
Loxl2 and Loxl3 Paralogues Play Redundant Roles during Mouse Development

 ,
,
Abstract
:1. Introduction
2. Results
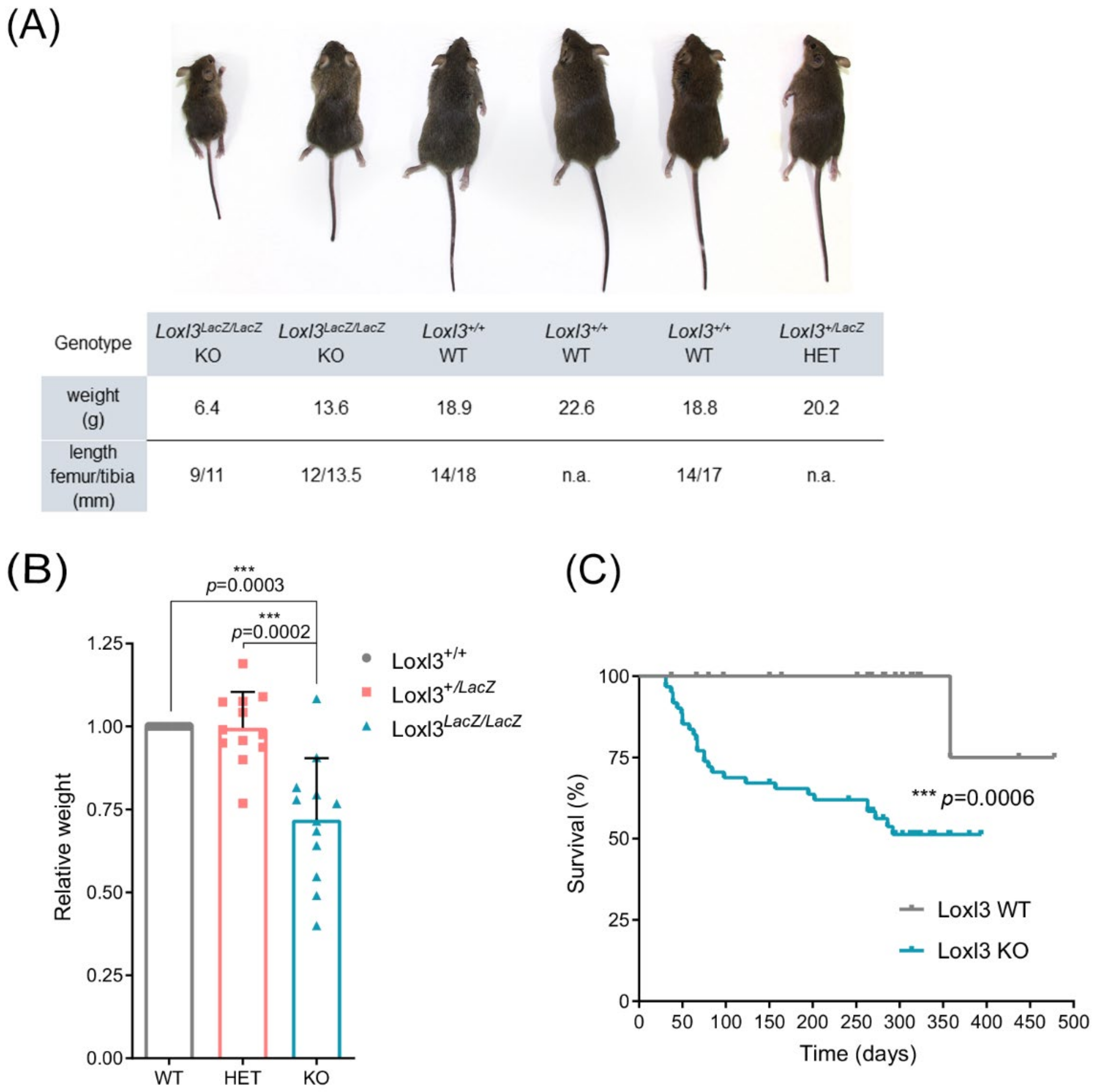
2.1. Deletion of Loxl3 Provokes Perinatal Lethality
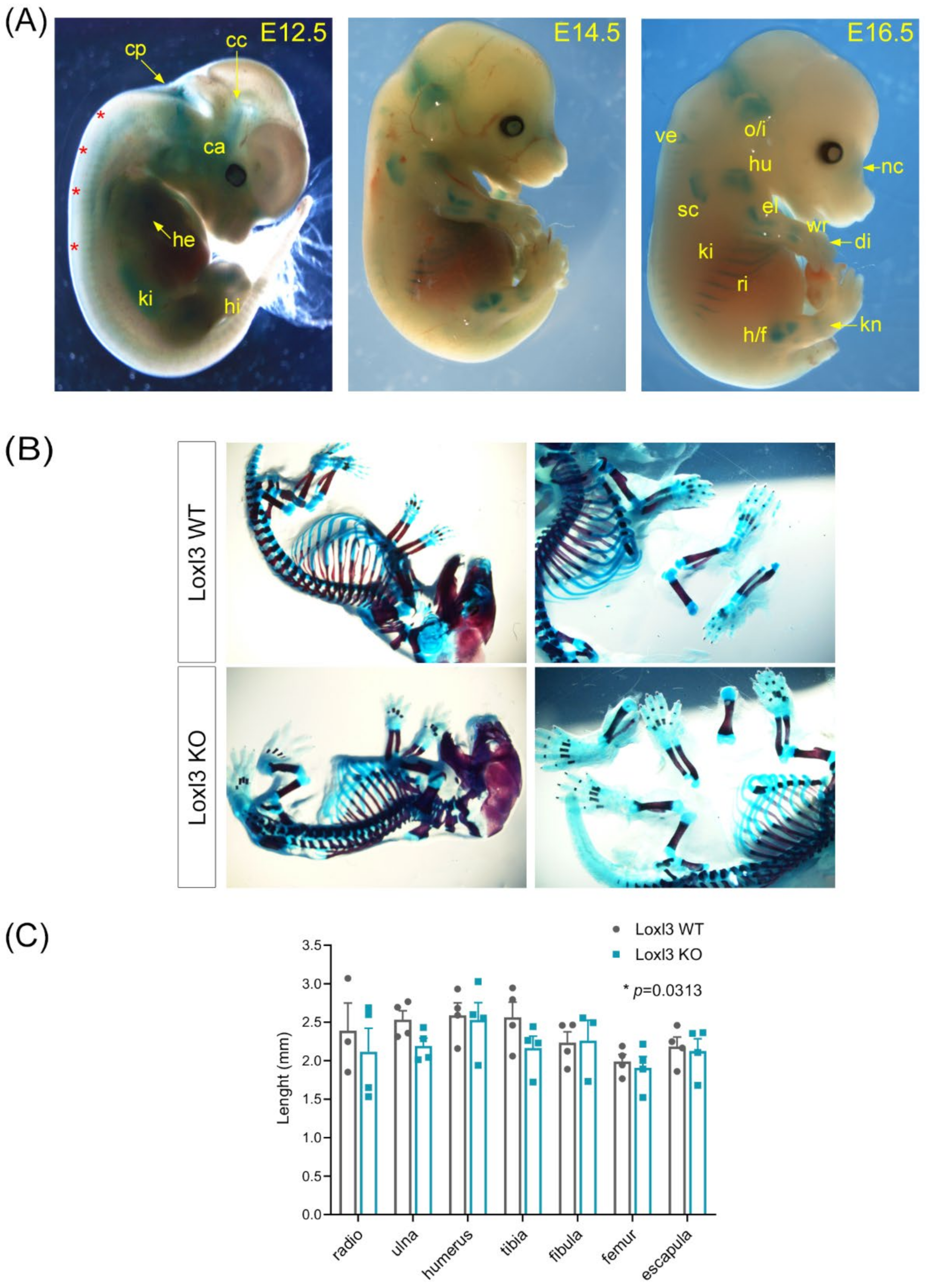
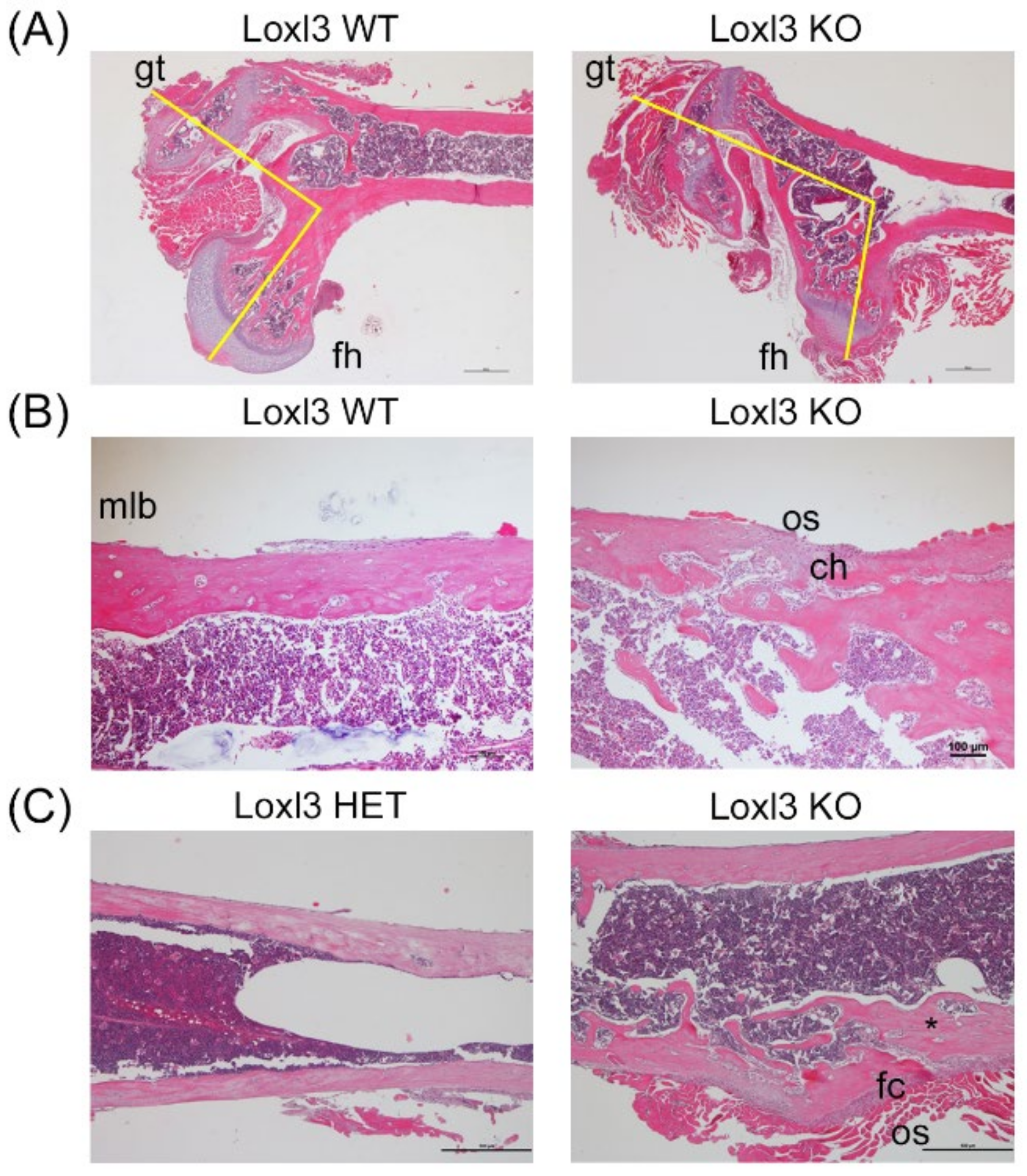
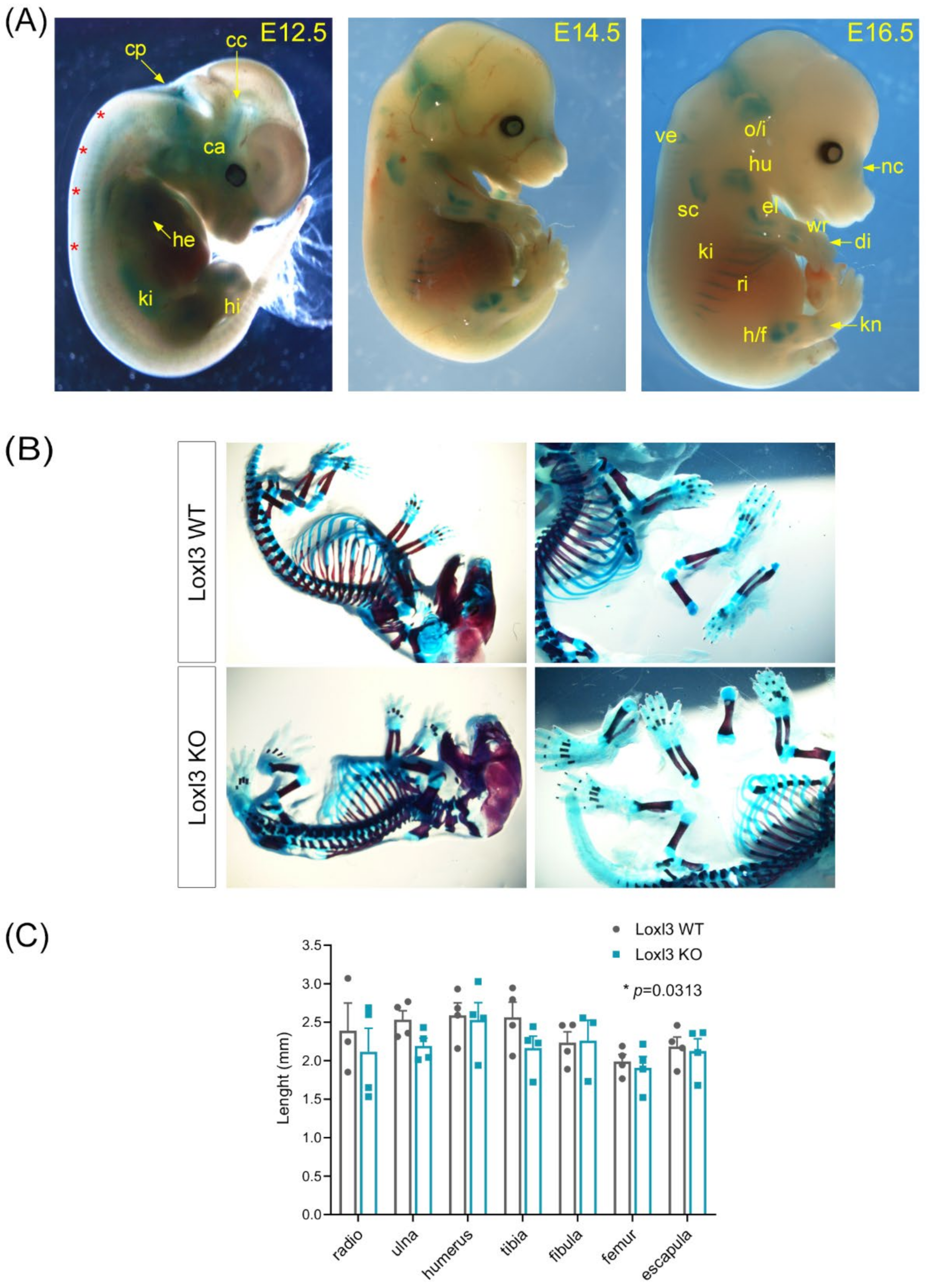
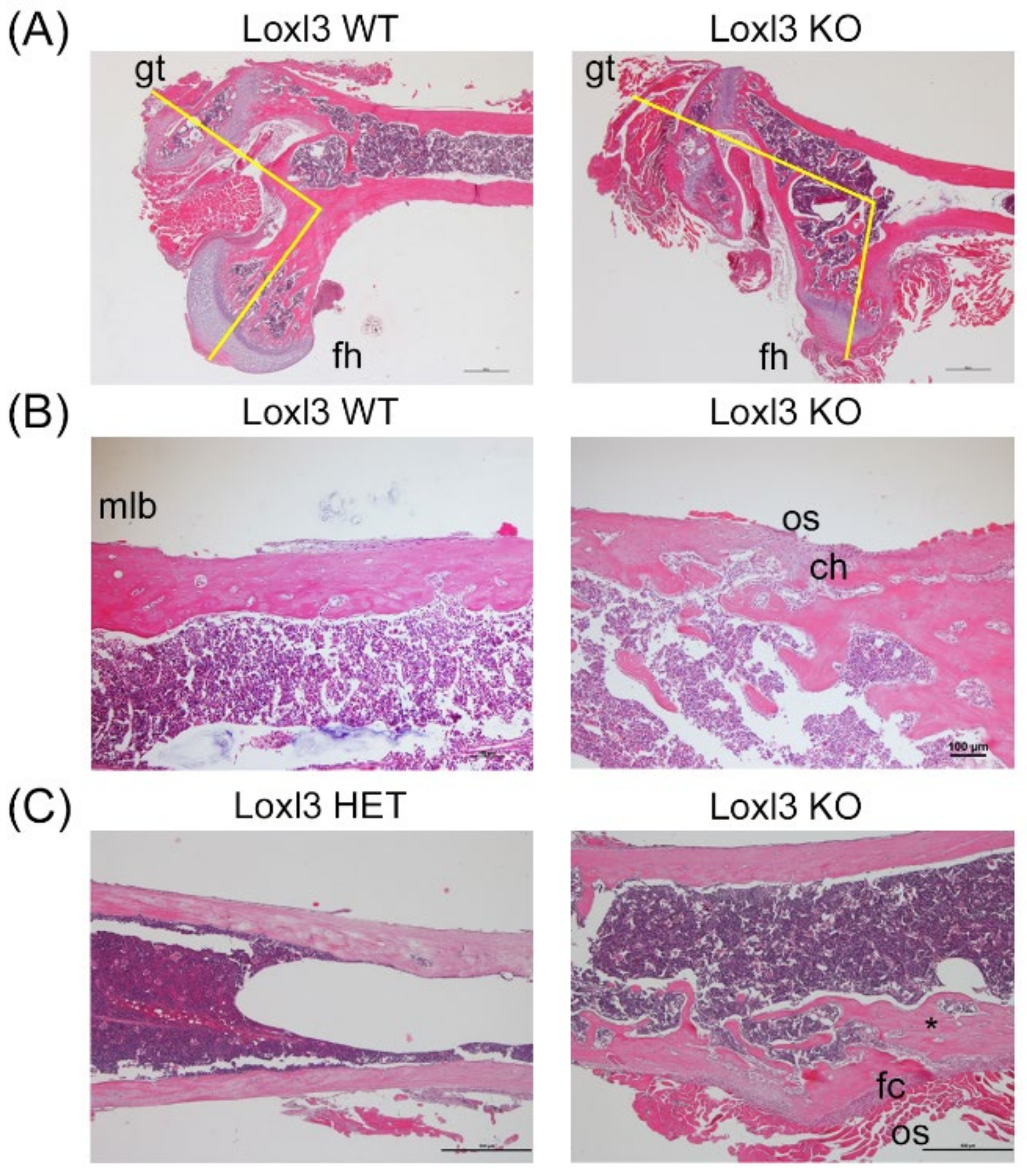
2.2. Loxl3 KO Mice Present Skeletal Abnormalities
2.3. Double Knockout of Loxl2 and Loxl3 Genes Leads to Embryonic Lethality
2.4. Ubiquitous Expression of Loxl2 Ameliorates Perinatal Lethality in Loxl3 KO Mice
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Histology
4.3. Whole-Mount X-Gal Staining of Embryos
4.4. Skeletal Stainings
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Finney, J.; Moon, H.-J.; Ronnebaum, T.; Lantz, M.; Mure, M. Human copper-dependent amine oxidases. Arch. Biochem. Biophys. 2014, 546, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Grau-Bové, X.; Ruiz-Trillo, I.; Rodriguez-Pascual, F. Origin and evolution of lysyl oxidases. Sci. Rep. 2015, 5, 10568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajdú, I.; Kardos, J.; Major, B.; Fabó, G.; Lőrincz, Z.; Cseh, S.; Dormán, G. Inhibition of the LOX enzyme family members with old and new ligands. Selectivity analysis revisited. Bioorg. Med. Chem. Lett. 2018, 28, 3113–3118. [Google Scholar] [CrossRef] [PubMed]
