
IgG from Adult Atopic Dermatitis (AD) Patients Induces Thymic IL-22 Production and CLA Expression on CD4+ T Cells: Possible Epigenetic Implications Mediated by miRNA

 , , ,
, , , 
Abstract
1. Introduction
2. Patients and Methods
2.1. Patient Samples
2.2. SPT
2.3. Serum Anti-Allergen IgE Determination
2.4. Thymic Tissue Dissociation, Cell Isolation, and Storage
2.5. IgG Purification, Isotypes Evaluation, and Labeling
2.6. Cell Culture and Flow Cytometry
2.7. RNA Extraction
2.8. sRNA Construction and Sequencing
2.9. Prediction, Gene Set Enrichment, and Functional Analysis of Target mRNA
3. Murine Methods
4. Statistical Analysis
5. Results
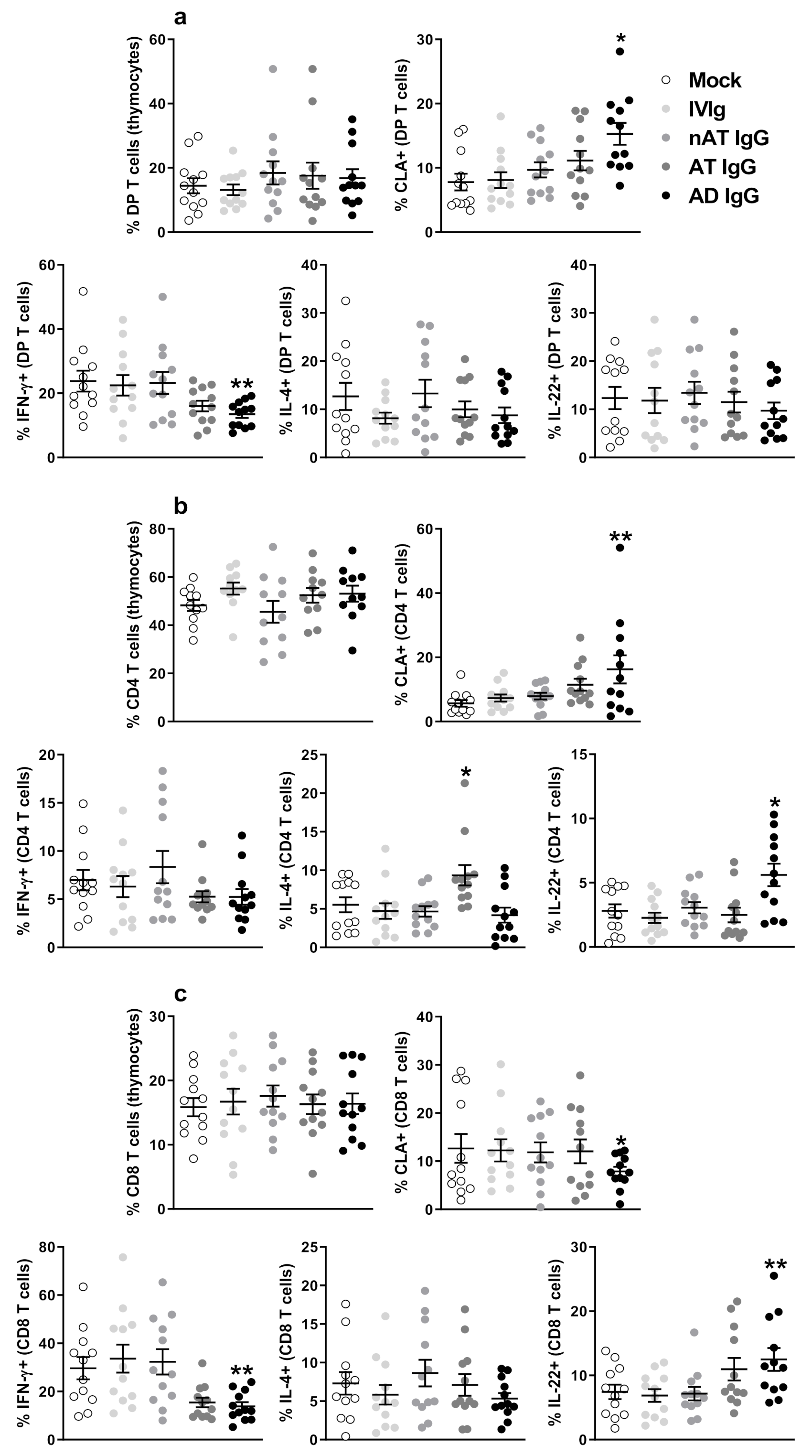
5.1. IgG from Adult AD Patients Induces CLA Expression and IL-22 Production by Infant Non-Atopic Intra-Thymic CD4 T Cells with Similar Implications on Murine Cells

5.2. IgG Isotypes Are Not Related to the Modulatory Effect and the Possible Involvement of Direct Interactions with Maturing T Cells
5.3. Identification of Differentially Expressed miRNAs and Their Target Genes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Hits | Pop Hits | List Total | Pop Total | Genes | p-Value | Adjusted p-Value (BH) |
|---|---|---|---|---|---|---|---|
| R-HSA-381340_Transcriptional regulation of white adipocyte differentiation | 7 | 133 | 69 | 10812 | CDK8;CDK19;TBL1XR1;PPARG | 0.0 | 0.0 |
| R-HSA-8986944_Transcriptional Regulation by MECP2 | 8 | 74 | 69 | 10812 | TNRC6B;TRPC3;AGO1;TBL1XR1;PPARG;LBR | 0.0 | 0.0 |
| R-HSA-73857_RNA Polymerase II Transcription | 27 | 1628 | 69 | 10812 | ESR1;TNRC6B;NABP1;RRM2;CBX5;KRBOX4;TRPC3;ZNF566;ZNF268;MDM4;CCNT2;CDK8;NR6A1;PHC3;AGO1;TBL1XR1;CPSF2;PPARG;LBR;RRAGD | 0.0001 | 0.0008 |
| R-HSA-74160_Gene expression (Transcription) | 30 | 1838 | 69 | 10812 | ESR1;TNRC6B;NABP1;RRM2;TET2;CBX5;KRBOX4;TRPC3;ZNF566;ZNF268;NFIB;MDM4;CCNT2;CDK8;NR6A1;PHC3;AGO1;TBL1XR1;CPSF2;PPARG;LBR;MBD2;RRAGD | 0.0001 | 0.0008 |
| R-HSA-212436_Generic Transcription Pathway | 23 | 1439 | 69 | 10812 | ESR1;TNRC6B;RRM2;CBX5;KRBOX4;TRPC3;ZNF566;ZNF268;MDM4;CCNT2;CDK8;NR6A1;PHC3;AGO1;TBL1XR1;PPARG;LBR;RRAGD | 0.0003 | 0.0015 |
| R-HSA-6807070_PTEN Regulation | 7 | 177 | 69 | 10812 | TNRC6B;PHC3;AGO1;PPARG;RRAGD | 0.0003 | 0.0015 |
| R-HSA-8943724_Regulation of PTEN gene transcription | 5 | 98 | 69 | 10812 | PHC3;PPARG;RRAGD | 0.0006 | 0.0026 |
| R-HSA-1257604_PIP3 activates AKT signaling | 8 | 307 | 69 | 10812 | ESR1;TNRC6B;PHC3;AGO1;PPARG;RRAGD | 0.0014 | 0.0052 |
| R-HSA-1989781_PPARA activates gene expression | 5 | 134 | 69 | 10812 | CDK8;CDK19;TBL1XR1;PPARG | 0.0024 | 0.0075 |
| R-HSA-400206_Regulation of lipid metabolism by PPARalpha | 5 | 136 | 69 | 10812 | CDK8;CDK19;TBL1XR1;PPARG | 0.0025 | 0.0075 |
| R-HSA-9006925_Intracellular signaling by second messengers | 8 | 360 | 69 | 10812 | ESR1;TNRC6B;PHC3;AGO1;PPARG;RRAGD | 0.0036 | 0.0098 |
| R-HSA-3108232_SUMO E3 ligases SUMOylate target proteins | 6 | 231 | 69 | 10812 | ESR1;CBX5;SMC1A;PHC3;PPARG | 0.0052 | 0.013 |
| R-HSA-2990846_SUMOylation | 6 | 239 | 69 | 10812 | ESR1;CBX5;SMC1A;PHC3;PPARG | 0.0061 | 0.0141 |
| R-HSA-2559583_Cellular Senescence | 5 | 208 | 69 | 10812 | TNRC6B;MDM4;HMGA2;PHC3;AGO1 | 0.0137 | 0.0294 |
