
Meta-Analysis of Two Human RNA-seq Datasets to Determine Periodontitis Diagnostic Biomarkers and Drug Target Candidates

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
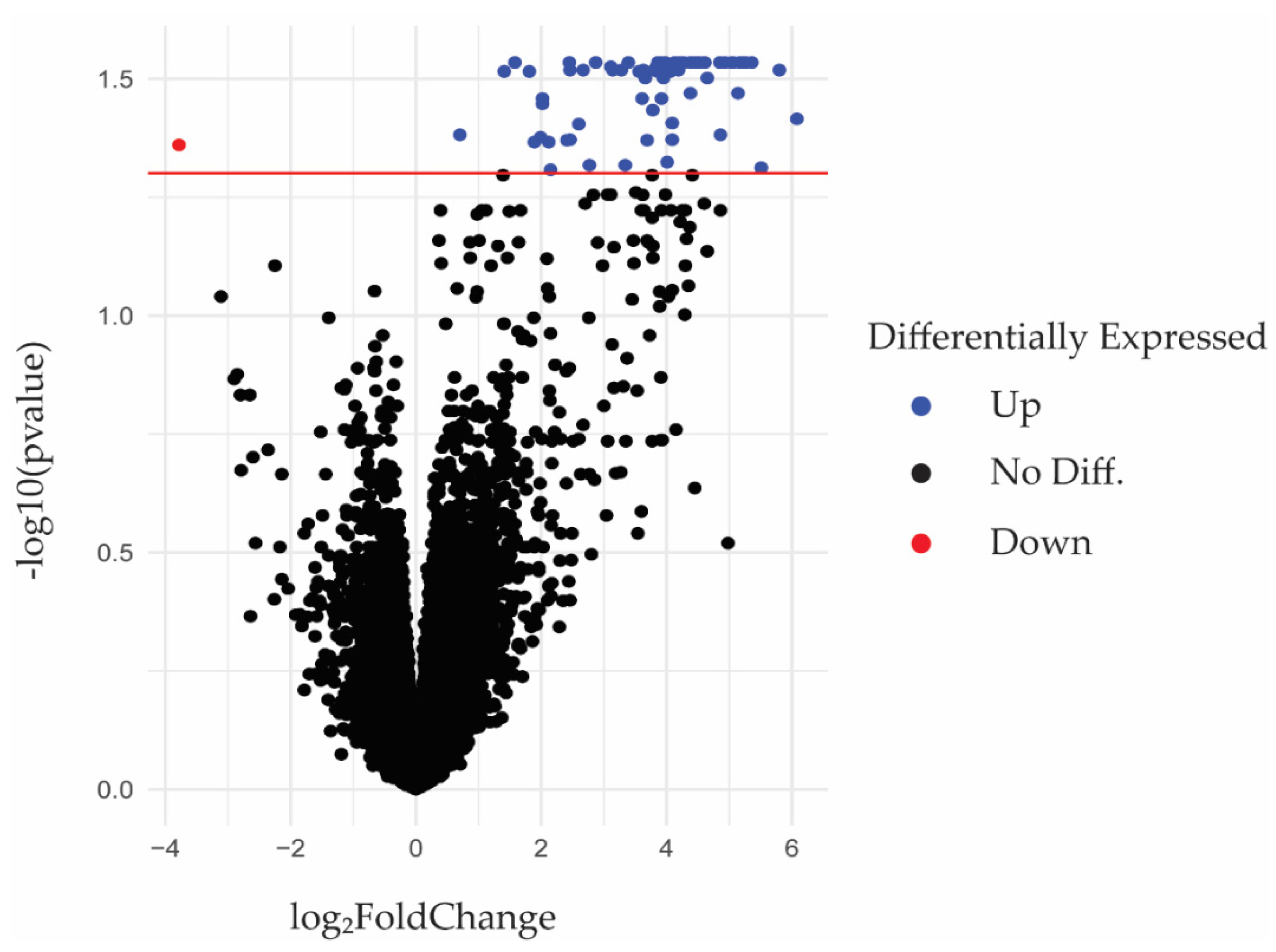
2.1. RNA-seq Identification of Differentially Expressed Genes in Periodontal Gingival Epithelial Cells
2.2. Signaling Pathway Impact Analysis Identified 10 Significantly Impacted Pathways
2.3. Drug Target Analysis Identified 500 Drugs That May Be Repurposed to Treat Periodontitis
2.4. Protein-Protein Interaction Identification of Candidate Drug Targets against Top 10 Hub Proteins
3. Discussion
4. Materials and Methods
4.1. RNA-seq Analysis
4.2. Signaling Pathway Analysis
4.3. Drug Target Analysis
4.4. Protein-Protein Interactions Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eke, P.I.; Thornton-Evans, G.O.; Wei, L.; Borgnakke, W.S.; Dye, B.A.; Genco, R.J. Periodontitis in US Adults: National Health and Nutrition Examination Survey 2009–2014. J. Am. Dent. Assoc. 2018, 149, 576–588.e6. [Google Scholar] [CrossRef]
- Vos, T.; Abajobir, A.A.; Abate, K.H.; Abbafati, C.; Abbas, K.M.; Abd-Allah, F.; Abdulkader, R.S.; Abdulle, A.M.; Abebo, T.A.; Abera, S.F.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1211–1259. [Google Scholar] [CrossRef] [Green Version]
- Botelho, J.; Machado, V.; Leira, Y.; Proenca, L.; Chambrone, L.; Mendes, J.J. Economic burden of periodontitis in the United States and Europe: An updated estimation. J. Periodontol. 2022, 93, 373–379. [Google Scholar] [CrossRef]
- Darveau, R.P. Periodontitis: A polymicrobial disruption of host homeostasis. Nat. Rev. Microbiol. 2010, 8, 481–490. [Google Scholar] [CrossRef]
- Holt, S.C.; Ebersole, J.L. Porphyromonas gingivalis, Treponema denticola, and Tannerella forsythia: The “red complex”, a prototype polybacterial pathogenic consortium in periodontitis. Periodontol. 2000 2005, 38, 72–122. [Google Scholar] [CrossRef]
- de Andrade, K.Q.; Almeida-da-Silva, C.L.C.; Coutinho-Silva, R. Immunological Pathways Triggered by Porphyromonas gingivalis and Fusobacterium nucleatum: Therapeutic Possibilities? Mediat. Inflamm. 2019, 2019, 7241312. [Google Scholar] [CrossRef] [Green Version]
- Hajishengallis, G.; Darveau, R.P.; Curtis, M.A. The keystone-pathogen hypothesis. Nat. Rev. Microbiol. 2012, 10, 717–725. [Google Scholar] [CrossRef]
- Yucel-Lindberg, T.; Bage, T. Inflammatory mediators in the pathogenesis of periodontitis. Expert Rev. Mol. Med. 2013, 15, e7. [Google Scholar] [CrossRef] [Green Version]
- Peres, M.A.; Macpherson, L.M.D.; Weyant, R.J.; Daly, B.; Venturelli, R.; Mathur, M.R.; Listl, S.; Celeste, R.K.; Guarnizo-Herreno, C.C.; Kearns, C.; et al. Oral diseases: A global public health challenge. Lancet 2019, 394, 249–260. [Google Scholar] [CrossRef]
- Nazir, M.; Al-Ansari, A.; Al-Khalifa, K.; Alhareky, M.; Gaffar, B.; Almas, K. Global Prevalence of Periodontal Disease and Lack of Its Surveillance. Sci. World J. 2020, 2020, 2146160. [Google Scholar] [CrossRef]
- Teughels, W.; Dhondt, R.; Dekeyser, C.; Quirynen, M. Treatment of aggressive periodontitis. Periodontol. 2000 2014, 65, 107–133. [Google Scholar] [CrossRef]
- Prakasam, A.; Elavarasu, S.S.; Natarajan, R.K. Antibiotics in the management of aggressive periodontitis. J. Pharm. Bioallied Sci. 2012, 4, S252–S255. [Google Scholar] [CrossRef]
- Graves, D. Cytokines that promote periodontal tissue destruction. J. Periodontol. 2008, 79, 1585–1591. [Google Scholar] [CrossRef] [Green Version]
- Bartold, P.M.; Cantley, M.D.; Haynes, D.R. Mechanisms and control of pathologic bone loss in periodontitis. Periodontol. 2000 2010, 53, 55–69. [Google Scholar] [CrossRef]
- Tanabe, N.; Maeno, M.; Suzuki, N.; Fujisaki, K.; Tanaka, H.; Ogiso, B.; Ito, K. IL-1 alpha stimulates the formation of osteoclast-like cells by increasing M-CSF and PGE2 production and decreasing OPG production by osteoblasts. Life Sci. 2005, 77, 615–626. [Google Scholar] [CrossRef]
- Reis, C.; AV, D.A.C.; Guimaraes, J.T.; Tuna, D.; Braga, A.C.; Pacheco, J.J.; Arosa, F.A.; Salazar, F.; Cardoso, E.M. Clinical improvement following therapy for periodontitis: Association with a decrease in IL-1 and IL-6. Exp. Med. 2014, 8, 323–327. [Google Scholar] [CrossRef] [Green Version]
- Machado, V.; Botelho, J.; Escalda, C.; Hussain, S.B.; Luthra, S.; Mascarenhas, P.; Orlandi, M.; Mendes, J.J.; D’Aiuto, F. Serum C-Reactive Protein and Periodontitis: A Systematic Review and Meta-Analysis. Front. Immunol. 2021, 12, 706432. [Google Scholar] [CrossRef]
