
Identification of a Steric Zipper Motif in the Amyloidogenic Core of Human Cystatin C and Its Use for the Design of Self-Assembling Peptides

 , , and
, , and
Abstract
1. Introduction
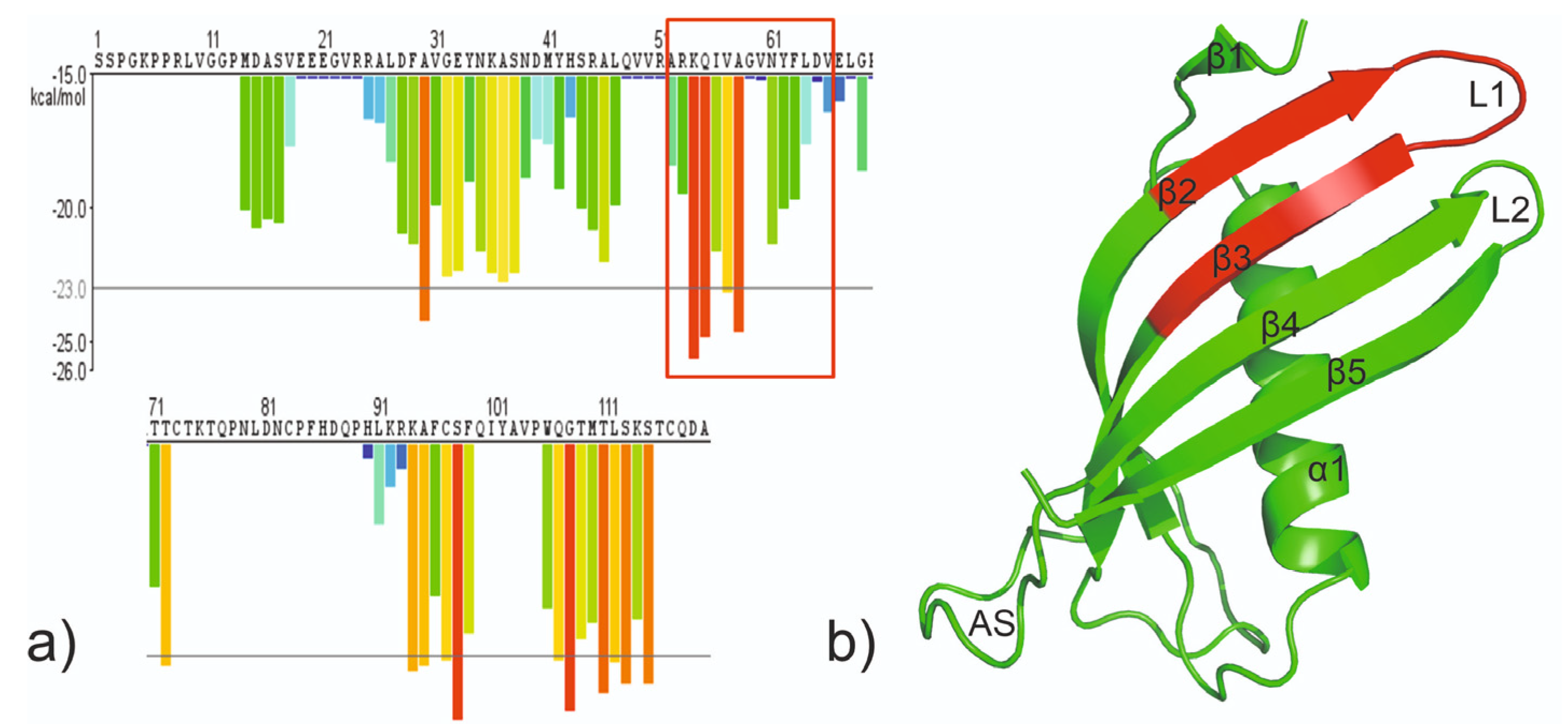
2. Results
2.1. Peptide Design, Synthesis and Purification
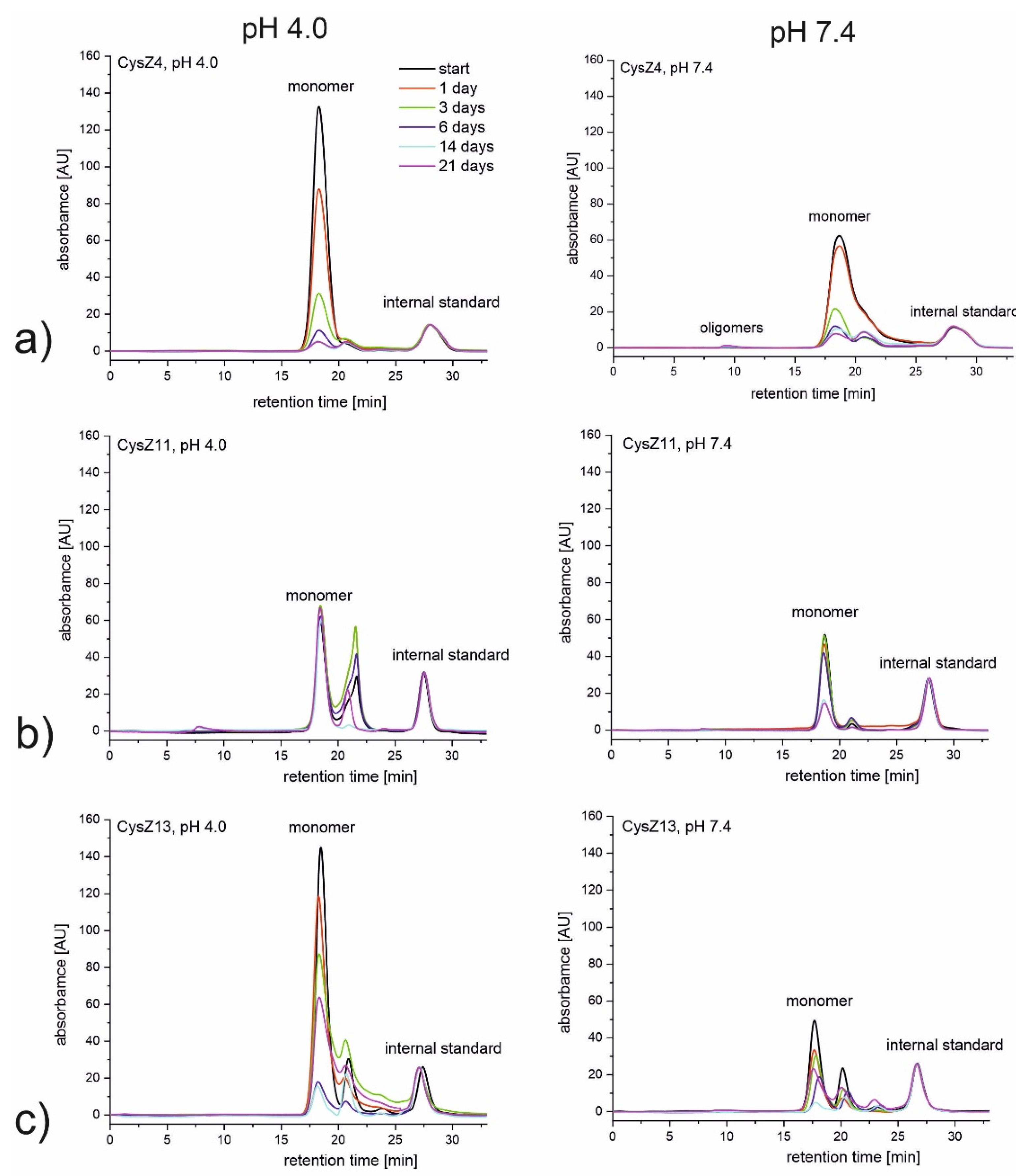
2.2. Size Exclusion Chromatography (SEC)
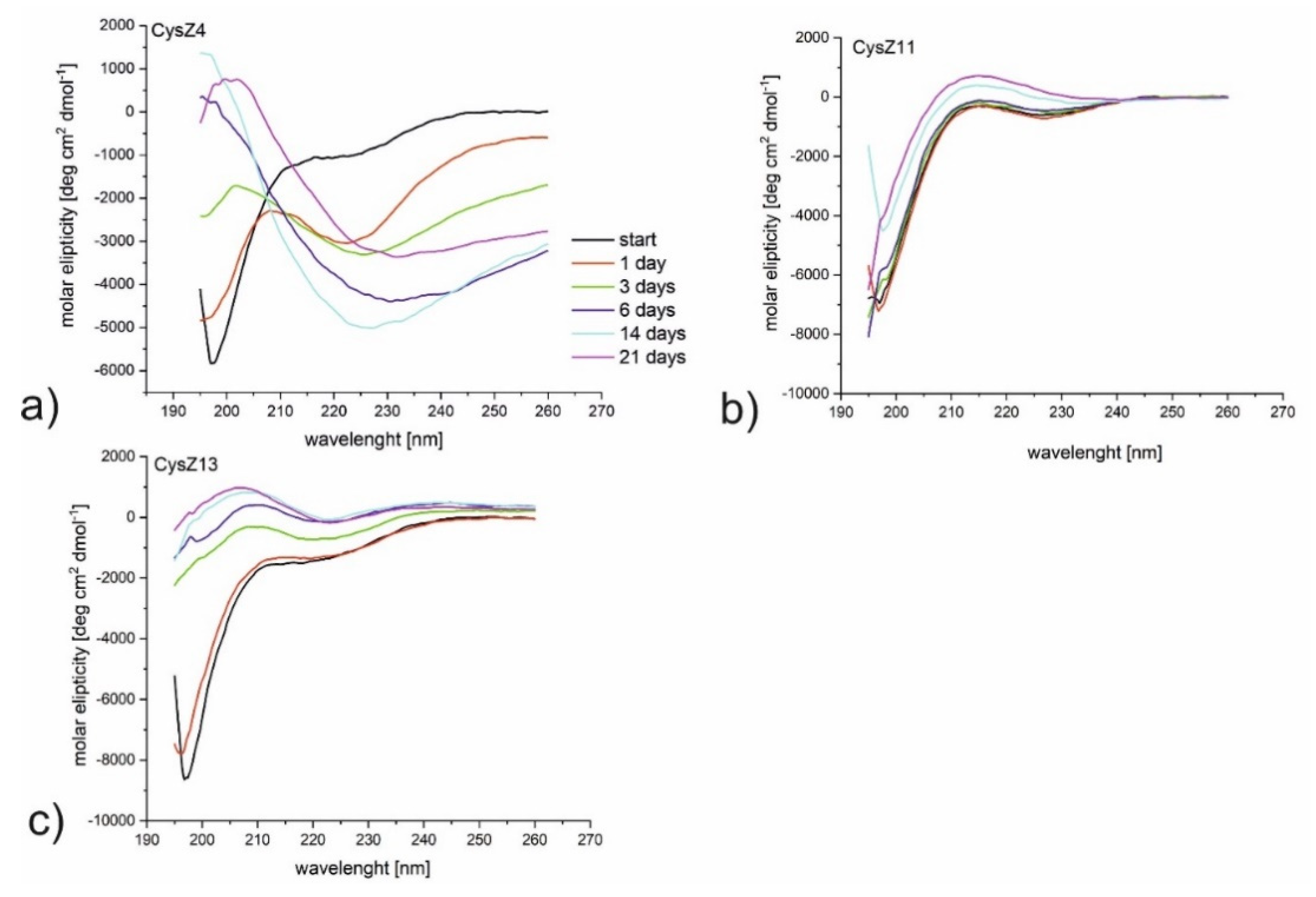
2.3. Circular Dichroism Measurements
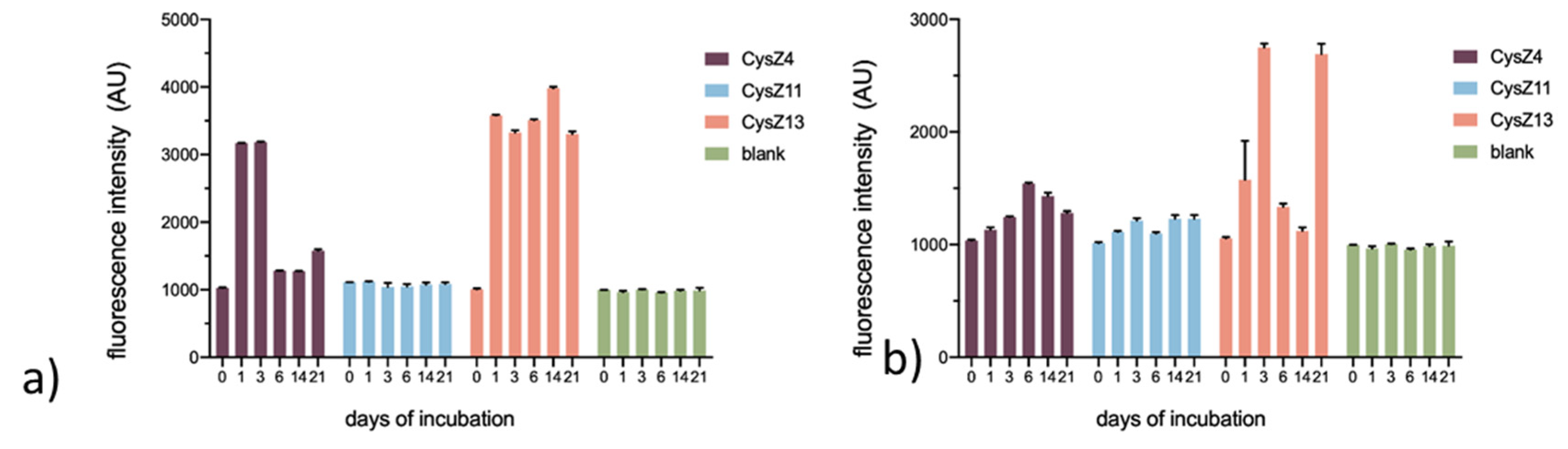
2.4. Thioflavin T Assay
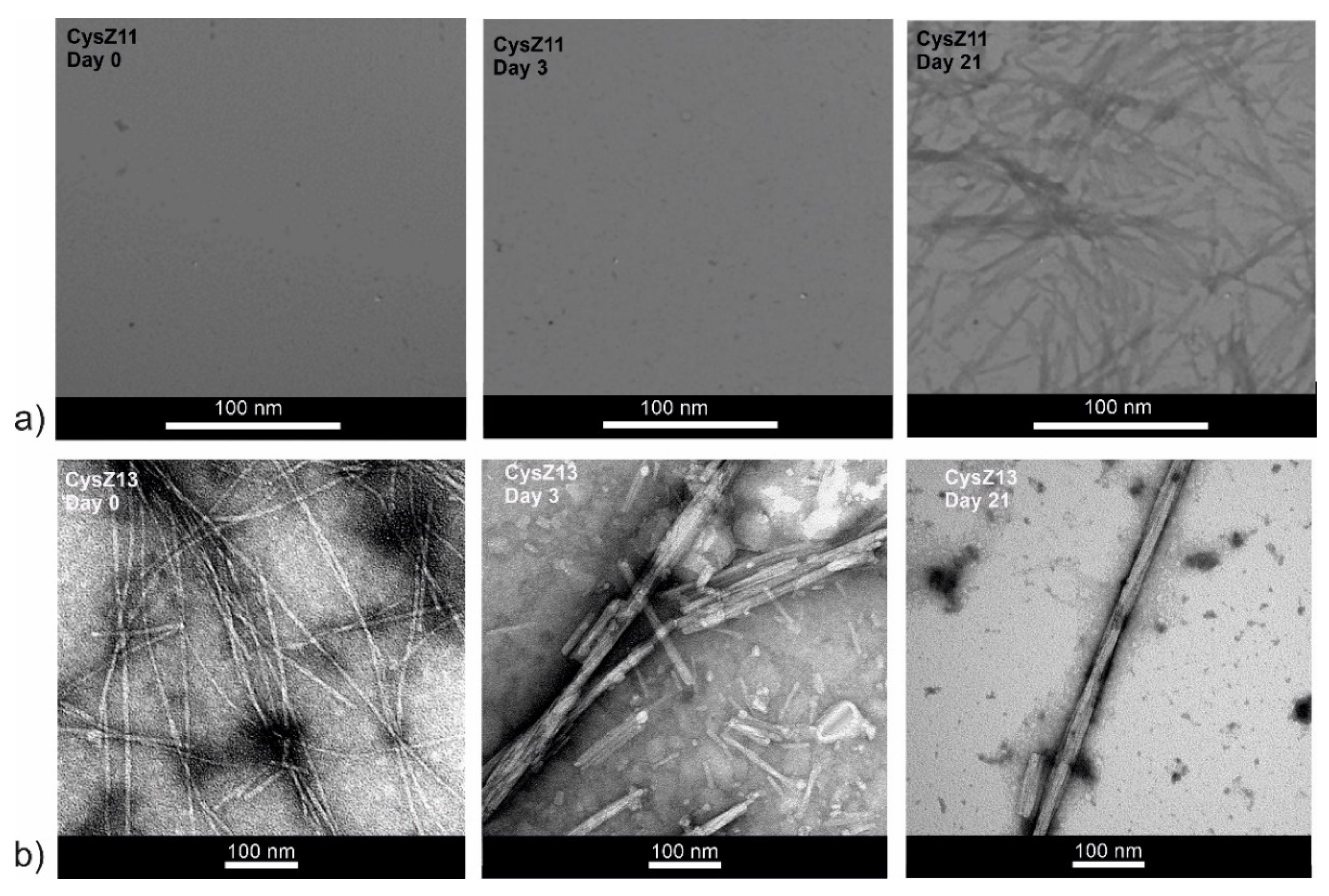
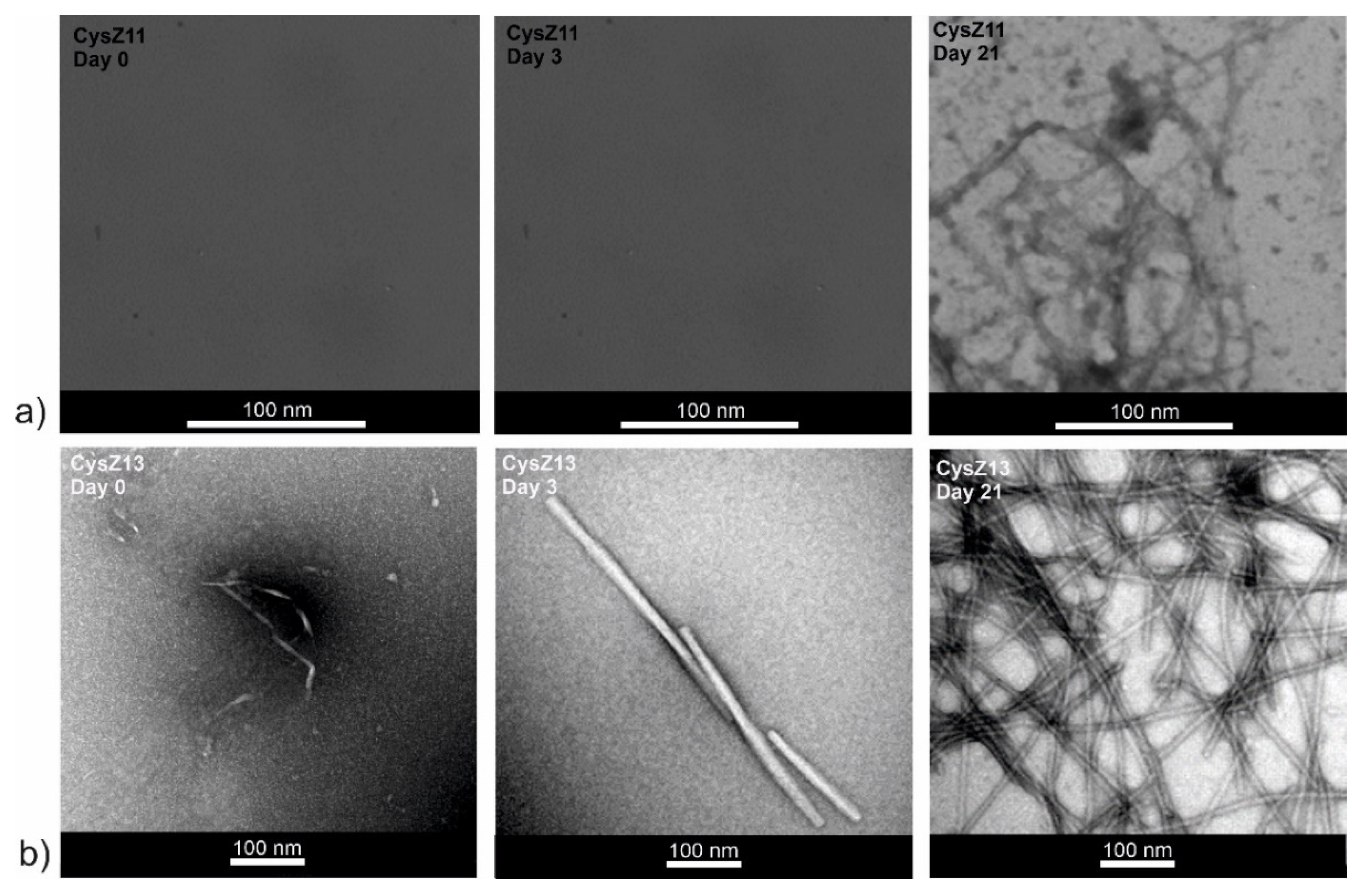
2.5. Transmission Electron Microscopy
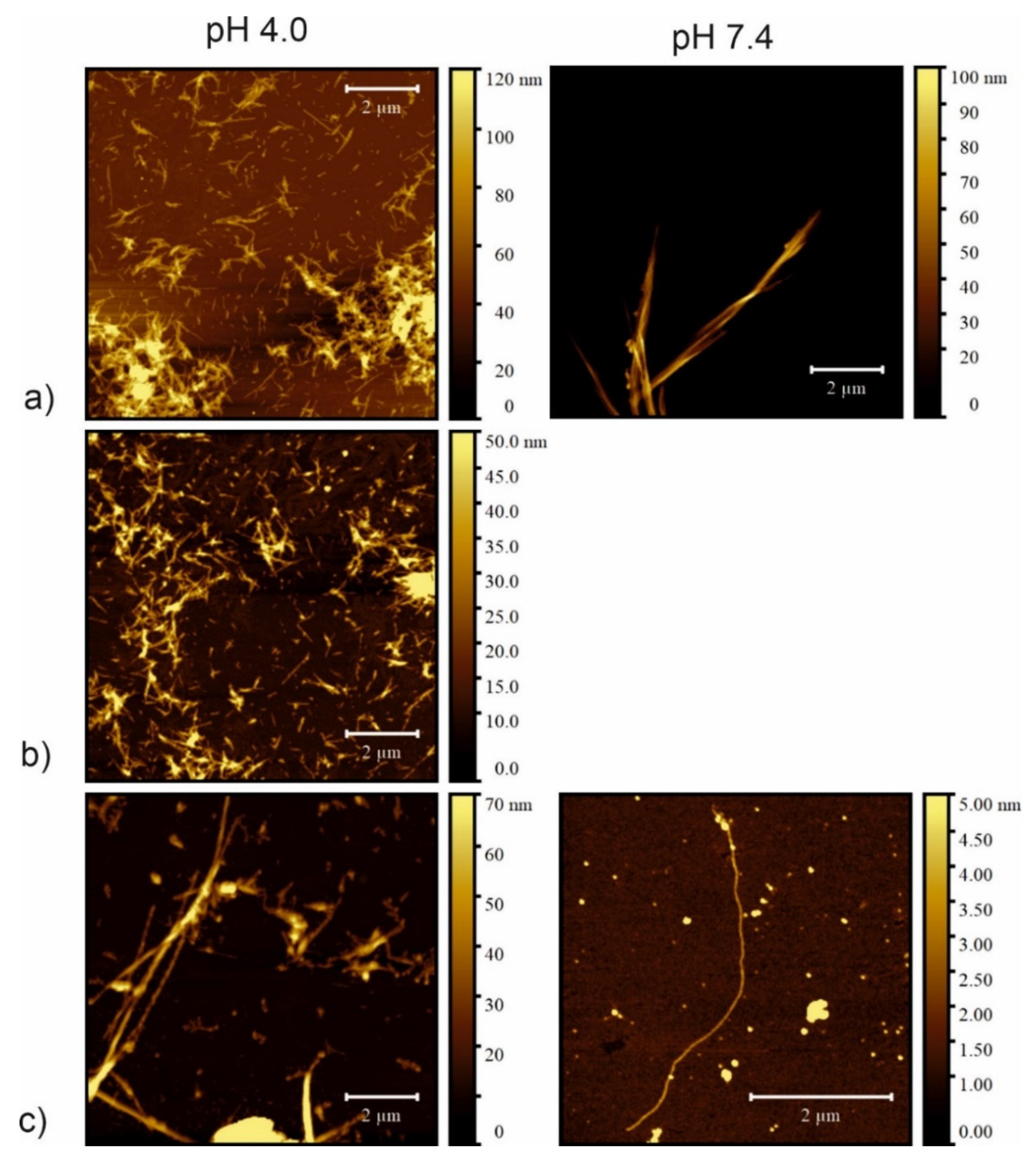
2.6. Atomic Force Microscopy
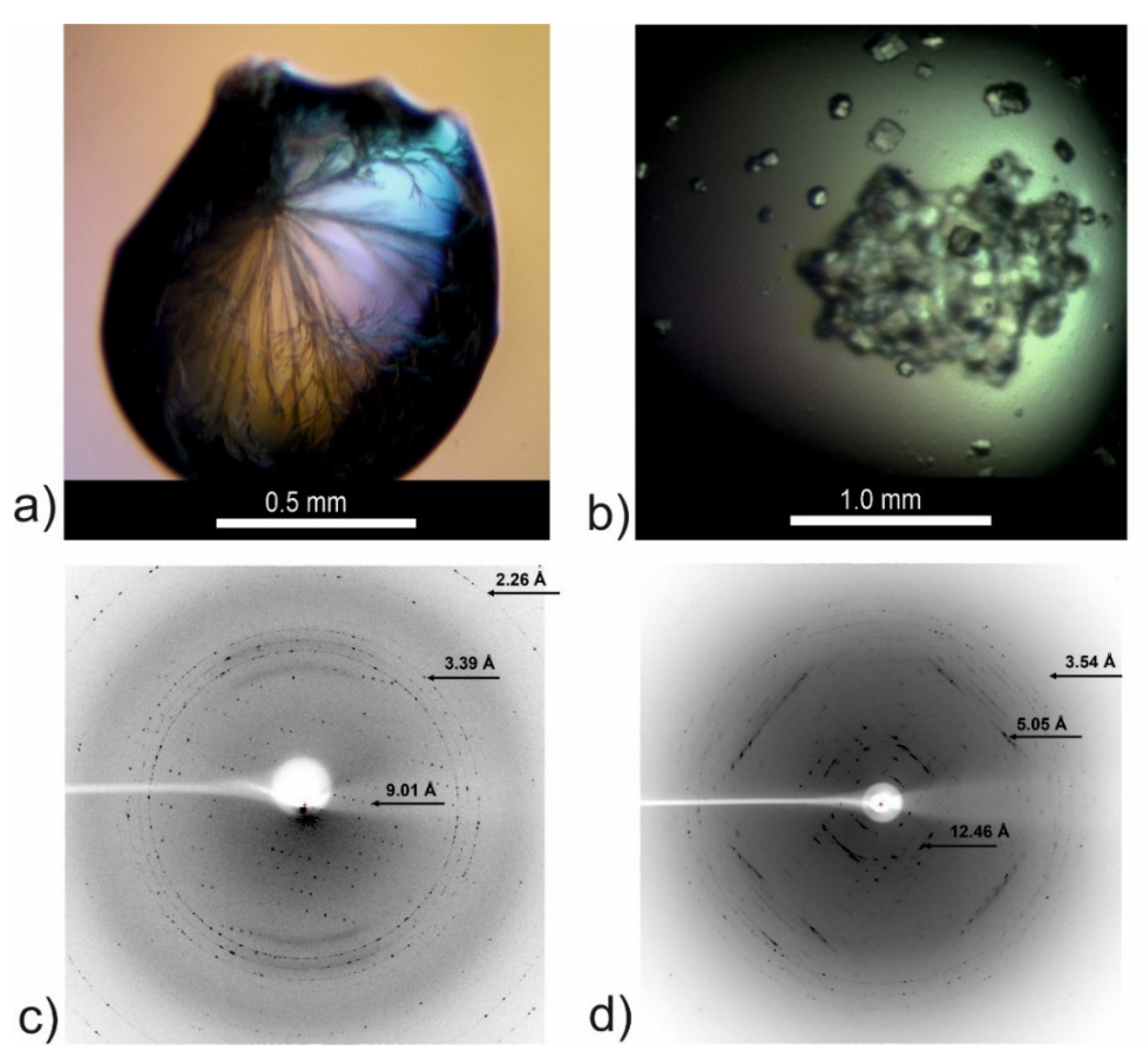
2.7. Attempts to Determine the Crystallographic Structure
2.8. X-Ray Fiber Diffraction
3. Discussion
4. Materials and Methods
4.1. Peptide Synthesis
4.2. Fibril Preparation
4.3. Size Exclusion Chromatography
4.4. Circular Dichroism
4.5. Thioflavin T Fluorescence Assay
4.6. Transmission Electron Microscopy
4.7. Atomic Force Microscopy
4.8. Crystallization and X-ray Diffraction
4.9. X-ray Fiber Diffraction
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schmidt, A.; Annamalai, K.; Schmidt, M.; Grigorieff, N.; Fändrich, M. Cryo-EM reveals the steric zipper structure of a light chain-derived amyloid fibril. Proc. Natl. Acad. Sci. USA 2016, 113, 6200–6205. [Google Scholar] [CrossRef]
- Bueno, J.; Demirci, F.; Baser, K.H.C. Antimicrobial Strategies in Novel Drug Delivery Systems, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; Volume 2, ISBN 9780128110799. [Google Scholar]
- Al-Halifa, S.; Babych, M.; Zottig, X.; Archambault, D.; Bourgault, S. Amyloid self-assembling peptides: Potential applications in nanovaccine engineering and biosensing. Pept. Sci. 2019, 111, e24095. [Google Scholar] [CrossRef]
- Wei, G.; Su, Z.; Reynolds, N.P.; Arosio, P.; Hamley, I.W.; Gazit, E.; Mezzenga, R. Self-assembling peptide and protein amyloids: From structure to tailored function in nanotechnology. Chem. Soc. Rev. 2017, 46, 4661–4708. [Google Scholar] [CrossRef]