| R-HSA-9006931_Signaling by Nuclear Receptors | 6 | 305 | 69 | 10812 | ESR1;TNRC6B;SMC1A;AGO1;TBL1XR1;SCD | 0.0178 | 0.0356 |
| R-HSA-556833_Metabolism of lipids | 11 | 786 | 69 | 10812 | OSBPL8;ACER2;CDK8;SLC44A1;CDK19;TBL1XR1;PPARG;LBR;PLPP3;SCD | 0.0195 | 0.0366 |

5.4. Functional and Pathway Enrichment Analyses
6. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kim, J.; Kim, B.E.; Leung, D.Y.M. Pathophysiology of atopic dermatitis: Clinical implications. Allergy Asthma Proc. 2019, 40, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Leung, D.Y.M. The microbiome and allergic diseases: A struggle between good and bad microbes. Ann. Allergy Asthma Immunol. 2019, 122, 231–232. [Google Scholar] [CrossRef] [PubMed]
- Goleva, E.; Berdyshev, E.; Leung, D.Y. Epithelial barrier repair and prevention of allergy. J. Clin. Investig. 2019, 129, 1463–1474. [Google Scholar] [CrossRef] [PubMed]
- Barbarot, S.; Auziere, S.; Gadkari, A.; Girolomoni, G.; Puig, L.; Simpson, E.L.; Margolis, D.J.; de Bruin-Weller, M.; Eckert, L. Epidemiology of atopic dermatitis in adults: Results from an international survey. Allergy 2018, 73, 1284–1293. [Google Scholar] [CrossRef] [PubMed]
- Czarnowicki, T.; Esaki, H.; Gonzalez, J.; Malajian, D.; Shemer, A.; Noda, S.; Talasila, S.; Berry, A.; Gray, J.; Becker, L.; et al. Early pediatric atopic dermatitis shows only a cutaneous lymphocyte antigen (CLA)(+) TH2/TH1 cell imbalance, whereas adults acquire CLA(+) TH22/TC22 cell subsets. J. Allergy Clin. Immunol. 2015, 136, 941–951.e943. [Google Scholar] [CrossRef]
- Gürkan, A.; Yücel, A.A.; Sönmez, C.; Keleş, Ş.; Bostancı, İ. Serum Cytokine Profiles in Infants with Atopic Dermatitis. Acta Dermatovenerol. Croat. 2016, 24, 268–273. [Google Scholar]
- Guttman-Yassky, E.; Brunner, P.M.; Neumann, A.U.; Khattri, S.; Pavel, A.B.; Malik, K.; Singer, G.K.; Baum, D.; Gilleaudeau, P.; Sullivan-Whalen, M.; et al. Efficacy and safety of fezakinumab (an IL-22 monoclonal antibody) in adults with moderate-to-severe atopic dermatitis inadequately controlled by conventional treatments: A randomized, double-blind, phase 2a trial. J. Am. Acad. Dermatol. 2018, 78, 872–881.e876. [Google Scholar] [CrossRef]
- de Sousa, T.R.; Victor, J.R. Natural Self-Ligand Gamma Delta T Cell Receptors (γδTCRs) Insight: The Potential of Induced IgG. Vaccines 2020, 8, 436. [Google Scholar] [CrossRef]
- Victor, J.R. Allergen-specific IgG as a mediator of allergy inhibition: Lessons from mother to child. Hum. Vaccines Immunother. 2017, 13, 507–513. [Google Scholar] [CrossRef]
- Victor, J.R. Influence of maternal immunization with allergens on the thymic maturation of lymphocytes with regulatory potential in children: A broad field for further exploration. J. Immunol. Res. 2014, 2014, 780386. [Google Scholar] [CrossRef]
- Santos, L.S.; Sgnotto, F.D.R.; Inoue, A.H.S.; Padreca, A.F.; Menghini, R.P.; Duarte, A.J.D.S.; Victor, J.R. IgG from Non-atopic Individuals Induces In Vitro IFN-γ and IL-10 Production by Human Intra-thymic γδT Cells: A Comparison with Atopic IgG and IVIg. Arch. Immunol. Ther. Exp. 2019, 67, 263–270. [Google Scholar] [CrossRef]
