1. Introduction
Mycobacterium tuberculosis, the primary cause of human tuberculosis (TB) remains a leading cause of morbidity and mortality in humans. The World Health Organization estimated that 10 million people developed TB disease in 2019 resulting in 1.2 million deaths [
1]. Fueling these active infections is a reservoir of latent infections of approximately 23% of the world population [
2]. This global burden of disease and the rise in multidrug-resistant TB emphasize the need to characterize novel processes in the pathogenesis of
M. tuberculosis as potential targets for treatment strategies and antimicrobial design [
1].
For replication,
M. tuberculosis must acquire trace amounts of copper and other essential metals from its host. Copper functions in single-electron transfer reactions in enzymes including the cytochrome
aa3-bc1 respiration supercomplex of
M. tuberculosis [
3]. The gene
Rv2043 encodes cytochrome
aa3 subunit 1, which contains the copper-B catalytic site. Multiple saturating transposon mutagenesis screens indicate that
Rv2043 is essential [
4,
5,
6]; thus, copper is essential for the replication of this pathogen. Due to its high reactivity, copper is universally toxic in excess in both prokaryotic and eukaryotic cells, necessitating levels to be tightly controlled. Among the mechanisms underlying its toxic effects is the disruption of protein function through the formation of thiolate bonds with iron–sulfur centers [
7]. The sequestration and release of copper and other essential trace elements in different niches within host organisms to protect against pathogens has been termed nutritional immunity [
8]. The level of free copper ions in unstressed eukaryotic cells is estimated to be extremely low, below 10
−18 M [
9]. In contrast, this metal is concentrated within phagosomes of macrophages stimulated with pro-inflammatory molecules to form an antimicrobial environment. The stimulation of RAW 264.7 murine macrophages by lipopolysaccharide or IFNγ was shown to upregulate the copper importer CTR and increase trafficking of the copper transporter ATP7A to post-Golgi vesicles; these vesicles fuse with phagosomal compartments resulting in the ATP7A- and copper-dependent killing of
E. coli mutants that are unable to produce the copper-exporting P-type ATPase CopA [
10]. Additionally, the infection of murine C57BL/6 peritoneal macrophages with
M. tuberculosis bacilli increased copper levels to as high as 0.4 mM within phagosomes [
11]. Mutants of
M. tuberculosis unable to synthesize the cytoplasmic copper-storage metallothionein MymT, the cytoplasmic-membrane copper-efflux P-type ATPase CtpV, or the outer-membrane protein MctB were more sensitive to macrophage killing (reviewed in [
12]). Regulators of copper-toxicity-response genes in
M. tuberculosis include the copper-sensing transcriptional repressors CsoR and RicR [
13,
14,
15].
How
M. tuberculosis bacteria acquire copper inside the host is less clear, but components have begun to be identified. One regulator of copper acquisition is transcription sigma factor SigC. The short-term artificial induction of
sigC in
M. tuberculosis strain Erdman elevated the transcription of genes
Rv0096-Rv0101 and nearby
Rv0103/
ctpB, while prolonged
sigC induction further increased the expression of the same genes but also elevated copper-toxicity-response genes:
ricR,
mymT, and
Rv0846–Rv0850 [
16]. The deletion of
sigC impaired replication in copper-starved medium and attenuated virulence in SCID mice, supporting SigC as an important regulator of copper acquisition and pathogenesis [
16]. The genes
Rv0097-Rv0101 contribute to the synthesis of isonitrile lipopeptides and aid in zinc acquisition [
17,
18]. In silico analysis supports CtpB as a transmembrane P-type ATPase with a copper/silver binding motif [
19]. It was recently reported that the constitutive heterologous expression of CtpB from
M. tuberculosis strain H37Rv in fast-growing, non-pathogenic
M. smegmatis resulted in elevated ATPase activity in the CtpB-enriched plasma membrane when incubated with copper (I) ions and a tolerance of high levels (2–2.5 mM) of copper (II) ions [
20]. The same study reported that deleting
ctpB in avirulent
M. tuberculosis strain H37Ra did not alter growth phenotypes in Sauton medium supplemented with CuSO
4 (5 to 100 µM) or intracellular copper levels in the presence of 50 µM CuSO
4, which led to a conclusion that CtpB functions in copper transport, but possibly not in copper detoxification.
To elucidate the functions of CtpB, we sought to examine its potential roles in copper acquisition and virulence. Our studies reveal that the controlled induction of M. tuberculosis ctpB in M. smegmatis increases sensitivity to copper, while ctpB-deletion mutants of M. tuberculosis and Mycobacterium bovis Bacille Calmette-Guérin (BCG) develop growth defects only after serial subculture in medium lacking copper, suggesting that internal stores of the metal must be depleted for phenotypes to manifest. Surprisingly, deleting M. tuberculosis ctpB enhanced replication and virulence in a murine infection model, but reduced replication in adipocytes cultured without copper supplementation. Homologs of CtpB are only conserved in intracellular pathogenic mycobacteria, supporting the evolution of this protein to counter copper nutritional-immunity mechanisms.