- Cherny, I.; Gazit, E. Amyloids: Not only pathological agents but also ordered nanomaterials. Angew. Chem. Int. Ed. Engl. 2008, 47, 4062–4069. [Google Scholar] [CrossRef]
- Gazit, E. Controlling molecular self-assembly: From amyloid oligomerization and therapy to novel biomaterials and technological applications in nanomedicine. Nanomedicine 2014, 9, 2433–2436. [Google Scholar] [CrossRef]
- Knowles, T.P.J.; White, D.A.; Abate, A.R.; Agresti, J.J.; Cohen, S.I.A.; Sperling, R.A.; De Genst, E.J.; Dobson, C.M.; Weitz, D.A. Observation of spatial propagation of amyloid assembly from single nuclei. Proc. Natl. Acad. Sci. USA 2011, 108, 14746–14751. [Google Scholar] [CrossRef]
- Nelson, R.; Sawaya, M.R.; Balbirnie, M.; Madsen, A.Ø.; Riekel, C.; Grothe, R.; Eisenberg, D. Structure of the cross-β spine of amyloid-like fibrils. Nature 2005, 435, 773–778. [Google Scholar] [CrossRef]
- Esposito, L.; Pedone, C.; Vitagliano, L. Molecular dynamics analyses of cross-beta-spine steric zipper models: Beta-sheet twisting and aggregation. Proc. Natl. Acad. Sci. USA 2006, 103, 11533–11538. [Google Scholar] [CrossRef]
- Sambashivan, S.; Liu, Y.; Sawaya, M.R.; Gingery, M.; Eisenberg, D. Amyloid-like fibrils of ribonuclease A with three-dimensional domain-swapped and native-like structure. Nature 2005, 437, 266–269. [Google Scholar] [CrossRef]
- Goldschmidt, L.; Teng, P.K.; Riek, R.; Eisenberg, D. Identifying the amylome, proteins capable of forming amyloid-like fibrils. Proc. Natl. Acad. Sci. USA 2010, 107, 3487–3492. [Google Scholar] [CrossRef]
- Grubb, A. Cystatin C—Properties and use as diagnositic marker. Adv. Clin. Chem. 2000, 35, 63–69. [Google Scholar]
- Ólafsson, Í.; Grubb, A. Hereditary cystatin C amyloid angiopathy. Amyloid 2000, 7, 70–79. [Google Scholar] [CrossRef]
- Wahlbom, M.; Wang, X.; Lindström, V.; Carlemalm, E.; Jaskolski, M.; Grubb, A. Fibrillogenic Oligomers of Human Cystatin C Are Formed by Propagated Domain Swapping. J. Biol. Chem. 2007, 282, 18318–18326. [Google Scholar] [CrossRef]
