
Green Synthesis of Silymarin–Chitosan Nanoparticles as a New Nano Formulation with Enhanced Anti-Fibrotic Effects against Liver Fibrosis

 and
and
Abstract
:1. Introduction
2. Results
2.1. Characterization of Nanoparticles
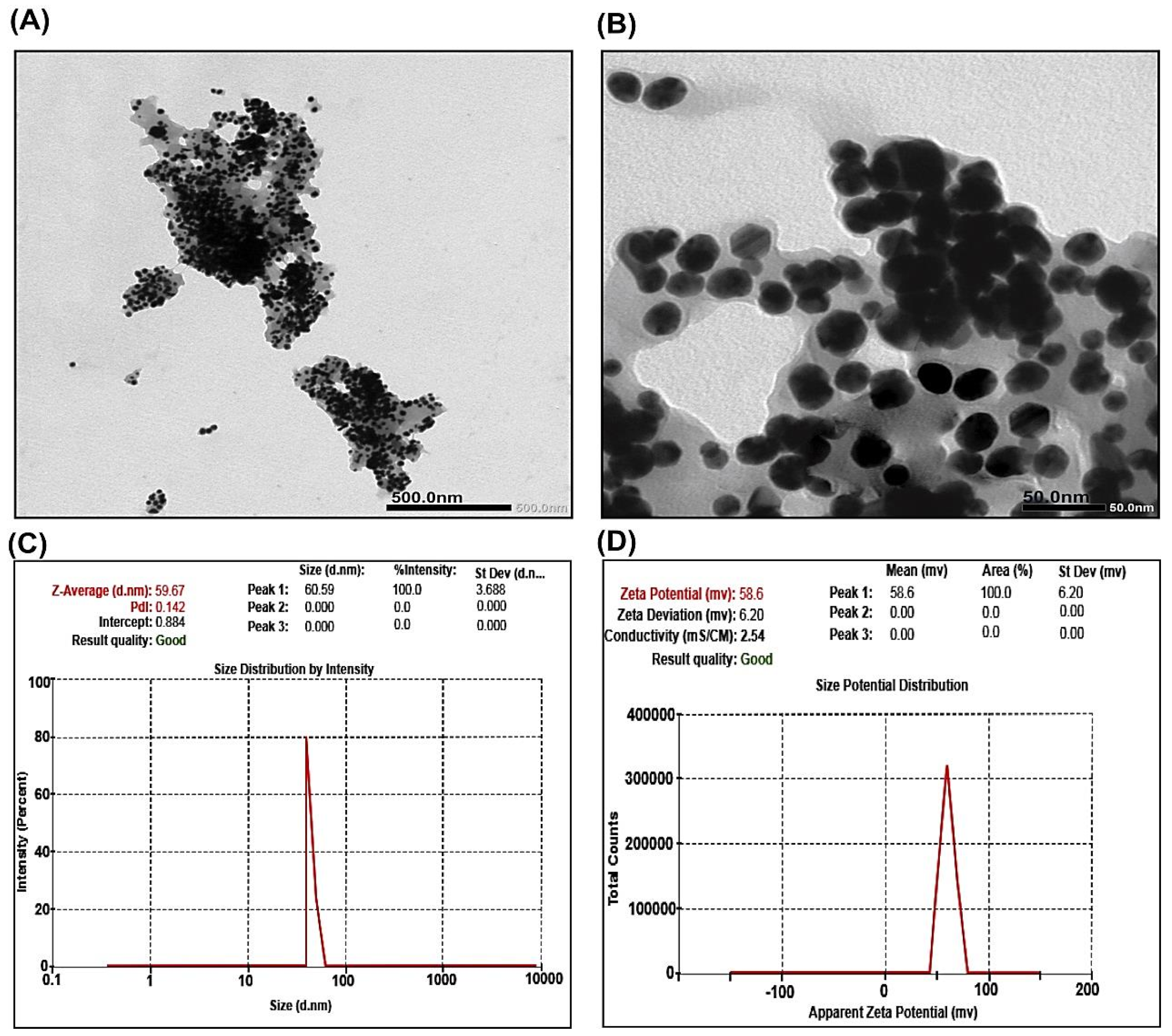
2.1.1. TEM Analysis of CNPs
2.1.2. DLS and Zeta Potential Measurements of CNPs
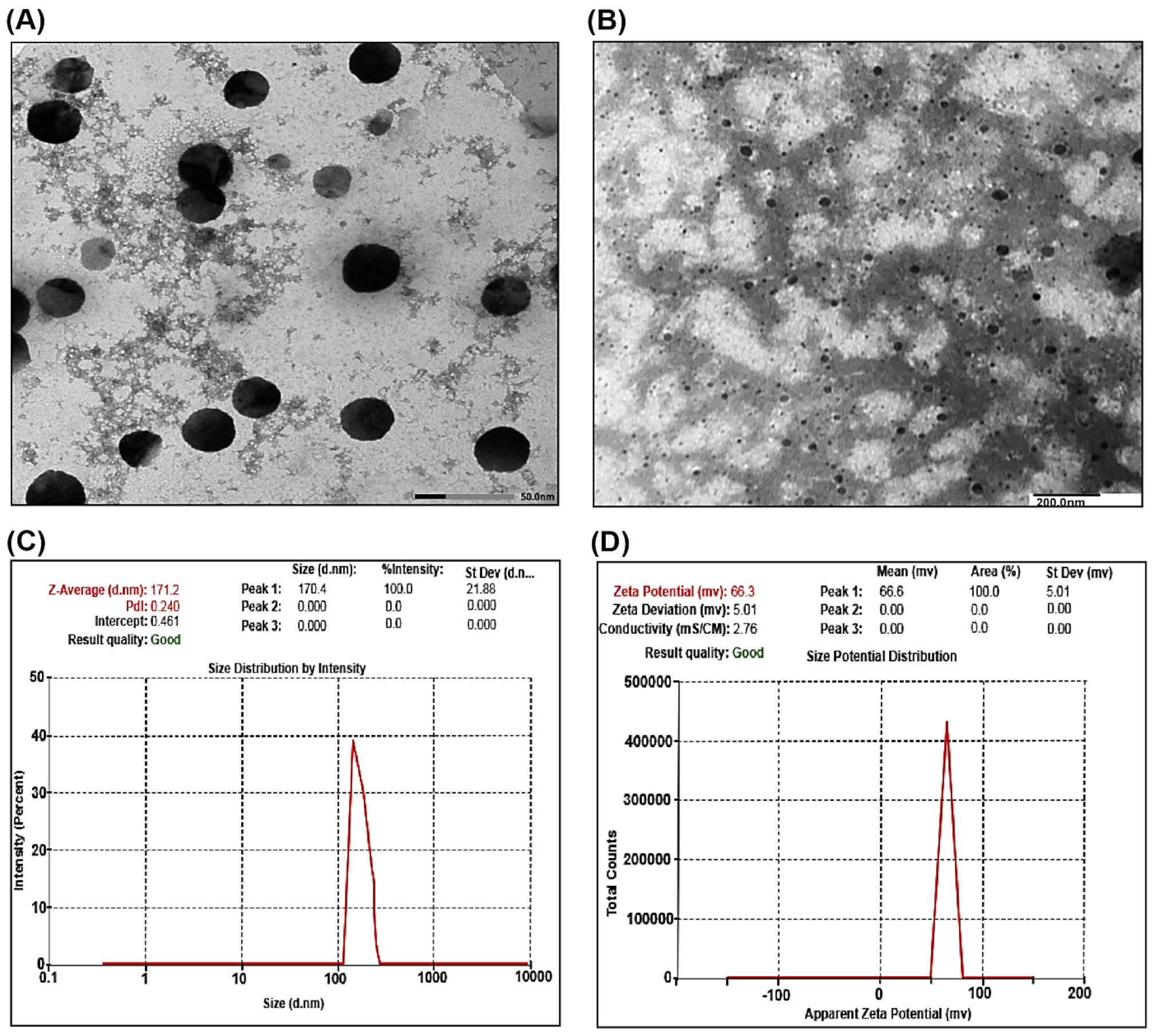
2.1.3. TEM Analysis of SCNPs
2.1.4. DLS and Zeta Potential Measurements of SCNPs
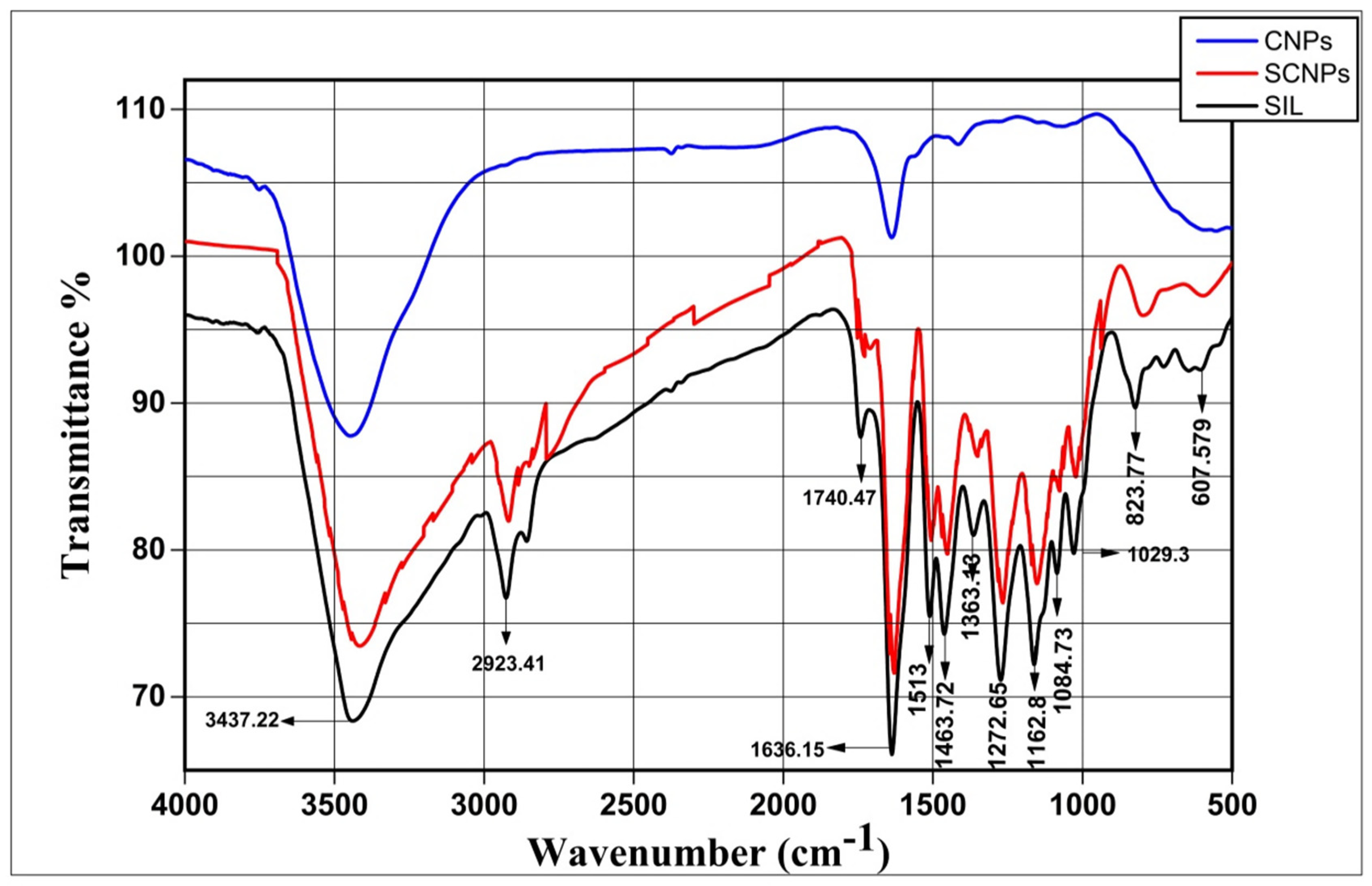
2.1.5. FT-IR Study
2.1.6. Drug Entrapment Efficiency and Loading Capacity of SIL in SCNPs
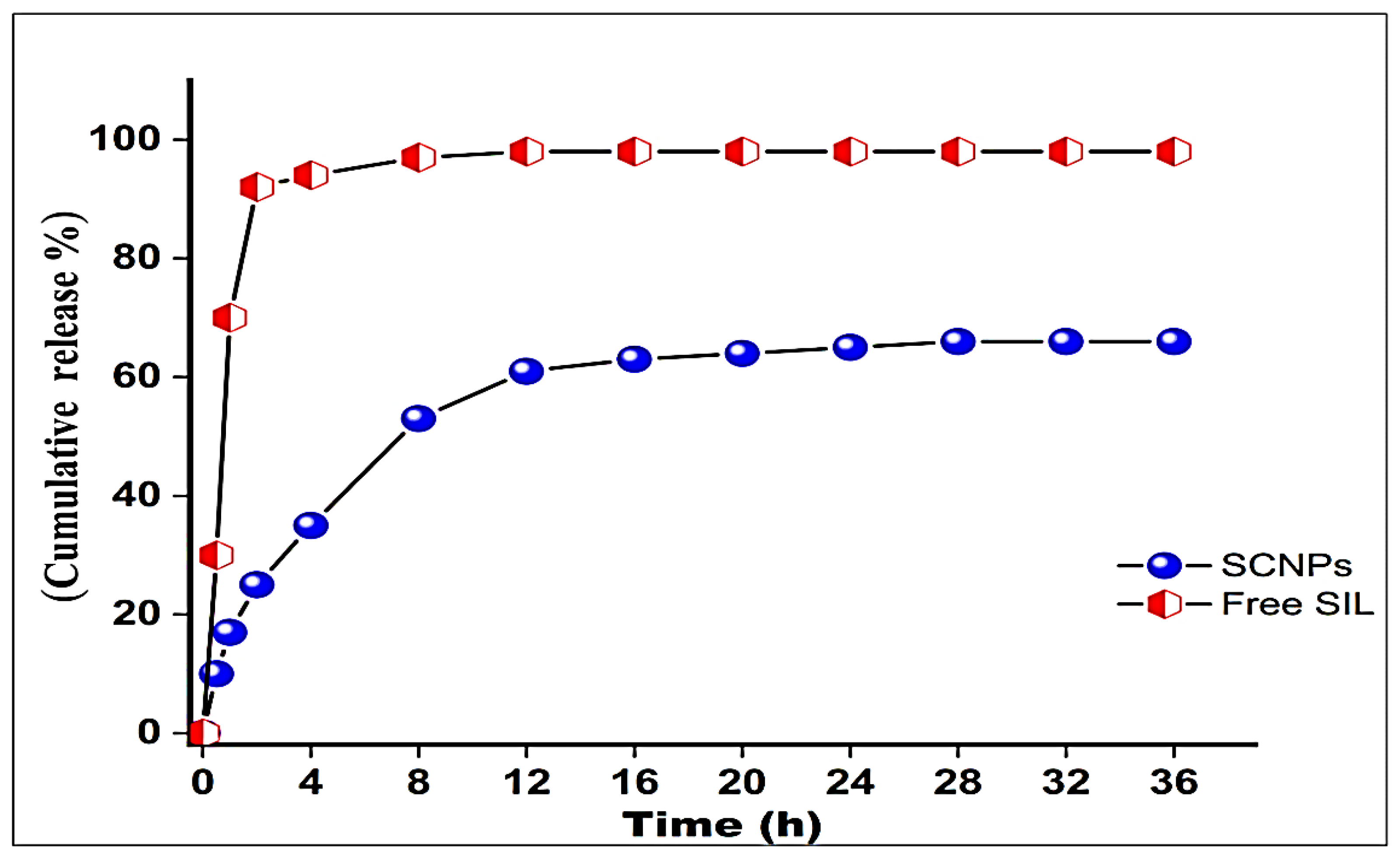
2.1.7. In Vitro Drug Release Study
2.2. In Vivo Studies
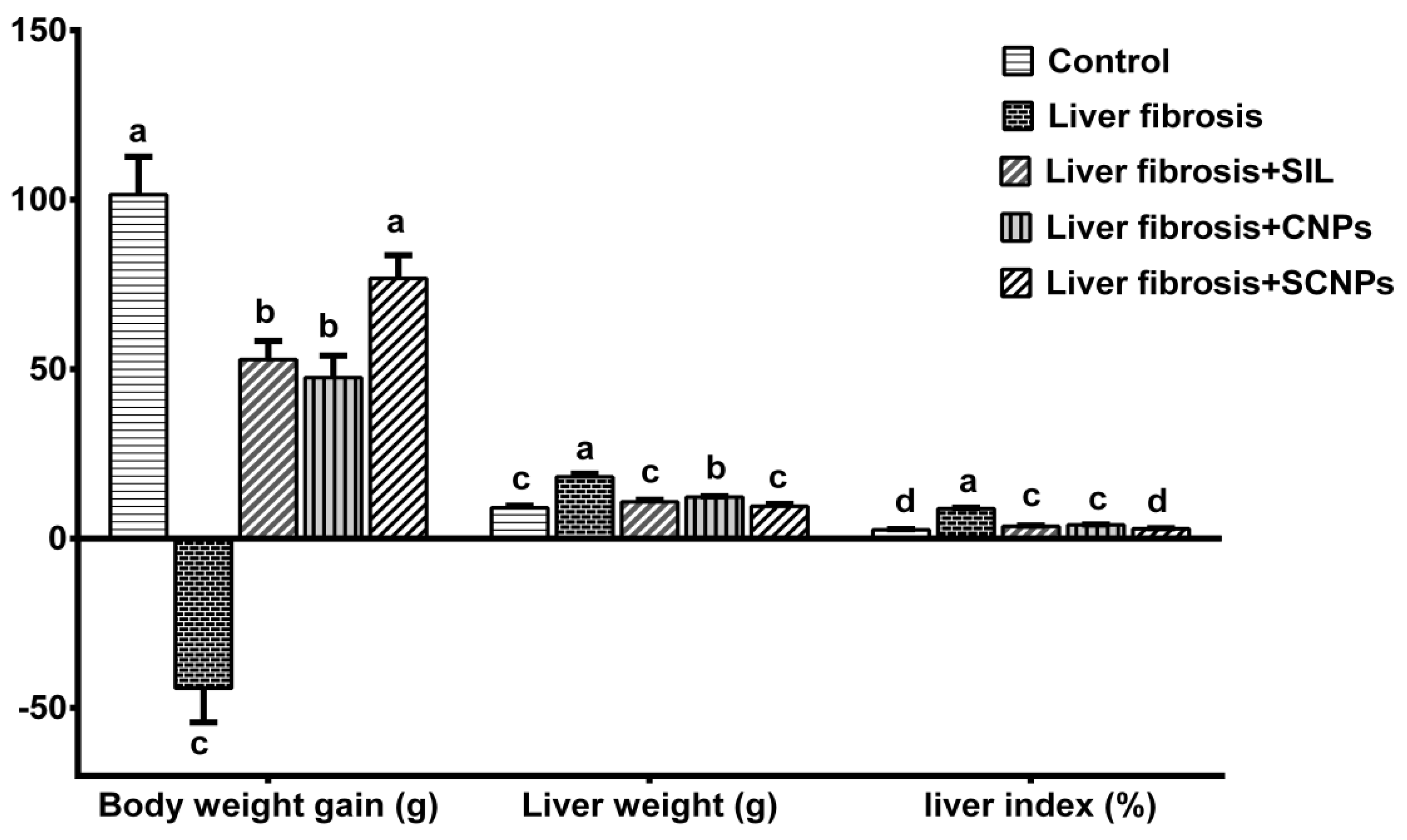
2.2.1. Body Weight Gain, Liver Weight, and Liver Index (%)
2.2.2. Serum Liver Function Markers