3. Discussion
The essentiality of copper for the activity of key redox enzymes and the potential toxicity of free copper ions necessitate networked systems for sensing, regulating, importing, and chaperoning this highly reactive metal to cuproenzymes, storage molecules, or efflux pumps, depending on copper levels. The hydrophobic character of biological membranes prevents the free diffusion of polar molecules. For copper traversal of these barriers in bacteria, pore-forming channels can be formed in the outer membrane, or substrate-specific transport systems can be utilized in either membrane. A family of related outer-membrane porin proteins MspA, MspB, and MspC in
M. smegmatis were found to function in copper acquisition. The growth defect of a
M. smegmatis mspA mspC mutant in trace copper conditions was abrogated by introducing
mspA,
mspB, or
mspC; moreover, the heterologous overexpression of
mspA in
M. tuberculosis increased copper sensitivity and production of the periplasmic multicopper oxidase, MmcO, which oxidized Cu (I) to the less reactive Cu (II) state [
25]. How copper traverses the outer membrane of
M. tuberculosis remains unclear. This pathogen lacks homologs of the MspA family, while the deletion of
Rv1698, which encodes an identified outer-membrane channel protein MctB, increased copper sensitivity, supporting its function in copper export rather than import [
26].
When abundant, copper is likely imported across the cytoplasmic membrane through ubiquitous major-facilitator superfamily (MSF)-type transporters, which utilize energy from the proton gradient to selectively transport a wide range of substrates [
27]. The range of substrates transported by the thirty MSF family homologs identified in
M. tuberculosis have not been studied in detail; however, most have been linked to antibiotic resistance [
28]. Members of the MSF family appear to have low substrate specificity and affinity, allowing the transport of a variety of substrates [
29]. The presence of glutathione and other reductants in the bacterial cytosol reduce copper (II) to copper (I) [
30].
Cytoplasmic levels of free copper are kept low in part by copper-storage proteins, such as MymT in
M. tuberculosis, which binds up to six copper (I) ions [
31], and copper-efflux pumps. Bacteria have two types of P-type ATPases that couple copper (I) export to ATP hydrolysis: the CopA1 family comprises low-affinity, high-turnover transporters that function to efflux excess copper, while the CopA2 family comprises high-affinity, low-turnover pumps that export copper (I) to the periplasm for insertion into cuproenzymes, and some species have multiple CopA2 proteins that export to different cuproenzymes [
32]. Of the three
