proteome Reveal Novel Stress Response Mechanisms. BMC Plant Biol. 2019, 19, 1–11. [Google Scholar] [CrossRef]
- Paz, I.; Kligun, E.; Bengad, B.; Mandel-Gutfreund, Y. BindUP: A Web Server for Non-Homology-Based Prediction of DNA and RNA Binding Proteins. Nucleic Acids Res. 2016, 44, W568–W574. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zhan, J.; Zhao, H.; Zhou, Y. A New Size-Independent Score for Pairwise Protein Structure Alignment and Its Application to Structure Classification and Nucleic-Acid Binding Prediction. Proteins 2012, 80, 2080–2088. [Google Scholar] [CrossRef] [Green Version]
- Si, J.; Cui, J.; Cheng, J.; Wu, R. Computational Prediction of RNA-Binding Proteins and Binding Sites. Int. J. Mol. Sci. 2015, 16, 26303–26317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, A.; Khanal, R.; Hoque, M.T. AIRBP: Accurate Identification of RNA-Binding Proteins Using Machine Learning Techniques. bioRxiv 2020. [Google Scholar] [CrossRef] [PubMed]
- Marondedze, C.; Thomas, L.; Serrano, N.L.; Lilley, K.S.; Gehring, C. The RNA-Binding Protein Repertoire of Arabidopsis Thaliana. Sci. Rep. 2016, 6, 29766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muthusamy, M.; Kim, J.-H.; Kim, J.A.; Lee, S.-I. Plant RNA Binding Proteins as Critical Modulators in Drought, High Salinity, Heat, and Cold Stress Responses: An Updated Overview. Int. J. Mol. Sci. 2021, 22, 6731. [Google Scholar] [CrossRef] [PubMed]
- Urdaneta, E.C.; Vieira-Vieira, C.H.; Hick, T.; Wessels, H.-H.; Figini, D.; Moschall, R.; Medenbach, J.; Ohler, U.; Granneman, S.; Selbach, M.; et al. Purification of Cross-Linked RNA-Protein Complexes by Phenol-Toluol Extraction. Nat. Commun. 2019, 10, 990. [Google Scholar] [CrossRef] [PubMed]
- Leppek, K.; Stoecklin, G. An Optimized Streptavidin-Binding RNA Aptamer for Purification of Ribonucleoprotein Complexes Identifies Novel ARE-Binding Proteins. Nucleic Acids Res. 2014, 42, e13. [Google Scholar] [CrossRef] [Green Version]
- Van Ende, R.; Balzarini, S.; Geuten, K. Single and Combined Methods to Specifically or Bulk-Purify RNA–Protein Complexes. Biomolecules 2020, 10, 1160. [Google Scholar] [CrossRef]
- Smith, J.M.; Sandow, J.J.; Webb, A.I. The Search for RNA-Binding Proteins: A Technical and Interdisciplinary Challenge. Biochem. Soc. Trans. 2021, 49, 393–403. [Google Scholar] [CrossRef]
- Queiroz, R.M.L.; Smith, T.; Villanueva, E.; Marti-Solano, M.; Monti, M.; Pizzinga, M.; Mirea, D.M.; Ramakrishna, M.; Harvey, R.F.; Dezi, V.; et al. Comprehensive Identification of RNA–Protein Interactions in Any Organism Using Orthogonal Organic Phase Separation (OOPS). Nat. Biotechnol. 2019, 37, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Villanueva, E.; Smith, T.; Queiroz, R.M.L.; Monti, M.; Pizzinga, M.; Elzek, M.; Dezi, V.; Harvey, R.F.; Ramakrishna, M.; Willis, A.E.; et al. Efficient Recovery of the RNA-Bound Proteome and Protein-Bound Transcriptome Using Phase Separation (OOPS). Nat. Protoc. 2020, 15, 2568–2588. [Google Scholar] [CrossRef]
- Smith, T.; Villanueva, E.; Queiroz, R.M.L.; Dawson, C.S.; Elzek, M.; Urdaneta, E.C.; Willis, A.E.; Beckmann, B.M.; Krijgsveld, J.; Lilley, K.S. Organic Phase Separation Opens up New Opportunities to Interrogate the RNA-Binding Proteome. Curr. Opin. Chem. Biol. 2020, 54, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Haroon, M.; Wang, X.; Afzal, R.; Zafar, M.M.; Idrees, F.; Batool, M.; Khan, A.S.; Imran, M. Novel Plant Breeding Techniques Shake Hands with Cereals to Increase Production. Plants 2022, 11, 1052. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.H.; Kwak, K.J.; Kim, M.K.; Park, S.J.; Yang, K.-Y.; Kang, H. Expression of Arabidopsis Glycine-Rich RNA-Binding Protein AtGRP2 or AtGRP7 Improves Grain Yield of Rice (Oryza Sativa) under Drought Stress Conditions. Plant Sci. 2014, 214, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Foley, S.W.; Gosai, S.J.; Wang, D.; Selamoglu, N.; Sollitti, A.C.; Köster, T.; Steffen, A.; Lyons, E.; Daldal, F.; Garcia, B.A. A Global View of RNA-Protein Interactions Identifies Post-Transcriptional Regulators of Root Hair Cell Fate. Dev. Cell 2017, 41, 204–220. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Wang, R.; Liang, D.; Ma, F.; Shu, H. Molecular Characterization and Expression Analysis of a Glycine-Rich RNA-Binding Protein Gene from Malus Hupehensis Rehd. Mol. Biol. Rep. 2012, 39, 4145–4153. [Google Scholar] [CrossRef] [PubMed]