- Futata, E.; de Brito, C.; Victor, J.; Fusaro, A.; Oliveira, C.; Maciel, M.; Duarte, A.; Sato, M. Long-term anergy in orally tolerized mice is linked to decreased B7.2 expression on B cells. Immunobiology 2006, 211, 157–166. [Google Scholar] [CrossRef]
- da Ressureição Sgnotto, F.; Santos, L.S.; de Sousa, T.R.; de Lima, J.F.; da Silva Oliveira, L.M.; Sanabani, S.S.; da Silva Duarte, A.J.; Victor, J.R. IgG From HIV-1-Exposed Seronegative and HIV-1-Infected Subjects Differently Modulates IFN-γ Production by Thymic T and B Cells. J. Acquir. Immune Defic. Syndr. 2019, 82, e56–e60. [Google Scholar] [CrossRef]
- de Oliveira, M.G.; Lira, A.A.L.; Sgnotto, F.D.R.; Inoue, A.H.S.; Beltrame, G.R.; da Silva, D.; Menghini, R.P.; Duarte, A.J.D.S.; Victor, J.R. Maternal immunization downregulates offspring TCD4 regulatory cells (Tregs) thymic maturation without implications for allergy inhibition. Scand. J. Immunol. 2018, 88, e12721. [Google Scholar] [CrossRef]
- da Ressureicao Sgnotto, F.; de Oliveira, M.G.; de Lima Lira, A.A.; Bento-de-Souza, L.; da Silva Duarte, A.J.; Victor, J.R. Low doses of IgG from atopic individuals can modulate in vitro IFN-gamma production by human intra-thymic TCD4 and TCD8 cells: An IVIg comparative approach. Hum. Vaccines Immunother. 2017, 13, 1563–1572. [Google Scholar] [CrossRef]
- de Oliveira, M.G.; Oliveira, L.M.; Lira, A.A.L.; Sgnotto, F.D.R.; Duarte, A.J.D.S.; Sato, M.N.; Victor, J.R. Preconception allergen sensitization can induce B10 cells in offspring: A potential main role for maternal IgG. Allergy Asthma Clin. Immunol. 2017, 13, 22. [Google Scholar] [CrossRef]
- de Oliveira, M.G.; da Ressureição Sgnotto, F.; de Sousa, T.R.; Fagundes, B.O.; Duarte, A.J.D.S.; Victor, J.R. Preconceptional immunization with an allergen inhibits offspring thymic Th17 cells maturation without influence on Th1 and Th2 cells. Eur. Cytokine Netw. 2020, 31, 113–117. [Google Scholar] [CrossRef]
- Inoue, A.H.S.; Lira, A.A.L.; de-Oliveira, M.G.; de Sousa, T.R.; Sgnotto, F.D.R.; Duarte, A.J.D.S.; Victor, J.R. The Potential of IgG to Induce Murine and Human Thymic Maturation of IL-10+ B Cells (B10) Revealed in a Pilot Study. Cells 2020, 9, 2239. [Google Scholar] [CrossRef]
- de Sousa, T.R.; da Ressureição Sgnotto, F.; Fagundes, B.O.; Santos, L.S.; da Silva Duarte, A.J.; Victor, J.R. IgG from atopic individuals can mediate non-atopic infant thymic and adult peripheral CD8 + TC2 skewing without influence on TC17 or TC22 cells. Eur. Ann. Allergy Clin. Immunol. 2020, 53, 161. [Google Scholar] [CrossRef]
- Santos, L.S.; Sgnotto, F.D.R.; Sousa, T.R.; Orfali, R.L.; Aoki, V.; Duarte, A.J.D.S.; Victor, J.R. IgG from atopic dermatitis patients induces non-atopic infant thymic invariant natural killer T (iNKT) cells to produce IL-4, IL-17, and IL-10. Int. J. Dermatol. 2019, 59, 359–364. [Google Scholar] [CrossRef]
- Sgnotto, F.D.R.; de Oliveira, M.G.; Lira, A.A.L.; Inoue, A.H.S.; Titz, T.O.; Orfali, R.L.; Bento-de-Souza, L.; Sato, M.N.; Aoki, V.; Duarte, A.J.S.; et al. IgG from atopic dermatitis patients induces IL-17 and IL-10 production in infant intrathymic TCD4 and TCD8 cells. Int. J. Dermatol. 2018, 57, 434–440. [Google Scholar] [CrossRef]
