
Salinity Tolerance of Halophytic Grass Puccinellia nuttalliana Is Associated with Enhancement of Aquaporin-Mediated Water Transport by Sodium




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
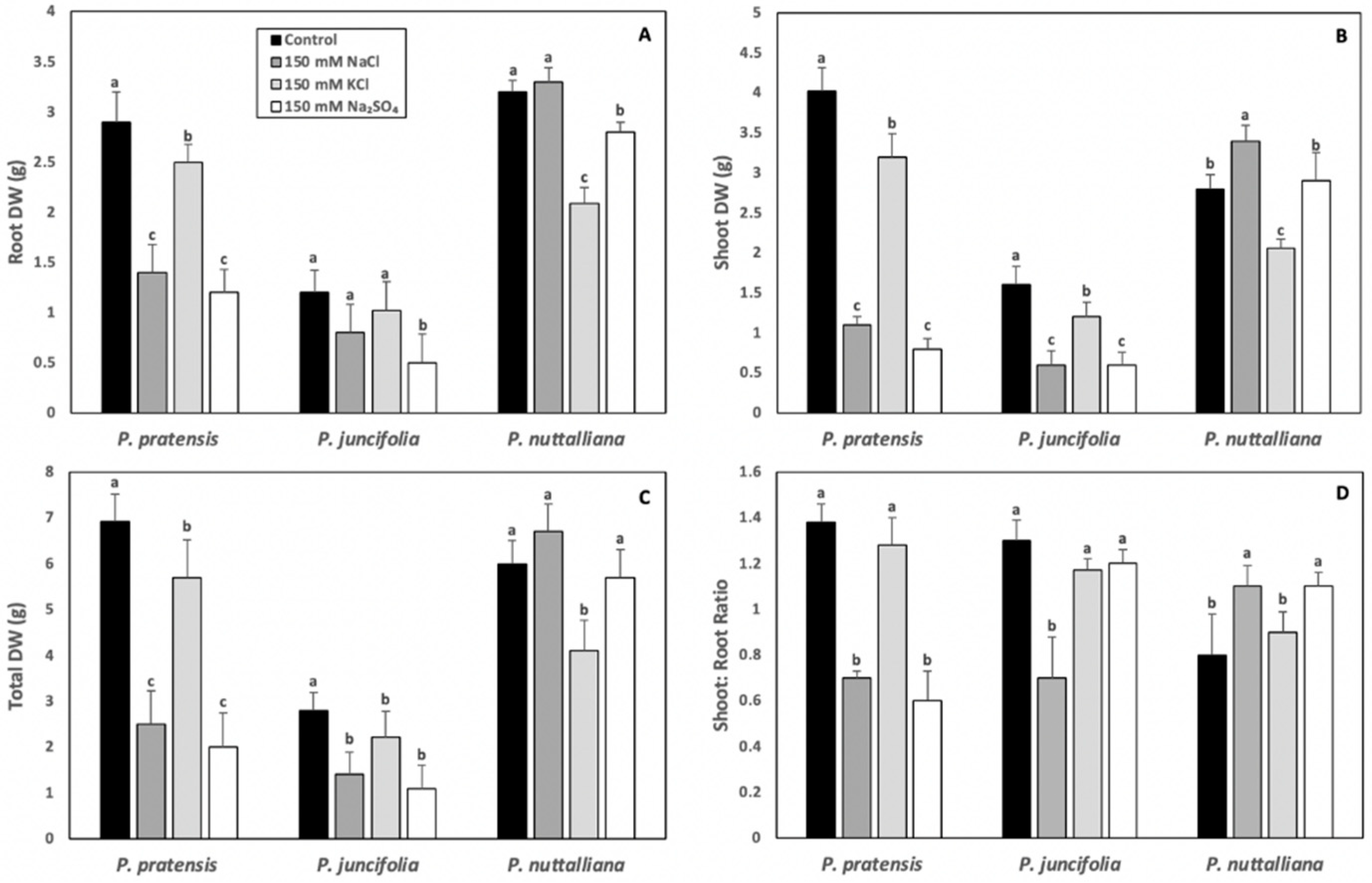
2.1. Plant Morphology and Dry Weights (DW)
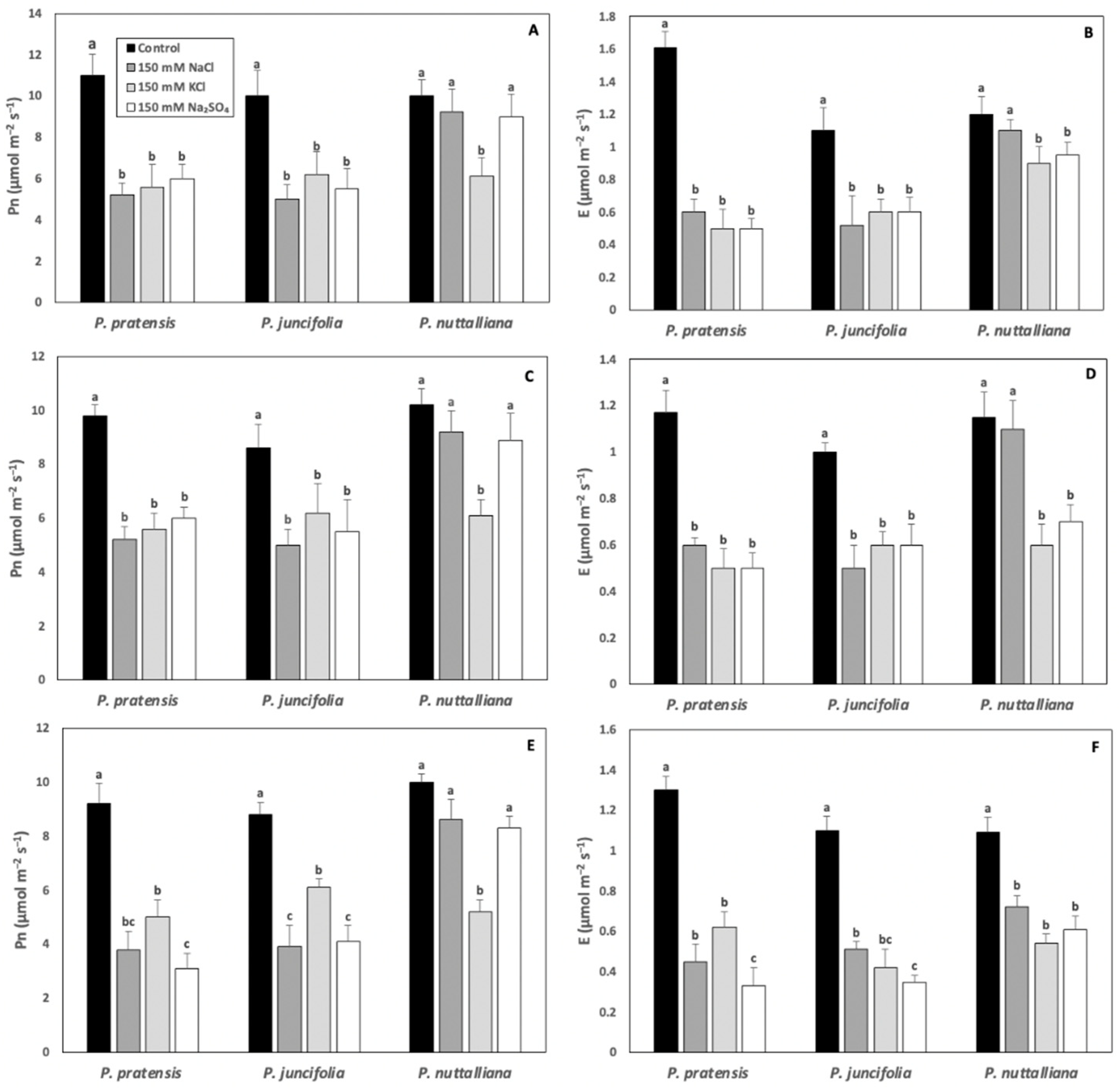
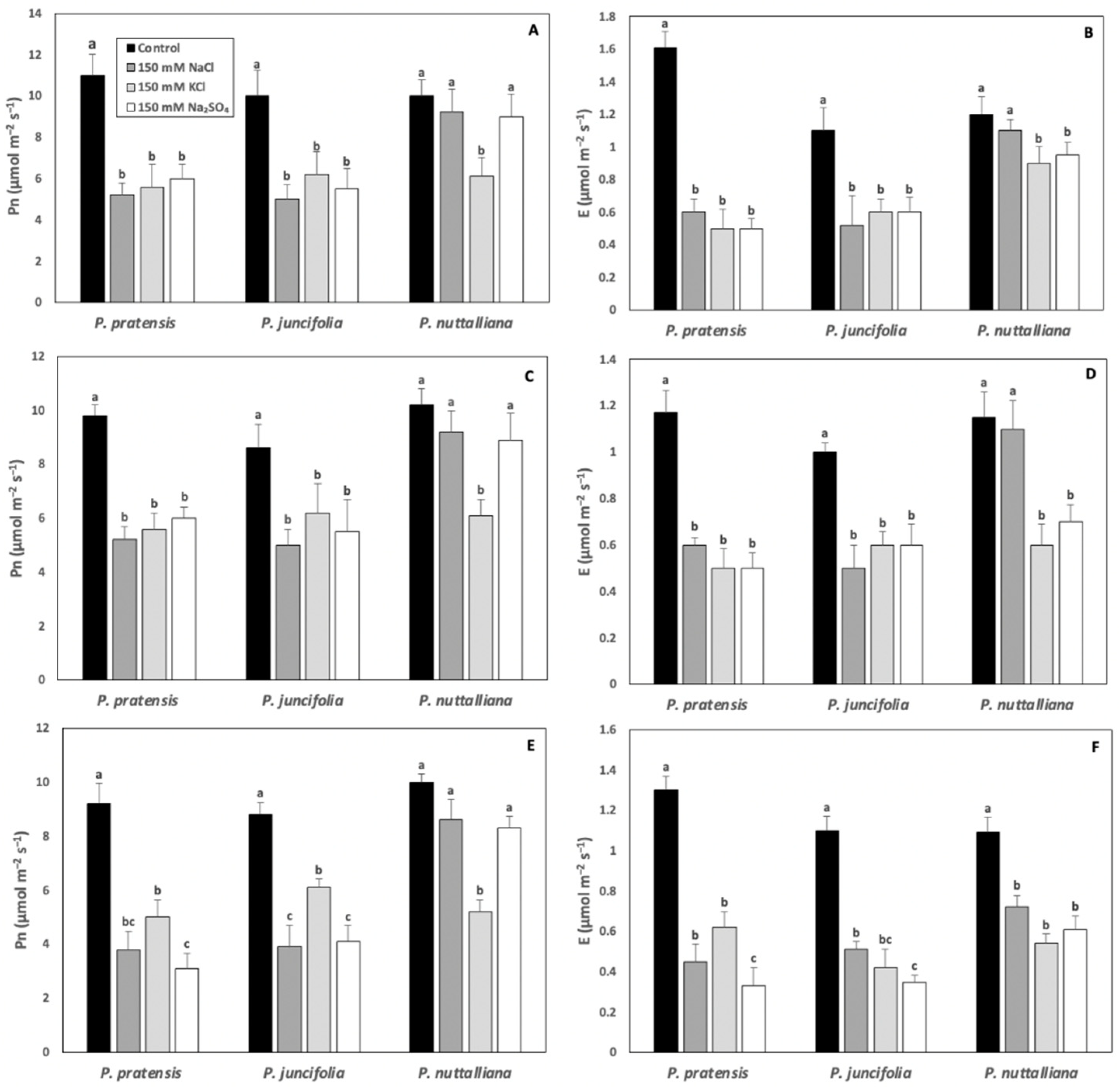
2.2. Net Photosynthesis (Pn) and Transpiration (E) Rates
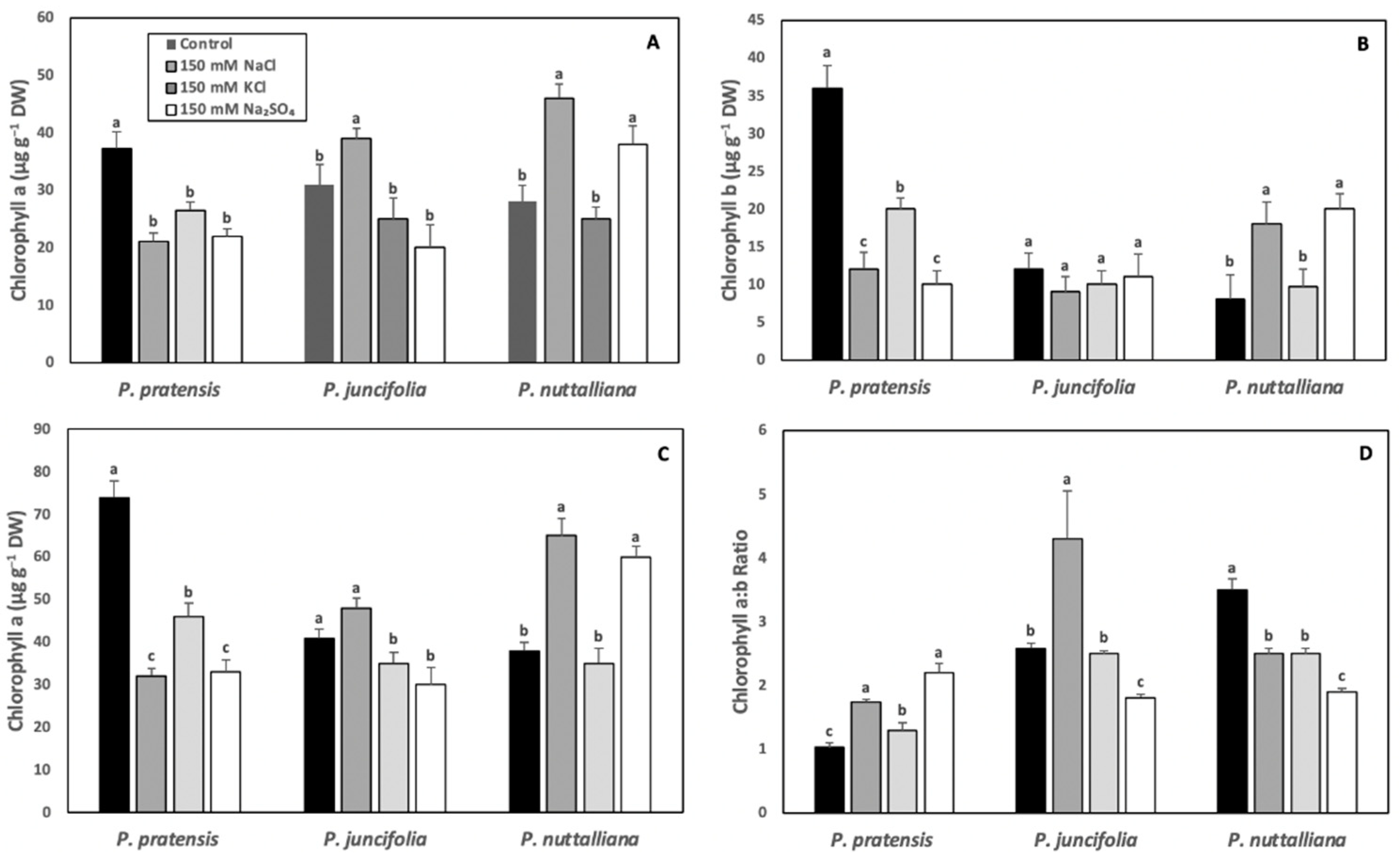
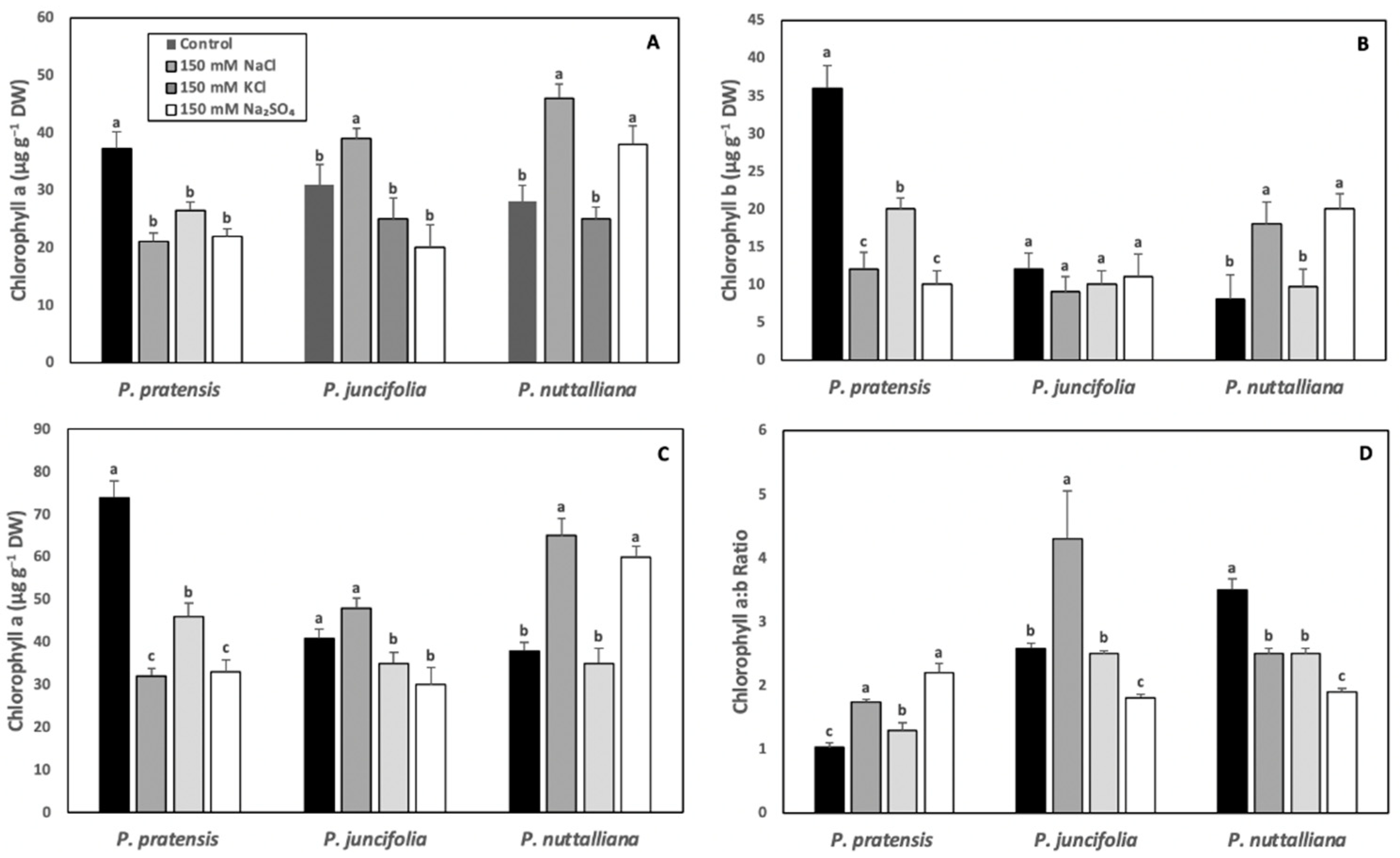
2.3. Leaf Chlorophyll Concentrations
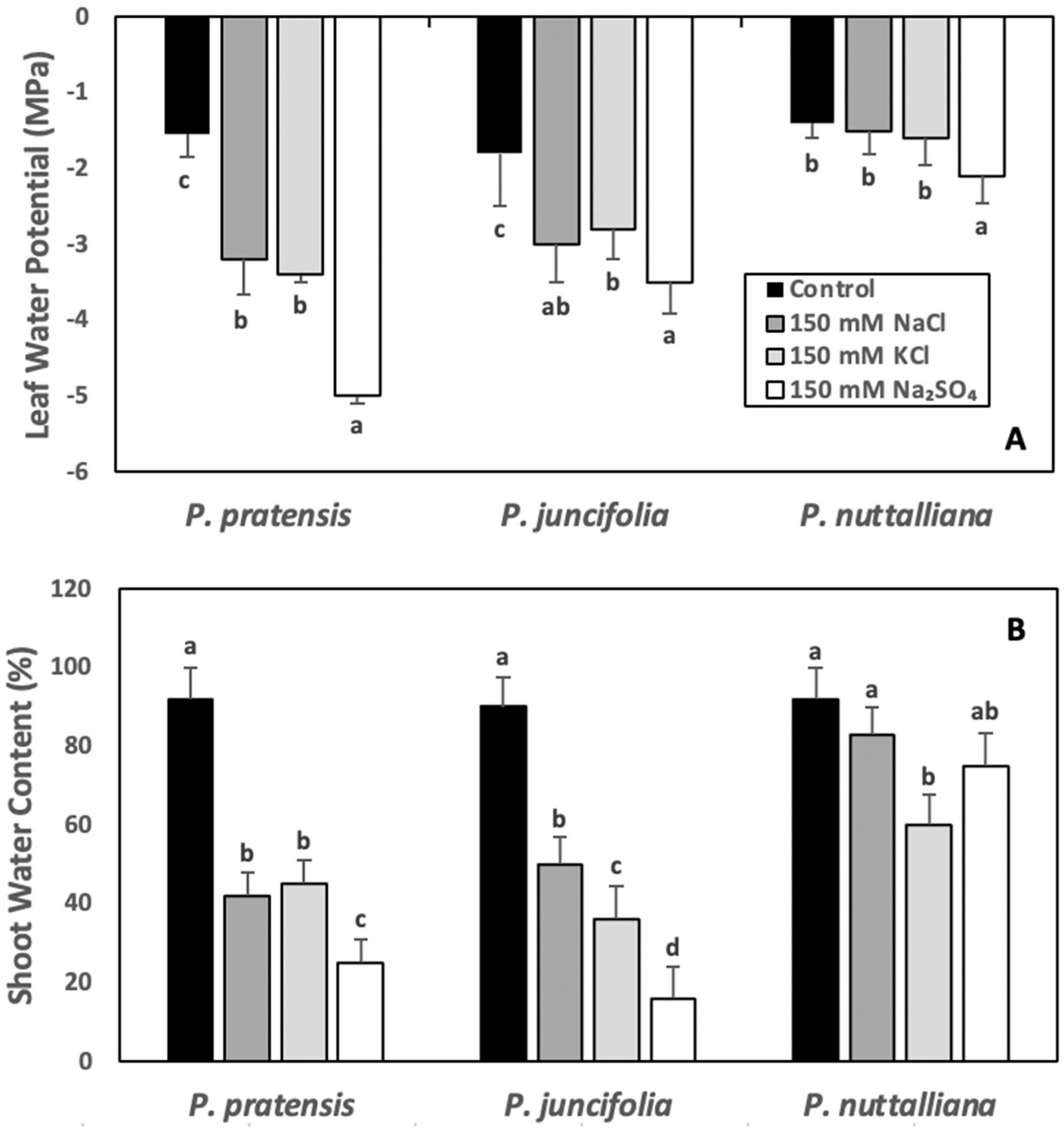
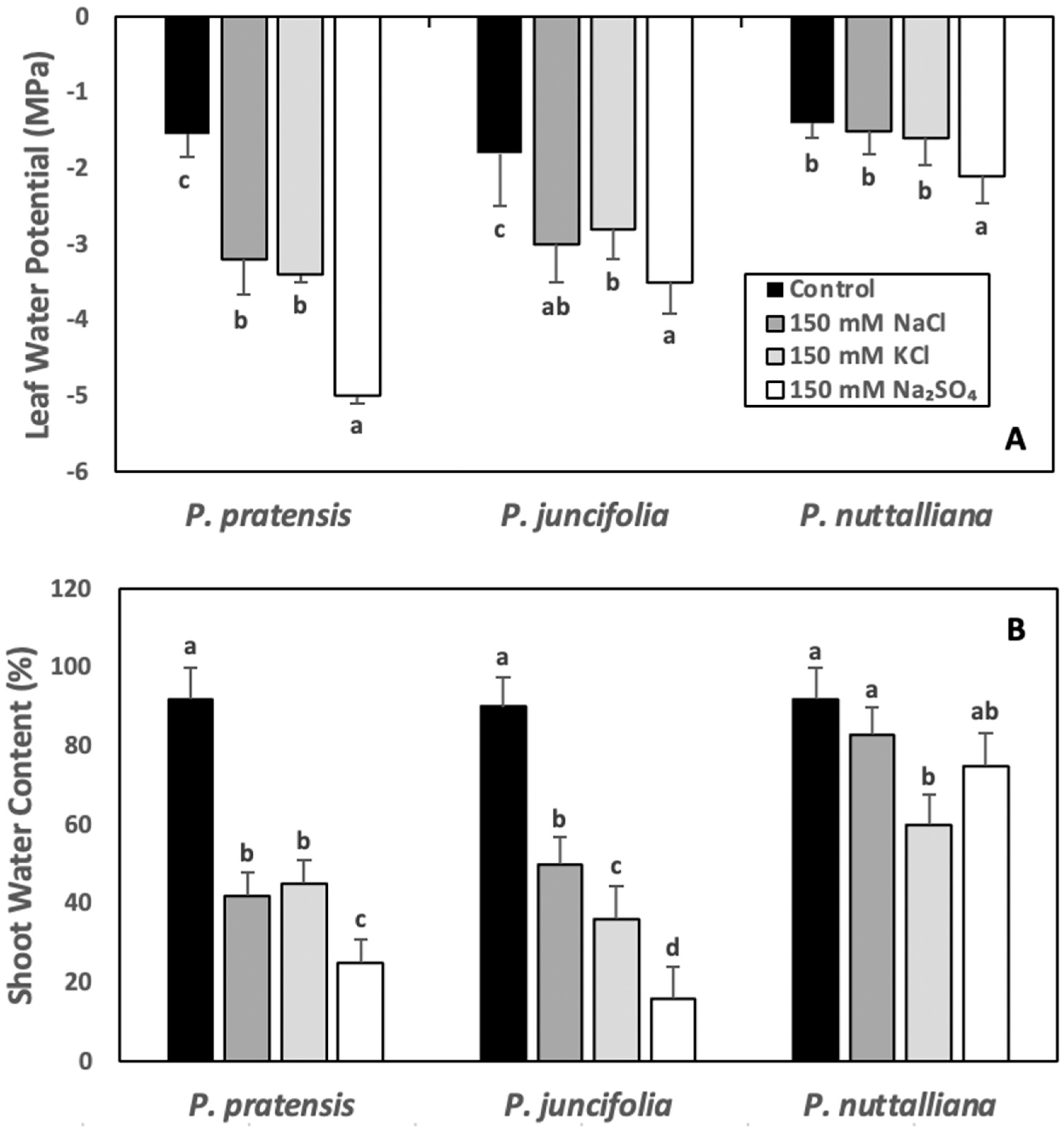
2.4. Leaf Water Potentials (ψw) and Shoot Water Contents (WC)
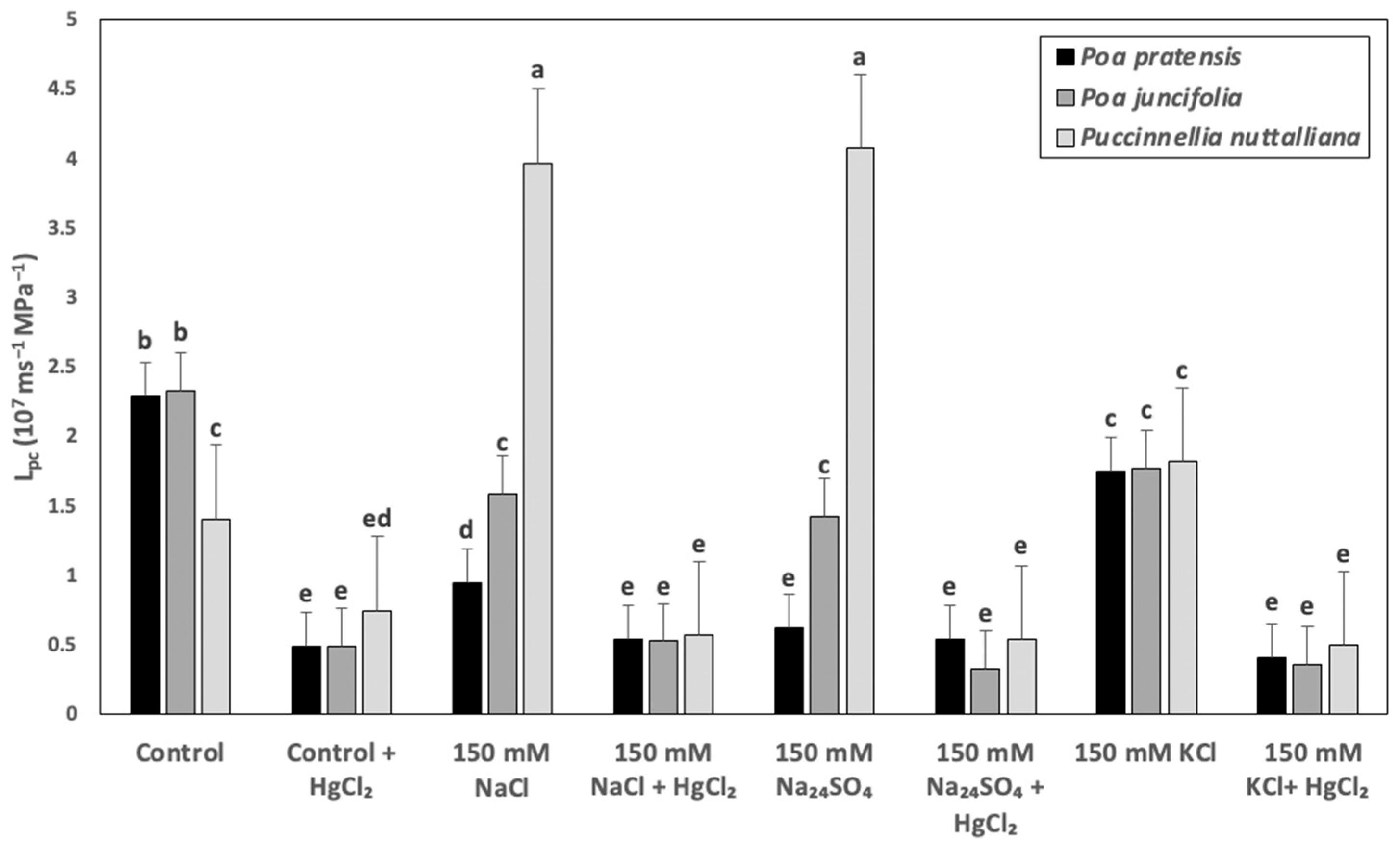
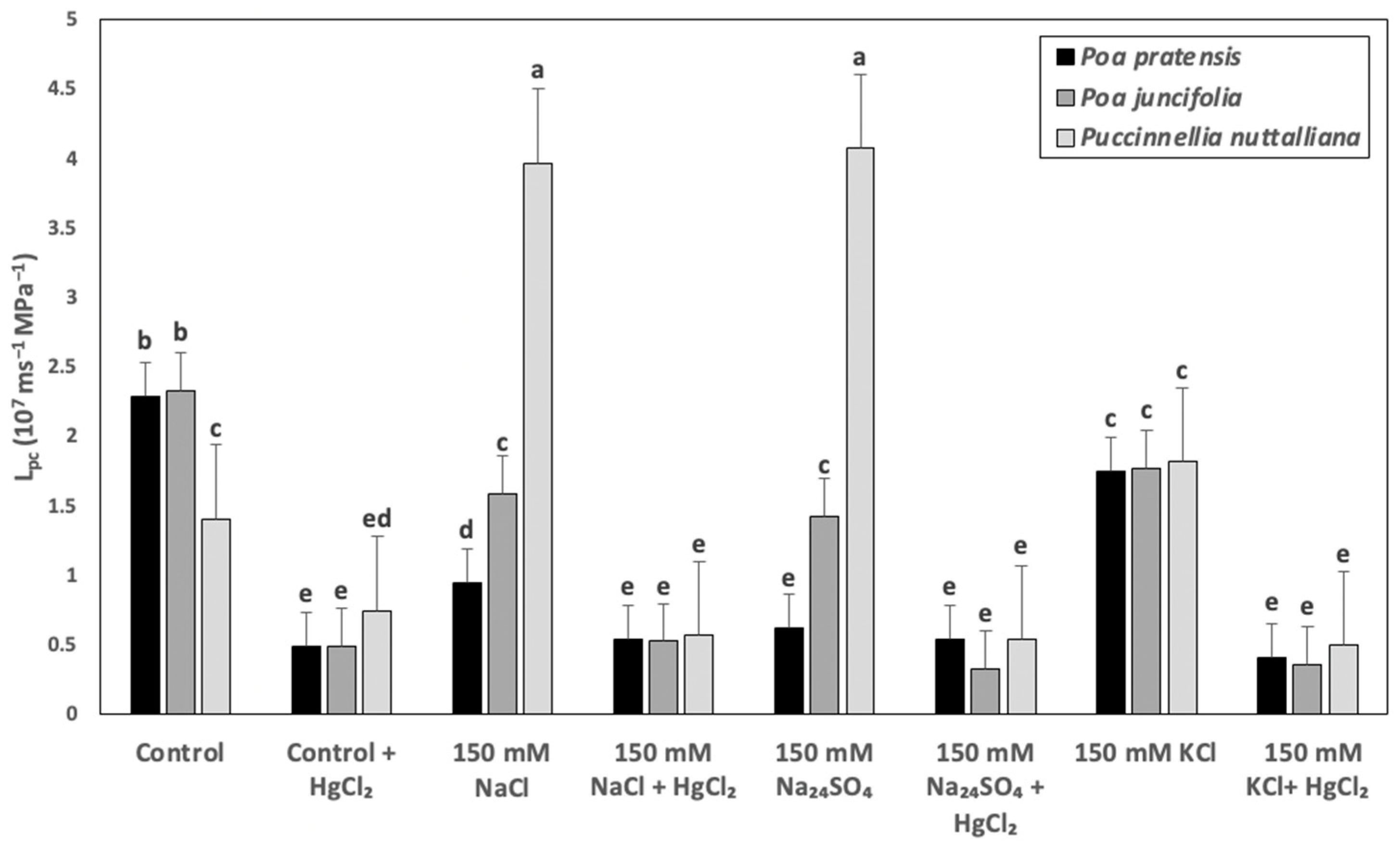
2.5. Cell Hydraulic Conductivity (Lpc)
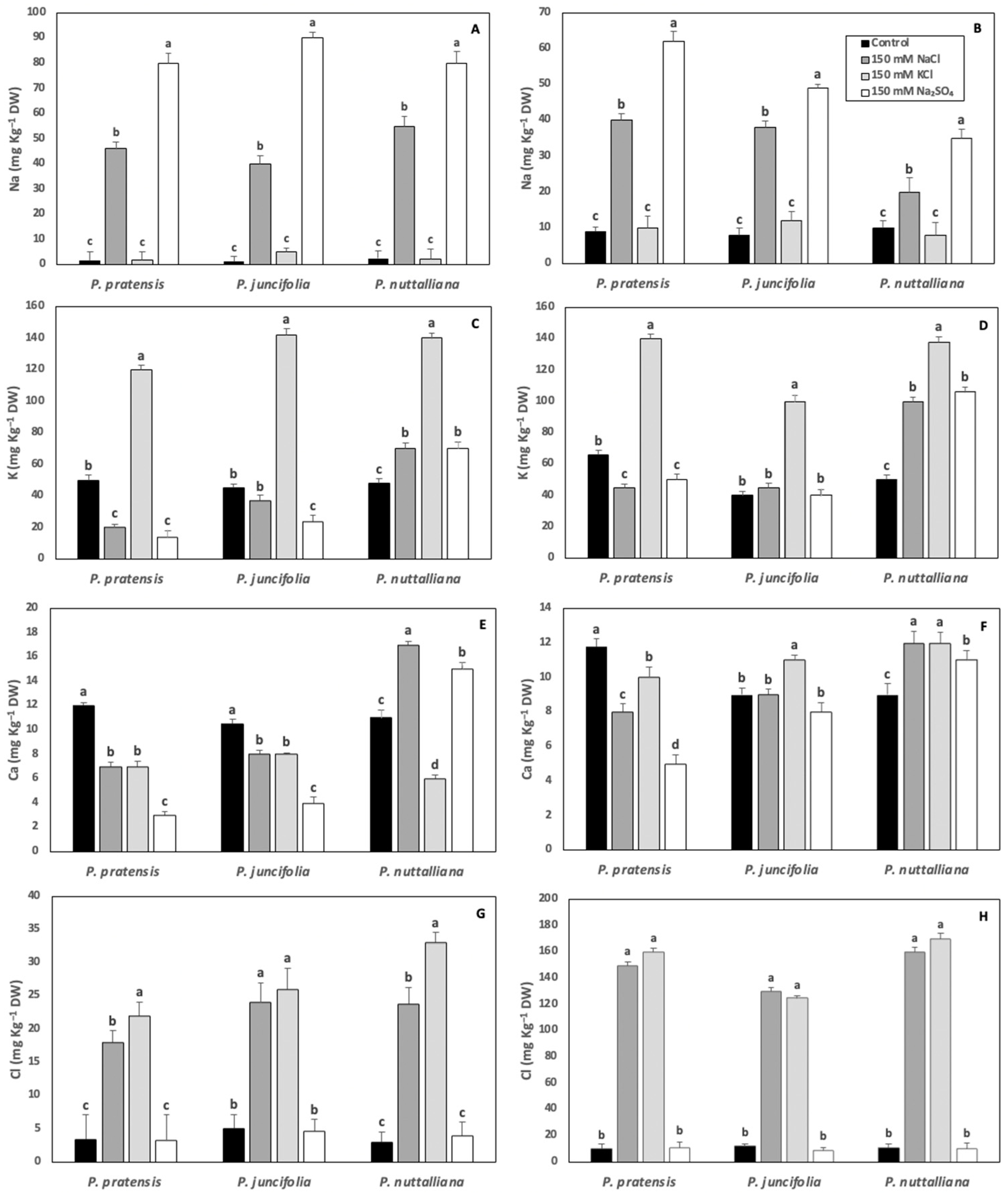
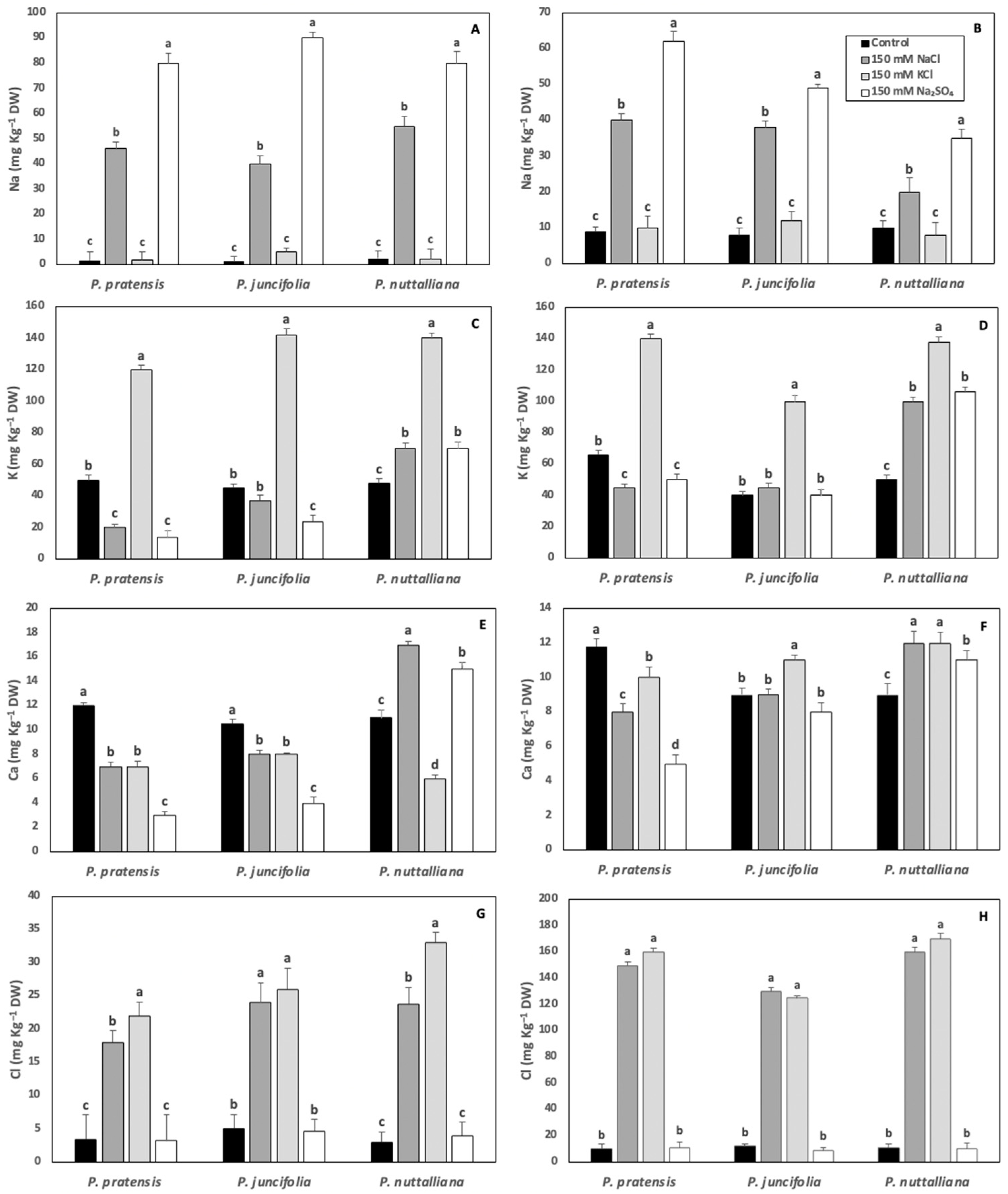
2.6. Root and Shoot Elemental Concentrations
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatments
4.2. Plant Morphology and Dry Weights (DW)
4.3. Net Photosynthesis (Pn) and Transpiration (E) Rates
4.4. Leaf Chlorophyll Concentrations
4.5. Leaf Water Potentials (ψw) and Shoot Water Contents (WC)
4.6. Cell Hydraulic Conductivity (Lpc)
4.7. Tissue Elemental Analyses
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant. Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkov, V.; Flowers, T.J. Mechanisms of ion transport in halophytes: From roots to leaves. In Tasks for Vegetation Science—49. Shabkha Ecosystems; Gul, B., Böer, B., Khan, A., Clüsener-Godt, M., Hameed, A., Eds.; Springer Nature: Cham, Switzerland, 2019; Volume 6, pp. 125–150. [Google Scholar]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Cheesman, J.M. The evolution of halophytes, glycophytes and crops, and its implications for food security under saline conditions. New Phytol. 2015, 206, 557–570. [Google Scholar] [CrossRef] [PubMed]
- Grigore, M.-N.; Toma, C. Anatomical Adaptations of Halophytes. A Review of Classic Literature and Recent Findings; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Mishra, A.; Tana, B. Halophytes: Potential resources for salt stress tolerance genes and promoters. Front. Plant Sci. 2017, 8, 829. [Google Scholar] [CrossRef]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Sun, J.; Chen, S.L.; Dai, S.X.; Wang, R.G.; Li, N.Y.; Shen, X.; Zhang, Z.K. Ion flux profiles and plant ion homeostasis control under salt stress. Plant Signal. Behav. 2009, 4, 261–264. [Google Scholar] [CrossRef] [Green Version]
- Steudle, E.; Peterson, C.A. How does water get through roots? J. Exp. Bot. 1998, 49, 775–788. [Google Scholar] [CrossRef]