- Schenkein, H.A.; Papapanou, P.N.; Genco, R.; Sanz, M. Mechanisms underlying the association between periodontitis and atherosclerotic disease. Periodontol. 2000 2020, 83, 90–106. [Google Scholar] [CrossRef]
- Loos, B.G.; Craandijk, J.; Hoek, F.J.; Wertheim-van Dillen, P.M.; van der Velden, U. Elevation of systemic markers related to cardiovascular diseases in the peripheral blood of periodontitis patients. J. Periodontol. 2000, 71, 1528–1534. [Google Scholar] [CrossRef]
- Pfeilschifter, J.; Chenu, C.; Bird, A.; Mundy, G.R.; Roodman, G.D. Interleukin-1 and tumor necrosis factor stimulate the formation of human osteoclastlike cells in vitro. J. Bone Min. Res. 1989, 4, 113–118. [Google Scholar] [CrossRef]
- Baker, P.J.; Dixon, M.; Evans, R.T.; Dufour, L.; Johnson, E.; Roopenian, D.C. CD4(+) T cells and the proinflammatory cytokines gamma interferon and interleukin-6 contribute to alveolar bone loss in mice. Infect. Immun. 1999, 67, 2804–2809. [Google Scholar] [CrossRef] [Green Version]
- Teng, Y.T.; Nguyen, H.; Gao, X.; Kong, Y.Y.; Gorczynski, R.M.; Singh, B.; Ellen, R.P.; Penninger, J.M. Functional human T-cell immunity and osteoprotegerin ligand control alveolar bone destruction in periodontal infection. J. Clin. Investig. 2000, 106, R59–R67. [Google Scholar] [CrossRef] [Green Version]
- Dutzan, N.; Kajikawa, T.; Abusleme, L.; Greenwell-Wild, T.; Zuazo, C.E.; Ikeuchi, T.; Brenchley, L.; Abe, T.; Hurabielle, C.; Martin, D.; et al. A dysbiotic microbiome triggers TH17 cells to mediate oral mucosal immunopathology in mice and humans. Sci. Transl. Med. 2018, 10, eaat0797. [Google Scholar] [CrossRef] [Green Version]
- Dutzan, N.; Abusleme, L.; Konkel, J.E.; Moutsopoulos, N.M. Isolation, Characterization and Functional Examination of the Gingival Immune Cell Network. J. Vis. Exp. 2016, 108, 53736. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Aitken, S.; Sodek, J.; McCulloch, C.A. Evidence of a direct relationship between neutrophil collagenase activity and periodontal tissue destruction in vivo: Role of active enzyme in human periodontitis. J. Periodontal Res. 1995, 30, 23–33. [Google Scholar] [CrossRef]
- Landzberg, M.; Doering, H.; Aboodi, G.M.; Tenenbaum, H.C.; Glogauer, M. Quantifying oral inflammatory load: Oral neutrophil counts in periodontal health and disease. J. Periodontal Res. 2015, 50, 330–336. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Chavakis, T. Local and systemic mechanisms linking periodontal disease and inflammatory comorbidities. Nat. Rev. Immunol. 2021, 21, 426–440. [Google Scholar] [CrossRef]
- Pihlstrom, B.L.; Michalowicz, B.S.; Johnson, N.W. Periodontal diseases. Lancet 2005, 366, 1809–1820. [Google Scholar] [CrossRef] [Green Version]
- Bui, F.Q.; Almeida-da-Silva, C.L.C.; Huynh, B.; Trinh, A.; Liu, J.; Woodward, J.; Asadi, H.; Ojcius, D.M. Association between periodontal pathogens and systemic disease. Biomed. J. 2019, 42, 27–35. [Google Scholar] [CrossRef]
- Atarashi, K.; Suda, W.; Luo, C.; Kawaguchi, T.; Motoo, I.; Narushima, S.; Kiguchi, Y.; Yasuma, K.; Watanabe, E.; Tanoue, T.; et al. Ectopic colonization of oral bacteria in the intestine drives TH1 cell induction and inflammation. Science 2017, 358, 359–365. [Google Scholar] [CrossRef]
- Kitamoto, S.; Nagao-Kitamoto, H.; Jiao, Y.; Gillilland, M.G., 3rd; Hayashi, A.; Imai, J.; Sugihara, K.; Miyoshi, M.; Brazil, J.C.; Kuffa, P.; et al. The Intermucosal Connection between the Mouth and Gut in Commensal Pathobiont-Driven Colitis. Cell 2020, 182, 447–462.e414. [Google Scholar] [CrossRef] [PubMed]
- Arimatsu, K.; Yamada, H.; Miyazawa, H.; Minagawa, T.; Nakajima, M.; Ryder, M.I.; Gotoh, K.; Motooka, D.; Nakamura, S.; Iida, T.; et al. Oral pathobiont induces systemic inflammation and metabolic changes associated with alteration of gut microbiota. Sci. Rep. 2014, 4, 4828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, S.C.; Shao, Y.P.; Zhao, X.J.; Hong, C.S.; Wang, F.B.; Lu, X.; Li, J.; Ye, G.Z.; Yan, M.; Zhuang, Z.P.; et al. Integration of Metabolomics and Transcriptomics Reveals Major Metabolic Pathways and Potential Biomarker Involved in Prostate Cancer. Mol. Cell Proteom. 2016, 15, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Zeng, J.; Yang, Y.; Wu, B. Diagnostic biomarker candidates for pulpitis revealed by bioinformatics analysis of merged microarray gene expression datasets. BMC Oral Health 2020, 20, 279. [Google Scholar] [CrossRef]