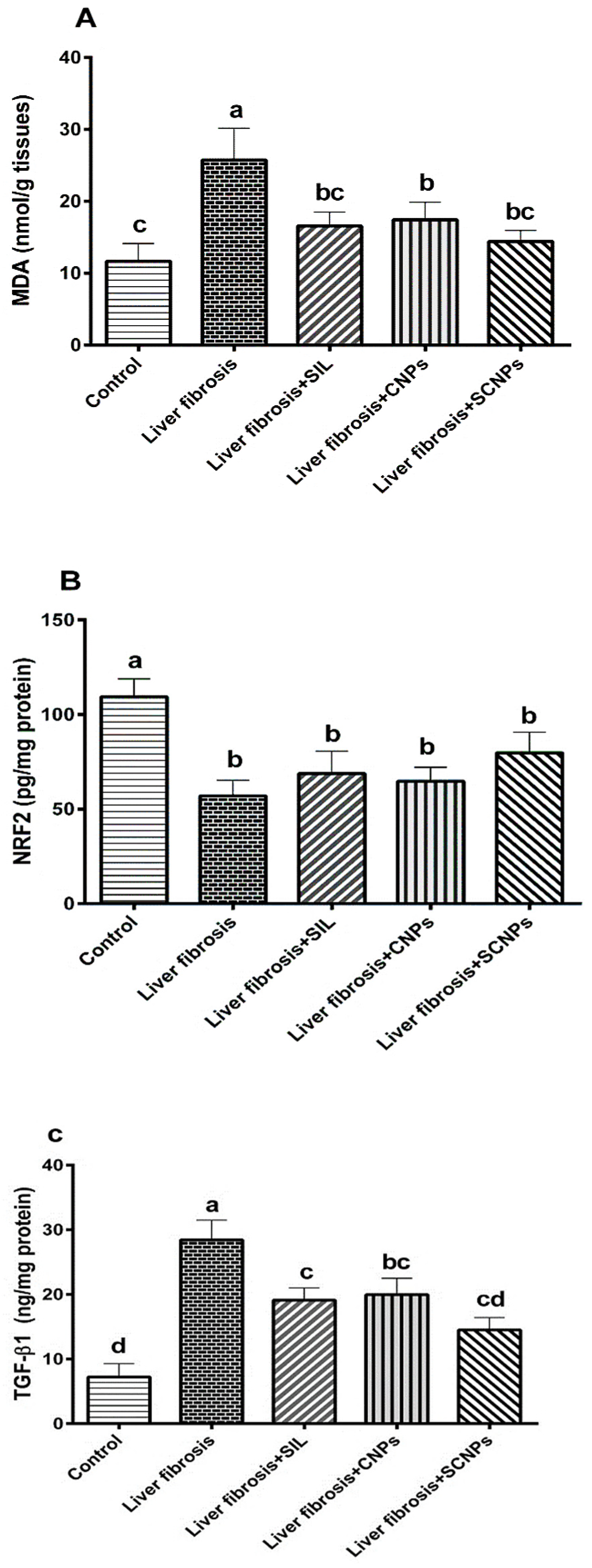
2.2.3. Hepatic Redox Parameters and TGFβ-1
2.3. Molecular Analysis
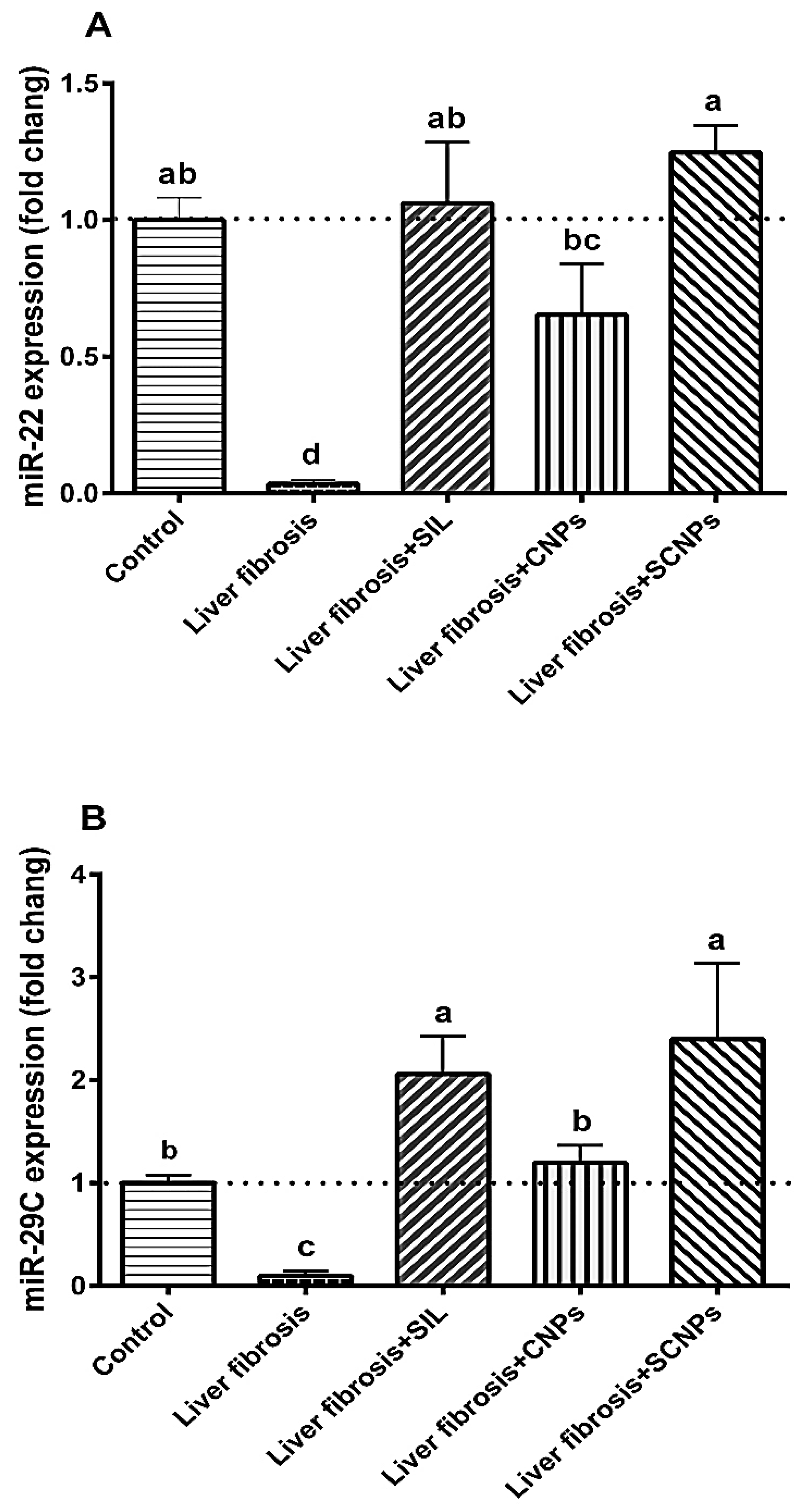
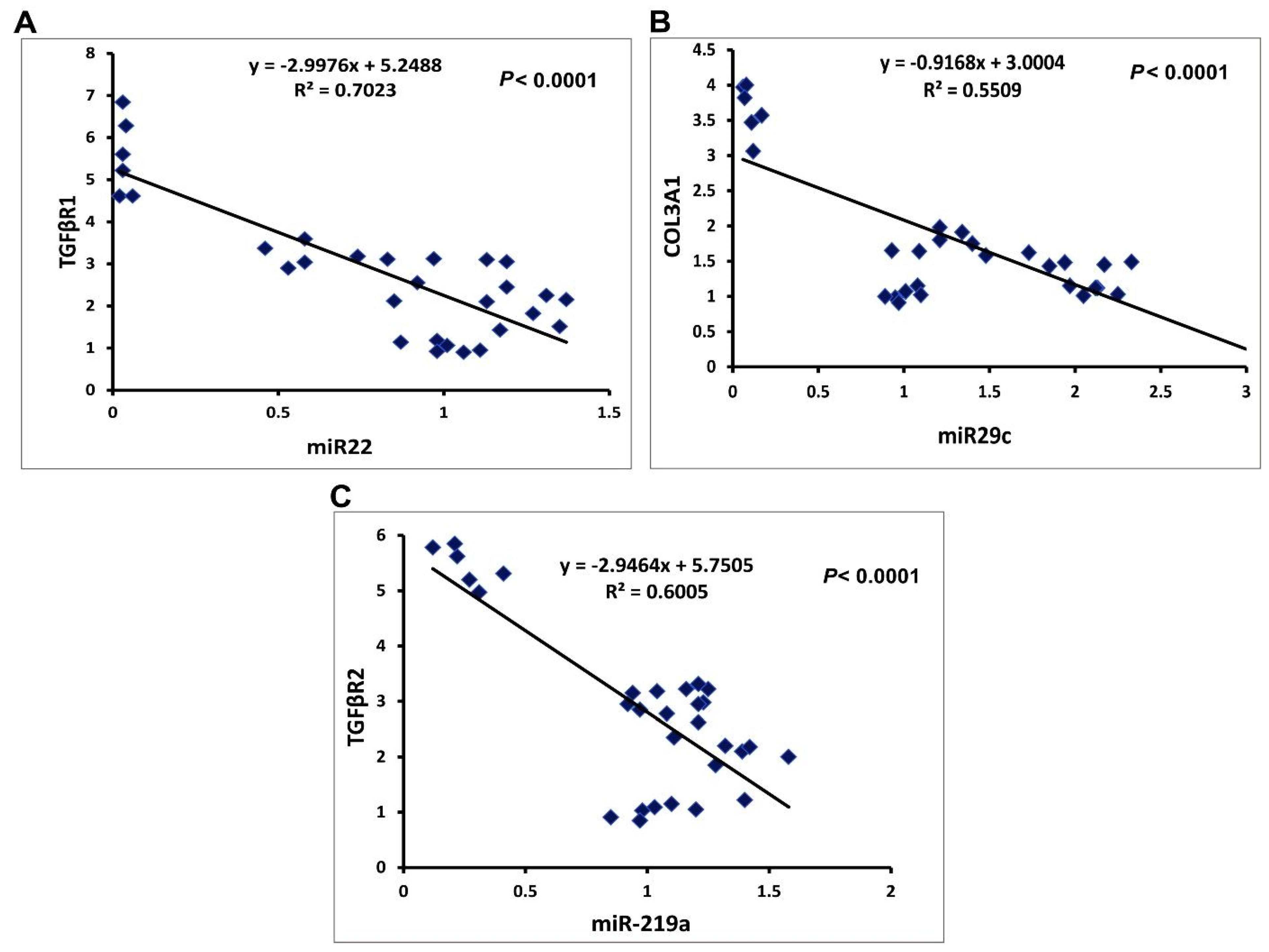
2.3.1. MicroRNAs Expression
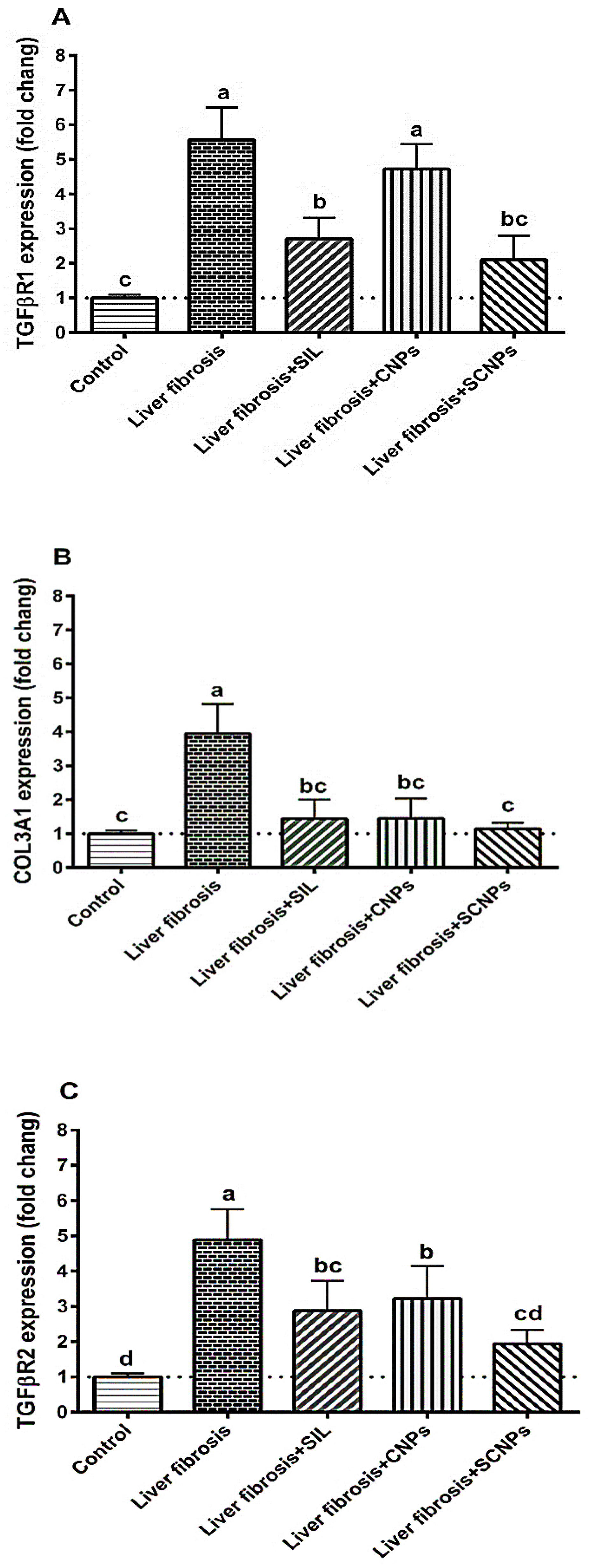
2.3.2. The Expression of the Target Genes
2.3.3. Correlation Studies
2.4. Histopathological Analysis
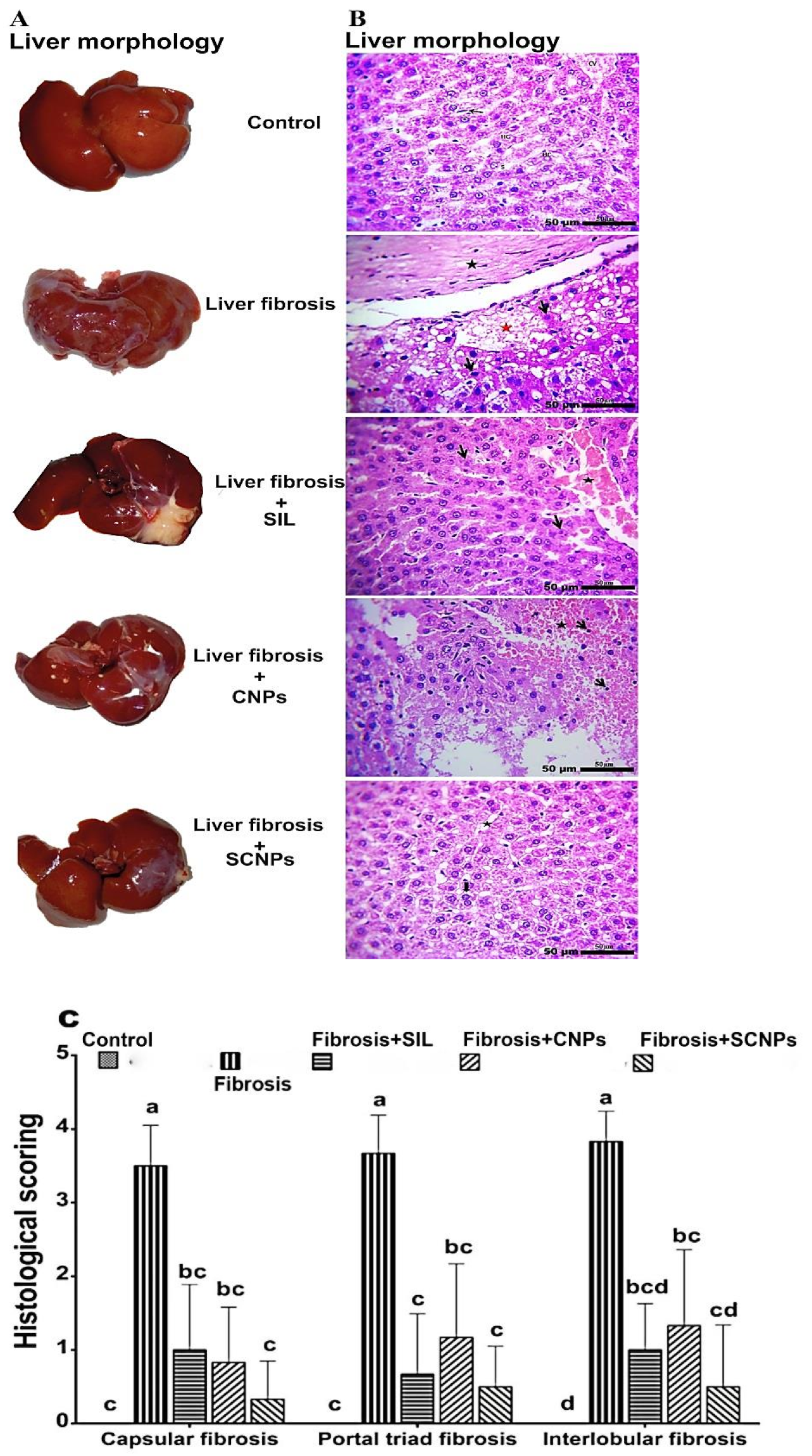
2.4.1. Liver Morphology
2.4.2. Histopathology of Liver Tissue
2.4.3. Histological Grading of Fibrosis
2.4.4. Masson’s Trichrome (MT) Staining of Liver Tissue
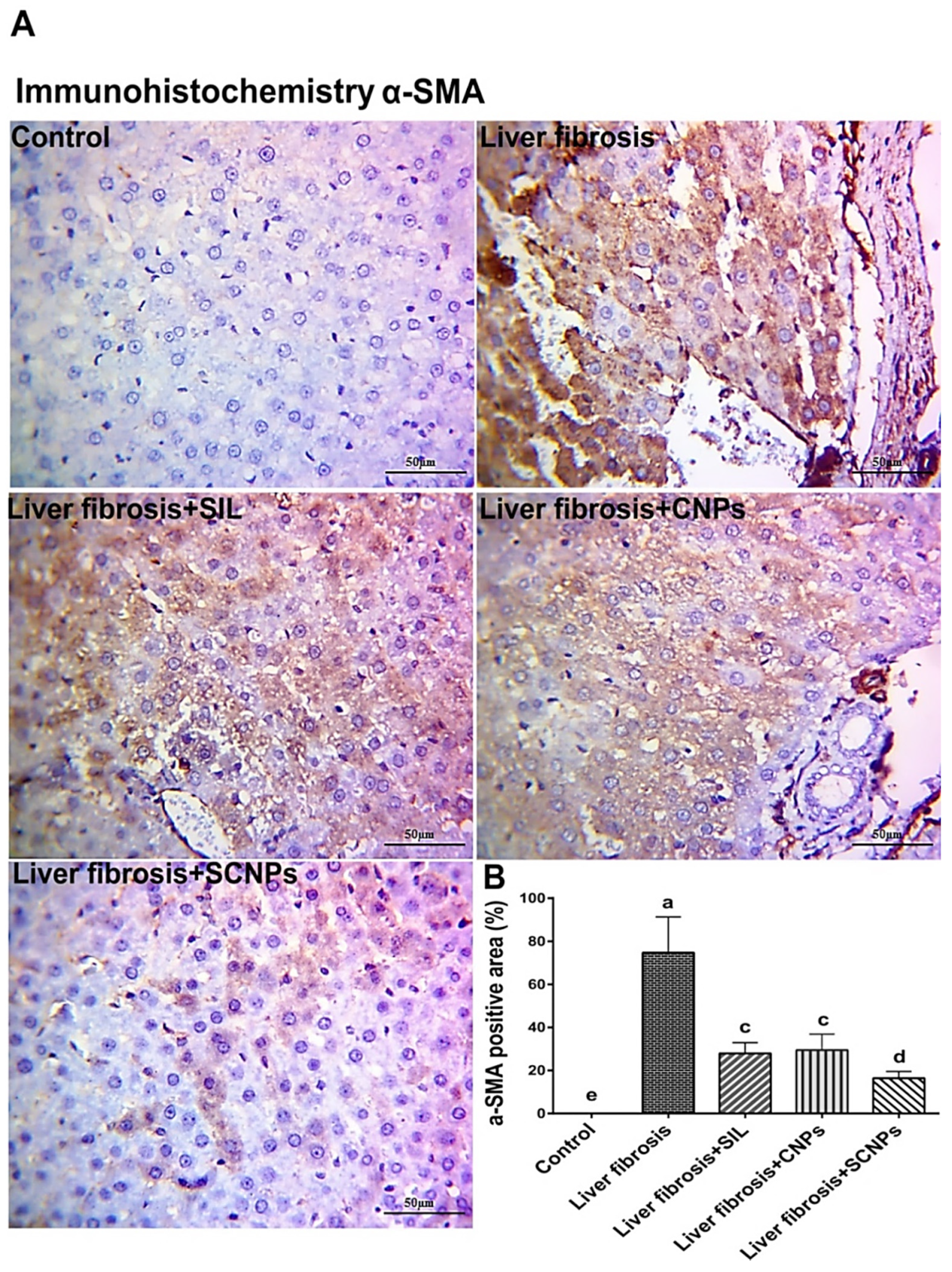
2.4.5. Immunohistochemistry (IHC) of αSMA
3. Discussion
4. Materials and Methods
4.1. Chemicals and Regents
4.2. Preparation and Characterization of Nanoparticles
4.2.1. Preparation of Chitosan Nanoparticles