M. tuberculosis genes encoding copper-transporting P-type ATPases, the transcription of
ctpV was upregulated 150-fold in response to 0.5 mM copper, while
ctpA and
ctpB were slightly downregulated, and the deletion of
ctpV increased copper sensitivity [
19]. These observations support CtpV as a CopA1-type pump for rapid copper efflux. CtpA is a probable CopA2-type exporter for copper delivery to cuproenzymes. The expression of
M. tuberculosis ctpA in
M. smegmatis enhanced copper tolerance and increased ATPase activity of the membrane fraction only in response to copper (I) [
33]. When
M. tuberculosis bacilli encounter a high-copper environment, it alleviates the need for a high-affinity CopA2-type copper (I) exporter for insertion into cuproenzymes, such as the cyt
aa3-bc1 respiration supercomplex.
Our data are consistent with the function of CtpB as a copper-importing P-type ATPase or as a high-affinity CopA2-type exporter to more-efficiently deliver limited cytoplasmic copper to the essential cyt
aa3-bc1 respiration complex. CtpB functioning as a copper importer would be rare as most bacterial copper-transporting P-type ATPases export copper (I) from the cytoplasm [
32,
34]. However, Lewinson and colleagues reported HmtA as a P-type ATPase in
Pseudomonas aeruginosa that increases intracellular copper and zinc pools [
35] (REF). The gradual development of growth defects in
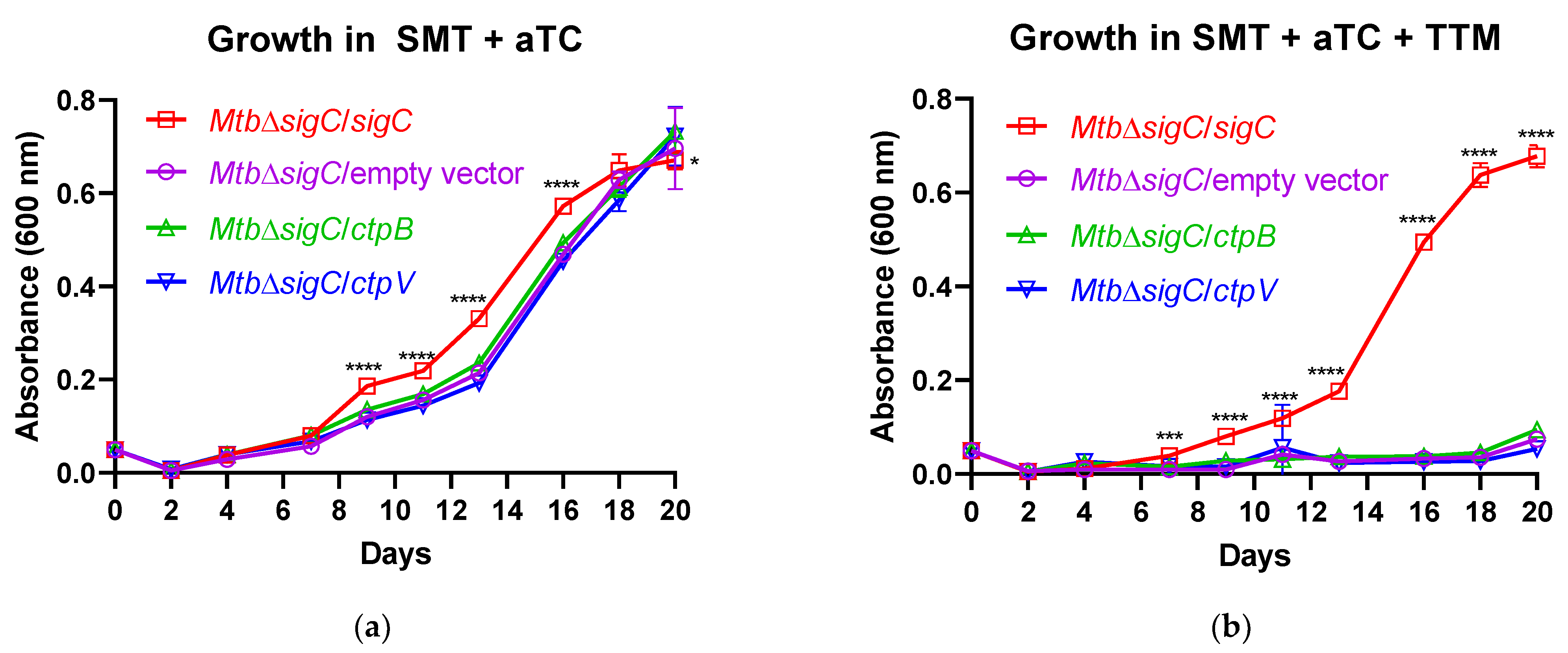
ctpB mutants after serial culture in copper-starved medium and reversal of the defects by copper supplementation (
Figure 3), and the reduced replication of the
M. tuberculosis ctpB mutant in adipocytes in the absence of copper supplementation (
Figure 4) support either hypothesis.
How does the loss of the ability to import or direct copper by the
ctpB mutant agree with the seemingly paradoxical increase in virulence and replication in the murine model? If CtpB enables
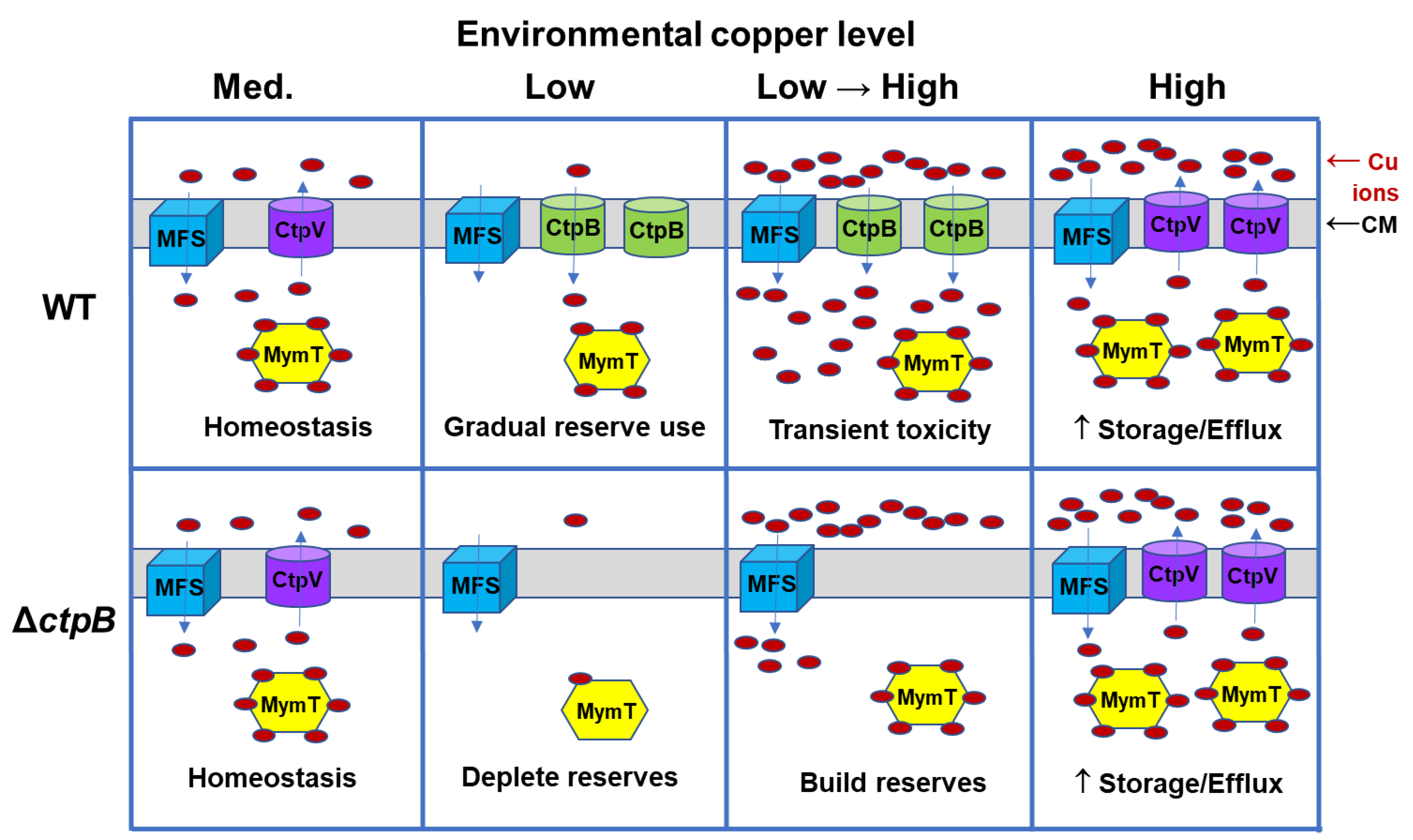
M. tuberculosis to import copper, it may contribute to copper overload within phagosomes. Cytosolic copper homeostasis relies on the balancing of copper import, storage, and export. Within a host, the pathogen will encounter environments with varied copper content, as illustrated with a simplistic model in
Figure 7. In a medium-copper niche, CtpB function is not required, as ample copper enters through MSF transporters, with some of the metal stored by binding to MymT, and the excess exported via CtpV (
Figure 7, column 1). In a low-copper environment, low-affinity MSF transporters are ineffective and production of CtpV is downregulated, while production of CtpB would occur to enable the import of scarce copper ions. Copper stores may begin to be used, but at a faster rate by a
ctpB mutant (
