
Intestinal Radiation Protection and Mitigation by Second-Generation Probiotic Lactobacillus-reuteri Engineered to Deliver Interleukin-22


 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
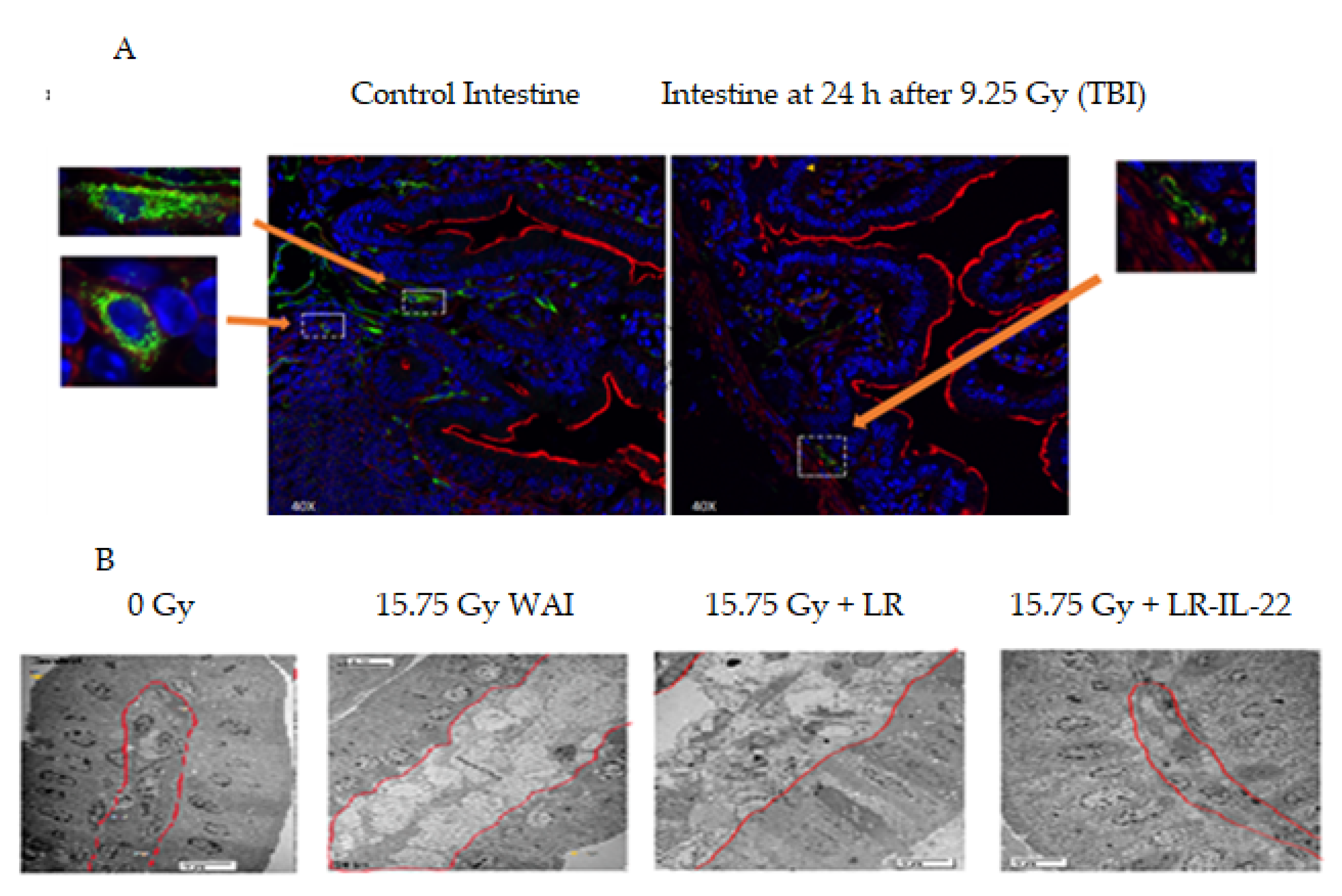
2.1. Effect of Gavage of LR-IL-22 on the Ionizing Irradiated Intestine
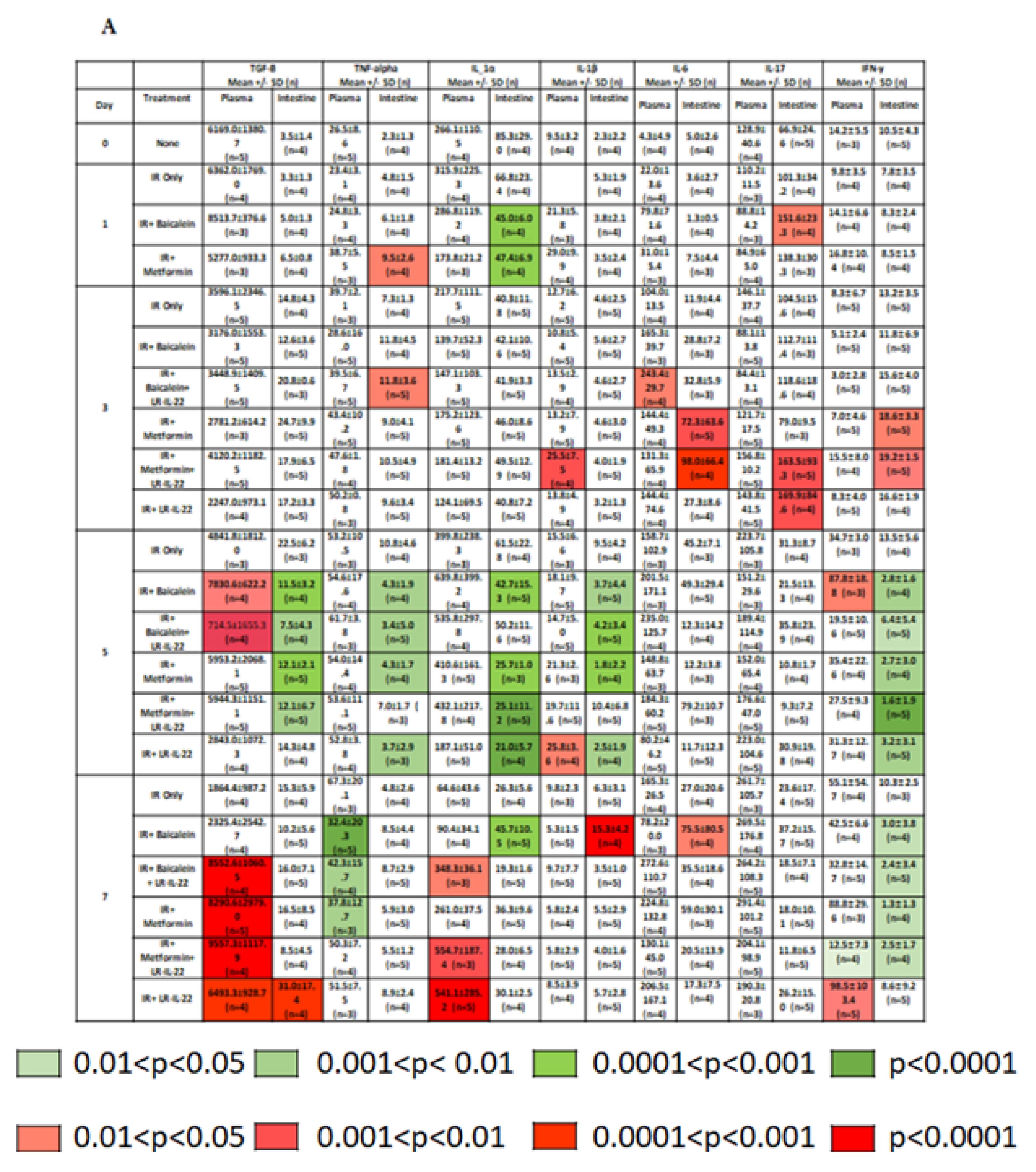
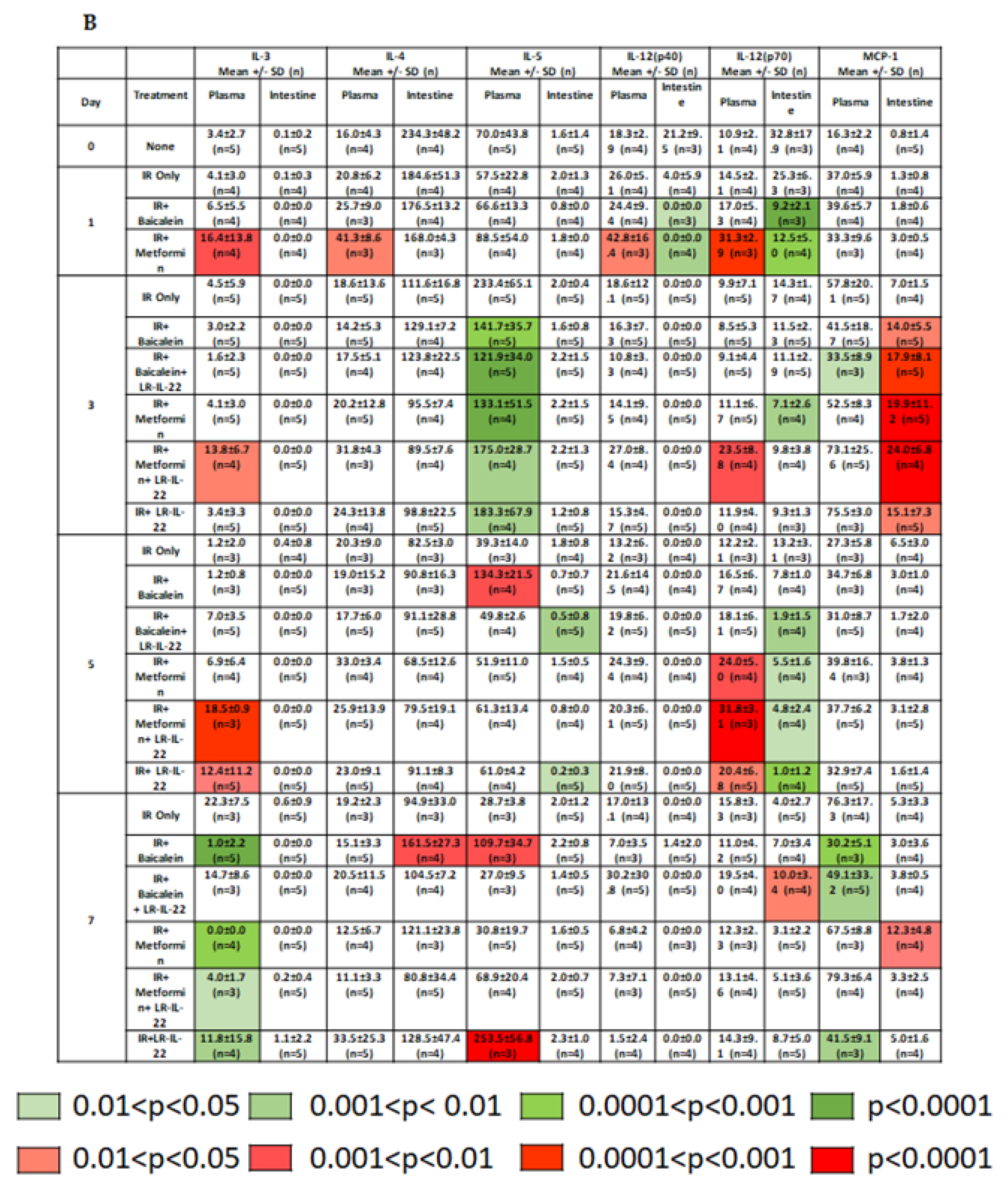
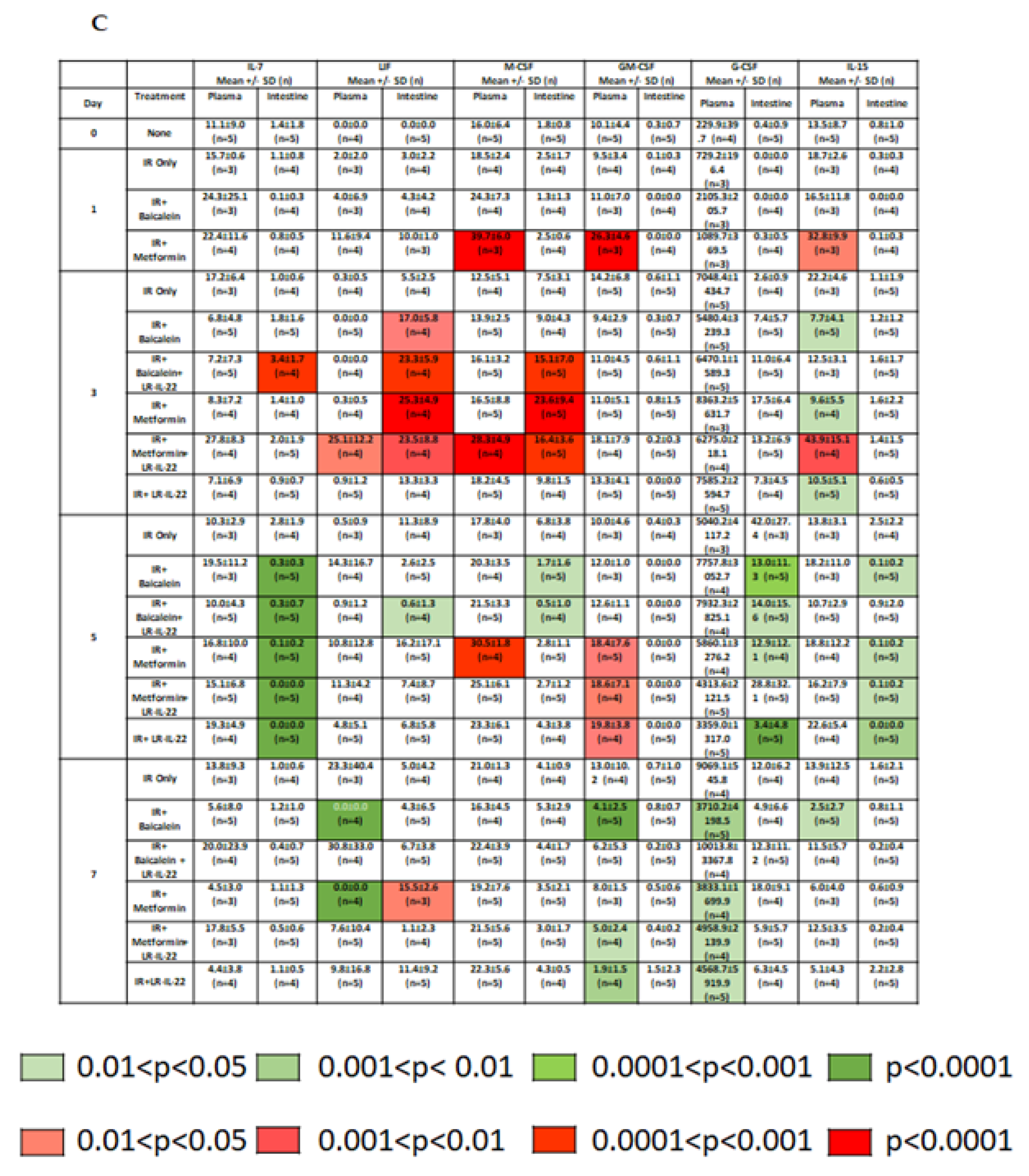
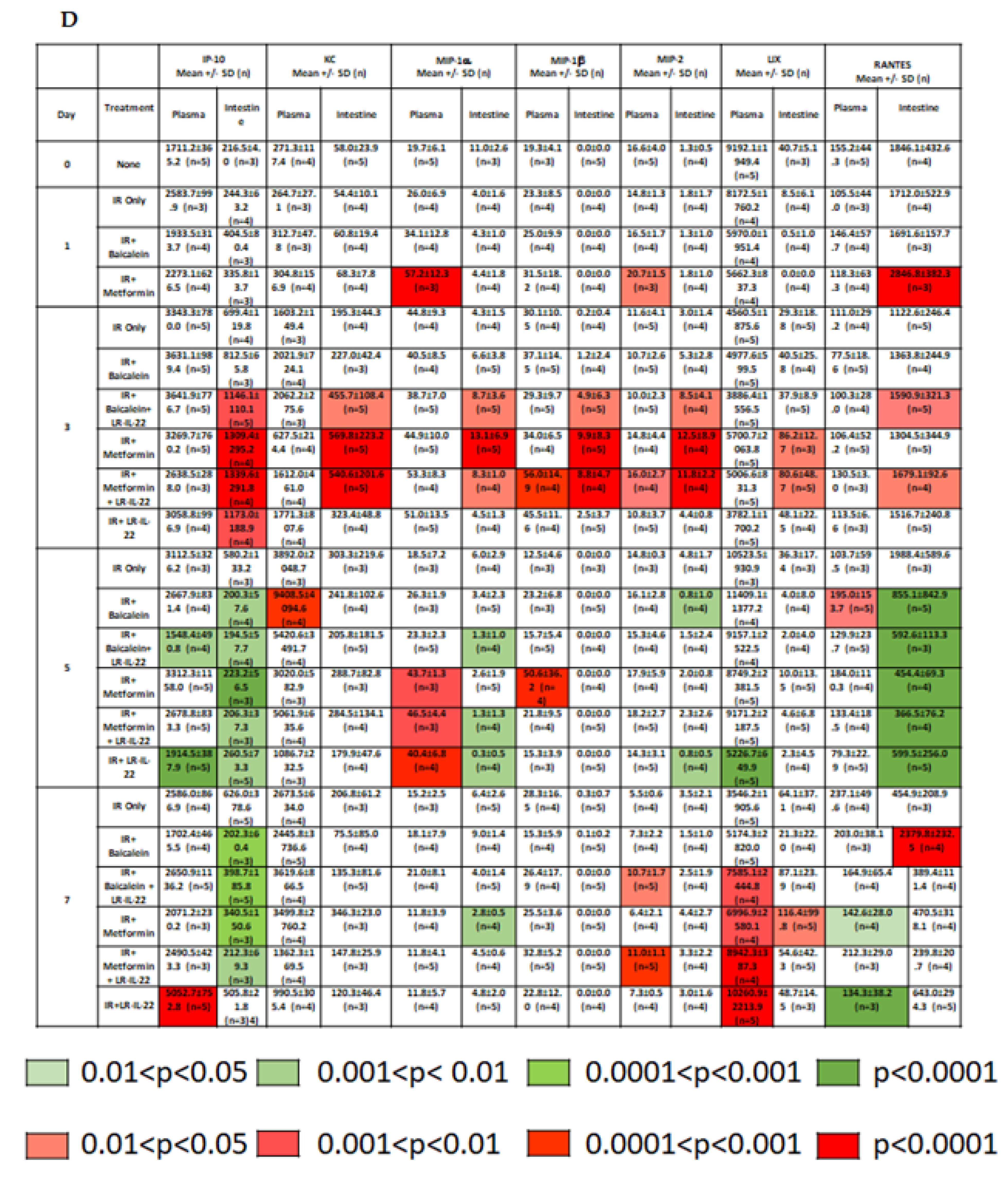
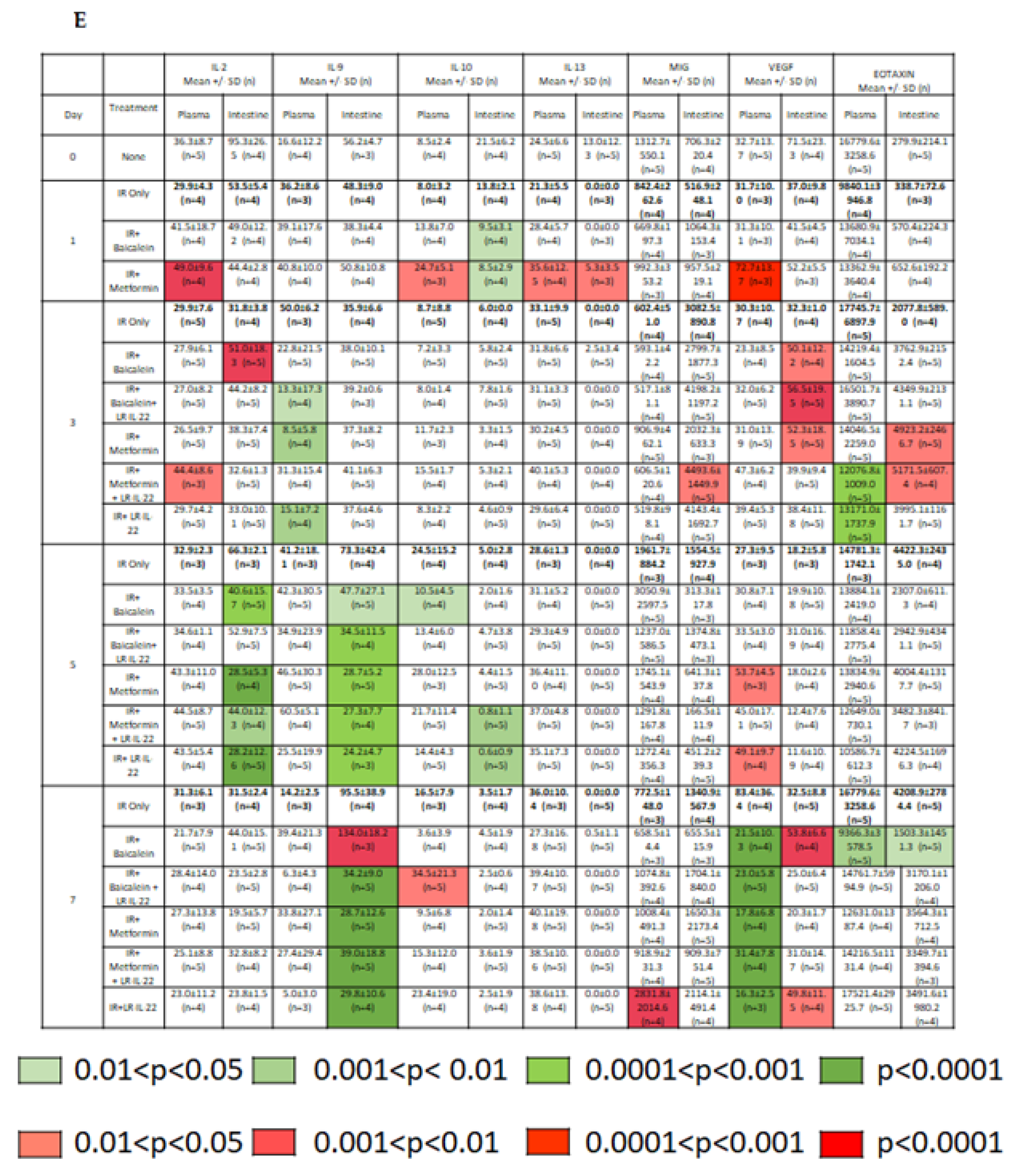
2.2. LR-IL-22 Produces Variable Alterations in Intestinal and Plasma Inflammatory Cytokines
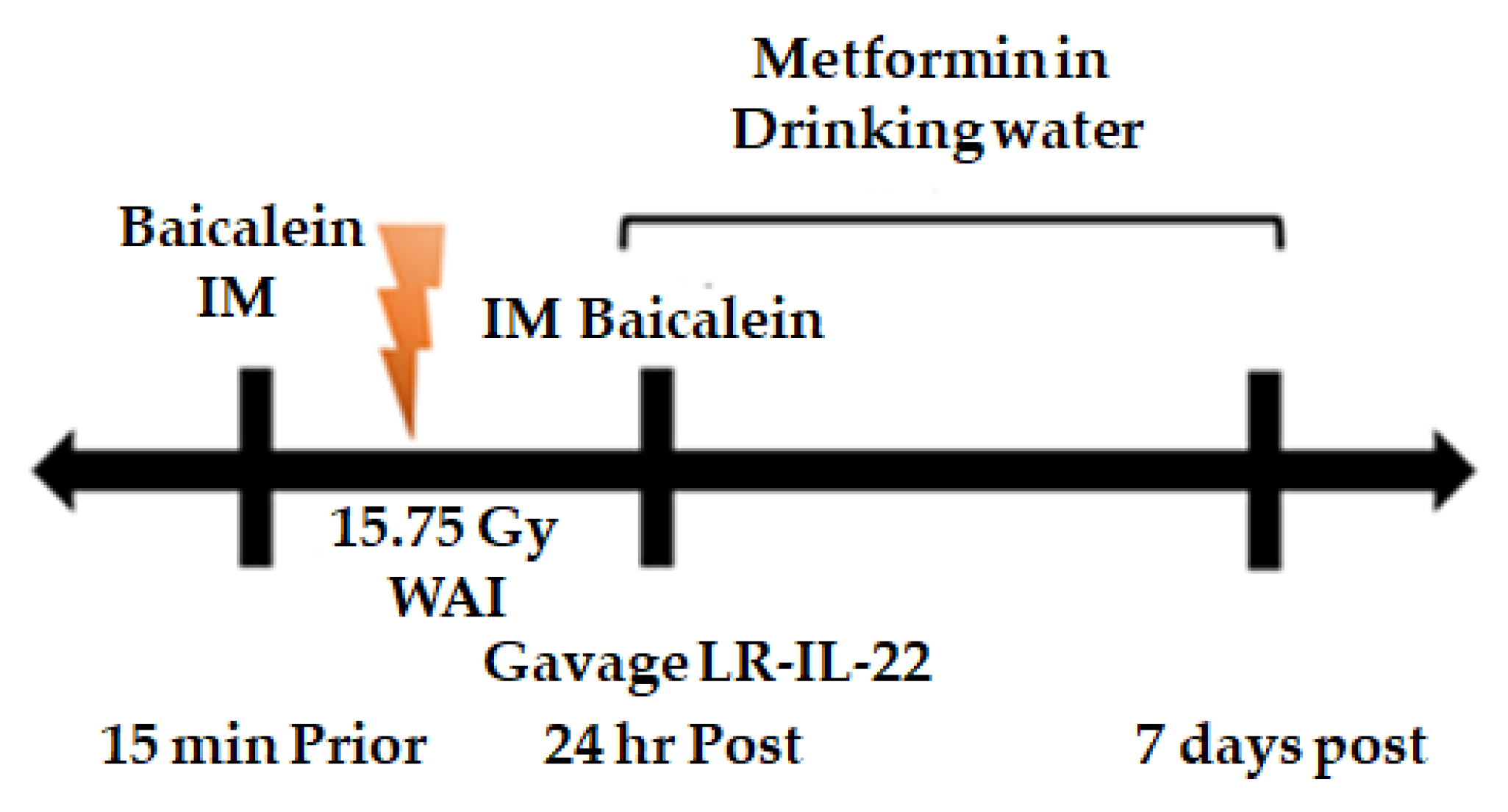
2.3. LR-IL-22 Gavage Mediated Radioprotection of the Intestine Is Enhanced by Systemic Administration of Small Molecule Radiation Mitigators
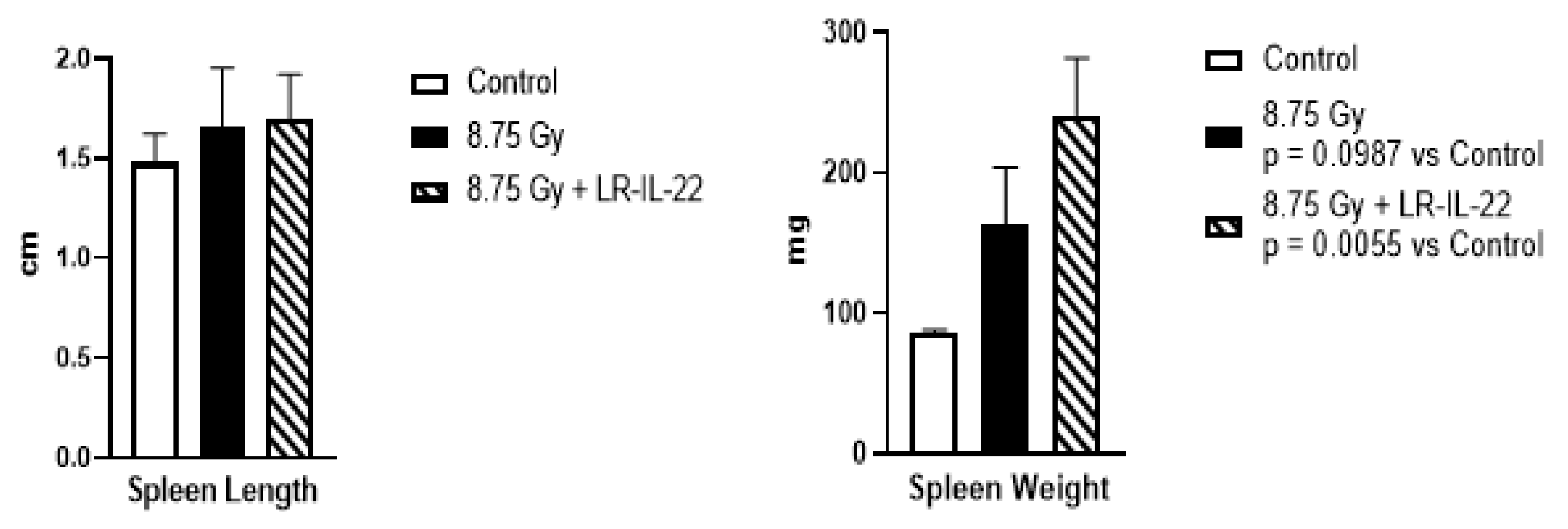
2.4. LR-IL-22 Effects on Spleen and the Immune System after Irradiation
2.5. Effect of LR-IL-22 on Bone Marrow
3. Discussion
Ionizing Irradiation Results in Dose and Volume Dependent Damage to Multiple Organs and Results in Discrete Syndromes Associated with Irradiation Exposure
4. Materials and Methods
4.1. Animal Care and Usage
4.2. Irradiation
4.3. Administration of Interleukin-22
4.4. Second Generation Probiotic Lactobacillus-reuteri-IL-22 (LR-IL-22)
4.5. Preparation and Delivery of Radiation Mitigator Drugs for Survival Analysis
4.6. Plasma and Intestinal Protein Level Measurements after WAI Irradiation and Administration of Small Molecule Radiation Mitigators for Luminex Assay
4.7. Preparation of Standards for Luminex Immunoassay
4.8. Procedure for 32-Multiplex Luminex Immunoassay and Standardization of Results
4.9. Procedures for TGF-beta-1 Immunoassay-Intestinal Samples
4.10. Procedures of TGF-beta-1 Immunoassay-Plasma Samples
4.11. Histopathologic Evaluation