- Martinez-Ballesta, M.C.; Aparicio, F.; Pallas, V.; Martinez, V.; Carvajal, M. Influence of saline stress on root hydraulic conductance and PIP expression in Arabidopsis. J. Plant Physiol. 2003, 160, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.; Calvo-Polanco, M.; Zwiazek, J.J. Gas exchange and growth responses of ectomycorrhizal Picea mariana, Picea glauca, and Pinus banksiana seedlings to NaCl and Na2SO4. Plant Biol. 2006, 8, 646–652. [Google Scholar] [CrossRef]
- Lee, S.H.; Calvo-Polanco, M.; Chung, G.C.; Zwiazek, J.J. Role of aquaporins in root water transport of ectomycorrhizal jack pine (Pinus banksiana) seedlings exposed to NaCl and fluoride. Plant Cell Environ. 2010, 33, 769–780. [Google Scholar] [CrossRef]
- Kapilan, R.; Vaziri, M.; Zwiazek, J.J. Regulation of aquaporins under stress. Biol. Res. 2018, 51, 4. [Google Scholar] [CrossRef] [PubMed]
- Carvajal, M.; Cerda, A.; Martinez, V. Does calcium ameliorate the negative effect of NaCl on melon root water transport by regulating aquaporin activity? New Phytol. 2000, 145, 439–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Ballesta, M.C.; Cabanero, F.; Olmos, E.; Periago, P.M.; Maurel, C.; Carvajal, M. Two different effects of calcium on aquaporins in salinity-stressed pepper plants. Planta 2008, 228, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Zwiazek, J.J. Regulation of aquaporin-mediated water transport in Arabidopsis roots exposed to NaCl. Plant Cell Physiol. 2015, 56, 750–758. [Google Scholar] [CrossRef] [Green Version]
- Vitali, V.; Bellati, J.; Soto, G.; Ayub, N.D.; Amodeo, G. Root hydraulic conductivity and adjustments in stomatal conductance: Hydraulic strategy in response to salt stress in a halotolerant species. AoB Plants 2015, 7, plv136. [Google Scholar] [CrossRef] [Green Version]
- Boursiac, Y.; Chen, S.; Luu, D.T.; Sorieul, M.; van den Dries, N.; Maurel, C. Early effects of salinity on water transport in Arabidopsis roots. Molecular and cellular features of aquaporin expression. Plant Physiol. 2005, 139, 790–805. [Google Scholar] [CrossRef] [Green Version]
- Horie, T.; Kaneko, T.; Sugimoto, G.; Sasanom, S.; Pandam, S.K.; Shibasaka, M.; Katsuhara, M. Mechanisms of water transport mediated by PIP aquaporins and their regulation via phosphorylation events under salinity stress in barley roots. Plant Cell Physiol. 2011, 52, 663–675. [Google Scholar] [CrossRef]
- Vaziriyeganeh, M.; Lee, S.H.; Zwiazek, J.J. Water transport properties of root cells contribute to salt tolerance in halophytic grasses Poa juncifolia and Puccinellia nuttalliana. Plant Sci. 2018, 276, 54–62. [Google Scholar] [CrossRef]