- Lucero, H.A.; Kagan, H.M. Lysyl oxidase: An oxidative enzyme and effector of cell function. Cell. Mol. Life Sci. 2006, 63, 2304–2316. [Google Scholar] [CrossRef]
- Martínez, V.G.; Moestrup, S.K.; Holmskov, U.; Mollenhauer, J.; Lozano, F. The Conserved Scavenger Receptor Cysteine-Rich Superfamily in Therapy and Diagnosis. Pharmacol. Rev. 2011, 63, 967–1000. [Google Scholar] [CrossRef] [Green Version]
- Trackman, P.C. Enzymatic and non-enzymatic functions of the lysyl oxidase family in bone. Matrix Biol. 2016, 52, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Cano, A.; Santamaría, P.G.; Moreno-Bueno, G. LOXL2 in epithelial cell plasticity and tumor progression. Future Oncol. 2012, 8, 1095–1108. [Google Scholar] [CrossRef]
- Barker, H.E.; Cox, T.R.; Erler, J.T. The rationale for targeting the LOX family in cancer. Nat. Rev. Cancer 2012, 12, 540–552. [Google Scholar] [CrossRef]
- Trackman, P.C. Lysyl Oxidase Isoforms and Potential Therapeutic Opportunities for Fibrosis and Cancer. Expert Opin. Ther. Targets 2016, 20, 935–945. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, C.; Martínez-González, J. The Role of Lysyl Oxidase Enzymes in Cardiac Function and Remodeling. Cells 2019, 8, 1483. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Pascual, F.; Rosell-Garcia, T. Lysyl Oxidases: Functions and Disorders. J. Glaucoma 2018, 27 (Suppl. 1), S15–S19. [Google Scholar] [CrossRef]