4.12. Transmission Electron Microscopy
4.13. Spleen Tissue Collection and Cell Separation
4.14. Antibodies and Flow Cytometry
4.15. Statistics
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leibowitz, B.J.; Wei, L.; Zhang, L.; Ping, X.; Epperly, M.; Greenberger, J.; Cheng, T.; Yu, J. Ionizing irradiation induces acute haematopoietic syndrome and gastrointestinal syndrome independently in mice. Nat. Commun. 2014, 5, 3494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, L.; Leibowitz, B.J.; Wang, X.; Epperly, M.; Greenberger, J.; Zhang, L.; Yu, J. Inhibition of CDK4/6 protects against radiation-induced intestinal injury in mice. J. Clin. Investig. 2016, 126, 4076–4086. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Leibowitz, B.J.; Epperly, M.; Bi, C.; Li, A.; Steinman, J.; Wipf, P.; Li, S.; Zhang, L.; Greenberger, J.; et al. The GS-nitroxide JP4-039 improves intestinal barrier and stem cell recovery in irradiated mice. Sci. Rep. 2018, 8, 2072. [Google Scholar] [CrossRef] [PubMed]
- Leibowitz, B.J.; Zhao, G.; Wei, L.; Ruan, H.; Epperly, M.; Chen, L.; Lu, X.; Greenberger, J.S.; Zhang, L.; Yu, J. Interferon β drives intestinal regeneration after radiation. Sci. Adv. 2021, 7, eabi5253. [Google Scholar] [CrossRef]
- Paris, F.; Fuks, Z.; Kang, A.; Capodieci, P.; Juan, G.; Ehleiter, D.; Haimovitz-Friedman, A.; Cordon-Cardo, C.; Kolesnick, R. Endothelial apoptosis as the primary lesion initiating intestinal radiation damage in mice. Science 2001, 293, 293–297. [Google Scholar] [CrossRef]
- Rotolo, J.A.; Maj, J.G.; Feldman, R.; Ren, D.; Haimovitz-Friedman, A.; Cordon-Cardo, C.; Cheng, E.H.-Y.; Kolesnick, R.; Fuks, Z. Bax and Bak do not exhibit functional redundancy in mediating radiation-induced endothelial apoptosis in the intestinal mucosa. Int. J. Radiat. Oncol. Biol. Phys. 2008, 70, 804–815. [Google Scholar] [CrossRef]
