
Mass Spectrometry-Based Disulfide Mapping of Lysyl Oxidase-like 2


Abstract
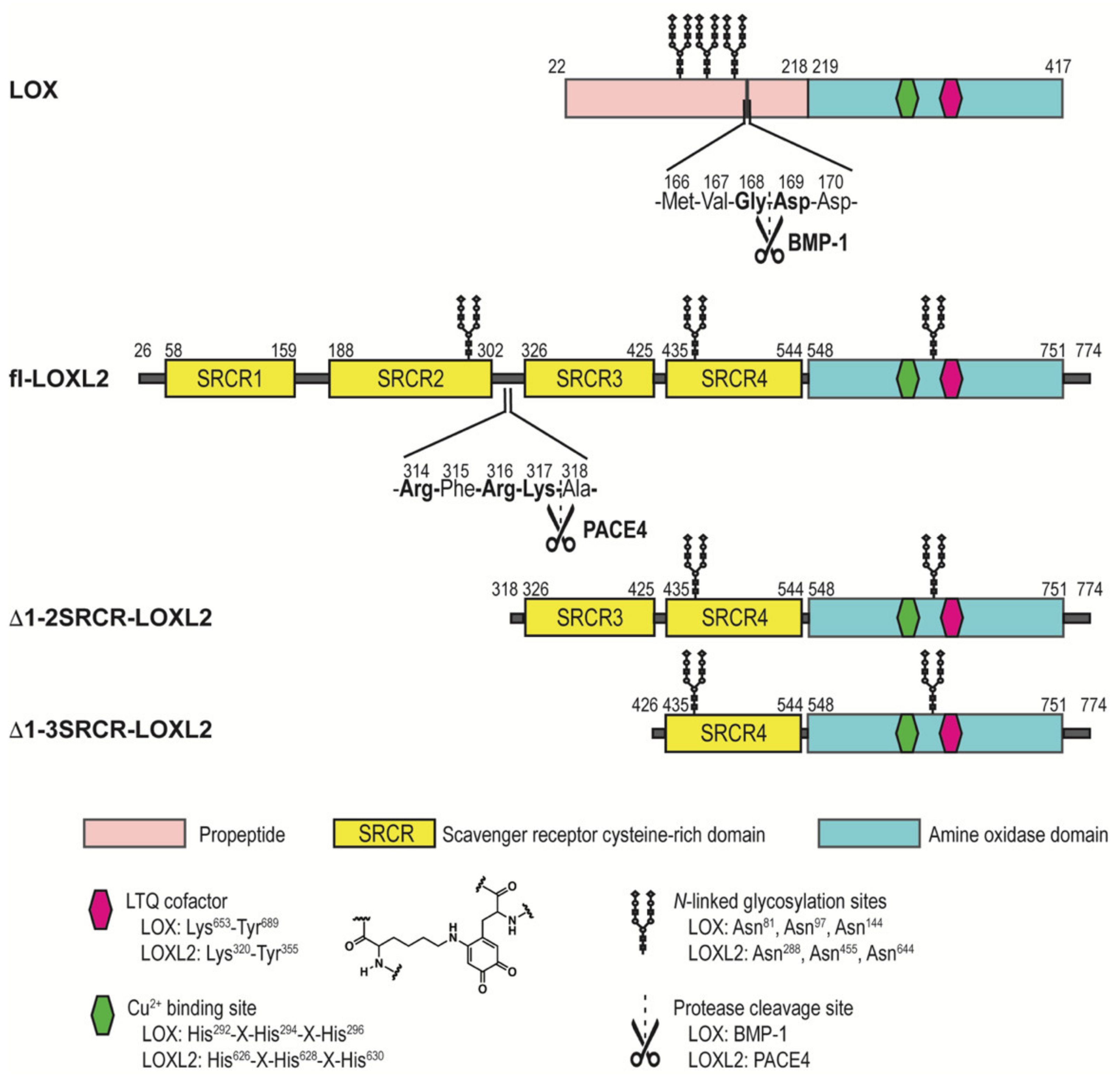
1. Introduction
2. Results
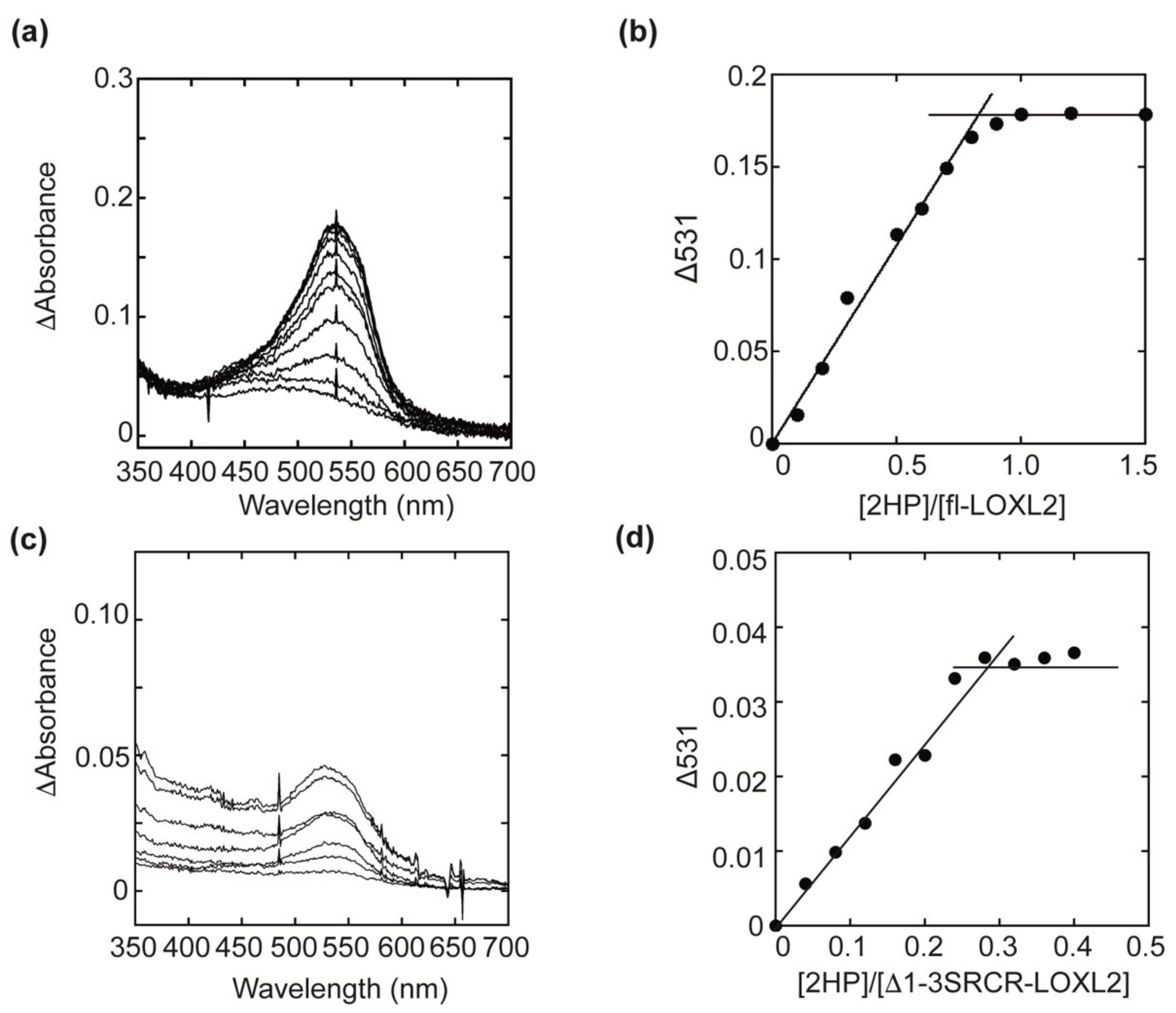
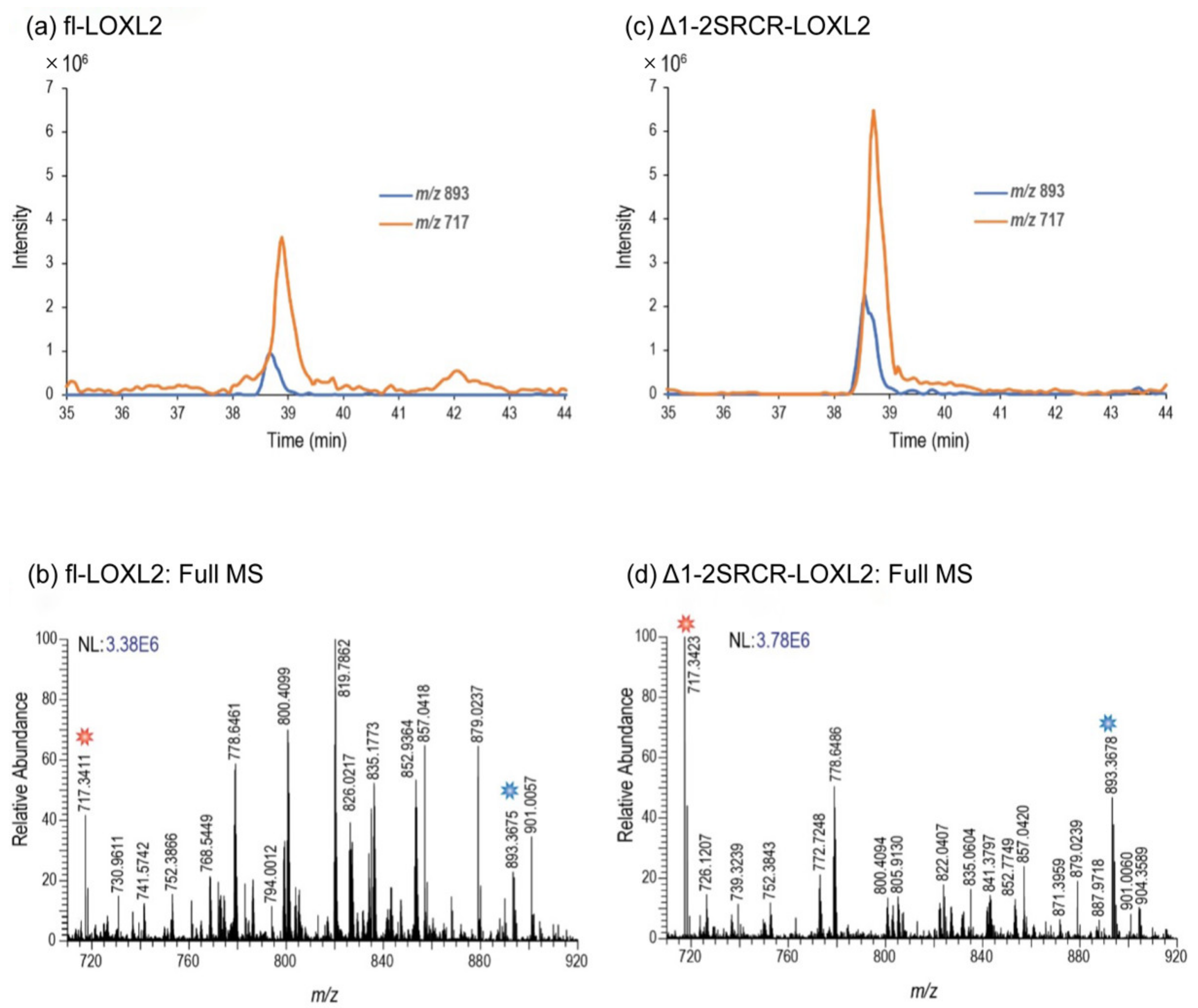
2.1. Quantitation of the LTQ Cofactor in fl-LOXL2 and Δ1-3SRCR-LOXL2
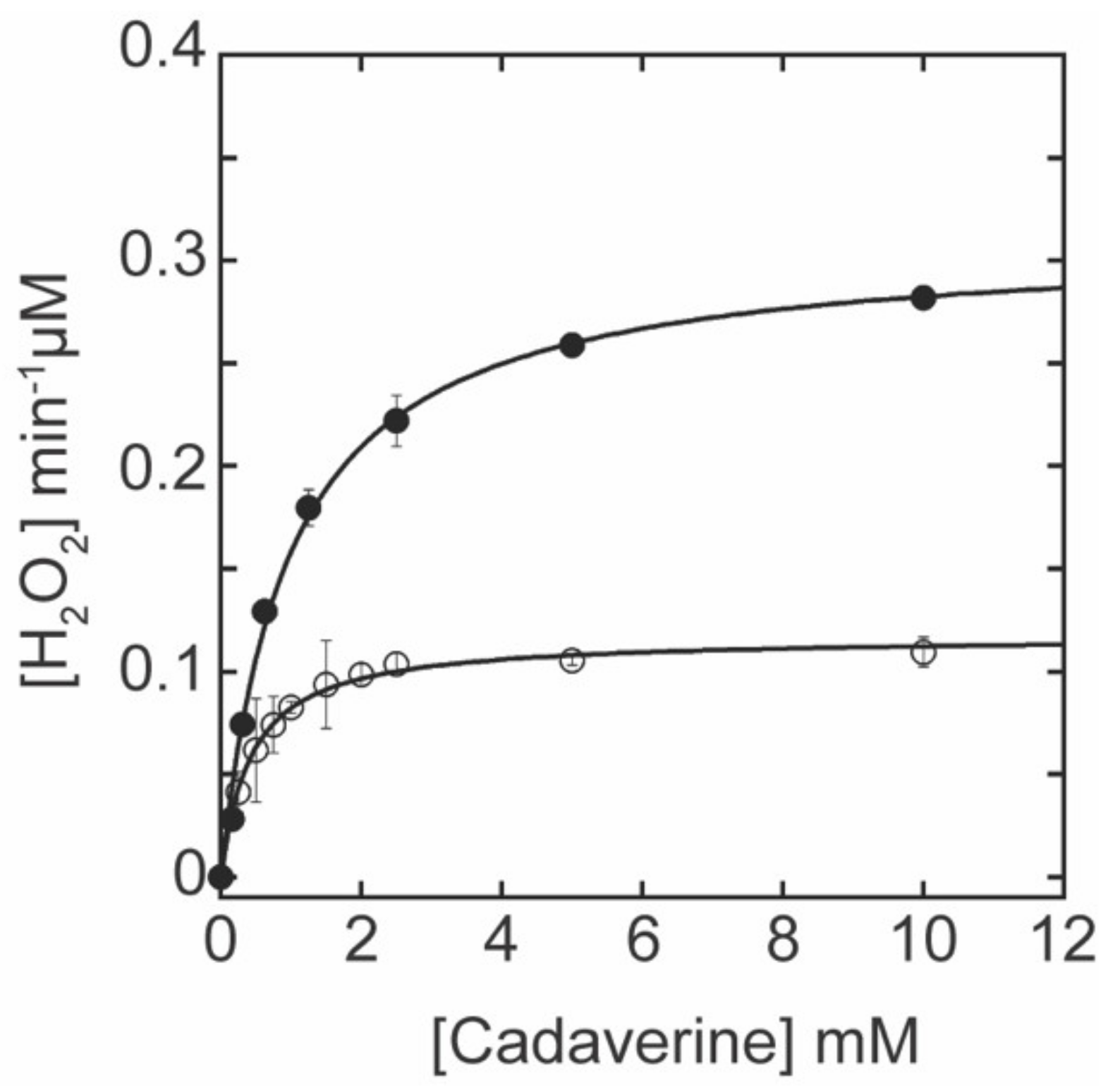
2.2. Correlation between the LTQ Content and the Amine Oxidase Activity of fl-LOXL2 and Δ1-3SRCR-LOXL2
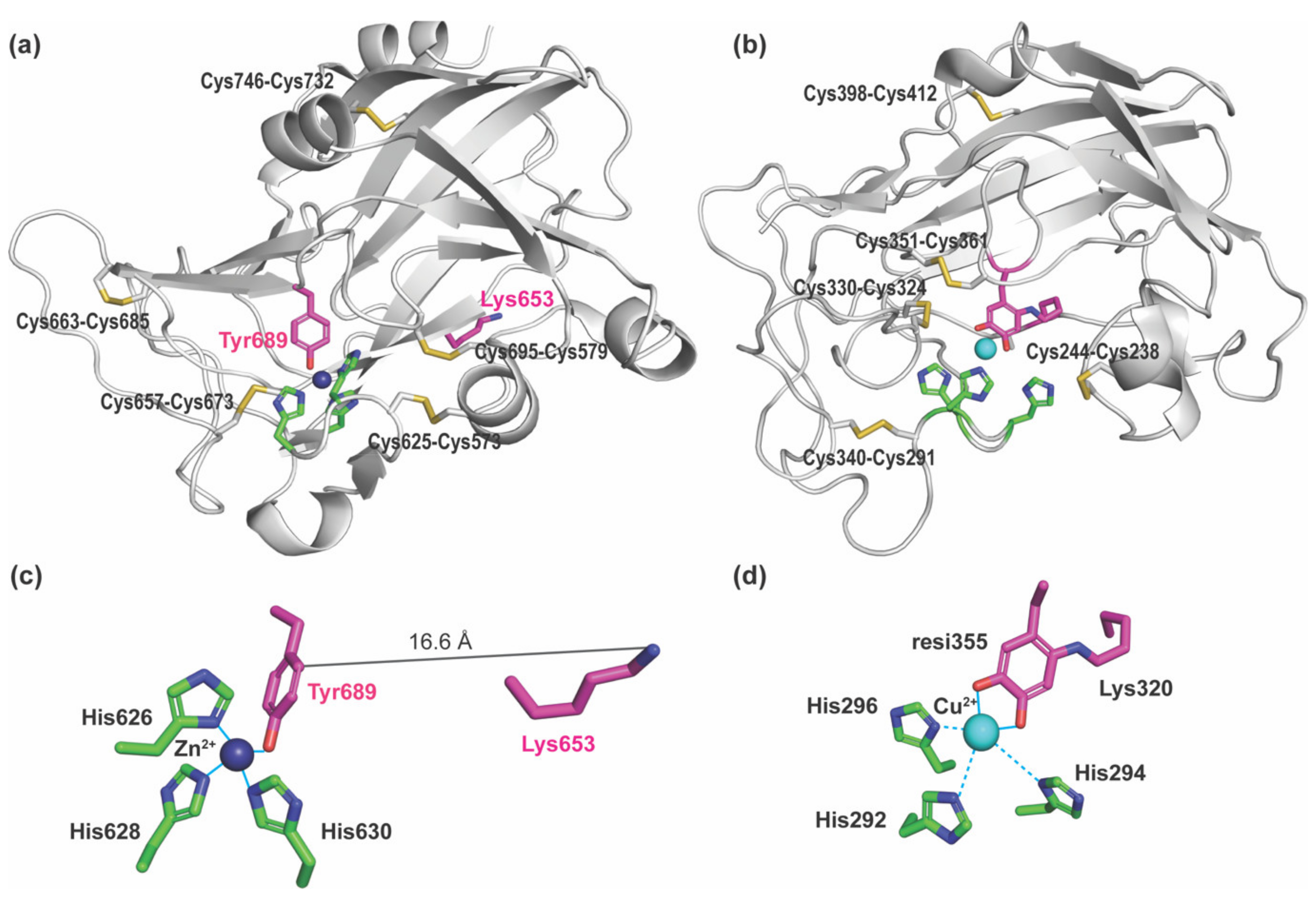
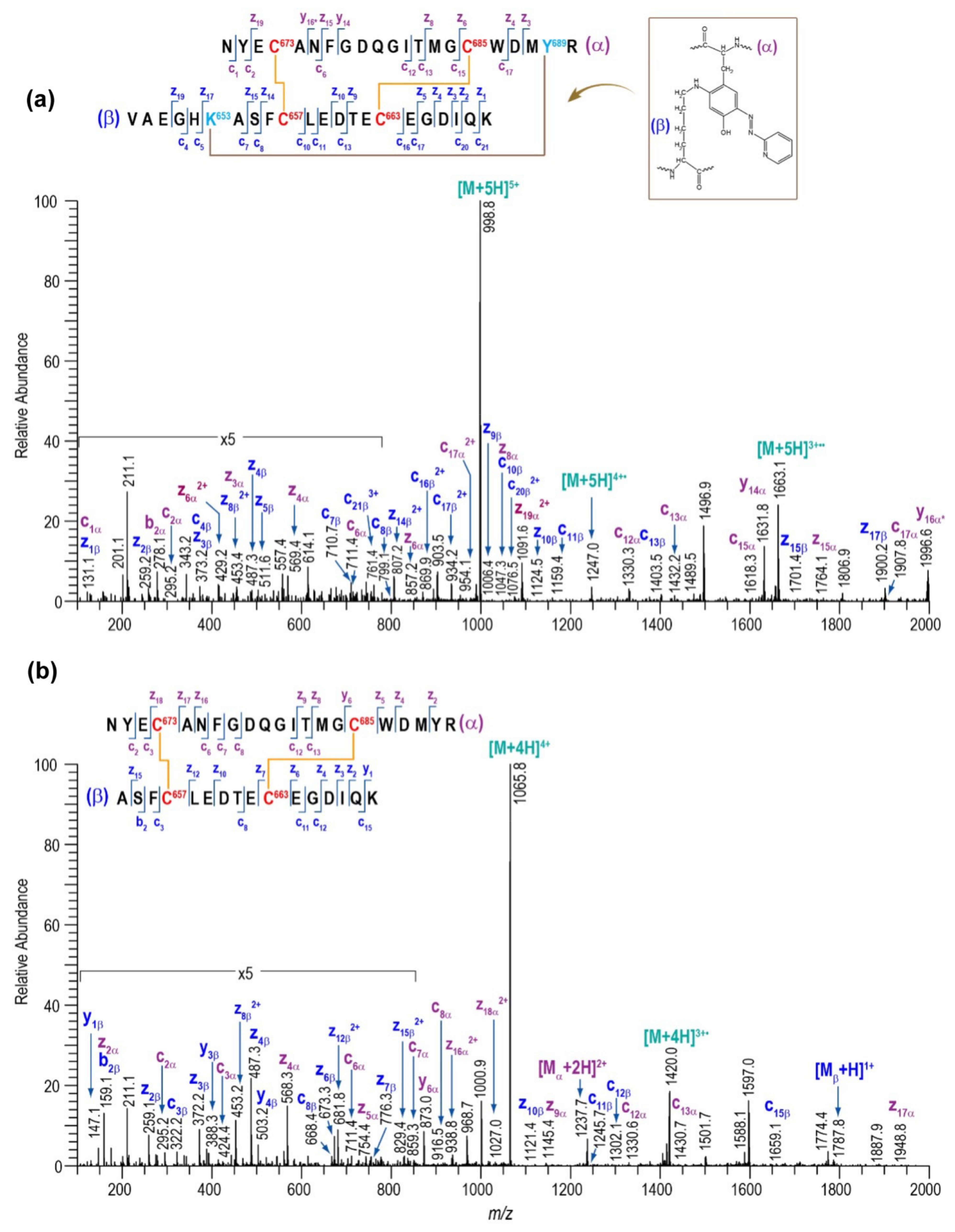
2.3. Assignment of Disulfide-Containing Peptides
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of LOXL2 Samples for Mass Spectrometry-Based Disulfide Mapping
4.3. HP Titration of LTQ in Mature LOXL2
4.4. Amine Oxidase Activity Assay of LOXL2
4.5. Protease Digestion LOXL2 Proteins for Disulfide Analysis
4.6. Chromatography and Mass Spectrometry
4.7. Disulfide Analysis of LOXL2 Proteins
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AGAO | Arthrobacter globiformis phenylethylamine oxidase |
| AUC | analytical ultracentrifugation |
| BMP-1 | bone morphogenetic protein-1 |
| CAO | copper amine oxidase |
| CH3CN | acetonitrile |
| Cys | cysteine |
| Da | Dalton |
| DES™ | Drosophila expression system |
| DPQ | dopaquinone |
| ECM | extracellular matrix |
| ETD | electron transfer dissociation |
| FDR | false discovery rate |
| Gln | glutamine |
| 2HP | 2-hydrazinopyridine |
| His | histidine |
| HRP | horseradish peroxidase |
| LC | liquid chromatography |
| Lys | lysine |
| LOX | lysyl oxidase |
| LOXL2 | lysyl oxidase-like 2 |
| LTQ | lysine tyrosylquinone |
| MS | mass spectrometry |
| MS/MS | tandem mass spectrometry |