- Yang, N.; Cao, D.-F.; Yin, X.-X.; Zhou, H.-H.; Mao, X.-Y. Lysyl oxidases: Emerging biomarkers and therapeutic targets for various diseases. Biomed. Pharmacother. 2020, 131, 110791. [Google Scholar] [CrossRef]
- Ye, M.; Song, Y.; Pan, S.; Chu, M.; Wang, Z.-W.; Zhu, X. Evolving roles of lysyl oxidase family in tumorigenesis and cancer therapy. Pharmacol. Ther. 2020, 215, 107633. [Google Scholar] [CrossRef]
- Mäki, J.M.; Räsänen, J.; Tikkanen, H.; Sormunen, R.; Mäkikallio, K.; Kivirikko, K.I.; Soininen, R. Inactivation of the Lysyl Oxidase Gene Lox Leads to Aortic Aneurysms, Cardiovascular Dysfunction, and Perinatal Death in Mice. Circulation 2002, 106, 2503–2509. [Google Scholar] [CrossRef] [Green Version]
- Hornstra, I.K.; Birge, S.; Starcher, B.; Bailey, A.J.; Mecham, R.P.; Shapiro, S.D. Lysyl Oxidase Is Required for Vascular and Diaphragmatic Development in Mice. J. Biol. Chem. 2003, 278, 14387–14393. [Google Scholar] [CrossRef] [Green Version]
- Galán, M.; Martí-Pàmies, I.; Varona, S.; Rodríguez-Calvo, R.; Briones, A.M.; Navarro, M.A.; de Diego, A.; Osada, J.; Orriols, M.; Guadall, A.; et al. Lysyl oxidase (LOX) in vascular remodelling. Insight from a new animal model. Thromb. Haemost. 2014, 112, 812–824. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, Y.; Gao, J.; Pawlyk, B.; Starcher, B.; Spencer, J.A.; Yanagisawa, H.; Zuo, J.; Li, T. Elastic fiber homeostasis requires lysyl oxidase–like 1 protein. Nat. Genet. 2004, 36, 178–182. [Google Scholar] [CrossRef]
- AlSofi, L.; Daley, E.; Hornstra, I.; Morgan, E.F.; Mason, Z.D.; Acevedo, J.F.; Word, R.A.; Gerstenfeld, L.C.; Trackman, P.C. Sex-Linked Skeletal Phenotype of Lysyl Oxidase Like-1 Mutant Mice. Calcif. Tissue Res. 2015, 98, 172–185. [Google Scholar] [CrossRef]
- Martín, A.; Salvador, F.; Moreno-Bueno, G.; Floristán, A.; Ruiz-Herguido, C.; Cuevas, E.P.; Morales, S.; Santos, V.; Csiszar, K.; Dubus, P.; et al. Lysyl oxidase-like 2 represses Notch1 expression in the skin to promote squamous cell carcinoma progression. EMBO J. 2015, 34, 1090–1109. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Savvatis, K.; Kang, J.S.; Fan, P.; Zhong, H.; Schwartz, K.; Barry, V.; Mikels-Vigdal, A.; Karpinski, S.; Kornyeyev, D.; et al. Targeting LOXL2 for cardiac interstitial fibrosis and heart failure treatment. Nat. Commun. 2016, 7, 13710. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, R.; Liu, Z.; Hou, C.; Zong, W.; Zhang, A.; Sun, X.; Gao, J. Loss of lysyl oxidase-like 3 causes cleft palate and spinal deformity in mice. Hum. Mol. Genet. 2015, 24, 6174–6185. [Google Scholar] [CrossRef] [Green Version]