- Bonnaud, S.; Niaudet, C.; Legoux, F.; Corre, I.; Delpon, G.; Saulquin, X.; Fuks, Z.; Gaugler, M.-H.; Kolesnick, R.; Paris, F. Sphingosine-1-phosphate activates the AKT pathway to protect small intestines from radiation-induced endothelial apoptosis. Cancer Res. 2010, 70, 9905–9915. [Google Scholar] [CrossRef] [Green Version]
- Rotolo, J.A.; Kolesnick, R.; Fuks, Z. Timing of lethality from gastrointestinal syndrome in mice revisited. Int. J. Radiat. Oncol. Biol. Phys. 2009, 73, 6–8. [Google Scholar] [CrossRef]
- Martin, M.L.; Adileh, M.; Hsu, K.-S.; Hua, G.; Lee, S.G.; Li, C.; Fuller, J.D.; Rotolo, J.A.; Bodo, S.; Klingler, S.; et al. Organoids reveal that inherent radiosensitivity of small and large intestinal stem cells determines organ sensitivity. Cancer Res. 2020, 80, 1219–1227. [Google Scholar] [CrossRef] [Green Version]
- Ch’ang, H.-J.; Maj, J.G.; Paris, F.; Xing, H.R.; Zhang, J.; Truman, J.-P.; Cardon-Cardo, C.; Haimovitz-Friedman, A.; Kolesnick, R.; Fuks, Z. ATM regulates target switching to escalating doses of radiation in the intestines. Nat. Med. 2005, 11, 484–490. [Google Scholar] [CrossRef]
- Steinman, J.; Epperly, M.; Hou, W.; Willis, J.; Wang, H.; Fisher, R.; Liu, B.; Bahar, I.; McCaw, T.; Kagan, V.; et al. Improved total-body irradiation survival by delivery of two radiation mitigators that target distinct cell death pathways. Radiat. Res. 2018, 189, 68–83. [Google Scholar] [CrossRef] [PubMed]
- Thermozier, S.; Hou, W.; Zhang, X.; Shields, D.; Fisher, R.; Bayir, H.; Kagan, V.; Yu, J.; Liu, B.; Bahar, I.; et al. Anti-ferroptosis drug enhances total body irradiation mitigation by drugs that block apoptosis and necroptosis. Radiat. Res. 2020, 193, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Greenberger, J.S.; Epperly, M.W. Radioprotective antioxidant gene therapy, Potential mechanisms of action. Gene Ther. Mol. Biol. 2004, 8, 31–44. [Google Scholar]