| N-term | N-terminus |
| PTM | post-translational modification |
| Pyro-Glu | pyroglutamic acid |
| RFU | relative fluorescent unit |
| SRCR | scavenger receptor cysteine rich |
| TPQ | topaquinone |
| Tyr | tyrosine |
| XIC | extracted ion chromatograms |
References
- Csiszar, K. Lysyl oxidases: A novel multifunctional amine oxidase family. Prog. Nucleic Acid. Res. Mol. Biol. 2001, 70, 1–32. [Google Scholar] [PubMed]
- Moon, H.J.; Finney, J.; Ronnebaum, T.; Mure, M. Human lysyl oxidase-like 2. Bioorg. Chem. 2014, 57, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Salvador, F.; Martin, A.; Lopez-Menendez, C.; Moreno-Bueno, G.; Santos, V.; Vazquez-Naharro, A.; Santamaria, P.G.; Morales, S.; Dubus, P.R.; Muinelo-Romay, L.; et al. Lysyl Oxidase-like Protein LOXL2 Promotes Lung Metastasis of Breast Cancer. Cancer Res. 2017, 77, 5846–5859. [Google Scholar] [CrossRef] [PubMed]
- Park, P.G.; Jo, S.J.; Kim, M.J.; Kim, H.J.; Lee, J.H.; Park, C.K.; Kim, H.; Lee, K.Y.; Kim, H.; Park, J.H.; et al. Role of LOXL2 in the epithelial-mesenchymal transition and colorectal cancer metastasis. Oncotarget 2017, 8, 80325–80335. [Google Scholar] [CrossRef]
- Hong, X.; Yu, J.J. Silencing of lysyl oxidaselike 2 inhibits the migration, invasion and epithelialtomesenchymal transition of renal cell carcinoma cells through the Src/FAK signaling pathway. Int. J. Oncol. 2019, 54, 1676–1690. [Google Scholar] [CrossRef]
- Ninomiya, G.; Yamada, S.; Tatami, H.; Iwata, N.; Kanda, M.; Niwa, Y.; Hayashi, M.; Tanaka, C.; Kobayashi, D.; Fuji, T.; et al. Significance of lysil oxidase-like 2 (LOXL2) expression in hepatocellular carcinoma. HPB 2017, 19, S88. [Google Scholar] [CrossRef][Green Version]