- Flint, S.M.; McKinney, E.F.; Lyons, P.A.; Smith, K.G.C. The Contribution of Transcriptomics to Biomarker Development in Systemic Vasculitis and SLE. Curr. Pharm. Des. 2015, 21, 2225–2235. [Google Scholar] [CrossRef] [Green Version]
- Altmae, S.; Koel, M.; Vosa, U.; Adler, P.; Suhorutsenko, M.; Laisk-Podar, T.; Kukushkina, V.; Saare, M.; Velthut-Meikas, A.; Krjutskov, K.; et al. Meta-signature of human endometrial receptivity: A meta-analysis and validation study of transcriptomic biomarkers. Sci. Rep. 2017, 7, 10077. [Google Scholar] [CrossRef]
- Ma, T.; Liang, F.; Oesterreich, S.; Tseng, G.C. A Joint Bayesian Model for Integrating Microarray and RNA Sequencing Transcriptomic Data. J. Comput. Biol. 2017, 24, 647–662. [Google Scholar] [CrossRef]
- Fagard, R.H.; Staessen, J.A.; Thijs, L. Advantages and disadvantages of the meta-analysis approach. J. Hypertens. Suppl. 1996, 14, S9–S12; discussion S13. [Google Scholar] [CrossRef]
- Orjuela, S.; Huang, R.; Hembach, K.M.; Robinson, M.D.; Soneson, C. ARMOR: An Automated Reproducible MOdular Workflow for Preprocessing and Differential Analysis of RNA-seq Data. G3 (Bethesda) 2019, 9, 2089–2096. [Google Scholar] [CrossRef] [Green Version]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarca, A.L.; Draghici, S.; Khatri, P.; Hassan, S.S.; Mittal, P.; Kim, J.S.; Kim, C.J.; Kusanovic, J.P.; Romero, R. A novel signaling pathway impact analysis. Bioinformatics 2009, 25, 75–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Otasek, D.; Morris, J.H.; Boucas, J.; Pico, A.R.; Demchak, B. Cytoscape Automation: Empowering workflow-based network analysis. Genome Biol. 2019, 20, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, C.H.; Chen, S.H.; Wu, H.H.; Ho, C.W.; Ko, M.T.; Lin, C.Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8 (Suppl. 4), S11. [Google Scholar] [CrossRef] [Green Version]
- Scott, T.M.; Jensen, S.; Pickett, B.E. A signaling pathway-driven bioinformatics pipeline for predicting therapeutics against emerging infectious diseases. F1000Research 2021, 10, 330. [Google Scholar] [CrossRef]
- Ochoa, D.; Hercules, A.; Carmona, M.; Suveges, D.; Gonzalez-Uriarte, A.; Malangone, C.; Miranda, A.; Fumis, L.; Carvalho-Silva, D.; Spitzer, M.; et al. Open Targets Platform: Supporting systematic drug-target identification and prioritisation. Nucleic Acids Res. 2021, 49, D1302–D1310. [Google Scholar] [CrossRef]
- Kim, Y.G.; Kim, M.; Kang, J.H.; Kim, H.J.; Park, J.W.; Lee, J.M.; Suh, J.Y.; Kim, J.Y.; Lee, J.H.; Lee, Y. Transcriptome sequencing of gingival biopsies from chronic periodontitis patients reveals novel gene expression and splicing patterns. Hum. Genom. 2016, 10, 28. [Google Scholar] [CrossRef] [Green Version]
- Aevermann, B.D.; Pickett, B.E.; Kumar, S.; Klem, E.B.; Agnihothram, S.; Askovich, P.S.; Bankhead, A., 3rd; Bolles, M.; Carter, V.; Chang, J.; et al. A comprehensive collection of systems biology data characterizing the host response to viral infection. Sci. Data 2014, 1, 140033. [Google Scholar] [CrossRef] [Green Version]
- Udagawa, N.; Koide, M.; Nakamura, M.; Nakamichi, Y.; Yamashita, T.; Uehara, S.; Kobayashi, Y.; Furuya, Y.; Yasuda, H.; Fukuda, C.; et al. Osteoclast differentiation by RANKL and OPG signaling pathways. J. Bone Min. Metab. 2021, 39, 19–26. [Google Scholar] [CrossRef]
- Kim, J.M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.H. Osteoblast-Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef] [PubMed]
- Hadjidakis, D.J.; Androulakis, I.I. Bone remodeling. Ann. N. Y. Acad. Sci. 2006, 1092, 385–396. [Google Scholar] [CrossRef]
- Hienz, S.A.; Paliwal, S.; Ivanovski, S. Mechanisms of Bone Resorption in Periodontitis. J. Immunol. Res. 2015, 2015, 615486. [Google Scholar] [CrossRef] [Green Version]
- de Molon, R.S.; Park, C.H.; Jin, Q.; Sugai, J.; Cirelli, J.A. Characterization of ligature-induced experimental periodontitis. Microsc. Res. Tech. 2018, 81, 1412–1421. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, E.M.; Reis, C.; Manzanares-Cespedes, M.C. Chronic periodontitis, inflammatory cytokines, and interrelationship with other chronic diseases. Postgrad. Med. 2018, 130, 98–104. [Google Scholar] [CrossRef]