- Nilsson, M.; Wang, X.; Rodziewicz-Motowidlo, S.; Janowski, R.; Lindström, V.; Önnerfjord, P.; Westermark, G.; Grzonka, Z.; Jaskolski, M.; Grubb, A. Prevention of domain swapping inhibits dimerization and amyloid fibril formation of cystatin C. Use of engineered disulfide bridges, antibodies, and carboxymethylpapain to stabilize the monomeric form of cystatin C. J. Biol. Chem. 2004, 279, 24236–24245. [Google Scholar] [CrossRef]
- Janowski, R.; Kozak, M.; Jankowska, E.; Grzonka, Z.; Grubb, A.; Abrahamson, M.; Jaskolski, M. Human cystatin C, an amyloidogenic protein, dimerizes through three-dimensional domain swapping. Nat. Struct. Biol. 2001, 8, 316–320. [Google Scholar] [CrossRef][Green Version]
- Kolodziejczyk, R.; Michalska, K.; Hernandez-Santoyo, A.; Wahlbom, M.; Grubb, A.; Jaskolski, M. Crystal structure of human cystatin C stabilized against amyloid formation. FEBS J. 2010, 277, 1726–1737. [Google Scholar] [CrossRef]
- Orlikowska, M.; Jankowska, E.; Borek, D.; Otwinowski, Z.; Skowron, P.; Szymaska, A. Crystallization and preliminary X-ray diffraction analysis of Val57 mutants of the amyloidogenic protein human cystatin C. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2011, 67, 1608–1611. [Google Scholar] [CrossRef]
- Maszota-Zieleniak, M.; Jurczak, P.; Orlikowska, M.; Zhukov, I.; Borek, D.; Otwinowski, Z.; Skowron, P.; Pietralik, Z.; Kozak, M.; Szymańska, A.; et al. NMR and crystallographic structural studies of the extremely stable monomeric variant of human cystatin C with single amino acid substitution. FEBS J. 2020, 287, 361–376. [Google Scholar] [CrossRef]
- Alvarez-Fernandez, M.; Barrett, A.J.; Gerhartz, B.; Dando, P.M.; Ni, J.; Abrahamson, M. Inhibition of mammalian legumain by some cystatins is due to a novel second reactive site. J. Biol. Chem. 1999, 274, 19195–19203. [Google Scholar] [CrossRef]