- Mahjour, F.; Dambal, V.; Shrestha, N.; Singh, V.; Noonan, V.; Kantarci, A.; Trackman, P.C. Mechanism for oral tumor cell lysyl oxidase like-2 in cancer development: Synergy with PDGF-AB. Oncogenesis 2019, 8, 34. [Google Scholar] [CrossRef]
- Findlay, A.D.; Foot, J.S.; Buson, A.; Deodhar, M.; Jarnicki, A.G.; Hansbro, P.M.; Liu, G.; Schilter, H.; Turner, C.I.; Zhou, W.; et al. Identification and Optimization of Mechanism-Based Fluoroallylamine Inhibitors of Lysyl Oxidase-like 2/3. J. Med. Chem. 2019, 62, 9874–9889. [Google Scholar] [CrossRef]
- Schilter, H.; Findlay, A.D.; Perryman, L.; Yow, T.T.; Moses, J.; Zahoor, A.; Turner, C.I.; Deodhar, M.; Foot, J.S.; Zhou, W.; et al. The lysyl oxidase like 2/3 enzymatic inhibitor, PXS-5153A, reduces crosslinks and ameliorates fibrosis. J. Cell Mol. Med. 2019, 23, 1759–1770. [Google Scholar] [CrossRef]
- Chang, J.; Lucas, M.C.; Leonte, L.E.; Garcia-Montolio, M.; Singh, L.B.; Findlay, A.D.; Deodhar, M.; Foot, J.S.; Jarolimek, W.; Timpson, P.; et al. Pre-clinical evaluation of small molecule LOXL2 inhibitors in breast cancer. Oncotarget 2017, 8, 26066–26078. [Google Scholar] [CrossRef]
- Hutchinson, J.H.; Rowbottom, M.W.; Lonergan, D.; Darlington, J.; Prodanovich, P.; King, C.D.; Evans, J.F.; Bain, G. Small Molecule Lysyl Oxidase-like 2 (LOXL2) Inhibitors: The Identification of an Inhibitor Selective for LOXL2 over LOX. ACS Med. Chem. Lett. 2017, 8, 423–427. [Google Scholar] [CrossRef]
- Rowbottom, M.W.; Bain, G.; Calderon, I.; Lasof, T.; Lonergan, D.; Lai, A.; Huang, F.; Darlington, J.; Prodanovich, P.; Santini, A.M.; et al. Identification of 4-(Aminomethyl)-6-(trifluoromethyl)-2-(phenoxy)pyridine Derivatives as Potent, Selective, and Orally Efficacious Inhibitors of the Copper-Dependent Amine Oxidase, Lysyl Oxidase-Like 2 (LOXL2). J. Med. Chem. 2017, 60, 4403–4423. [Google Scholar] [CrossRef]