Figure 7, column 2). A rapid transition from low to high copper may occur upon phagocytosis by an activated macrophage. Neither the wild-type nor the mutant strain would have time to synthesize new CtpV, but only the wild-type strain expresses CtpB, resulting in copper intoxication due to influx through both MSF and CtpB, with subtoxic influx occurring in the
ctpB mutant (
Figure 7, column 3). To adapt to the high-copper environment, CtpB production would cease and CtpV and MymT production would increase for export and storage of the metal, respectively (
Figure 7, column 4). Bacteria that escape the phagosome have copper reserves to sustain the pathogen, as it again encounters copper-restricted niches. If CtpB instead functions to export copper to the cyt
bc1-aa3 supercomplex, then increased respiration would be expected upon the rapid shift to a high-copper phagosomal environment, which could result in enhanced production of reactive oxygen species.
Our findings are in support of CtpB function in copper import or directing copper to the
cyt bc1-aa3 complex and are consistent with the published
ctpB data. While León-Torres and colleagues examined the effects of
M. tuberculosis ctpB expression in
M. smegmatis and observed enhanced copper resistance [
20], we observed copper sensitivity (
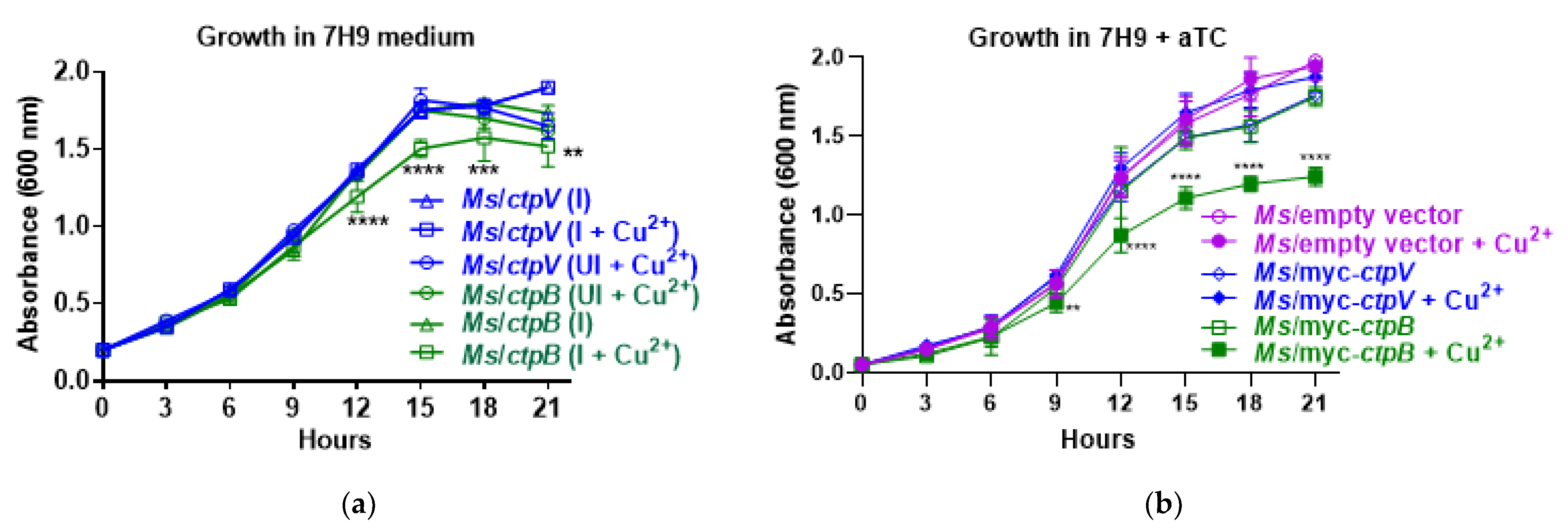
Figure 1). How can this be rationalized? A critical difference in our approach was the placement of the gene under control of an aTC-inducible promoter rather than the constitutive Hsp60 promoter. From a copper-scavenging perspective, the prolonged constitutive expression of
ctpB resulting in increased cytosolic copper would be sensed by the bacteria and lead to the upregulation of copper-efflux and detoxification mechanisms concomitant with the reported copper tolerance [
20]. In contrast, inducing
ctpB with aTC at the same time as copper supplementation enabled growth phenotypes to manifest before the bacteria had time to adapt to the elevated copper influx (
Figure 1). The lack of growth effects associated with the
M. tuberculosis H37Ra
ctpB mutant at varied copper levels [
20] is expected if the bacteria have not depleted internal copper stores. Growth defects of our
ctpB mutants of
M. tuberculosis Erdman and
M. bovis BCG only manifested after serial subculture in copper-deficient medium (
Figure 3). If we consider the phenotypes from the perspective of CtpB functioning to efficiently export scarce copper to respiration complexes, then increased reactive oxygen radicals would likely be generated when
ctpB is overexpressed, resulting in increased toxicity. Thus, the observations from both studies are consistent with CtpB functioning in either copper import or for directed export to cyt
bc1-aa3 supercomplex.