- Luckheeram, R.V.; Zhou, R.; Verma, A.D.; Xia, B. CD4(+)T cells: Differentiation and functions. Clin. Dev. Immunol. 2012, 2012, 925135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.S.; Xu, P.Z.; Gottlob, K.; Chen, M.L.; Sokol, K.; Shiyanova, T.; Roninson, I.; Weng, W.; Suzuki, R.; Tobe, K.; et al. Growth retardation and increased apoptosis in mice with homozygous disruption of the Akt1 gene. Genes Dev. 2001, 15, 2203–2208. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.; Thorvaldsen, J.L.; Chu, Q.; Feng, F.; Birnbaum, M.J. Akt1/PKBalpha is required for normal growth but dispensable for maintenance of glucose homeostasis in mice. J. Biol. Chem. 2001, 276, 38349–38352. [Google Scholar] [CrossRef] [Green Version]
- Hers, I.; Vincent, E.E.; Tavare, J.M. Akt signalling in health and disease. Cell Signal 2011, 23, 1515–1527. [Google Scholar] [CrossRef]
- Franke, T.F.; Yang, S.I.; Chan, T.O.; Datta, K.; Kazlauskas, A.; Morrison, D.K.; Kaplan, D.R.; Tsichlis, P.N. The protein kinase encoded by the Akt proto-oncogene is a target of the PDGF-activated phosphatidylinositol 3-kinase. Cell 1995, 81, 727–736. [Google Scholar] [CrossRef] [Green Version]
- Laouar, Y.; Welte, T.; Fu, X.Y.; Flavell, R.A. STAT3 is required for Flt3L-dependent dendritic cell differentiation. Immunity 2003, 19, 903–912. [Google Scholar] [CrossRef] [Green Version]
- Esashi, E.; Wang, Y.H.; Perng, O.; Qin, X.F.; Liu, Y.J.; Watowich, S.S. The signal transducer STAT5 inhibits plasmacytoid dendritic cell development by suppressing transcription factor IRF8. Immunity 2008, 28, 509–520. [Google Scholar] [CrossRef] [Green Version]
- D’Amico, A.; Wu, L. The early progenitors of mouse dendritic cells and plasmacytoid predendritic cells are within the bone marrow hemopoietic precursors expressing Flt3. J. Exp. Med. 2003, 198, 293–303. [Google Scholar] [CrossRef] [Green Version]
- Onai, N.; Obata-Onai, A.; Tussiwand, R.; Lanzavecchia, A.; Manz, M.G. Activation of the Flt3 signal transduction cascade rescues and enhances type I interferon-producing and dendritic cell development. J. Exp. Med. 2006, 203, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Hillmer, E.J.; Zhang, H.; Li, H.S.; Watowich, S.S. STAT3 signaling in immunity. Cytokine Growth Factor Rev. 2016, 31, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, W.C.; Levy, D.E.; Lee, C.K. STAT3 positively regulates an early step in B-cell development. Blood 2006, 108, 3005–3011. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Ivanov, I.I.; Spolski, R.; Min, R.; Shenderov, K.; Egawa, T.; Levy, D.E.; Leonard, W.J.; Littman, D.R. IL-6 programs T(H)-17 cell differentiation by promoting sequential engagement of the IL-21 and IL-23 pathways. Nat. Immunol. 2007, 8, 967–974. [Google Scholar] [CrossRef]
- Nurieva, R.; Yang, X.O.; Martinez, G.; Zhang, Y.; Panopoulos, A.D.; Ma, L.; Schluns, K.; Tian, Q.; Watowich, S.S.; Jetten, A.M.; et al. Essential autocrine regulation by IL-21 in the generation of inflammatory T cells. Nature 2007, 448, 480–483. [Google Scholar] [CrossRef]
- Nurieva, R.I.; Chung, Y.; Hwang, D.; Yang, X.O.; Kang, H.S.; Ma, L.; Wang, Y.H.; Watowich, S.S.; Jetten, A.M.; Tian, Q.; et al. Generation of T follicular helper cells is mediated by interleukin-21 but independent of T helper 1, 2, or 17 cell lineages. Immunity 2008, 29, 138–149. [Google Scholar] [CrossRef] [Green Version]
- Batten, M.; Ramamoorthi, N.; Kljavin, N.M.; Ma, C.S.; Cox, J.H.; Dengler, H.S.; Danilenko, D.M.; Caplazi, P.; Wong, M.; Fulcher, D.A.; et al. IL-27 supports germinal center function by enhancing IL-21 production and the function of T follicular helper cells. J. Exp. Med. 2010, 207, 2895–2906. [Google Scholar] [CrossRef]
- Cui, W.; Liu, Y.; Weinstein, J.S.; Craft, J.; Kaech, S.M. An interleukin-21-interleukin-10-STAT3 pathway is critical for functional maturation of memory CD8+ T cells. Immunity 2011, 35, 792–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaech, S.M.; Cui, W. Transcriptional control of effector and memory CD8+ T cell differentiation. Nat. Rev. Immunol. 2012, 12, 749–761. [Google Scholar] [CrossRef] [PubMed]
- Vergadi, E.; Ieronymaki, E.; Lyroni, K.; Vaporidi, K.; Tsatsanis, C. Akt Signaling Pathway in Macrophage Activation and M1/M2 Polarization. J. Immunol. 2017, 198, 1006–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pompura, S.L.; Dominguez-Villar, M. The PI3K/AKT signaling pathway in regulatory T-cell development, stability, and function. J. Leukoc. Biol. 2018, 103, 1065–1076. [Google Scholar] [CrossRef]