4.2.2. Synthesis of Silymarin–Chitosan Nanoparticles
4.3. Physicochemical Characterizations of Nanoparticles
4.3.1. Dynamic Light Scattering and Zeta Potential Analysis
4.3.2. Transmission Electron Microscopy Analysis
4.3.3. Fourier Transforms Infrared Spectroscopy Study
4.3.4. Drug Entrapment Efficiency and Loading Capacity
4.3.5. In Vitro Drug Release Study
4.4. In Vivo Study
4.4.1. Animals
4.4.2. Ethical Statement
4.4.3. Establishment of Liver Fibrosis Model
4.4.4. Experimental Design
4.4.5. Sample Collection and Tissue Preparation
4.5. Methods
4.5.1. Serum Biomarkers for Liver Function Tests
4.5.2. Malondialdehyde (MDA) as Index of Lipid Peroxidation
4.5.3. ELISA Measurements
4.5.4. Gene Expression Analysis
Assessment of Hepatic MicroRNAs (miRNA-22, miRNA-29c, and miRNA-219a)
Assessment of Hepatic Expression of TGFβR1, TGFβR2, and COL3A1 Genes
Relative Quantification of Gene Expression
4.6. Histopathology Study
4.7. Immunohistochemistry (IHC) Study
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- GBD 2017 Cirrhosis Collaborators. The global, regional, and national burden of cirrhosis by cause in 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2020, 5, 245–266. [Google Scholar]
- Aydın, M.M.; Akçalı, K.C. Liver fibrosis. Turk. J. Gastroenterol. 2018, 29, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.-S.; Kim, W.R. The global impact of hepatic fibrosis and end-stage liver disease. Clin. Liver Dis. 2008, 12, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Geervliet, E.; Bansal, R. Matrix metalloproteinases as potential biomarkers and therapeutic targets in liver diseases. Cells 2020, 9, 1212. [Google Scholar] [CrossRef] [PubMed]