- Smithen, D.A.; Leung, L.M.H.; Challinor, M.; Lawrence, R.; Tang, H.; Niculescu-Duvaz, D.; Pearce, S.P.; Mcleary, R.; Lopes, F.; Aljarah, M.; et al. 2-Aminomethylene-5-sulfonylthiazole Inhibitors of Lysyl Oxidase (LOX) and LOXL2 Show Significant Efficacy in Delaying Tumor Growth. J. Med. Chem. 2020, 63, 2308–2324. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Q.; Wu, J.; Wang, J.; Shi, Y.; Liu, M. Crystal structure of human lysyl oxidase-like 2 (hLOXL2) in a precursor state. Proc. Natl. Acad. Sci. USA 2018, 115, 3828–3833. [Google Scholar] [CrossRef]
- Mure, M.; Wang, S.X.; Klinman, J.P. Synthesis and characterization of model compounds of the lysine tyrosyl quinone cofactor of lysyl oxidase. J. Am. Chem. Soc. 2003, 125, 6113–6125. [Google Scholar] [CrossRef]
- Moore, R.H.; Spies, M.A.; Culpepper, M.B.; Murakawa, T.; Hirota, S.; Okajima, T.; Tanizawa, K.; Mure, M. Trapping of a dopaquinone intermediate in the TPQ cofactor biogenesis in a copper-containing amine oxidase from Arthrobacter globiformis. J. Am. Chem. Soc. 2007, 129, 11524–11534. [Google Scholar] [CrossRef]
- Vallet, S.D.; Guéroult, M.; Belloy, N.; Dauchez, M.; Ricard-Blum, S. A Three-Dimensional Model of Human Lysyl Oxidase, a Cross-Linking Enzyme. ACS Omega 2019, 4, 8495–8505. [Google Scholar] [CrossRef]
- Chen, X.; Greenaway, F.T. Identification of the disulfide bonds of lysyl oxidase. J. Neural Transm. 2011, 118, 1111–1114. [Google Scholar] [CrossRef]