- Xu, K.; Yin, N.; Peng, M.; Stamatiades, E.G.; Shyu, A.; Li, P.; Zhang, X.; Do, M.H.; Wang, Z.; Capistrano, K.J.; et al. Glycolysis fuels phosphoinositide 3-kinase signaling to bolster T cell immunity. Science 2021, 371, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Ouyang, G.; Bao, S. The activation of Akt/PKB signaling pathway and cell survival. J. Cell. Mol. Med. 2005, 9, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.X.; Wang, B.; Li, B. IL-10 and IL-22 in Mucosal Immunity: Driving Protection and Pathology. Front. Immunol. 2020, 11, 1315. [Google Scholar] [CrossRef]
- Ho, I.C.; Miaw, S.C. Regulation of IL-4 Expression in Immunity and Diseases. Adv. Exp. Med. Biol. 2016, 941, 31–77. [Google Scholar] [CrossRef]
- Gadani, S.P.; Cronk, J.C.; Norris, G.T.; Kipnis, J. IL-4 in the brain: A cytokine to remember. J. Immunol. 2012, 189, 4213–4219. [Google Scholar] [CrossRef]
- Howard, M.; Farrar, J.; Hilfiker, M.; Johnson, B.; Takatsu, K.; Hamaoka, T.; Paul, W.E. Identification of a T cell-derived b cell growth factor distinct from interleukin 2. J. Exp. Med. 1982, 155, 914–923. [Google Scholar] [CrossRef] [Green Version]
- Geha, R.S.; Jabara, H.H.; Brodeur, S.R. The regulation of immunoglobulin E class-switch recombination. Nat. Rev. Immunol. 2003, 3, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.S.; Heimberger, A.B.; Gold, J.S.; O’Garra, A.; Murphy, K.M. Differential regulation of T helper phenotype development by interleukins 4 and 10 in an alpha beta T-cell-receptor transgenic system. Proc. Natl. Acad. Sci. USA 1992, 89, 6065–6069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges da Silva, H.; Fonseca, R.; Alvarez, J.M.; D’Imperio Lima, M.R. IFN-gamma Priming Effects on the Maintenance of Effector Memory CD4(+) T Cells and on Phagocyte Function: Evidences from Infectious Diseases. J. Immunol. Res. 2015, 2015, 202816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, P.L.; de Jong, E.C.; Wierenga, E.A.; Kapsenberg, M.L.; Kalinski, P. Development of Th1-inducing capacity in myeloid dendritic cells requires environmental instruction. J. Immunol. 2000, 164, 4507–4512. [Google Scholar] [CrossRef] [Green Version]
- Taylor, K.R.; Trowbridge, J.M.; Rudisill, J.A.; Termeer, C.C.; Simon, J.C.; Gallo, R.L. Hyaluronan fragments stimulate endothelial recognition of injury through TLR4. J. Biol. Chem. 2004, 279, 17079–17084. [Google Scholar] [CrossRef] [Green Version]
- Traboulsee, A.; Greenberg, B.M.; Bennett, J.L.; Szczechowski, L.; Fox, E.; Shkrobot, S.; Yamamura, T.; Terada, Y.; Kawata, Y.; Wright, P.; et al. Safety and efficacy of satralizumab monotherapy in neuromyelitis optica spectrum disorder: A randomised, double-blind, multicentre, placebo-controlled phase 3 trial. Lancet Neurol. 2020, 19, 402–412. [Google Scholar] [CrossRef]
- Heo, Y.A. Satralizumab: First Approval. Drugs 2020, 80, 1477–1482. [Google Scholar] [CrossRef]
- Bone, H.G.; Wagman, R.B.; Brandi, M.L.; Brown, J.P.; Chapurlat, R.; Cummings, S.R.; Czerwinski, E.; Fahrleitner-Pammer, A.; Kendler, D.L.; Lippuner, K.; et al. 10 years of denosumab treatment in postmenopausal women with osteoporosis: Results from the phase 3 randomised FREEDOM trial and open-label extension. Lancet Diabetes Endocrinol. 2017, 5, 513–523. [Google Scholar] [CrossRef]
- Papp, K.A.; Reich, K.; Paul, C.; Blauvelt, A.; Baran, W.; Bolduc, C.; Toth, D.; Langley, R.G.; Cather, J.; Gottlieb, A.B.; et al. A prospective phase III, randomized, double-blind, placebo-controlled study of brodalumab in patients with moderate-to-severe plaque psoriasis. Br. J. Derm. 2016, 175, 273–286. [Google Scholar] [CrossRef]
- Henry, D.H.; Costa, L.; Goldwasser, F.; Hirsh, V.; Hungria, V.; Prausova, J.; Scagliotti, G.V.; Sleeboom, H.; Spencer, A.; Vadhan-Raj, S.; et al. Randomized, double-blind study of denosumab versus zoledronic acid in the treatment of bone metastases in patients with advanced cancer (excluding breast and prostate cancer) or multiple myeloma. J. Clin. Oncol. 2011, 29, 1125–1132. [Google Scholar] [CrossRef] [Green Version]
- Van den Wyngaert, T.; Wouters, K.; Huizing, M.T.; Vermorken, J.B. RANK ligand inhibition in bone metastatic cancer and risk of osteonecrosis of the jaw (ONJ): Non bis in idem? Support Care Cancer 2011, 19, 2035–2040. [Google Scholar] [CrossRef] [PubMed]
- Papapoulos, S.; Chapurlat, R.; Libanati, C.; Brandi, M.L.; Brown, J.P.; Czerwinski, E.; Krieg, M.A.; Man, Z.; Mellstrom, D.; Radominski, S.C.; et al. Five years of denosumab exposure in women with postmenopausal osteoporosis: Results from the first two years of the FREEDOM extension. J. Bone Min. Res. 2012, 27, 694–701. [Google Scholar] [CrossRef]