- Nemali, K.S.; Bonin, C.; Dohleman, F.G.; Stephens, M.; Reeves, W.R.; Nelson, D.E.; Castiglioni, P.; Whitsel, J.E.; Sammons, B.; Silady, R.A. Physiological Responses Related to Increased Grain Yield under Drought in the First Biotechnology-derived Drought-tolerant Maize. Plant. Cell Environ. 2015, 38, 1866–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambrosone, A.; Batelli, G.; Nurcato, R.; Aurilia, V.; Punzo, P.; Bangarusamy, D.K.; Ruberti, I.; Sassi, M.; Leone, A.; Costa, A. The Arabidopsis RNA-Binding Protein AtRGGA Regulates Tolerance to Salt and Drought Stress. Plant Physiol. 2015, 168, 292–306. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Guo, T.; Wang, P.; Sun, X.; Shao, Y.; Jia, X.; Liang, B.; Gong, X.; Ma, F. MhYTP1 and MhYTP2 from Apple Confer Tolerance to Multiple Abiotic Stresses in Arabidopsis Thaliana. Front. Plant Sci. 2017, 8, 1367. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhang, C.; Jia, X.; Wang, W.; Yin, H. Comparative Analysis of RNA-Binding Proteomes under Arabidopsis Thaliana-Pst DC3000-PAMP Interaction by Orthogonal Organic Phase Separation. Int. J. Biol. Macromol. 2020, 160, 47–54. [Google Scholar] [CrossRef]
- Reichel, M.; Liao, Y.; Rettel, M.; Ragan, C.; Evers, M.; Alleaume, A.-M.; Horos, R.; Hentze, M.W.; Preiss, T.; Millar, A.A. In Planta Determination of the MRNA-Binding Proteome of Arabidopsis Etiolated Seedlings. Plant Cell 2016, 28, 2435–2452. [Google Scholar] [CrossRef] [Green Version]
- Xing, D.; Wang, Y.; Hamilton, M.; Ben-Hur, A.; Reddy, A.S.N. Transcriptome-Wide Identification of RNA Targets of Arabidopsis SERINE/ARGININE-RICH45 Uncovers the Unexpected Roles of This RNA Binding Protein in RNA Processing. Plant Cell 2015, 27, 3294–3308. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gu, L.; Hou, Y.; Wang, L.; Deng, X.; Hang, R.; Chen, D.; Zhang, X.; Zhang, Y.; Liu, C.; et al. Integrative Genome-Wide Analysis Reveals HLP1, a Novel RNA-Binding Protein, Regulates Plant Flowering by Targeting Alternative Polyadenylation. Cell Res. 2015, 25, 864–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
| Method | Purpose | Plant Specie | Identified RBPs/RNAs | Ref |
|---|---|---|---|---|
| RIC | Discovery of the RNA-binding proteome of plant leaves with an improved RNA interactome capture method | Arabidopsis | 717 | [22] |
| RIC | Determination of the mRNA-binding proteome of Arabidopsis etiolated seedlings | Arabidopsis | 700 | [82] |
| RIC | It was used in cells from different ecotypes grown in cultures and leaves to find the RBPs | Arabidopsis | 1145 | [65] |
| RIC | To capture the mRNA interactome from plant protoplasts | Arabidopsis | 325 | [56] |
| PtRIC | To check the change in RBPs in response to environmental cues | Arabidopsis | 717 | [22] |
| RIC | To check the effect of severe drought stress on the RNA-binding proteome | Arabidopsis | 1408 | [60] |
| RIP-seq | To identify the RNAs bound with the protein of interest | Arabidopsis | 4000 | [65,83] |
| iCLIP-seq | To unravel genome-wide RNA–protein interactions in vivo and the landscape of AtGRP7 | Arabidopsis | [14,17] | |
| HITS-CLIP | To study the function of RBP (HLP1) in flowering by targeting alternative polyadenylation | Arabidopsis | [84] | |
| OOPS | To find all RBPs in plant extracts (both coding and non-coding) | Arabidopsis | 468 | [81] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haroon, M.; Afzal, R.; Zafar, M.M.; Zhang, H.; Li, L. Ribonomics Approaches to Identify RBPome in Plants and Other Eukaryotes: Current Progress and Future Prospects. Int. J. Mol. Sci. 2022, 23, 5923. https://doi.org/10.3390/ijms23115923
Haroon M, Afzal R, Zafar MM, Zhang H, Li L. Ribonomics Approaches to Identify RBPome in Plants and Other Eukaryotes: Current Progress and Future Prospects. International Journal of Molecular Sciences. 2022; 23(11):5923. https://doi.org/10.3390/ijms23115923
Chicago/Turabian StyleHaroon, Muhammad, Rabail Afzal, Muhammad Mubashar Zafar, Hongwei Zhang, and Lin Li. 2022. "Ribonomics Approaches to Identify RBPome in Plants and Other Eukaryotes: Current Progress and Future Prospects" International Journal of Molecular Sciences 23, no. 11: 5923. https://doi.org/10.3390/ijms23115923
APA StyleHaroon, M., Afzal, R., Zafar, M. M., Zhang, H., & Li, L. (2022). Ribonomics Approaches to Identify RBPome in Plants and Other Eukaryotes: Current Progress and Future Prospects. International Journal of Molecular Sciences, 23(11), 5923. https://doi.org/10.3390/ijms23115923