- Papadopoulos, J.S.; Agarwala, R. COBALT: Constraint-based alignment tool for multiple protein sequences. Bioinformatics 2007, 23, 1073–1079. [Google Scholar] [CrossRef]
- Okada, K.; Moon, H.J.; Finney, J.; Meier, A.; Mure, M. Extracellular Processing of Lysyl Oxidase-like 2 and Its Effect on Amine Oxidase Activity. Biochemistry 2018, 57, 6973–6983. [Google Scholar] [CrossRef]
- Meier, A.A.; Moon, H.J.; Toth, R.t.; Folta-Stogniew, E.; Kuczera, K.; Middaugh, C.R.; Mure, M. Oligomeric States and Hydrodynamic Properties of Lysyl Oxidase-Like 2. Biomolecules 2021, 11, 1846. [Google Scholar] [CrossRef]
- Lakbub, J.C.; Shipman, J.T.; Desaire, H. Recent mass spectrometry-based techniques and considerations for disulfide bond characterization in proteins. Anal. BioAnal. Chem. 2018, 410, 2467–2484. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.F.; Go, E.P.; Desaire, H. Simple approach to assign disulfide connectivity using extracted ion chromatograms of electron transfer dissociation spectra. Anal. Chem. 2013, 85, 1192–1199. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lakbub, J.C.; Clark, D.F.; Shah, I.S.; Zhu, Z.; Go, E.P.; Tolbert, T.J.; Desaire, H. Disulfide Bond Characterization of Endogenous IgG3 Monoclonal Antibodies Using LC-MS: An Investigation of IgG3 Disulfide-mediated Isoforms. Anal. Methods 2016, 8, 6046–6055. [Google Scholar] [CrossRef] [PubMed]
- Rebecchi, K.R.; Go, E.P.; Xu, L.; Woodin, C.L.; Mure, M.; Desaire, H. A general protease digestion procedure for optimal protein sequence coverage and post-translational modifications analysis of recombinant glycoproteins: Application to the characterization of human lysyl oxidase-like 2 glycosylation. Anal. Chem. 2011, 83, 8484–8491. [Google Scholar] [CrossRef]