- Aljohani, S.; Fliefel, R.; Ihbe, J.; Kuhnisch, J.; Ehrenfeld, M.; Otto, S. What is the effect of anti-resorptive drugs (ARDs) on the development of medication-related osteonecrosis of the jaw (MRONJ) in osteoporosis patients: A systematic review. J. Craniomaxillofac. Surg. 2017, 45, 1493–1502. [Google Scholar] [CrossRef] [PubMed]
- Bagan, J.; Peydro, A.; Calvo, J.; Leopoldo, M.; Jimenez, Y.; Bagan, L. Medication-related osteonecrosis of the jaw associated with bisphosphonates and denosumab in osteoporosis. Oral Dis. 2016, 22, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Diniz-Freitas, M.; Fernandez-Feijoo, J.; Diz Dios, P.; Pousa, X.; Limeres, J. Denosumab-related osteonecrosis of the jaw following non-surgical periodontal therapy: A case report. J. Clin. Periodontol. 2018, 45, 570–577. [Google Scholar] [CrossRef]
- McGowan, K.; McGowan, T.; Ivanovski, S. Risk factors for medication-related osteonecrosis of the jaws: A systematic review. Oral Dis. 2018, 24, 527–536. [Google Scholar] [CrossRef]
- Yamazaki, T.; Yamori, M.; Ishizaki, T.; Asai, K.; Goto, K.; Takahashi, K.; Nakayama, T.; Bessho, K. Increased incidence of osteonecrosis of the jaw after tooth extraction in patients treated with bisphosphonates: A cohort study. Int. J. Oral Maxillofac Surg. 2012, 41, 1397–1403. [Google Scholar] [CrossRef] [Green Version]
- Kappos, L.; Polman, C.H.; Freedman, M.S.; Edan, G.; Hartung, H.P.; Miller, D.H.; Montalban, X.; Barkhof, F.; Bauer, L.; Jakobs, P.; et al. Treatment with interferon beta-1b delays conversion to clinically definite and McDonald MS in patients with clinically isolated syndromes. Neurology 2006, 67, 1242–1249. [Google Scholar] [CrossRef]
- Edan, G.; Kappos, L.; Montalban, X.; Polman, C.H.; Freedman, M.S.; Hartung, H.P.; Miller, D.; Barkhof, F.; Herrmann, J.; Lanius, V.; et al. Long-term impact of interferon beta-1b in patients with CIS: 8-year follow-up of BENEFIT. J. Neurol. Neurosurg. Psychiatry 2014, 85, 1183–1189. [Google Scholar] [CrossRef]
- Liu, Z.; Pelfrey, C.M.; Cotleur, A.; Lee, J.C.; Rudick, R.A. Immunomodulatory effects of interferon beta-1a in multiple sclerosis. J. Neuroimmunol. 2001, 112, 153–162. [Google Scholar] [CrossRef]
- Kieseier, B.C. The mechanism of action of interferon-beta in relapsing multiple sclerosis. CNS Drugs 2011, 25, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Chen, G.; Nie, H.; Zhang, X.; Niu, X.; Zang, Y.C.; Skinner, S.M.; Zhang, J.Z.; Killian, J.M.; Hong, J. Regulatory effects of IFN-beta on production of osteopontin and IL-17 by CD4+ T Cells in MS. Eur. J. Immunol. 2009, 39, 2525–2536. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, N.; Tsuchimori, N.; Matsumoto, T.; Ii, M. TAK-242 (resatorvid), a small-molecule inhibitor of Toll-like receptor (TLR) 4 signaling, binds selectively to TLR4 and interferes with interactions between TLR4 and its adaptor molecules. Mol. Pharm. 2011, 79, 34–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, T.W.; Wheeler, A.P.; Bernard, G.R.; Vincent, J.L.; Angus, D.C.; Aikawa, N.; Demeyer, I.; Sainati, S.; Amlot, N.; Cao, C.; et al. A randomized, double-blind, placebo-controlled trial of TAK-242 for the treatment of severe sepsis. Crit. Care Med. 2010, 38, 1685–1694. [Google Scholar] [CrossRef]
- Smedley, D.; Haider, S.; Ballester, B.; Holland, R.; London, D.; Thorisson, G.; Kasprzyk, A. BioMart--biological queries made easy. BMC Genom. 2009, 10, 22. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| GEO Gene Set ID | GSE173082 | GSE80715 |
|---|---|---|
| Title | Differential DNA methylation and mRNA expression in gingival tissues in periodontal health and disease | Transcriptome analysis of chronic periodontitis patients’ gingival tissue |
| Platform | Illumina HiSeq 4000 | Illumina HiSeq 2000 |
| Library Construction Protocol | Gingival tissue samples were harvested in conjunction with an invasive oral surgical procedure required for the participant’s oral care after administration of local anesthesia. Poly-A pull-down was carried out to enrich mRNAs from total RNA samples (200 ng–1 g per sample) followed by library preparation using the Illumina TruSeq RNA prep kit. | Frozen tissues were disrupted in the lysis solution of mirVana RNA isolation kit (Thermo Fisher Scientific) using disposable pestle grinder system (Thermo Fisher Scientific). After purification of mRNA molecules by poly-T oligo-attached magnetic beads followed by fragmentation, the RNA of approximately 300 bp size was isolated using gel electrophoresis. The cDNA synthesis and library construction were performed using the Illumina Truseq RNA sample preparation kit (Illumina, San Diego, CA, USA) following the manufacturer’s protocol. |