- Vaziriyeganeh, M.; Khan, S.; Zwiazek, J.J. Transcriptome and metabolome analyses reveal potential salt tolerance mechanisms contributing to maintenance of water balance by the halophytic grass Puccinellia nuttalliana. Front. Plant Sci. 2021, 12, 760863. [Google Scholar] [CrossRef]
- Isayenkov, S.V.; Maathuis, F.J. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Maurel, C.; Boursiac, Y.; Luu, D.T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T.; Horie, T.; Nakahara, Y.; Tsuji, N.; Shibasaka, M.; Katsuhara, T. Dynamic regulation of root hydraulic conductivity of barley plants in response to salinity/osmotic stress. Plant Cell Physiol. 2015, 56, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Lüttge, U.; Ratajczak, R. Effects of salt treatment and osmotic stress on V-ATPase and V-PPase in leaves of the halophyte Suaeda salsa. J. Exp. Bot. 2001, 52, 2355–2365. [Google Scholar] [CrossRef] [PubMed]
- Ramos, J.; Lopez, M.J.; Benlloch, M. Effect of NaCl and KCl salts on the growth and solute accumulation of the halophyte Atriplex nummularia. Plant Soil 2004, 259, 163–168. [Google Scholar] [CrossRef]
- Fatemi, F.; Hashemi-Petroudi, S.H.; Nematzadeh, G.; Askari, H.; Abdollahi, M.R. Exploiting differential gene expression to discover ionic and osmotic-associated transcripts in the halophyte grass Aeluropus littoralis. Biol. Proced. Online 2019, 21, 14. [Google Scholar] [CrossRef] [Green Version]
- Harivandi, M.A.; Butler, J.D.; Wu, L. Salinity and turfgrass culture. In Turfgrass; Waddington, D.V., Carrow, R.N., Shearman, R.C., Eds.; American Society of Agronomy: Madison, WI, USA, 1992; Series No. 32; pp. 207–229. [Google Scholar]
- Silva, C.; Martínez, V.; Carvajal, M. Osmotic versus toxic effects of NaCl on pepper plants. Biol. Plant. 2008, 52, 72–79. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Hsiao, T.C.; Xu, L.K. Sensitivity of growth of roots versus leaves to water stress: Biophysical analysis and relation to water transport. J. Exp. Bot. 2000, 51, 1595–1616. [Google Scholar] [CrossRef]
- Wang, D.; Grieve, C.M.; Suarez, D.L. Composition of irrigation water salinity affects growth characteristics and uptake of selenium and salt ions by soybean. J. Plant. Nutr. 2005, 28, 1073–1088. [Google Scholar] [CrossRef] [Green Version]
- Andrés, Z.; Pérez-Hormaeche, J.; Leidi, E.O.; Schlücking, K.; Steinhorst, L.; McLachlan, D.H.; Schumacher, K.; Hetherington, A.M.; Kudla, J.; Cubero, E.; et al. Control of vacuolar dynamics and regulation of stomatal aperture by tonoplast potassium uptake. Proc. Natl. Acad. Sci. USA 2014, 111, E1806–E1814. [Google Scholar] [CrossRef] [Green Version]