- Orfali, R.L.; Sato, M.N.; Takaoka, R.; Azor, M.H.; Rivitti, E.A.; Hanifin, J.M.; Aoki, V. Atopic dermatitis in adults: Evaluation of peripheral blood mononuclear cells proliferation response to Staphylococcus aureus enterotoxins A and B and analysis of interleukin-18 secretion. Exp. Dermatol. 2009, 18, 628–633. [Google Scholar] [CrossRef]
- Czarnowicki, T.; Gonzalez, J.; Shemer, A.; Malajian, D.; Xu, H.; Zheng, X.; Khattri, S.; Gilleaudeau, P.; Sullivan-Whalen, M.; Suárez-Fariñas, M.; et al. Severe atopic dermatitis is characterized by selective expansion of circulating TH2/TC2 and TH22/TC22, but not TH17/TC17, cells within the skin-homing T-cell population. J. Allergy Clin. Immunol. 2015, 136, 104–115.e107. [Google Scholar] [CrossRef]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Baltimore, D.; Boldin, M.P.; O’Connell, R.M.; Rao, D.S.; Taganov, K.D. MicroRNAs: New regulators of immune cell development and function. Nat. Immunol. 2008, 9, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Wells, A.C.; Daniels, K.A.; Angelou, C.C.; Fagerberg, E.; Burnside, A.S.; Markstein, M.; Alfandari, D.; Welsh, R.M.; Pobezinskaya, E.L.; Pobezinsky, L.A. Modulation of let-7 miRNAs controls the differentiation of effector CD8 T cells. Elife 2017, 6, e26398. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Z.; Li, L.; Lodish, H.F.; Bartel, D.P. MicroRNAs modulate hematopoietic lineage differentiation. Science 2004, 303, 83–86. [Google Scholar] [CrossRef]
- Hanifin, J.M.; Thurston, M.; Omoto, M.; Cherill, R.; Tofte, S.J.; Graeber, M. The eczema area and severity index (EASI): Assessment of reliability in atopic dermatitis. EASI Evaluator Group. Exp. Dermatol. 2001, 10, 11–18. [Google Scholar] [CrossRef]
- Heinzerling, L.; Mari, A.; Bergmann, K.C.; Bresciani, M.; Burbach, G.; Darsow, U.; Durham, S.; Fokkens, W.; Gjomarkaj, M.; Haahtela, T.; et al. The skin prick test-European standards. Clin. Transl. Allergy 2013, 3, 3. [Google Scholar] [CrossRef]
- Valadao de Souza, D.R.; Pessoa, R.; Nascimento, A.; Nukui, Y.; Pereira, J.; Casseb, J.; Penalva de Oliveira, A.C.; da Silva Duarte, A.J.; Clissa, P.B.; Sanabani, S.S. Small RNA profiles of HTLV-1 asymptomatic carriers with monoclonal and polyclonal rearrangement of the T-cell antigen receptor gamma-chain using massively parallel sequencing: A pilot study. Oncol. Lett. 2020, 20, 2311–2321. [Google Scholar] [CrossRef]
- Bento-de-Souza, L.; Victor, J.R.; Bento-de-Souza, L.C.; Arrais-Santos, M.; Rangel-Santos, A.C.; Pereira-Costa, É.; Raniero-Fernandes, E.; Seixas-Duarte, M.I.; Oliveira-Filho, J.B.; Duarte, A.J.S. Constitutive expression of genes encoding notch receptors and ligands in developing lymphocytes, nTreg cells and dendritic cells in the human thymus. Results Immunol. 2016, 6, 15–20. [Google Scholar] [CrossRef]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. miRWalk: An online resource for prediction of microRNA binding sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef]
- Sheth, U.; Parker, R. Decapping and decay of messenger RNA occur in cytoplasmic processing bodies. Science 2003, 300, 805–808. [Google Scholar] [CrossRef]
- Lemieux, R.; Bazin, R.; Néron, S. Therapeutic intravenous immunoglobulins. Mol. Immunol. 2005, 42, 839–848. [Google Scholar] [CrossRef]