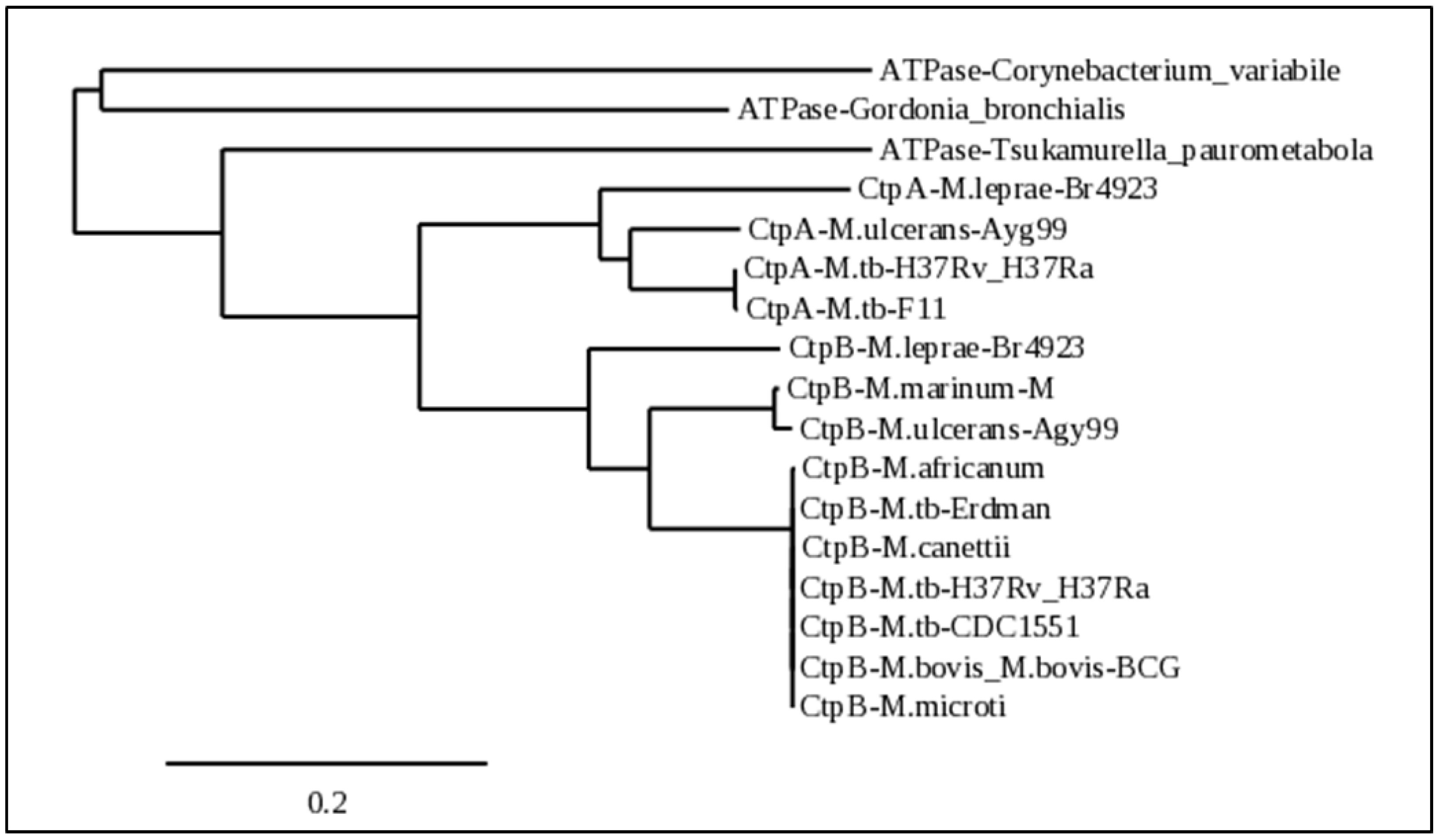
Phylogenetically, CtpB is highly conserved only among intracellular pathogenic Mycobacterium species (MTC, mycolactone clade, and M. leprae). It is also noteworthy that these same groups also have a closely-related copper (I)-exporting CtpA homolog. The evolution of a high-affinity copper (I)-importing function from the duplication and mutation of ctpA correlates with the enhanced ability of these mycobacteria to adapt to host nutritional immunity. In a copper-starved environment, these pathogens will eventually develop cytosolic reductive stress, which may lead to a greater reducing environment in the periplasm, where copper (II) may be reduced to copper (I) for import by CtpB, so that the metal can be more efficiently delivered to CtpA for export to the cyt aa3-bc1 respiration supercomplex. Alternatively, CtpB evolved as a second high-affinity CopA2-type exporter that outcompetes CtpA for cytoplasmic copper for directed export to the essential cyt aa3-bc1 respiration supercomplex, which would suggest that CtpA functions to transfer the metal to the supercomplex and to other cuproenzymes.