- Hedrich, R.; Shabala, S. Stomata in a saline world. Curr. Opin. Plant Biol. 2018, 46, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Rasouli, F.; Kiani-Pouya, A.; Tahir, A.; Shabala, L.; Chen, Z.; Shabala, S. A comparative analysis of stomatal traits and photosynthetic responses in closely related halophytic and glycophytic species under saline conditions. Environ. Exp. Bot. 2021, 181, 104300. [Google Scholar] [CrossRef]
- Hu, L.; Wang, Z.; Huang, B. Effects of cytokinin and potassium on stomatal and photosynthetic recovery of Kentucky bluegrass from drought stress. Crop Sci. 2013, 53, 221–231. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Kwon, O.K.; Mekapogu, M.; Kim, K.S. Effect of salinity stress on photosynthesis and related physiological responses in carnation (Dianthus caryophyllus). Hortic. Environ. Biotechnol. 2019, 60, 831–839. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful conditions: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Stepien, P.; Johnson, G.N. Contrasting responses of photosynthesis to salt stress in the glycophyte Arabidopsis and the halophyte Thellungiella: Role of the plastid terminal oxidase as an alternative electron sink. Plant Physiol. 2009, 149, 1154–1165. [Google Scholar] [CrossRef] [Green Version]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Bose, J.; Munns, R.; Shabala, S.; Gilliham, M.; Pogson, B.; Tyerman, S.D. Chloroplast function and ion regulation in plants growing on saline soils: Lessons from halophytes. J. Exp. Bot. 2017, 68, 3129–3143. [Google Scholar] [CrossRef]
- Percey, W.J.; McMinn, A.; Bose, J.; Breadmore, M.C.; Guijt, R.M.; Shabala, S. Salinity effects on chloroplast PSII performance in glycophytes and halophytes. Funct. Plant Biol. 2016, 43, 1003–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, M.; Kunz, H.H.; Schroeder, J.I.; Kemp, G.; Young, H.S.; Neuhaus, H.E. Decreased capacity for sodium export out of Arabidopsis chloroplasts impairs salt tolerance, photosynthesis and plant performance. Plant J. 2014, 78, 646–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhsin, T.M.; Zwiazek, J.J. Ectomycorrhizae increase water conductance and protect white spruce (Picea glauca) seedlings against salt stress. Plant Soil 2002, 238, 217–225. [Google Scholar] [CrossRef]
- Apostol, K.G.; Zwiazek, J.J. Hypoxia affects root sodium and chloride concentrations and alters water conductance in salt-treated jack pine (Pinus banksiana) seedlings. Trees 2003, 17, 251–257. [Google Scholar] [CrossRef]
- Calvo-Polanco, M.C.; Zwiazek, J.J.; Voicu, M.C. Responses of ectomycorrhizal American elm (Ulmus americana) seedlings to salinity and soil compaction. Plant Soil 2008, 308, 189–200. [Google Scholar] [CrossRef]