- Nascimento, A.; Valadao de Souza, D.R.; Pessoa, R.; Pietrobon, A.J.; Nukui, Y.; Pereira, J.; Casseb, J.; de Oliveira, A.C.P.; Loureiro, P.; da Silva Duarte, A.J.; et al. Global expression of noncoding RNome reveals dysregulation of small RNAs in patients with HTLV-1-associated adult T-cell leukemia: A pilot study. Infect. Agents Cancer 2021, 16, 4. [Google Scholar] [CrossRef]
- Esaki, H.; Brunner, P.M.; Renert-Yuval, Y.; Czarnowicki, T.; Huynh, T.; Tran, G.; Lyon, S.; Rodriguez, G.; Immaneni, S.; Johnson, D.B.; et al. Early-onset pediatric atopic dermatitis is TH2 but also TH17 polarized in skin. J. Allergy Clin. Immunol. 2016, 138, 1639–1651. [Google Scholar] [CrossRef]
- Czarnowicki, T.; Santamaria-Babí, L.F.; Guttman-Yassky, E. Circulating CLA. Allergy 2017, 72, 366–372. [Google Scholar] [CrossRef]
- Nomura, T.; Wu, J.; Kabashima, K.; Guttman-Yassky, E. Endophenotypic Variations of Atopic Dermatitis by Age, Race, and Ethnicity. J. Allergy Clin. Immunol. Pract. 2020, 8, 1840–1852. [Google Scholar] [CrossRef]
- Nahm, D.H.; Ye, Y.M.; Shin, Y.S.; Park, H.S.; Kim, M.E.; Kwon, B.; Cho, S.M.; Han, J. Efficacy, Safety, and Immunomodulatory Effect of the Intramuscular Administration of Autologous Total Immunoglobulin G for Atopic Dermatitis: A Randomized Clinical Trial. Allergy Asthma Immunol. Res. 2020, 12, 949–963. [Google Scholar] [CrossRef]
- Cho, S.M.; Kim, M.E.; Kwon, B.; Nahm, D.H. Immunomodulatory effects induced by intramuscular administration of autologous total immunoglobulin G in patients with atopic dermatitis. Int. Immunopharmacol. 2017, 52, 1–6. [Google Scholar] [CrossRef]
- Kim, S.M.; Park, C.O. Alternative Immunomodulatory and Disease-Modifying Treatment for Atopic Dermatitis: Autologous Total Immunoglobulin G. Allergy Asthma Immunol. Res. 2020, 12, 903–905. [Google Scholar] [CrossRef] [PubMed]
- Aalberse, R.C.; Van Milligen, F.; Tan, K.Y.; Stapel, S.O. Allergen-specific IgG4 in atopic disease. Allergy 1993, 48, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Aalberse, R.C.; Stapel, S.O.; Schuurman, J.; Rispens, T. Immunoglobulin G4: An odd antibody. Clin. Exp. Allergy 2009, 39, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Smoldovskaya, O.; Feyzkhanova, G.; Voloshin, S.; Arefieva, A.; Chubarova, A.; Pavlushkina, L.; Filatova, T.; Antonova, E.; Timofeeva, E.; Butvilovskaya, V.; et al. Allergen-specific IgE and IgG4 patterns among patients with different allergic diseases. World Allergy Organ. J. 2018, 11, 35. [Google Scholar] [CrossRef]
- de Oliveira, M.G.; de Lima Lira, A.A.; da Ressureição Sgnotto, F.; Inoue, A.H.S.; Santos, L.S.; Nakamatsu, B.Y.; Duarte, A.J.D.S.; Leite-de-Moraes, M.; Victor, J.R. Maternal IgG impairs the maturation of offspring intrathymic IL-17-producing γδT cells: Implications for murine and human allergies. Clin. Exp. Allergy 2019, 49, 1000–1012. [Google Scholar] [CrossRef]
- Eichmann, K.; Rajewsky, K. Induction of T and B cell immunity by anti-idiotypic antibody. Eur. J. Immunol. 1975, 5, 661–666. [Google Scholar] [CrossRef]
- Schluter, S.F.; Adelman, M.K.; Taneja, V.; David, C.; Yocum, D.E.; Marchalonis, J.J. Natural autoantibodies to TCR public idiotopes: Potential roles in immunomodulation. Cell Mol. Biol. 2003, 49, 193–207. [Google Scholar]