- Kraft-Sheleg, O.; Zaffryar-Eilot, S.; Genin, O.; Yaseen, W.; Soueid-Baumgarten, S.; Kessler, O.; Smolkin, T.; Akiri, G.; Neufeld, G.; Cinnamon, Y.; et al. Localized LoxL3-Dependent Fibronectin Oxidation Regulates Myofiber Stretch and Integrin-Mediated Adhesion. Dev. Cell 2016, 36, 550–561. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Guo, J.; Jia, Y.; Kong, W.; Li, W. LOXL4 Abrogation Does Not Exaggerate Angiotensin II-Induced Thoracic or Abdominal Aortic Aneurysm in Mice. Genes 2021, 12, 513. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Z.; Zhang, T.; Lin, Z.; Li, Z.; Zhang, A.; Sun, X.; Gao, J. Loss of Lysyl Oxidase-like 3 Attenuates Embryonic Lung Development in Mice. Sci. Rep. 2016, 6, 33856. [Google Scholar] [CrossRef]
- Vázquez-Naharro, A.; Bustos-Tauler, J.; Floristán, A.; Yuste, L.; Oltra, S.S.; Vinyals, A.; Moreno-Bueno, G.; Fabra, À.; Portillo, F.; Cano, A.; et al. Loxl3 Promotes Melanoma Progression and Dissemination Influencing Cell Plasticity and Survival. Cancers 2022, 14, 1200. [Google Scholar] [CrossRef]
- van Boxtel, A.L.; Gansner, J.M.; Hakvoort, H.W.; Snell, H.; Legler, J.; Gitlin, J.D. Lysyl oxidase-like 3b is critical for cartilage maturation during zebrafish craniofacial development. Matrix Biol. 2011, 30, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Bai, X.; Wan, P.; Mo, F.; Chen, G.; Zhang, J.; Gao, J. Targeted Deletion of Loxl3 by Col2a1-Cre Leads to Progressive Hearing Loss. Front. Cell Dev. Biol. 2021, 9, 683495. [Google Scholar] [CrossRef]