- Nardini, A.; Gasco, A.; Trifilo, P.; Lo Gullo, M.A.; Salleo, S. Ion-mediated enhancement of xylem hydraulic conductivity is not always suppressed by the presence of Ca2+ in the sap. J. Exp. Bot. 2007, 58, 2609–2615. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Equiza, M.A.; Navarro-Rodenas, A.; Lee, S.H.; Zwiazek, J.J. Hydraulic adjustments in aspen (Populus tremuloides) seedlings following defoliation involve root and leaf aquaporins. Planta 2014, 240, 553–564. [Google Scholar] [CrossRef]
- Alleva, K.; Niemietz, C.M.; Sutka, M.; Maurel, C.; Parisi, M.; Tyerman, S.D.; Amodeo, G. Plasma membrane of Beta vulgaris storage root shows high water channel activity regulated by cytoplasmic pH and a dual range of calcium concentrations. J. Exp. Bot. 2006, 57, 609–621. [Google Scholar] [CrossRef] [Green Version]
- Abideen, Z.; Koyro, H.W.; Huchzermeyer, B.; Ahmed, M.Z.; Gul, B.; Khan, M.A. Moderate salinity stimulates growth and photosynthesis of Phragmites karka by water relations and tissue specific ion regulation. Environ. Exp. Bot. 2014, 105, 70–76. [Google Scholar] [CrossRef]
- Tada, Y.; Komatsubara, S.; Kurusu, T. Growth and physiological adaptation of whole plants and cultured cells from a halophyte turf grass under salt stress. AoB Plants 2014, 6, plu041. [Google Scholar] [CrossRef] [Green Version]
- White, P.J.; Broadley, M.R. Chloride in soils and its uptake and movement within the plant: A review. Ann. Bot. 2001, 88, 967–988. [Google Scholar] [CrossRef] [Green Version]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant. 2008, 133, 651–669. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Pottosin, I.I.; Cuin, T.A.; Fuglsang, A.T.; Tester, M.; Jha, D.; Zepeda-Jazo, I.; Zhou, M.; Palmgren, M.G.; Newman, I.A.; et al. Root plasma membrane transporters controlling K+/Na+ homeostasis in salt-stressed barley. Plant Physiol. 2007, 145, 1714–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuin, T.A.; Betts, S.A.; Chalmandrier, R.; Shabala, S. A root’s ability to retain K+ correlates with salt tolerance in wheat. J. Exp. Bot. 2008, 59, 2697–2706. [Google Scholar] [CrossRef] [Green Version]
- Hauser, F.; Horie, T.A. conserved primary salt tolerance mechanism mediated by HKT transporters: A mechanism for sodium exclusion and maintenance of high K+/Na+ ratio in leaves during salinity stress. Plant Cell Environ. 2010, 33, 552–565. [Google Scholar] [CrossRef]
- Roy, S.; Chakraborty, U. Salt tolerance mechanisms in salt tolerant grasses (STGs) and their prospects in cereal crop improvement. Bot. Stud. 2014, 55, 31. [Google Scholar] [CrossRef] [Green Version]
- Volkov, V.; Wang, B.; Dominy, P.J.; Fricke, W.; Amtmann, A. Thellungiella halophila, a salt-tolerant relative of Arabidopsis thaliana, possesses effective mechanisms to discriminate between potassium and sodium. Plant Cell Environ. 2004, 27, 1–14. [Google Scholar] [CrossRef]