- Bennett, M.J.; Schlunegger, M.P.; Eisenberg, D. 3D domain swapping: A mechanism for oligomer assembly. Protein Sci. 1995, 4, 2455–2468. [Google Scholar] [CrossRef]
- Jaskólski, M. 3D domain swapping, protein oligomerization, and amyloid formation. Acta Biochim. Pol. 2001, 48, 807–827. [Google Scholar] [CrossRef]
- Nelson, R.; Eisenberg, D. Recent atomic models of amyloid fibril structure. Curr. Opin. Struct. Biol. 2006, 16, 260–265. [Google Scholar] [CrossRef]
- Sawaya, M.R.; Sambashivan, S.; Nelson, R.; Ivanova, M.I.; Sievers, S.A.; Apostol, M.I.; Thompson, M.J.; Balbirnie, M.; Wiltzius, J.J.W.; McFarlane, H.T.; et al. Atomic structures of amyloid cross-β spines reveal varied steric zippers. Nature 2007, 447, 453–457. [Google Scholar] [CrossRef]
- Iłowska, E.; Sawicka, J.; Szymańska, A. Synthesis and physicochemical studies of amyloidogenic hexapeptides derived from human cystatin C. J. Pept. Sci. 2018, 24, e3073. [Google Scholar] [CrossRef]
- Iłowska, E. Studies of the Steric Zipper Motif in the Sequence of Human Cystatin C. Ph.D. Thesis, University of Gdansk, Gdansk, Poland, 2014; p. 171. [Google Scholar]
- Thompson, M.J.; Sievers, S.; Karanicolas, J.; Ivanova, M.I.; Baker, D.; Eisenberg, D. The 3D profile method for identifying fibril-forming segments of proteins. Proc. Natl. Acad. Sci. USA 2006, 103, 4074–4078. [Google Scholar] [CrossRef]
- Nina, B.; Koji, N.; Woody, W.R. Circular Dichroism:Principles and Applications, 2nd ed.; Wiley Online Library: New York, NY, USA, 2000; ISBN 978-0-471-33003-5. [Google Scholar]
- Colletier, J.-P.; Laganowsky, A.; Landau, M.; Zhao, M.; Soriaga, A.B.; Goldschmidt, L.; Flot, D.; Cascio, D.; Sawaya, M.R.; Eisenberg, D. Molecular Basis for Amyloid- Polymorphism. Proc. Natl. Acad. Sci. 2011, 108, 16938–16943. [Google Scholar] [CrossRef]
- Marshall, K.E.; Serpell, L.C. Insights into the Structure of Amyloid Fibrils; 2009; Volume 2, Available online: https://www.researchgate.net/publication/228675516_Insights_into_the_Structure_of_Amyloid_Fibrils (accessed on 15 March 2022).