- Xu, L.; Go, E.P.; Finney, J.; Moon, H.; Lantz, M.; Rebecchi, K.; Desaire, H.; Mure, M. Post-translational Modifications of Recombinant Human Lysyl Oxidase-like 2 (rhLOXL2) Secreted from Drosophila S2 Cells. J. Biol. Chem. 2013, 288, 5357–5363. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- Misra, H.P.; Fridovich, I. The oxidation of phenylhydrazine: Superoxide and mechanism. Biochemistry 1976, 15, 681–687. [Google Scholar] [CrossRef]
- Hill, H.A.; Thornalley, P.J. Phenyl radical production during the oxidation of phenylhydrazine and in phenylphydrazine-induced haemolysis. FEBS Lett. 1981, 125, 235–238. [Google Scholar] [CrossRef]
- Go, E.P.; Cupo, A.; Ringe, R.; Pugach, P.; Moore, J.P.; Desaire, H. Native Conformation and Canonical Disulfide Bond Formation Are Interlinked Properties of HIV-1 Env Glycoproteins. J. Virol. 2015, 90, 2884–2894. [Google Scholar] [CrossRef]
- Dey, A.K.; Cupo, A.; Ozorowski, G.; Sharma, V.K.; Behrens, A.J.; Go, E.P.; Ketas, T.J.; Yasmeen, A.; Klasse, P.J.; Sayeed, E.; et al. cGMP production and analysis of BG505 SOSIP.664, an extensively glycosylated, trimeric HIV-1 envelope glycoprotein vaccine candidate. Biotechnol. Bioeng. 2018, 115, 885–899. [Google Scholar] [CrossRef]
- Go, E.P.; Zhang, Y.; Menon, S.; Desaire, H. Analysis of the disulfide bond arrangement of the HIV-1 envelope protein CON-S gp140 DeltaCFI shows variability in the V1 and V2 regions. J. Proteome Res. 2011, 10, 578–591. [Google Scholar] [CrossRef]