- Shabala, S.; Demidchik, V.; Shabala, L.; Cuin, T.A.; Smith, S.J.; Miller, A.J.; Davies, J.M.; Newman, I.A. Extracellular Ca2+ ameliorates NaCl-induced K+ loss from Arabidopsis root and leaf cells by controlling plasma membrane K+-permeable channels. Plant Physiol. 2006, 141, 1653–1665. [Google Scholar] [CrossRef] [Green Version]
- Carvajal, M.; Martínez, V.; Alcaraz, C.F. Physiological function of water channels as affected by salinity in roots of paprika pepper. Physiol. Plant. 1999, 105, 95–101. [Google Scholar] [CrossRef]
- Franklin, J.A.; Zwiazek, J.J. Ion uptake in Pinus banksiana treated with sodium chloride and sodium sulphate. Physiol. Plant. 2004, 120, 482–490. [Google Scholar] [CrossRef]
- Qian, Z.-J.; Song, J.-J.; Chaumont, F.; Ye, Q. Differential responses of plasma membrane aquaporins in mediating water transport of cucumber seedlings under osmotic and salt stresses. Plant Cell Environ. 2015, 38, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Zwiazek, J.J. Mercuric chloride effects on root water transport in aspen (Populus tremuloides) seedlings. Plant Physiol. 1999, 121, 939–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, Y.; Okimoto, N.; Kadohira, I.; Suematsu, M.; Yasuoka, K.; Yasui, M. Molecular mechanisms of how mercury inhibits water permeation through aquaporin-1: Understanding by molecular dynamics simulation. Biophys. J. 2010, 98, 1512–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeira, A.; Moura, T.F.; Soveral, G. Detecting aquaporin function and regulation. Front. Chem. 2016, 4, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwiazek, J.J.; Tan, X.; Xu, H.; Navarro-Ródenas, A.; Morte, A. Functional significance of oxygen transport through aquaporins. Sci. Rep. 2017, 17, 40411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyerman, S.D.; McGaughey, S.A.; Qiu, J.; Yool, A.J.; Byrt, C.S. Adaptable and multifunctional ion-conducting aquaporins. Annu. Rev. Plant Biol. 2021, 72, 8.1–8.34. [Google Scholar] [CrossRef]
- Katsuhara, M.; Hanba, Y.T. Barley plasma membrane intrinsic proteins (PIP aquaporins) as water and CO2 transporters. Pflug. Arch. Eur. J. Physiol. 2008, 456, 687–691. [Google Scholar] [CrossRef] [Green Version]
- Katsuhara, M.; Rhee, J.Y.; Sugimoto, G.; Chung, G.C. Early response in water relations influenced by NaCl reflects tolerance or sensitivity of barley plants to salinity stress via aquaporins. Soil Sci. Plant Nutr. 2011, 57, 50–60. [Google Scholar] [CrossRef]