Structural and mutational analyses will help elucidate CtpB function and to identify chaperones that participate in copper trafficking in intracellular pathogenic mycobacteria. If CtpB imports copper, it will be particularly interesting to understand how the Post–Albers model for P-type ATPase function is influenced by the binding of a periplasmic copper chaperone. Additionally, in light of the essentiality of copper for
M. tuberculosis aerobic respiration, examination of the
ctpB mutant in an animal model that develops hypoxic granulomas will be informative. This human physiological niche is absent in most murine models, but is generated in response to
M. tuberculosis infection of nonhuman primates, rabbits, guinea pigs, and C3HeB/FeJ (Kramnik) mice [
36,
37].
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
Strains of mycobacteria were initially cultured in Middlebrook 7H9 broth or 7H10 agar (Thermo Scientific) supplemented with 0.5% glycerol, 0.05% Tween 80, and albumin–dextrose–saline (ADS) or oleic-acid–albumin–dextrose–saline (OADS) enrichment [
38]. For copper-response studies, strains were cultured in copper-free Sauton medium supplemented with 0.05% tyloxapol (SMT) [
39] or SMT supplemented with the indicated concentration of the copper (II) chelator tetrathiomolybdate (TTM) and/or copper salts at concentrations that were consistent with those reported for inactivated (limiting) or activated (inhibitory) intraphagosomal environments [
11]. Plasmids, strains, and primers used in these studies can be found in
Tables S1–S3, respectively. Transformation of
Mycobacterium species by electroporation employed standard techniques [
37]. Cultivation of
E. coli strains used Luria–Bertani broth and agar. When required, kanamycin was used at 50 µg/mL for
E. coli and 25 µg/mL for mycobacteria. Hygromycin was used at 50 µg/mL and gentamycin at 20 µg/mL in either
E. coli or mycobacteria. For induced expression of genes controlled from anhydrotetracycline-controlled promoters, the drug was used at the subclinical concentrations indicated.
Controlled expression of
M. tuberculosis genes in mycobacterial recipient host strains utilized the plasmid pOSN20. It was derived from plasmid pSR173 that was used previously for inducible biosynthesis of
M. tuberculosis SigC with an N-terminal
myc tag under control of the anhydrotetracycline-inducible promoter, P
myctetO [
22]. Plasmid pOSN20 was constructed by annealing primers P1705 and P1706 to yield the
E. coli lacZ promoter lacking the binding site for the catabolite-activator protein but containing the
lac operator site. The annealed fragment was ligated into the
SphI site of pSR173 upstream of the mycobacterial P
tb21 promoter preceding the tetracycline repressor gene,
tetR(B). This was performed to limit leaky expression in
E. coli of genes inserted into pOSN20 to replace
sigC. Plasmid pOSN19 was generated by removal of
sigC from plasmid pOSN20 by digestion with
SphI and
NsiI, blunting the DNA ends and re-ligating the vector. Plasmid pOSN37 was produced by PCR of
M. tuberculosis ctpB with primers P2059 and P2041 and insertion into pOSN20 digested with
PacI and
NsiI to replace
sigC, yielding a plasmid encoding a Myc-tagged CtpB translational fusion protein. Plasmid pOSN38 was similarly constructed by amplification of
M. tuberculosis ctpV with primers P2060 and P2061 and insertion into the pOSN20 digested with
PacI and
NsiI. DNA sequencing confirmed the absence of mutations in the inserted genes. Plasmid pOSN26 was produced by PCR of
M. tuberculosis ctpV with primers P1880 and P1881 and insertion pOSN20 after removal of
myc-sigC by
ScaI/
NsiI digestion. Plasmid pOSN30 was similarly made by PCR of
ctpB with primers P1605 and P1606 and insertion into the
ScaI/
NsiI-digested pOSN20.
For deletion of ctpB from M. tuberculosis and from M. bovis BCG, plasmid pOSN21 was created by sequential insertion of PCR products from ~1-kb genomic regions upstream and downstream of ctpB in M. tuberculosis, generated with primer pairs P2042/P2043 and P2044/P2045, respectively, into the AflI-XbaI and NcoI-BglII regions of pYUB854 flanking the hyg cassette. The ΔctpB::hyg allelic exchange substrate (AES) for deleting ctpB through recombineering was obtained by digestion of pOSN21 with BglII and XbaI. For ctpB complementation, plasmid pOSN5 was generated by PCR of the M. tuberculosis ctpB region from 209 bp upstream to 49 bp downstream with primers P1575 and P1576 and insertion into the HindIII site of plasmid pMV306. Plasmid pOSN40 was produced by replacement of the kanamycin-resistance gene of pOSN5 with the gentamycin-resistance element obtained from plasmid pPR27 by PCR with primers P1250 and P2251 and insertion into the NheI-SpeI region of pOSN5.
Deletion of
ctpB from
M. bovis BCG and
M. tuberculosis utilized mycobacterial recombineering [
39]. In brief, parent strains were transformed with plasmid pJV53 encoding mycobacteriophage Che9 recombinase proteins gp60 and gp61 under control of an acetamide-inducible promoter [
40]. The Δ
ctpB::hyg AES was electroporated into competent cells prepared after 12–16 h of growth in the presence of acetamide; hygromycin-resistant transformants were screened by PCR.