- Noyan, S.; Çavuşoğlu, İ.; Minbay, F.Z. The effect of vitamin A on CCl4-induced hepatic injuries in rats: A histochemical, immunohistochemical and ultrastructural study. Acta Histochem. 2006, 107, 421–434. [Google Scholar] [CrossRef]
- Jiao, J.; Friedman, S.L.; Aloman, C. Hepatic fibrosis. Curr. Opin. Gastroenterol. 2009, 25, 223. [Google Scholar] [CrossRef]
- Arfmann-Knübel, S.; Struck, B.; Genrich, G.; Helm, O.; Sipos, B.; Sebens, S.; Schäfer, H. The crosstalk between Nrf2 and TGF-β1 in the epithelial-mesenchymal transition of pancreatic duct epithelial cells. PLoS ONE 2015, 10, e0132978. [Google Scholar] [CrossRef]
- Choi, H.-K.; Pokharel, Y.R.; Lim, S.C.; Han, H.-K.; Ryu, C.S.; Kim, S.K.; Kwak, M.K.; Kang, K.W. Inhibition of liver fibrosis by solubilized coenzyme Q10: Role of Nrf2 activation in inhibiting transforming growth factor-β1 expression. Toxicol. Appl. Pharmacol. 2009, 240, 377–384. [Google Scholar] [CrossRef]
- O’Reilly, S. MicroRNAs in fibrosis: Opportunities and challenges. Arthritis Res. Ther. 2016, 18, 11. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.-P.; Ai, W.-B.; Wan, L.-Y.; Zhang, Y.-Q.; Wu, J.-F. The roles of microRNA families in hepatic fibrosis. Cell Biosci. 2017, 7, 34. [Google Scholar] [CrossRef] [Green Version]
- Hobert, O. Gene regulation by transcription factors and microRNAs. Science 2008, 319, 1785–1786. [Google Scholar] [CrossRef] [PubMed]
- Ji, D.; Li, B.; Shao, Q.; Li, F.; Li, Z.; Chen, G. Mir-22 suppresses bmp7 in the development of cirrhosis. Cell. Physiol. Biochem. 2015, 36, 1026–1036. [Google Scholar] [CrossRef] [PubMed]