- Knipfer, T.; Danjou, M.; Vionne, C.; Fricke, W. Salt stress reduces water uptake in barley (Hordeum vulgare L.) through modification of the transcellular path. Plant Cell Environ. 2021, 44, 458–475. [Google Scholar] [CrossRef]
- Luu, D.T.; Martiniere, A.; Sorieul, M.; Runions, J.; Maurel, C. Fluorescence recovery after photobleaching reveals high cycling dynamics of plasma membrane aquaporins in Arabidopsis roots under salt stress. Plant J. 2012, 69, 894–905. [Google Scholar] [CrossRef]
- Kourghi, M.; Nourmohammadi, S.; Pei, J.V.; Qiu, J.; McGaughey, S.; Tyerman, S.D.; Byrt, C.S.; Yool, A. Divalent cations regulate the ion conductance properties of diverse classes of aquaporins. Int. J. Mol. Sci. 2017, 16, 2323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, S.T.H.; Imran, S.; Horie, T.; Qiu, J.; McGaughey, S.; Byrt, C.S.; Tyerman, S.D.; Katsuhara, M. A survey of barley PIP aquaporin ionic conductance reveals Ca2+-sensitive HvPIP2;8 Na+ and K+ conductance. Int. J. Mol. Sci. 2020, 21, 7135. [Google Scholar] [CrossRef] [PubMed]
- Barzegargolchini, B.; Movafeghi, A.; Dehestani, A.; Mehrabanjoubani, P. Increased cell wall thickness of endodermis and protoxylem in Aeluropus littoralis roots under salinity: The role of LAC4 and PER64 genes. J. Plant Physiol. 2017, 218, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Pardo, F.E.; Hilal, M.B.; Albornoz, P.L.; Gallardo, M.G.; Ruiz, V.E. Anatomical and physiological responses of four quinoa cultivars to salinity at seedling stage. Ind. J. Sci. Technol. 2017, 10, 1–12. [Google Scholar] [CrossRef]
- Fischer, M.; Kaldenhoff, R. On the pH regulation of plant aquaporins. J. Biol. Chem. 2008, 283, 33889–33892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geilfus, C.M.; Muehling, K.H. Transient alkalinization in the leaf apoplast of Vicia faba L. depends on NaCl stress intensity: An in situ ratio imaging study. Plant Cell Environ. 2012, 35, 578–587. [Google Scholar] [CrossRef]
- Morgan, S.H.; Lindberg, S.; Maity, P.J.; Geilfus, C.M.; Plieth, C.; Mühling, K.H. Calcium improves apoplastic-cytosolic ion homeostasis in salt-stressed Vicia faba leaves. Funct. Plant Biol. 2017, 44, 515–524. [Google Scholar] [CrossRef] [Green Version]
- Morgan, S.H.; Maity, P.J.; Geilfus, C.M.; Lindberg, S.; Mühling, K.H. Leaf ion homeostasis and plasma membrane H+-ATPase activity in Vicia faba change after extra calcium and potassium supply under salinity. Plant Physiol. Biochem. 2014, 82, 244–253. [Google Scholar] [CrossRef]
- Hadi, M.R.; Karimi, N. The role of calcium in plant’s salt tolerance. J. Plant. Nutr. 2012, 35, 2037–2054. [Google Scholar] [CrossRef]
- Demidchik, V.; Shabala, S.; Isayenkov, S.; Cuin, T.A.; Pottosin, I. Calcium transport across plant membranes: Mechanisms and functions. New Phytol. 2018, 220, 49–69. [Google Scholar] [CrossRef] [Green Version]