Strains of M. tuberculosis used for infection experiments were passaged three times in SMT and stored in 1 mL aliquots at −80 °C upon reaching OD600 = 1. To determine the bacterial counts of the M. tuberculosis infection stocks, all strains were thawed at 37 °C, harvested by centrifugation (10,000× g, 5 min), resuspended in phosphate-buffered saline (PBS), and sonicated in an ultrasonic water bath twice for 30 s. Serial dilutions of each strain were performed in PBS + 0.05% Tween 80 and plated onto 7H10ADStg agar. Colonies were enumerated after 3 to 4 weeks of incubation at 37 °C to determine the titers of the infection stocks.
4.2. Culture and Infection of Cells Lines
3T3-L1 fibroblast cells (ATCC
® CL-173) were cultured in DMEM basal medium supplemented with 10% heat-inactivated bovine calf serum (DMEM-BCS) at 37 °C and 5% CO
2 and replaced every three days with fresh pre-warmed medium. Differentiation of pre-adipocytes to adipocytes was started two days after confluency by stimulation of cells with 0.5 mM 3-isobutyl-1-methylxanthine (IBMX), 0.25 μM dexamethasone, 2 μM rosiglitazone, and 1 μg/mL insulin in DMEM plus 10% heat-inactivated fetal bovine serum (DMEM-FBS) using the methodology of Zebisch and colleagues [
41]. Two days later, the medium was replaced with DMEM-FBS with 1 μg/mL insulin. Adipocytes were maintained in DMEM-FBS that was changed twice per week. Adipocytes were infected at MOI = 1 using infection stocks prepared for the indicated
M. tuberculosis strains and cultured in DMEM-FBS alone or medium supplemented with 0.1 mM CuCl
2 or ZnCl
2. After 24 h incubation at 37 °C, monolayers were washed three times with PBS and fresh medium was applied. At the indicated time points, cells were lysed with 1% Triton X-100. To enumerate viable bacteria, cell lysates were serially diluted in phosphate-buffered saline (PBS) + 0.05% Tween 80, plated onto 7H10ADStg, and colonies counted after a 3-to-4-week incubation at 37 °C.
4.3. RNA Isolation and Quantitative RT-PCR
For RNA isolation from in vitro bacterial cultures,
M. tuberculosis strain Erdman was passaged three times in SMT, then replicate cultures were grown in SMT inoculated to OD
600 = 0.05. At OD
600 = 0.35, TTM was added to 20 µM to the half of the cultures. At OD
600 = 0.45, bacteria were harvested by centrifugation (10,000×
g, 5 min) and the pellets stored at −80 °C. RNA was isolated as described previously [
16]. For RNA isolation from the
M. tuberculosis-infected 3T3-L1 adipocytes, host cells infected at MOI = 1 were cultured in basal medium (DMEM-FBS 10%) alone or with addition of either 0.1 mM CuCl
2 or ZnSO
4 and incubated at 37 °C, 5% CO
2 with medium replaced every 3 days. At day 10 post-infection, cells were lysed by suspension in 4 M guanidine isothiocyanate and bacteria harvested by centrifugation (10,000×
g, 45 min). Bacterial RNA was isolated from the pellets using the methods described earlier for isolation from axenic cultures. For cDNA synthesis, 0.5 µg RNA was used with 200 ng random hexamers and Superscript III Reverse Transcriptase (Invitrogen) according to the manufacturer instructions. Quantitative PCR was performed using SYBR Green PCR Master Mix (Applied Biosystems), gene-specific primers, and prepared cDNA on a BioRad iCyler. For ΔΔCt analysis, gene expression was normalized to the
M. tuberculosis housekeeping gene
sigA [
16].