- Minor, W.; Cymborowski, M.; Otwinowski, Z.; Chruszcz, M. HKL-3000: The integration of data reduction and structure solution–from diffraction images to an initial model in minutes. Acta Crystallogr. Sect. D 2006, 62, 859–866. [Google Scholar] [CrossRef]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzym. 1997, 276, 307–326. [Google Scholar]
- Matthews, B.W. Solvent content of protein crystals. J. Mol. Biol. 1968, 33, 491–497. [Google Scholar] [CrossRef]
- Kantardjieff, K.A.; Rupp, B. Matthews coefficient probabilities: Improved estimates for unit cell contents of proteins, DNA, and protein-nucleic acid complex crystals. Protein Sci. 2003, 12, 1865–1871. [Google Scholar] [CrossRef]
- Louros, N.N.; Chrysina, E.D.; Baltatzis, G.E.; Patsouris, E.S.; Hamodrakas, S.J.; Iconomidou, V.A. A common aggregation-prone interface possibly participates in the self-assembly of human zona pellucida proteins. FEBS Lett. 2016, 590, 619–630. [Google Scholar] [CrossRef]
- Janowski, R.; Kozak, M.; Abrahamson, M.; Grubb, A.; Jaskolski, M. 3D domain-swapped human cystatin C with amyloidlike intermolecular ?-sheets. Proteins Struct. Funct. Genet. 2005, 61, 570–578. [Google Scholar] [CrossRef]
- Orlikowska, M.; Jankowska, E.; Kołodziejczyk, R.; Jaskólski, M.; Szymańska, A. Hinge-loop mutation can be used to control 3D domain swapping and amyloidogenesis of human cystatin C. J. Struct. Biol. 2011, 173, 406–413. [Google Scholar] [CrossRef]
- Rodziewicz-Motowidło, S.; Iwaszkiewicz, J.; Sosnowska, R.; Czaplewska, P.; Sobolewski, E.; Szymańska, A.; Stachowiak, K.; Liwo, A. The role of the Val57 amino-acid residue in the hinge loop of the human cystatin C. Conformational studies of the Beta2-L1-Beta3 segments of wild-type human cystatin C and its mutants. Biopolymers 2009, 91, 373–383. [Google Scholar] [CrossRef]
- Szymańska, A.; Jankowska, E.; Orlikowska, M.; Behrendt, I.; Czaplewska, P.; Rodziewicz-Motowidło, S. Influence of point mutations on the stability, dimerization, and oligomerization of human cystatin C and its L68Q variant. Front. Mol. Neurosci. 2012, 5, 82. [Google Scholar] [CrossRef]