- Greenberger, J.S.; Epperly, M.W. Pleiotrophic stem cell and tissue effects of ionizing irradiation protection by MnSOD-plasmid liposome gene therapy. In Progress in Gene Therapy; Columbus, F., Ed.; Nova Science Publications: Hauppauge, NY, USA, 2005; pp. 110–118. [Google Scholar]
- Greenberger, J.S. Gene therapy approaches for stem cell protection. Gene Ther. 2008, 15, 100–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epperly, M.W.; Melendez, A.; Zhang, X.; Franicola, D.; Smith, T.; Greenberger, B.A.; Komanduri, P.; Greenberger, J.S. Mitochondrial targeting of a catalase transgene product by plasmid liposomes increases radioresistance in vitro and in vivo. Radiat. Res. 2009, 171, 588–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, E.J.; Giaccia, A.J. Radiobiology for the Radiologist, 6th ed.; Lippincott Williams & Wilkins: New York, NY, USA, 2006; pp. 106–117. [Google Scholar]
- Kalash, R.; Epperly, M.W.; Goff, J.; Dixon, T.; Sprachman, M.M.; Zhang, X.; Shields, D.; Cao, S.; Wipf, P.; Franicola, D.; et al. Amelioration of irradiation pulmonary fibrosis by a water-soluble bi-functional sulfoxide radiation mitigator (MMS350). Radiat. Res. 2013, 180, 474–490. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Fisher, R.; Hou, W.; Shields, D.; Epperly, M.W.; Wang, H.; Wei, L.; Leibowitz, B.J.; Yu, J.; Alexander, L.M.; et al. Second generation probiotics producing IL-22 increase survival of mice after total body irradiation. In Vivo 2020, 4, 39–50. [Google Scholar] [CrossRef]