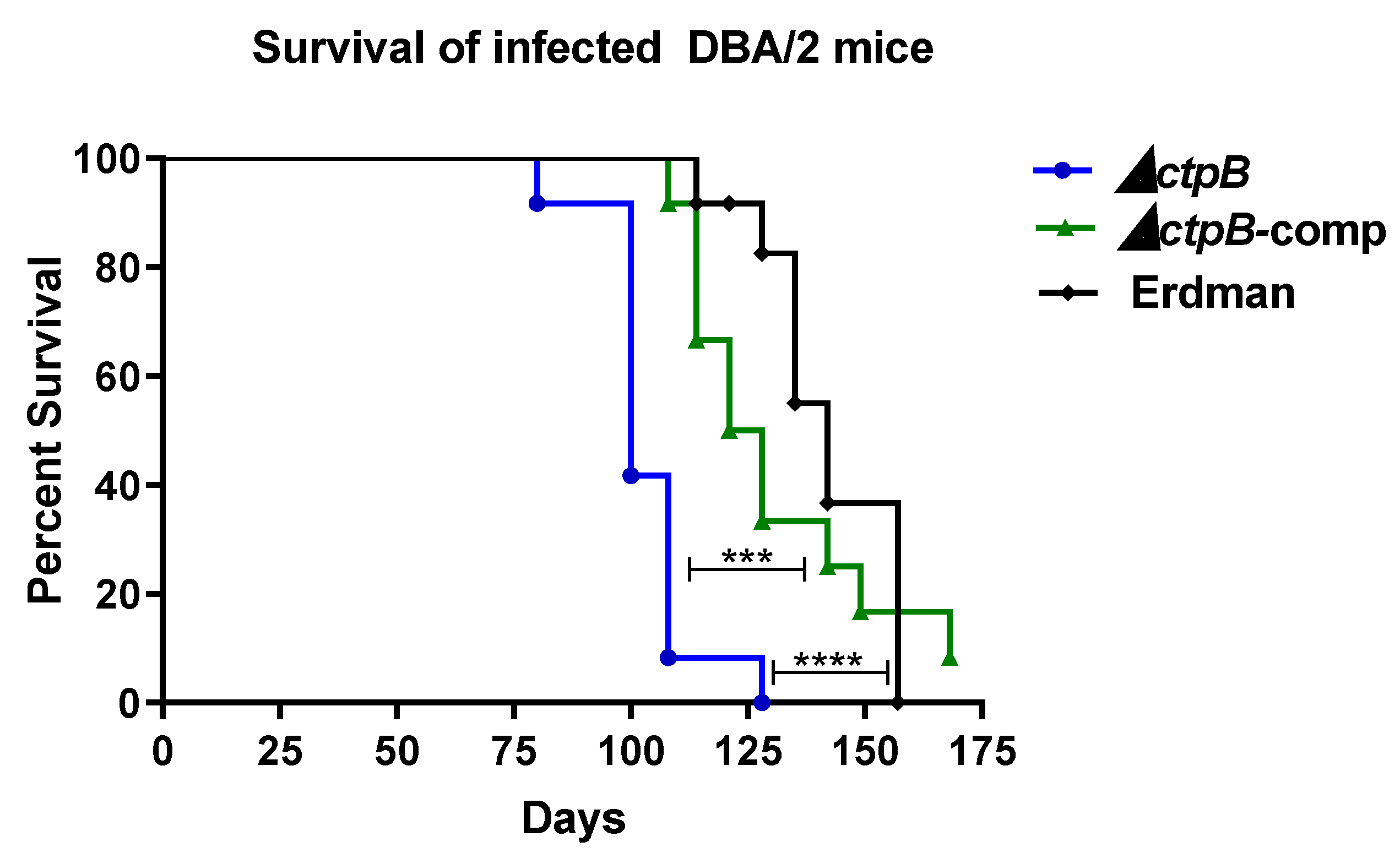
4.4. Murine Infection Studies
Groups of female, 8-week-old, DBA/2J mice (Jackson Laboratories, Bar Harbor, ME 04609) mice were intravenously infected via tail vein injection with 1–2 × 105 CFU M. tuberculosis strain ΔctpB, the complemented mutant (ΔctpB-comp), or Erdman. One day after infection, the mycobacterial lung burden was enumerated for one mouse in each group as follows: the lungs were harvested post-euthanasia, homogenized and plated on Middlebrook 7H11 agar supplemented with 10% ADS, 0.5% glycerol, and 0.05% Tween 80 (7H11gtADS), and resulting colonies counted after a 3-to-4-week incubation at 37 °C. The health of all animals was monitored daily, and mice were weighed weekly. Mice were euthanized when moribund. At necropsy, gross pathology of the lungs was examined. Half of the lungs were fixed in 10% neutral-buffered formalin for histopathological assessment, while the other half of the lungs as well as the spleen and liver were homogenized in PBS–Tween and serial dilutions plated on 7H11tgADS agar and bacterial counts enumerated after a 3-to-4-week incubation at 37 °C. For histopathology, formalin-fixed lungs were routinely processed for paraffin embedding, sectioning at 4 µm thickness, and staining with hematoxylin and eosin. Lung slides were assessed by a board-certified, veterinary pathologist using the following scoring system. Neutrophil (PMN) score: 1 = up to 25% of alveolar infiltrate composed of neutrophils, 2 = 26–50%, 3 = 51–75%, 4 = more than 75%; necrosis, alveolar, and alveolar edema scores: 1 = focal, 2 = multifocal, 3 = coalescing, 4 = diffuse; Vasculitis score: 1 = leukocytes infiltrating vessel wall, 2 = changes in 1 + clear spaces separating smooth muscle cells, 3 = changes in 2 + fibrinoid change, 4 = vessel wall effacement; perivascular cuffing (PVC) score: 1 = 1 layer of leukocytes surrounding vessel, 2 = 2–5 cells thick, 3 = 6–9 cells thick, 4 = 10 or more cells thick.
4.5. Bioinformatics
Sequences with homology to CtpB from
M. tuberculosis strain H37Rv were identified by a NCBI protein BLAST search launched through the Mycobrowser server (
https://mycobrowser.epfl.ch (accessed on 2 April 2022)). The searched database consisted of all non-redundant GenBank CDS translations + PDB + SwissProt + PIR + PRF excluding environmental samples from WGS projects. Additional CtpB homologs from mycobacterial strains of interest were obtained from Genbank. Sequences of representative homologs from various species were aligned using Phylogeny (
http://www.phylogeny.fr/index.cgi, accessed on 2 April 2022) to generate a phylogenetic tree. Colorized alignments of the mycobacterial strains used the phylogenetic tree were obtained by first aligning with Phylogeny and then pasting the fasta format alignment output into the Sequence Manipulator Site (
https://www.bioinformatics.org/sms2/index.html, accessed on 2 April 2022).
4.6. Statistical Analyses
Statistically significant differences between groups in bacterial growth studies utilized ANOVA with pair-wise comparisons. For the animal-survival comparisons, a log-rank (Mantel–Cox) test was applied to the Kaplan–Meier curves. All statistical analyses were performed using GraphPad Prism software. Differences between groups were deemed significant when p < 0.05.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}