- Nikalje, G.C.; Nikam, T.D.; Suprasanna, P. Looking at halophytic adaptation to high salinity through genomics landscape. Curr. Genom. 2017, 18, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Chung, G.C.; Jang, J.Y.; Ahn, S.J.; Hong, S.-W.; Zwiazek, J.J. Overexpression of PIP2;5 aquaporin alleviates effects of low root temperature on cell hydraulic conductivity and growth in Arabidopsis. Plant Physiol. 2012, 159, 479–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, F.; Song, C.-P.; He, J.; Zhu, H. Polyamines improve K+/Na+ homeostasis in barley seedlings by regulating root ion channel activities. Plant Physiol. 2007, 145, 1061–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, J.N.; Burris, J.N.; Stewart, C.N.; Lenaghan, S.C. Improved tissue culture conditions for the emerging C4 model Panicum hallii. BMC Biotechnol. 2017, 17, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, E.X. Mineral Nutrition of Plants: Principles and Perspectives; Wiley: New York, NY, USA, 1972. [Google Scholar]
- Šesták, Z.; Catský, J.; Jarvis, P.G. Plant Photosynthetic Production. Manual of Methods; Dr. W. Junk NV Publishers: The Hague, The Netherlands, 1971. [Google Scholar]
- Lee, S.H.; Zwiazek, J.J. Regulation of water transport in Arabidopsis by methyl jasmonate. Plant Physiol. Biochem. 2019, 139, 540–547. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaziriyeganeh, M.; Carvajal, M.; Du, N.; Zwiazek, J.J. Salinity Tolerance of Halophytic Grass Puccinellia nuttalliana Is Associated with Enhancement of Aquaporin-Mediated Water Transport by Sodium. Int. J. Mol. Sci. 2022, 23, 5732. https://doi.org/10.3390/ijms23105732
Vaziriyeganeh M, Carvajal M, Du N, Zwiazek JJ. Salinity Tolerance of Halophytic Grass Puccinellia nuttalliana Is Associated with Enhancement of Aquaporin-Mediated Water Transport by Sodium. International Journal of Molecular Sciences. 2022; 23(10):5732. https://doi.org/10.3390/ijms23105732
Chicago/Turabian StyleVaziriyeganeh, Maryamsadat, Micaela Carvajal, Ning Du, and Janusz J. Zwiazek. 2022. "Salinity Tolerance of Halophytic Grass Puccinellia nuttalliana Is Associated with Enhancement of Aquaporin-Mediated Water Transport by Sodium" International Journal of Molecular Sciences 23, no. 10: 5732. https://doi.org/10.3390/ijms23105732
APA StyleVaziriyeganeh, M., Carvajal, M., Du, N., & Zwiazek, J. J. (2022). Salinity Tolerance of Halophytic Grass Puccinellia nuttalliana Is Associated with Enhancement of Aquaporin-Mediated Water Transport by Sodium. International Journal of Molecular Sciences, 23(10), 5732. https://doi.org/10.3390/ijms23105732