| Sample Type | Single End | Paired End |
| Diagnostic criteria | Not recorded | On the basis of clinical and radiographic criteria, periodontitis-affected site had a probing depth of ≥4 mm, clinical attachment level of ≥4 mm, and bleeding on probing. |
| Sample Prep | Not recorded | The size of 3 mm2 gingival biopsies were obtained from the marginal gingiva during periodontal flap surgery and immediately stored in RNAlater solution (Thermo Fisher Scientific, Waltham, MA, USA) at −70 °C after removal of blood by brief washing in phosphate-buffered saline. |
| Number of healthy samples vs. periodontitis samples | 12 vs. 12 | 10 vs. 10 |
| Number of healthy patients | Not recorded | nine periodontal healthy patients with pocket depth < 4 mm |
| Number of periodontitis patients | Not recorded | four periodontitis patients with pocket depth of 4–6 mm; three severe periodontitis patients with pocket depth of 7 mm or deeper |
| PubMed ID | Not published | 27531006 [48] |
| Ensembl Gene ID | Symbol | Description | logFC * | logCPM ** | p-Value | FDR *** | |
|---|---|---|---|---|---|---|---|
| 1 | ENSG00000099958 | DERL3 | Derlin 3 | 3.92 | 4.40 | 3.45 × 10−5 | 2.92 × 10−2 |
| 2 | ENSG00000170476 | MZB1 | Marginal zone B and B1 cell specific protein | 3.98 | 5.29 | 4.48 × 10−5 | 2.92 × 10−2 |
| 3 | ENSG00000153208 | MERTK | MER proto-oncogene, tyrosine kinase | 1.58 | 1.32 | 4.56 × 10−5 | 2.92 × 10−2 |
| 4 | ENSG00000183508 | TENT5C | Terminal nucleotidyltransferase 5C | 3.11 | 5.28 | 6.44 × 10−5 | 2.98 × 10−2 |
| 5 | ENSG00000198794 | SCAMP5 | Secretory carrier membrane protein 5 | 2.67 | 2.54 | 6.79 × 10−5 | 3.03 × 10−2 |
| 6 | ENSG00000137265 | IRF4 | Interferon regulatory factor 4 | 3.14 | 4.02 | 7.33 × 10−5 | 3.03 × 10−2 |
| 7 | ENSG00000061656 | SPAG4 | Sperm associated antigen 4 | 3.28 | 1.63 | 7.87 × 10−5 | 3.03 × 10−2 |
| 8 | ENSG00000112936 | C7 | Complement C7 | 2.46 | −0.06 | 8.31 × 10−5 | 3.03 × 10−2 |
| 9 | ENSG00000100219 | XBP1 | X-box binding protein 1 | 1.81 | 7.87 | 8.99 × 10−5 | 3.05 × 10−2 |
| 10 | ENSG00000065413 | ANKRD44 | Ankyrin repeat domain 44 | 1.41 | 3.31 | 9.72 × 10−5 | 3.05 × 10−2 |
| 11 | ENSG00000117322 | CR2 | Complement C3d receptor 2 | 5.14 | 0.83 | 1.19 × 10−4 | 3.39 × 10−2 |
| 12 | ENSG00000189233 | NUGGC | Nuclear GTPase, germinal center associated | 2.02 | 0.63 | 1.25 × 10−4 | 3.48 × 10−2 |
| 13 | ENSG00000134285 | FKBP11 | FKBP prolyl isomerase 11 | 2.02 | 4.53 | 1.34 × 10−4 | 3.57 × 10−2 |
| 14 | ENSG00000102096 | PIM2 | Pim-2 proto-oncogene, serine/threonine kinase | 2.60 | 4.33 | 1.58 × 10−4 | 3.94 × 10−2 |
| 15 | ENSG00000198018 | ENTPD7 | Ectonucleoside triphosphate diphosphohydrolase 7 | 0.70 | 4.47 | 1.70 × 10−4 | 4.15 × 10−2 |
| 16 | ENSG00000130768 | SMPDL3B | Sphingomyelin phosphodiesterase acid like 3B | 1.99 | 1.39 | 1.76 × 10−4 | 4.20 × 10−2 |
| 17 | ENSG00000101194 | SLC17A9 | Solute carrier family 17 member 9 | 2.46 | 1.75 | 1.84 × 10−4 | 4.25 × 10−2 |
| 18 | ENSG00000153162 | BMP6 | Bone morphogenetic protein 6 | 1.89 | 2.26 | 1.95 × 10−4 | 4.30 × 10−2 |
| 19 | ENSG00000073849 | ST6GAL1 | ST6 beta-galactoside alpha-2,6-sialyltransferase 1 | 2.12 | 5.13 | 1.97 × 10−4 | 4.30 × 10−2 |
| 20 | ENSG00000198854 | C1orf68 | Chromosome 1 open reading frame 68 | −3.78 | 1.50 | 2.03 × 10−4 | 4.36 × 10−2 |
| 21 | ENSG00000122188 | LAX1 | Lymphocyte transmembrane adaptor 1 | 2.77 | 2.44 | 2.33 × 10−4 | 4.81 × 10−2 |
| 22 | ENSG00000091490 | SEL1L3 | SEL1L family member 3 | 2.15 | 4.70 | 2.45 × 10−4 | 4.92 × 10−2 |
| Name | pSize | NDE | pNDE | tA | pPERT | pG | pGFdr | pGFWER | Status | SourceDB | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Cytokine–cytokine receptor interaction | 177 | 39 | 1.27 × 10−5 | 13.49 | 1.20 × 10−3 | 2.90 × 10−7 | 5.01 × 10−5 | 5.01 × 10−5 | Activated | KEGG |
| 2 | Staphylococcus aureus infection | 29 | 13 | 3.66 × 10−6 | 9.22 | 1.55 × 10−1 | 8.72 × 10−6 | 7.55 × 10−4 | 1.51 × 10−3 | Activated | KEGG |