- Alzahrani, F.; Al Hazzaa, S.A.; Tayeb, H.; Alkuraya, F.S. LOXL3, encoding lysyl oxidase-like 3, is mutated in a family with autosomal recessive Stickler syndrome. Qual. Life Res. 2015, 134, 451–453. [Google Scholar] [CrossRef]
- Robin, N.H.; Moran, R.T.; Ala-Kokko, L. Stickler Syndrome. In GeneReviews® [Internet]; Adam, M.P., Ardinger, H.H., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Gripp, K.W., Mirzaa, G.M., Amemiya, A., Eds.; University of Washington, Seattle: Seattle, WA, USA, 2021. [Google Scholar]
- Tissières, V.; Geier, F.; Kessler, B.; Wolf, E.; Zeller, R.; Lopez-Rios, J. Gene Regulatory and Expression Differences between Mouse and Pig Limb Buds Provide Insights into the Evolutionary Emergence of Artiodactyl Traits. Cell Rep. 2020, 31, 107490. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Genotype | After Weaning (%) | E18.5 (%) | Expected (%) |
|---|---|---|---|
| Loxl3+/+ | 30.09 | 18.52 | 25 |
| Loxl3+/LacZ | 64.26 | 55.56 | 50 |
| Loxl3LacZ/LacZ | 5.64 | 25.93 | 25 |
| Total number of animals analysed | 319 | 81 |
| Genotype | After Weaning (%) | Expected (%) |
|---|---|---|
| Loxl2+/+; Loxl3+/+ | 13.31 | 6.25 |