- Janowski, R.; Abrahamson, M.; Grubb, A.; Jaskolski, M. Domain Swapping in N-truncated Human Cystatin C. J. Mol. Biol. 2004, 341, 151–160. [Google Scholar] [CrossRef]
- Tsiolaki, P.L.; Louros, N.N.; Hamodrakas, S.J.; Iconomidou, V.A. Exploring the “aggregation-prone” core of human Cystatin C: A structural study. J. Struct. Biol. 2015, 191, 272–280. [Google Scholar] [CrossRef]
- Tsiolaki, P.L.; Hamodrakas, S.J.; Iconomidou, V.A. The pentapeptide LQVVR plays a pivotal role in human cystatin C fibrillization. FEBS Lett. 2015, 589, 159–164. [Google Scholar] [CrossRef]
- Frousios, K.K.; Iconomidou, V.A.; Karletidi, C.-M.; Hamodrakas, S.J. Amyloidogenic determinants are usually not buried. BMC Struct. Biol. 2009, 9, 44. [Google Scholar] [CrossRef]
- Makin, O.S.; Serpell, L.C. X-ray diffraction studies of amyloid structure. Methods Mol. Biol. 2005, 299, 67–80. [Google Scholar]
- Marshall, K.E.; Morris, K.L.; Charlton, D.; O’Reilly, N. Hydrophobic, Aromatic, and Electrostatic Interactions Play a Central Role in Amyloid Fibril Formation and Stability—Biochemistry (ACS Publications). Biochemistry 2011, 50, 2061–2071. [Google Scholar] [CrossRef]
- Sikorski, P.; Atkins, E.D.T.; Serpell, L.C. Structure and texture of fibrous crystals formed by Alzheimer’s Aβ (11–25) peptide fragment. Structure 2003, 11, 915–926. [Google Scholar] [CrossRef]
- Goux, W.J.; Kopplin, L.; Nguyen, A.D.; Leak, K.; Rutkofsky, M.; Shanmuganandam, V.D.; Sharma, D.; Inouye, H.; Kirschner, D.A. The Formation of Straight and Twisted Filaments from Short Tau Peptides. J. Biol. Chem. 2004, 279, 26868–26875. [Google Scholar] [CrossRef]
- Perutz, M.F.; Finch, J.T.; Berriman, J.; Lesk, A. Amyloid fibers are water-filled nanotubes. Proc. Natl. Acad. Sci. USA 2002, 99, 5591–5595. [Google Scholar] [CrossRef]
- Makin, O.S.; Atkins, E.; Sikorski, P.; Johansson, J.; Serpell, L.C. Molecular basis for amyloid fibril formation and stability. Proc. Natl. Acad. Sci. USA 2005, 102, 315–320. [Google Scholar] [CrossRef]