| 3 | Natural killer cell-mediated cytotoxicity | 95 | 23 | 1.70 × 10−4 | 47.51 | 1.28 × 10−2 | 3.06 × 10−5 | 1.47 × 10−3 | 5.29 × 10−3 | Activated | KEGG |
| 4 | Chemokine signaling pathway | 157 | 30 | 1.52 × 10−3 | 31.64 | 1.60 × 10−3 | 3.39 × 10−5 | 1.47 × 10−3 | 5.87 × 10−3 | Activated | KEGG |
| 5 | Osteoclast differentiation | 108 | 25 | 1.94 × 10−4 | 13.41 | 6.88 × 10−2 | 1.63 × 10−4 | 5.65 × 10−3 | 2.83 × 10−2 | Activated | KEGG |
| 6 | Leukocyte transendothelial migration | 76 | 19 | 3.94 × 10−4 | 19.35 | 4.92 × 10−2 | 2.30 × 10−4 | 6.63 × 10−3 | 3.98 × 10−2 | Activated | KEGG |
| 7 | Keratinization | 90 | 27 | 6.00 × 10−7 | −1.90 | 2.00 × 10−1 | 2.03 × 10−6 | 1.43 × 10−3 | 1.43 × 10−3 | Inhibited | Reactome |
| 8 | Innate Immune System | 633 | 101 | 4.65 × 10−5 | 67.93 | 7.80 × 10−2 | 4.90 × 10−5 | 1.01 × 10−2 | 3.47 × 10−2 | Activated | Reactome |
| 9 | Assembly of collagen fibrils and other multimeric structures | 45 | 12 | 2.51 × 10−3 | 7.31 | 1.60 × 10−3 | 5.38 × 10−5 | 1.01 × 10−2 | 3.80 × 10−2 | Activated | Reactome |
| 10 | Formation of the cornified envelope | 62 | 19 | 1.96 × 10−5 | −1.89 | 2.18 × 10−1 | 5.71 × 10−5 | 1.01 × 10−2 | 4.03 × 10−2 | Inhibited | Reactome |
| Target Symbol | Target Name(s) | Drug ID | Drug Name | Approved by FDA | Highest Clinical Trial Phase | Health Condition Investigated | |
|---|---|---|---|---|---|---|---|
| 1 | IL6R; IL6ST | Interleukin 6 receptor; Interleukin 6 cytokine family signal transducer | CHEMBL3833307 | Satralizumab | TRUE | 4 | AQP4 antibody-positive Neuromyelitis optica spectrum disorder (NMOSD) |
| 2 | TNFSF11 | TNF superfamily member 11 (RANKL) | CHEMBL1237023 | Denosumab | TRUE | 4 | Postmenopausal osteoporosis |
| 3 | IFNAR2 | Interferon alpha and beta receptor subunit 2 | CHEMBL1201563 | Interferon Beta-1B | TRUE | 4 | Relapsing-remitting forms of multiple sclerosis |
| 4 | IL17RA | Interleukin 17 receptor A | CHEMBL1742996 | Brodalumab | TRUE | 4 | Moderate to severe plaque psoriasis |
| 5 | TLR4 | Toll-like receptor 4 | CHEMBL225157 | Resatorvid | FALSE | 3 | Severe sepsis |
| 6 | IL6 | Interleukin 6 | CHEMBL2108589 | Clazakizumab | FALSE | 3 | Kidney failure, antibody-mediated rejection of kidney transplants, rheumatoid arthritis, asthma, Crohn’s disease, psoriatic arthritis, and COVID-19. |
| 7 | IL1B | Interleukin 1 beta | CHEMBL1743026 | Gevokizumab | FALSE | 3 | Scleritis, colon cancer, osteoarthritis, chronic uveitis, Pyoderma Gangrenosum, gastroesophageal cancer, renal cell carcinoma, rheumatoid arthritis, Muckle–Wells syndrome, Behcet’s disease, and Type I and Type II Diabetes |
| 8 | TGFBR1 | Transforming growth factor beta receptor 1 | CHEMBL2364611 | Galunisertib | FALSE | 2 | Metastatic pancreatic cancer, colorectal cancer, advanced hepatocellular carcinoma, prostate cancer, ovarian carcinosarcoma, rectal adenocarcinoma, breast cancer, nasopharyngeal cancer, and glioblastoma |
| 9 | CSF2RB | Colony stimulating factor 2 receptor subunit beta | CHEMBL1743039 | Mavrilimumab | FALSE | 2 | Rheumatoid arthritis; acute respiratory failure and hyperinflammation in COVID-19 |
| 10 | CSF2 | Colony stimulating factor 2 | CHEMBL2109430 | Gimsilumab | FALSE | 2 | Ankylosing spondylitis; COVID-19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno, C.; Bybee, E.; Tellez Freitas, C.M.; Pickett, B.E.; Weber, K.S. Meta-Analysis of Two Human RNA-seq Datasets to Determine Periodontitis Diagnostic Biomarkers and Drug Target Candidates. Int. J. Mol. Sci. 2022, 23, 5580. https://doi.org/10.3390/ijms23105580
Moreno C, Bybee E, Tellez Freitas CM, Pickett BE, Weber KS. Meta-Analysis of Two Human RNA-seq Datasets to Determine Periodontitis Diagnostic Biomarkers and Drug Target Candidates. International Journal of Molecular Sciences. 2022; 23(10):5580. https://doi.org/10.3390/ijms23105580
Chicago/Turabian StyleMoreno, Carlos, Ellie Bybee, Claudia M. Tellez Freitas, Brett E. Pickett, and K. Scott Weber. 2022. "Meta-Analysis of Two Human RNA-seq Datasets to Determine Periodontitis Diagnostic Biomarkers and Drug Target Candidates" International Journal of Molecular Sciences 23, no. 10: 5580. https://doi.org/10.3390/ijms23105580
APA StyleMoreno, C., Bybee, E., Tellez Freitas, C. M., Pickett, B. E., & Weber, K. S. (2022). Meta-Analysis of Two Human RNA-seq Datasets to Determine Periodontitis Diagnostic Biomarkers and Drug Target Candidates. International Journal of Molecular Sciences, 23(10), 5580. https://doi.org/10.3390/ijms23105580