- Adelman, M.K.; Schluter, S.F.; Robey, I.F.; Marchalonis, J.J. Natural and autoantibodies to human T-cell receptor Vbeta segments: Potential roles in immunomodulation. Crit. Rev. Immunol. 2007, 27, 221–232. [Google Scholar] [CrossRef]
- Ballow, M. Mechanisms of immune regulation by IVIG. Curr. Opin. Allergy Clin. Immunol. 2014, 14, 509–515. [Google Scholar] [CrossRef]
- Victor, J.R. Do different IgG repertoires play a role in B- and T-cell functional modulation during ontogeny? The “hooks without bait” theory. Immunol. Cell Biol. 2020, 98, 540–548. [Google Scholar] [CrossRef]
- Indrieri, A.; Carrella, S.; Carotenuto, P.; Banfi, S.; Franco, B. The Pervasive Role of the miR-181 Family in Development, Neurodegeneration, and Cancer. Int. J. Mol. Sci. 2020, 21, 2092. [Google Scholar] [CrossRef]
- Li, L.; Feng, J.; Zhang, D.; Yong, J.; Wang, Y.; Yao, J.; Huang, R. Differential expression of miR-4492 and IL-10 is involved in chronic rhinosinusitis with nasal polyps. Exp. Ther. Med. 2019, 18, 3968–3976. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, Y.; Huang, J.; Gao, L.; Wu, Z.; Wang, L.; Fan, L. Let-7b-5p promotes cell apoptosis in Parkinson’s disease by targeting HMGA2. Mol. Med. Rep. 2021, 24, 820. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, L.; Tang, S. MicroRNA-4497 functions as a tumor suppressor in laryngeal squamous cell carcinoma via negatively modulation the GBX2. Auris Nasus Larynx 2019, 46, 106–113. [Google Scholar] [CrossRef]
- Chen, Z.; Ying, J.; Shang, W.; Ding, D.; Guo, M.; Wang, H. miR-342-3p Regulates the Proliferation and Apoptosis of NSCLC Cells by Targeting. Technol. Cancer Res. Treat. 2021, 20, 15330338211041193. [Google Scholar] [CrossRef]
- Huang, Z.; Xing, S.; Liu, M.; Deng, W.; Wang, Y.; Huang, Y.; Huang, X.; Wu, C.; Guo, X.; Pan, X.; et al. MiR-26a-5p enhances cells proliferation, invasion, and apoptosis resistance of fibroblast-like synoviocytes in rheumatoid arthritis by regulating PTEN/PI3K/AKT pathway. Biosci. Rep. 2019, 39, BSR20182192. [Google Scholar] [CrossRef]
- Ghorbani, S.; Talebi, F.; Ghasemi, S.; Abad, A.J.J.; Vojgani, M.; Noorbakhsh, F. miR-181 interacts with signaling adaptor molecule DENN/MADD and enhances TNF-induced cell death. PLoS ONE 2017, 12, e0174368. [Google Scholar] [CrossRef]
- Huo, X.; Zhang, K.; Yi, L.; Mo, Y.; Liang, Y.; Zhao, J.; Zhang, Z.; Xu, Y.; Zhen, G. Decreased epithelial and plasma miR-181b-5p expression associates with airway eosinophilic inflammation in asthma. Clin. Exp. Allergy 2016, 46, 1281–1290. [Google Scholar] [CrossRef]
- Acevedo, N.; Benfeitas, R.; Katayama, S.; Bruhn, S.; Andersson, A.; Wikberg, G.; Lundeberg, L.; Lindvall, J.M.; Greco, D.; Kere, J.; et al. Epigenetic alterations in skin homing CD4. Sci. Rep. 2020, 10, 18020. [Google Scholar] [CrossRef]
- Li, H.M.; Xiao, Y.J.; Min, Z.S.; Tan, C. Identification and interaction analysis of key genes and microRNAs in atopic dermatitis by bioinformatics analysis. Clin. Exp. Dermatol. 2019, 44, 257–264. [Google Scholar] [CrossRef]
- Gu, C.; Li, Y.; Wu, J.; Xu, J. IFN-γ-induced microRNA-29b up-regulation contributes tokeratinocyte apoptosis in atopic dermatitis through inhibiting Bcl2L2. Int. J. Clin. Exp. Pathol. 2017, 10, 10117–10126. [Google Scholar]