- Maurer-Stroh, S.; Debulpaep, M.; Kuemmerer, N.; de la Paz, M.L.; Martins, I.C.; Reumers, J.; Morris, K.L.; Copland, A.; Serpell, L.; Serrano, L.; et al. Exploring the sequence determinants of amyloid structure using position-specific scoring matrices. Nat. Methods 2010, 7, 237–242. [Google Scholar] [CrossRef]
- Chatani, E.; Yuzu, K.; Ohhashi, Y.; Goto, Y. Current understanding of the structure, stability and dynamic properties of amyloid fibrils. Int. J. Mol. Sci. 2021, 22, 4349. [Google Scholar] [CrossRef]
- Jahn, T.R.; Makin, O.S.; Morris, K.L.; Marshall, K.E.; Tian, P.; Sikorski, P.; Serpell, L.C. The Common Architecture of Cross-β Amyloid. J. Mol. Biol. 2010, 395, 717–727. [Google Scholar] [CrossRef]
- Knowles, T.P.J.; Buehler, M.J. Nanomechanics of functional and pathological amyloid materials. Nat. Nanotechnol. 2011, 6, 469–479. [Google Scholar] [CrossRef]
- Fan, T.; Yu, X.; Shen, B.; Sun, L. Peptide Self-Assembled Nanostructures for Drug Delivery Applications. J. Nanomater. 2017, 2017. [Google Scholar] [CrossRef]
- Marta, D.; Navarro, S.; Fuentes, I.; Teixidor, F.; Ventura, S. Minimalist Prion-Inspired Polar Self- Assembling Peptides. ACS Nano. 2018, 12, 5394–5407. [Google Scholar]
- Rufo, C.M.; Moroz, Y.S.; Moroz, O.V.; Stöhr, J.; Smith, T.A.; Hu, X.; DeGrado, W.F.; Korendovych, I. V Short peptides self-assemble to produce catalytic amyloids. Nat. Chem. 2014, 6, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, S.K.; Siddiqi, M.K.; Alam, P.; Khan, R.H. Protein misfolding and aggregation: Mechanism, factors and detection. Process Biochem. 2016, 51, 1183–1192. [Google Scholar] [CrossRef]
- Nečas, D.; Klapetek, P. Gwyddion: An open-source software for SPM data analysis. Open Phys. 2012, 10, 181–188. [Google Scholar] [CrossRef]
- Arvai, A. ADXV—A Program to Display X-ray Diffraction Images; Scripps Research Institute: La Jolla, CA, USA, 2012. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide ID | Kind of the Sequence | Amino Acid Sequence |