- Crawford, P.A.; Gordon, J.I. Microbial regulation of intestinal radiosensitivity. Proc. Natl. Acad. Sci. USA. 2005, 102, 13254–13259. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Chou, W.-C.; Lai, Y.; Liang, K.; Tam, J.W.; Brickey, W.J.; Chen, L.; Montgomery, N.D.; Li, X.; Bohannon, L.M.; et al. Multi-omics analyses of radiation survivors identify radioprotective microbes and metabolites. Science 2020, 370, 549. [Google Scholar] [CrossRef]
- Li, K.; Epperly, M.W.; Barreto, G.A.; Greenberger, J.S.; Methe, B.A. Longitudinal fecal microbiome study of total body irradiated mice treated with radiation mitigators identify bacterial associations with survival. Front. Cell Infect. Microbiol. 2021, 11, 34621689. [Google Scholar] [CrossRef]
- Hamade, D.F.; Espinal, A.; Yu, J.; Leibowitz, B.; Fisher, R.; Hou, W.; Shields, D.; van Pijkeren, J.-P.; Mukherjee, A.; Epperly, M.W.; et al. Intestinal radioprotection by Lactobacillus reuteri synthesizing IL-22 (LR-IL-22) facilitates therapeutic whole abdomen irradiation (WAI). Radiat. Res. 2022. [Google Scholar] [CrossRef]
- Mortezaee, K.; Shabeeb, D.; Musa, A.E.; Najabi, M.; Farhood, B. Metformin as a radiation modifier: Implications to normal tissue protection and tumor sensitization. Curr. Clin. Pharmacol. 2019, 14, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Teng, M.; Xiao, T.; Baoding, Z.; Mengqi, L.; Yu, W.; Chunyan, Y.; Jianfeng, W.; Xiaoyan, W.; Qi, Q.; Yaxin, Y.; et al. Low-dose metformin targets the lysosomal AMPK pathway through PEN2. Nature 2022, 603, 159–165. [Google Scholar] [CrossRef]
- Dunn, C.J.; Peters, D.H. “Metformin. A review of its pharmacological properties and therapeutic use in non-insulin-dependent diabetes mellitus”. Drugs 1995, 49, 721–749. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Vogelzang, A.; Miyajima, M.; Sugiura, Y.; Wu, Y.; Chamoto, K.; Nakano, R.; Hatae, R.; Menzies, R.J.; Sonomura, K.; et al. B cell-derived GABA elicits IL-10+ macrophages to limit anti-tumour immunity. Nature 2021, 599, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Berhane, H.; Epperly, M.W.; Goff, J.; Kalash, R.; Cao, S.; Franicola, D.; Zhang, X.; Shields, D.; Houghton, F.; Wang, H.; et al. Radiobiologic differences between bone marrow stromal and hematopoietic progenitor cell lines from Fanconi Anemia (Fancd2-/-) mice. Radiat. Res. 2014, 181, 76–89. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.-B.; Holmgren, J.; Cragg, M.S.; Xiang, Z. Lack of Fc gamma receptor IIIA promotes rather than suppresses humoral and cellular immune responses after mucosal or parenteral immunization with antigen and adjuvants. Scand. J. Immunol. 2017, 85, 264–271. [Google Scholar] [CrossRef] [Green Version]
- Dittmann, K.H.; Rothmund, M.C.; Paasch, A.; Mayer, C.; Fehrenbacher, B.; Schaller, M.; Frauenstein, K.; Fritsche, E.; Haarmann-Stemmann, T.; Braeuning, A.; et al. The nuclear aryl hydrocarbon receptor is involved in regulation of DNA repair and cell survival following treatment with ionizing radiation. Toxicol. Lett. 2016, 240, 122–129. [Google Scholar] [CrossRef]
- Quintana, F.; Sherr, D.H. Aryl hydrocarbon receptor control of adaptive immunity. Pharmacol. Rev. 2013, 65, 1148–1161. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.S.; Cella, M.; McDonald, K.G.; Garlanda, C.; Kennedy, G.D.; Nukaya, M.; Mantovani, A.; Kopan, R.; Bradfield, C.A.; Newberry, R.D.; et al. AHR drives the development of gut ILC22 cells and postnatal lymphoid tissues via pathways dependent on and independent of Notch. Nat. Immunol. 2012, 13, 144–156. [Google Scholar] [CrossRef]