| Loxl2+/+; Loxl3+/LacZ | 23.89 | 12.25 |
| Loxl2+/+; Loxl3LacZ/LacZ | 3.75 | 6.25 |
| Loxl2+/−; Loxl3+/+ | 20.14 | 12.5 |
| Loxl2+/−; Loxl3+/LacZ | 34.81 | 25 |
| Loxl2+/−; Loxl3LacZ/LacZ | 0.68 | 12.5 |
| Loxl2−/−; Loxl3+/+ | 2.05 | 6.25 |
| Loxl2−/−; Loxl3+/LacZ | 1.37 | 12.5 |
| Loxl2−/−; Loxl3LacZ/LacZ | 0 | 6.25 |
| Total number of mice analysed | 293 |
| Genotype | E9.5 (%) | E11.5 (%) | E13.5 (%) | Expected (%) |
|---|---|---|---|---|
| Loxl2+/+; Loxl3+/+ | 4.82 | 9.38 | 11.11 | 6.25 |
| Loxl2+/+; Loxl3+/LacZ | 10.84 | 12.5 | 15.38 | 12.25 |
| Loxl2+/+; Loxl3LacZ/LacZ | 4.82 | 12.5 | 3.42 | 6.25 |
| Loxl2+/−; Loxl3+/+ | 14.46 | 28.13 | 16.24 | 12.5 |
| Loxl2+/−; Loxl3+/LacZ | 39.76 | 28.13 | 38.46 | 25 |
| Loxl2+/−; Loxl3LacZ/LacZ | 9.64 | 6.25 | 8.55 | 12.5 |
| Loxl2−/−; Loxl3+/+ | 7.23 | 3.13 | 3.42 | 6.25 |
| Loxl2−/−; Loxl3+/LacZ | 3.61 | 0 | 3.42 | 12.5 |
| Loxl2−/−; Loxl3LacZ/LacZ | 4.82 | 0 | 0 | 6.25 |
| Total number of mice analysed | 83 | 32 | 117 |
| Genotype | After Weaning (%) | Expected (%) |
|---|---|---|
| R26Loxl2/Loxl2; Loxl3LacZ/LacZ | 2 | 6.25 |
| R26Loxl2/+; Loxl3LacZ/LacZ | 12.67 | 12.25 |
| R26+/+; Loxl3LacZ/LacZ | 0 | 6.25 |
| R26Loxl2/Loxl2; Loxl3+/LacZ | 5.33 | 12.5 |
| R26Loxl2/+; Loxl3+/LacZ | 44 | 25 |
| R26+/+; Loxl3+/LacZ | 7.33 | 12.5 |
| R26Loxl2/Loxl2; Loxl3+/+ | 12.67 | 6.25 |
| R26Loxl2/+; Loxl3+/+ | 14 | 12.5 |