- Sonkoly, E.; Janson, P.; Majuri, M.L.; Savinko, T.; Fyhrquist, N.; Eidsmo, L.; Xu, N.; Meisgen, F.; Wei, T.; Bradley, M.; et al. MiR-155 is overexpressed in patients with atopic dermatitis and modulates T-cell proliferative responses by targeting cytotoxic T lymphocyte-associated antigen 4. J. Allergy Clin. Immunol. 2010, 126, 581–589.e20. [Google Scholar] [CrossRef]
- Malaisse, J.; Bourguignon, V.; De Vuyst, E.; de Rouvroit, C.L.; Nikkels, A.F.; Flamion, B.; Poumay, Y. Hyaluronan metabolism in human keratinocytes and atopic dermatitis skin is driven by a balance of hyaluronan synthases 1 and 3. J. Investig. Dermatol. 2014, 134, 2174–2182. [Google Scholar] [CrossRef]
- Kärner, J.; Wawrzyniak, M.; Tankov, S.; Runnel, T.; Aints, A.; Kisand, K.; Altraja, A.; Kingo, K.; Akdis, C.A.; Akdis, M.; et al. Increased microRNA-323-3p in IL-22/IL-17-producing T cells and asthma: A role in the regulation of the TGF-β pathway and IL-22 production. Allergy 2017, 72, 55–65. [Google Scholar] [CrossRef]
- Yang, L.; Hu, Z.; Jin, Y.; Huang, N.; Xu, S. MiR-4497 mediates oxidative stress and inflammatory injury in keratinocytes induced by ultraviolet B radiation through regulating NF-κB expression. Ital. J. Dermatol. Venerol. 2022, 157, 84–91. [Google Scholar] [CrossRef]
- Rebane, A.; Runnel, T.; Aab, A.; Maslovskaja, J.; Rückert, B.; Zimmermann, M.; Plaas, M.; Kärner, J.; Treis, A.; Pihlap, M.; et al. MicroRNA-146a alleviates chronic skin inflammation in atopic dermatitis through suppression of innate immune responses in keratinocytes. J. Allergy Clin. Immunol. 2014, 134, 836–847.e811. [Google Scholar] [CrossRef]
- Sonkoly, E.; Wei, T.; Janson, P.C.; Sääf, A.; Lundeberg, L.; Tengvall-Linder, M.; Norstedt, G.; Alenius, H.; Homey, B.; Scheynius, A.; et al. MicroRNAs: Novel regulators involved in the pathogenesis of psoriasis? PLoS ONE 2007, 2, e610. [Google Scholar] [CrossRef]
- Zheng, R.; Du, M.; Tian, M.; Zhu, Z.; Wei, C.; Chu, H.; Gan, C.; Liang, J.; Xue, R.; Gao, F.; et al. Fine Particulate Matter Induces Childhood Asthma Attacks via Extracellular Vesicle-Packaged Let-7i-5p-Mediated Modulation of the MAPK Signaling Pathway. Adv. Sci. 2022, 9, e2102460. [Google Scholar] [CrossRef] [PubMed]
- Soares, C.T.; Trombone, A.P.F.; Fachin, L.R.V.; Rosa, P.S.; Ghidella, C.C.; Ramalho, R.F.; Pinilla, M.G.; Carvalho, A.F.; Carrara, D.N.; Soares, F.A.; et al. Differential Expression of MicroRNAs in Leprosy Skin Lesions. Front. Immunol. 2017, 8, 1035. [Google Scholar] [CrossRef] [PubMed]
- Chiba, T.; Kooka, A.; Kowatari, K.; Yoshizawa, M.; Chiba, N.; Takaguri, A.; Fukushi, Y.; Hongo, F.; Sato, H.; Wada, S. Expression profiles of hsa-miR-148a-3p and hsa-miR-125b-5p in human breast milk and infant formulae. Int. Breastfeed J. 2022, 17, 1. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Chu, C.; Wang, M. Correlation of microRNA profiles with disease risk and severity of allergic rhinitis. Int. J. Clin. Exp. Pathol. 2018, 11, 1791–1802. [Google Scholar]
- Kim, B.S.; Jung, J.Y.; Jeon, J.Y.; Kim, H.A.; Suh, C.H. Circulating hsa-miR-30e-5p, hsa-miR-92a-3p, and hsa-miR-223-3p may be novel biomarkers in systemic lupus erythematosus. HLA 2016, 88, 187–193. [Google Scholar] [CrossRef]