|---|---|---|
| CysZ4 | HCC 55–60 | NH2-QIVAGV-NH2 |
| CysZ9 | HCC 60–65 | NH2-VNYFLD-NH2 |
| CysZ11 | shuffle HCC 55–60 | NH2-VIGAQV-NH2 |
| CysZ13 | shuffle HCC 55–60 | NH2-QAGIVV-NH2 |
| Synchrotron/Beamline | BESSY II/14.2 |
| Temperature (K) | 100 K |
| Space group | C2 |
| Cell parameters (Å,°) | a = 71.56; b = 65.15; c = 48.01, β = 102.20 |
| Wavelength (Å) | 0.91841 |
| Resolution (Å) | 50.0−2.94 (2.99−2.94) a |
| Reflection collected | 9550 |
| Unique reflections | 4775 |
| Rmerge | 0.233 (0.646) |
| <I>/<σI> | 3.7 (1.2) |
| Completeness (%) | 49.6 (43.7) |
| Redundancy | 2.0 (1.7) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iłowska, E.; Barciszewski, J.; Jaskólski, M.; Moliński, A.; Kozak, M.; Szymańska, A. Identification of a Steric Zipper Motif in the Amyloidogenic Core of Human Cystatin C and Its Use for the Design of Self-Assembling Peptides. Int. J. Mol. Sci. 2022, 23, 5800. https://doi.org/10.3390/ijms23105800
Iłowska E, Barciszewski J, Jaskólski M, Moliński A, Kozak M, Szymańska A. Identification of a Steric Zipper Motif in the Amyloidogenic Core of Human Cystatin C and Its Use for the Design of Self-Assembling Peptides. International Journal of Molecular Sciences. 2022; 23(10):5800. https://doi.org/10.3390/ijms23105800
Chicago/Turabian StyleIłowska, Emilia, Jakub Barciszewski, Mariusz Jaskólski, Augustyn Moliński, Maciej Kozak, and Aneta Szymańska. 2022. "Identification of a Steric Zipper Motif in the Amyloidogenic Core of Human Cystatin C and Its Use for the Design of Self-Assembling Peptides" International Journal of Molecular Sciences 23, no. 10: 5800. https://doi.org/10.3390/ijms23105800
APA StyleIłowska, E., Barciszewski, J., Jaskólski, M., Moliński, A., Kozak, M., & Szymańska, A. (2022). Identification of a Steric Zipper Motif in the Amyloidogenic Core of Human Cystatin C and Its Use for the Design of Self-Assembling Peptides. International Journal of Molecular Sciences, 23(10), 5800. https://doi.org/10.3390/ijms23105800