| R26+/+; Loxl3+/+ | 2 | 6.25 |
| Total number of animals analysed | 319 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santamaría, P.G.; Dubus, P.; Bustos-Tauler, J.; Floristán, A.; Vázquez-Naharro, A.; Morales, S.; Cano, A.; Portillo, F. Loxl2 and Loxl3 Paralogues Play Redundant Roles during Mouse Development. Int. J. Mol. Sci. 2022, 23, 5730. https://doi.org/10.3390/ijms23105730
Santamaría PG, Dubus P, Bustos-Tauler J, Floristán A, Vázquez-Naharro A, Morales S, Cano A, Portillo F. Loxl2 and Loxl3 Paralogues Play Redundant Roles during Mouse Development. International Journal of Molecular Sciences. 2022; 23(10):5730. https://doi.org/10.3390/ijms23105730
Chicago/Turabian StyleSantamaría, Patricia G., Pierre Dubus, José Bustos-Tauler, Alfredo Floristán, Alberto Vázquez-Naharro, Saleta Morales, Amparo Cano, and Francisco Portillo. 2022. "Loxl2 and Loxl3 Paralogues Play Redundant Roles during Mouse Development" International Journal of Molecular Sciences 23, no. 10: 5730. https://doi.org/10.3390/ijms23105730
APA StyleSantamaría, P. G., Dubus, P., Bustos-Tauler, J., Floristán, A., Vázquez-Naharro, A., Morales, S., Cano, A., & Portillo, F. (2022). Loxl2 and Loxl3 Paralogues Play Redundant Roles during Mouse Development. International Journal of Molecular Sciences, 23(10), 5730. https://doi.org/10.3390/ijms23105730