- Go, E.P.; Hua, D.; Desaire, H. Glycosylation and disulfide bond analysis of transiently and stably expressed clade C HIV-1 gp140 trimers in 293T cells identifies disulfide heterogeneity present in both proteins and differences in O-linked glycosylation. J. Proteome Res. 2014, 13, 4012–4027. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LOXL2 1 | LOX 2 | LOX 3 |
|---|---|---|
| Cys573–Cys625 | Cys238–Cys244 | Cys573–Cys579 |
| Cys579–Cys695 | Cys291–Cys340 | Cys625–Cys673 |
| Cys657–Cys673 | Cys324–Cys330 | Cys657–Cys663 |
| Cys663–Cys685 | Cys351–Cys361 | Cys685–Cys695 |
| Cys732–Cys746 | Cys398–Cys412 | Cys732–Cys746 |
| LOXL2 | LTQ (%) 1 | kcat (min−1) | KM (mM) | kcat/KM (min−1mM−1) |
|---|---|---|---|---|
| fl-LOXL2 | 95 | 7.74 ± 0.19 | 0.96 ± 0.08 | 8.08 ± 0.69 |
| Δ1-3SRCR-LOXL2 | 27 | 2.93 ± 0.04 | 0.43 ± 0.03 | 6.86 ± 0.44 |
| Peptide (#) | Domain | Disulfide | Disulfide-Linked Peptides |
|---|---|---|---|
| 1 | 1st SRCR | Cys84–Cys148 |  |
| 2 | Cys97–Cys158 |  | |
| 3 | Cys128–Cys138 |  | |
| 4 | 2nd SRCR | Cys218–Cys291 + Cys231–Cys301 |  |
| 5 | Cys265–Cys275 |  | |
| 6 | 3rd SRCR | Cys351–Cys414 |  |
| 7 | Cys364–Cys424 |  | |
| 8 | Cys395–Cys405 |  | |
| 9 | 4th SRCR | Cys464–Cys530 + Cys477–Cys543 |  |
| 10 | Cys511–Cys521 |  | |
| 11 | Amine oxidase domain | Cys573–Cys625 + Cys579–Cys695 |  |
| 12a 12b | Cys657–Cys673 + Cys663–Cys685 |  | |
| 13 | Cys732–Cys746 |  |
| Peptide (#) | Domain | Disulfide | Δ1-2SRCR-LOXL2 | Δ1-3SRCR-LOXL2 |
|---|---|---|---|---|
| Precursor | Mixture | |||
| 6 | SRCR3 | Cys351–Cys414 | ✓ | − |
| 7 | Cys364–Cys424 | ✓ | − | |
| 8 | Cys395–Cys405 | ✓ | − | |
| 9 | SRCR4 | Cys464–Cys530 + Cys477–Cys543 | ✓ | ✓ |
| 10 | Cys511–Cys521 | ✓ | ✓ | |
| 11 | Amine oxidase domain | Cys573–Cys625 + Cys579–Cys695 | ✓ | ✓ |