- Zelante, T.; Iannitti, R.G.; Cunha, C.; De Luca, A.; Giovannini, G.; Pieraccini, G.; Zecchi, R.; D’Angelo, C.; Massi-Benedetti, C.; Fallarino, F.; et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 2013, 39, 372–385. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.; Heller, J.J.; Guo, X.; Chen, Z.-M.E.; Fish, K.; Fu, Y.-X.; Zhou, L. The aryl hydrocarbon receptor regulates gut immunity through modulation of innate lymphoid cells. Immunity 2012, 36, 92–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Liu, Z.; Bloomer, B.J.; Clark, D.S.; Mukhopadhyay, A.; Keasling, J.D.; Hartwig, J.F. Unnatural biosynthesis by an engineered microorganism with heterologously expressed natural enzymes and an artificial metalloenzyme. Nat. Chem. 2021, 13, 1186–1192. [Google Scholar] [CrossRef] [PubMed]
- Canale, F.P.; Basso, C.; Antonini, G.; Perotti, M.; Li, N.; Sokolovska, A.; Neumann, J.; James, M.J.; Geiger, S.; Jin, W.; et al. Metabolic modulation of tumours with engineered bacteria for immunotherapy. Nature 2021, 598, 662–666. [Google Scholar] [CrossRef]
- Progatzky, F.; Shapiro, M.; Chng, S.H.; Garcia-Cassani, B.; Classon, C.H.; Sevgi, S.; Laddach, A.; Bon-Frauches, A.C.; Lasrado, R.; Rahim, M.; et al. Regulation of intestinal immunity and tissue repair by enteric glia. Nature 2021, 599, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Berhane, H.; Shinde, A.; Kalash, R.; Xu, K.; Epperly, M.W.; Goff, J.; Franicola, D.; Zhang, X.; Dixon, T.; Shields, D.; et al. Amelioration of irradiation induced oral cavity mucositis and distant bone marrow suppression in Fancd2-/- (FVB/N) mice by intraoral JP4-039/F15. Radiat. Res. 2014, 182, 35–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parks, O.B.; Pociask, D.A.; Hodzic, Z.; Kolls, J.K.; Good, M. Interleukin-22 signaling in the regulation of intestinal health and disease. Front. Cell Dev. Biol. 2016, 3, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meisel, M.; Hinterleitner, R.; Pacis, A.; Chen, L.; Earley, Z.M.; Mayassi, T.; Pierre, J.F.; Ernest, J.D.; Galipeau, H.J.; Thuille, N.; et al. Microbial signals drive pre-leukaemic myeloproliferation in a Tet2-deficien host. Nature 2018, 557, 580–584. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espinal, A.; Epperly, M.W.; Mukherjee, A.; Fisher, R.; Shields, D.; Wang, H.; Huq, M.S.; Hamade, D.F.; Vlad, A.M.; Coffman, L.; et al. Intestinal Radiation Protection and Mitigation by Second-Generation Probiotic Lactobacillus-reuteri Engineered to Deliver Interleukin-22. Int. J. Mol. Sci. 2022, 23, 5616. https://doi.org/10.3390/ijms23105616
Espinal A, Epperly MW, Mukherjee A, Fisher R, Shields D, Wang H, Huq MS, Hamade DF, Vlad AM, Coffman L, et al. Intestinal Radiation Protection and Mitigation by Second-Generation Probiotic Lactobacillus-reuteri Engineered to Deliver Interleukin-22. International Journal of Molecular Sciences. 2022; 23(10):5616. https://doi.org/10.3390/ijms23105616
Chicago/Turabian StyleEspinal, Alexis, Michael W. Epperly, Amitava Mukherjee, Renee Fisher, Donna Shields, Hong Wang, M. Saiful Huq, Diala Fatima Hamade, Anda M. Vlad, Lan Coffman, and et al. 2022. "Intestinal Radiation Protection and Mitigation by Second-Generation Probiotic Lactobacillus-reuteri Engineered to Deliver Interleukin-22" International Journal of Molecular Sciences 23, no. 10: 5616. https://doi.org/10.3390/ijms23105616
APA StyleEspinal, A., Epperly, M. W., Mukherjee, A., Fisher, R., Shields, D., Wang, H., Huq, M. S., Hamade, D. F., Vlad, A. M., Coffman, L., Buckanovich, R., Yu, J., Leibowitz, B. J., van Pijkeren, J.-P., Patel, R. B., Stolz, D., Watkins, S., Ejaz, A., & Greenberger, J. S. (2022). Intestinal Radiation Protection and Mitigation by Second-Generation Probiotic Lactobacillus-reuteri Engineered to Deliver Interleukin-22. International Journal of Molecular Sciences, 23(10), 5616. https://doi.org/10.3390/ijms23105616