- Zhang, L.J.; Guerrero-Juarez, C.F.; Hata, T.; Bapat, S.P.; Ramos, R.; Plikus, M.V.; Gallo, R.L. Innate immunity. Dermal adipocytes protect against invasive Staphylococcus aureus skin infection. Science 2015, 347, 67–71. [Google Scholar] [CrossRef]
- Ong, P.Y.; Ohtake, T.; Brandt, C.; Strickland, I.; Boguniewicz, M.; Ganz, T.; Gallo, R.L.; Leung, D.Y. Endogenous antimicrobial peptides and skin infections in atopic dermatitis. N. Engl. J. Med. 2002, 347, 1151–1160. [Google Scholar] [CrossRef]
- Li, H.; Yamagata, T.; Mori, M.; Yasuhara, A.; Momoi, M.Y. Mutation analysis of methyl-CpG binding protein family genes in autistic patients. Brain Dev. 2005, 27, 321–325. [Google Scholar] [CrossRef]
- Koelsch, K.A.; Webb, R.; Jeffries, M.; Dozmorov, M.G.; Frank, M.B.; Guthridge, J.M.; James, J.A.; Wren, J.D.; Sawalha, A.H. Functional characterization of the MECP2/IRAK1 lupus risk haplotype in human T cells and a human MECP2 transgenic mouse. J. Autoimmun. 2013, 41, 168–174. [Google Scholar] [CrossRef][Green Version]
- Ausió, J.; Martínez de Paz, A.; Esteller, M. MeCP2: The long trip from a chromatin protein to neurological disorders. Trends. Mol. Med. 2014, 20, 487–498. [Google Scholar] [CrossRef]
- Wu, K.; Volke, A.; Lund, M.; Bang, K.; Thestrup-Pedersen, K. Telomerase activity is spontaneously increased in lymphocytes from patients with atopic dermatitis and correlates with cellular proliferation. J. Dermatol. Sci. 1999, 22, 24–30. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Sousa, T.R.; Fagundes, B.O.; Nascimento, A.; Fernandes, L.A.; Sgnotto, F.d.R.; Orfali, R.L.; Aoki, V.; Duarte, A.J.d.S.; Sanabani, S.S.; Victor, J.R. IgG from Adult Atopic Dermatitis (AD) Patients Induces Thymic IL-22 Production and CLA Expression on CD4+ T Cells: Possible Epigenetic Implications Mediated by miRNA. Int. J. Mol. Sci. 2022, 23, 6867. https://doi.org/10.3390/ijms23126867
de Sousa TR, Fagundes BO, Nascimento A, Fernandes LA, Sgnotto FdR, Orfali RL, Aoki V, Duarte AJdS, Sanabani SS, Victor JR. IgG from Adult Atopic Dermatitis (AD) Patients Induces Thymic IL-22 Production and CLA Expression on CD4+ T Cells: Possible Epigenetic Implications Mediated by miRNA. International Journal of Molecular Sciences. 2022; 23(12):6867. https://doi.org/10.3390/ijms23126867
Chicago/Turabian Stylede Sousa, Thamires Rodrigues, Beatriz Oliveira Fagundes, Andrezza Nascimento, Lorena Abreu Fernandes, Fábio da Ressureição Sgnotto, Raquel Leão Orfali, Valéria Aoki, Alberto José da Silva Duarte, Sabri Saeed Sanabani, and Jefferson Russo Victor. 2022. "IgG from Adult Atopic Dermatitis (AD) Patients Induces Thymic IL-22 Production and CLA Expression on CD4+ T Cells: Possible Epigenetic Implications Mediated by miRNA" International Journal of Molecular Sciences 23, no. 12: 6867. https://doi.org/10.3390/ijms23126867
APA Stylede Sousa, T. R., Fagundes, B. O., Nascimento, A., Fernandes, L. A., Sgnotto, F. d. R., Orfali, R. L., Aoki, V., Duarte, A. J. d. S., Sanabani, S. S., & Victor, J. R. (2022). IgG from Adult Atopic Dermatitis (AD) Patients Induces Thymic IL-22 Production and CLA Expression on CD4+ T Cells: Possible Epigenetic Implications Mediated by miRNA. International Journal of Molecular Sciences, 23(12), 6867. https://doi.org/10.3390/ijms23126867