| 12 | Cys657–Cys673 + Cys663–Cys685 | 12a | 12a + 12b | |
| 13 | Cys732–Cys746 | ✓ | ✓ |
| Peptide (#) | Domain | Disulfide | Disulfide-Linked Peptides |
|---|---|---|---|
| 14 | 4th SRCR | Cys464–Cys477 |  |
| 15 | Cys530–Cys543 |  | |
| 16 | Amine oxidase domain | Cys573–Cys579 |  |
| 17 | Cys625–Cys695 |  | |
| 18 | Cys657–Cys663 |  | |
| 19 | Cys673–Cys685 |  |
| Peptide (#) | Domain | Disulfide | Δ1-3SRCR-LOXL2 | Δ1-2SRCR-LOXL2 | fl-LOXL2 |
|---|---|---|---|---|---|
| 14 | SRCR4 | Cys464–Cys477 | ✓ | ✓ | ✓ |
| 15 | Cys530–Cys543 | ✓ | ND | ND | |
| 16 | Amine oxidase domain | Cys573–Cys579 | ✓ | ✓ | ND |
| 17 | Cys625–Cys695 | ✓ | ND | ND | |
| 18 | Cys657–Cys663 | ✓ | ✓ | ✓ | |
| 19 | Cys673–Cys685 | ✓ | ✓ | ✓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meier, A.A.; Go, E.P.; Moon, H.-J.; Desaire, H.; Mure, M. Mass Spectrometry-Based Disulfide Mapping of Lysyl Oxidase-like 2. Int. J. Mol. Sci. 2022, 23, 5879. https://doi.org/10.3390/ijms23115879
Meier AA, Go EP, Moon H-J, Desaire H, Mure M. Mass Spectrometry-Based Disulfide Mapping of Lysyl Oxidase-like 2. International Journal of Molecular Sciences. 2022; 23(11):5879. https://doi.org/10.3390/ijms23115879
Chicago/Turabian StyleMeier, Alex A., Eden P. Go, Hee-Jung Moon, Heather Desaire, and Minae Mure. 2022. "Mass Spectrometry-Based Disulfide Mapping of Lysyl Oxidase-like 2" International Journal of Molecular Sciences 23, no. 11: 5879. https://doi.org/10.3390/ijms23115879
APA StyleMeier, A. A., Go, E. P., Moon, H.-J., Desaire, H., & Mure, M. (2022). Mass Spectrometry-Based Disulfide Mapping of Lysyl Oxidase-like 2. International Journal of Molecular Sciences, 23(11), 5879. https://doi.org/10.3390/ijms23115879