- Roderburg, C.; Urban, G.W.; Bettermann, K.; Vucur, M.; Zimmermann, H.; Schmidt, S.; Janssen, J.; Koppe, C.; Knolle, P.; Castoldi, M. Micro-RNA profiling reveals a role for miR-29 in human and murine liver fibrosis. Hepatology 2011, 53, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Ma, J.; Ou, H. MicroRNA-219 overexpression serves a protective role during liver fibrosis by targeting tumor growth factor β receptor 2. Mol. Med. Rep. 2019, 19, 1543–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Zhang, Q.; Wang, B.; Wu, W.; Wei, J.; Li, P.; Huang, R. miR-22 regulates C2C12 myoblast proliferation and differentiation by targeting TGFBR1. Eur. J. Cell Biol. 2018, 97, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.-D.; Khorram, O. Mechanisms underlying aberrant expression of miR-29c in uterine leiomyoma. Fertil. Steril. 2016, 105, 236–245.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdullah, A.S.; El Sayed, I.E.T.; El-Torgoman, A.M.A.; Alghamdi, N.A.; Ullah, S.; Wageh, S.; Kamel, M.A. Preparation and Characterization of Silymarin-Conjugated Gold Nanoparticles with Enhanced Anti-Fibrotic Therapeutic Effects against Hepatic Fibrosis in Rats: Role of MicroRNAs as Molecular Targets. Biomedicines 2021, 9, 1767. [Google Scholar] [CrossRef]
- Woo, J.S.; Kim, T.-S.; Park, J.-H.; Chi, S.-C. Formulation and biopharmaceutical evaluation of silymarin using SMEDDS. Arch. Pharmacal Res. 2007, 30, 82–89. [Google Scholar] [CrossRef]
- Clichici, S.; David, L.; Moldovan, B.; Baldea, I.; Olteanu, D.; Filip, M.; Nagy, A.; Luca, V.; Crivii, C.; Mircea, P.; et al. Hepatoprotective effects of silymarin coated gold nanoparticles in experimental cholestasis. Mater. Sci. Eng. C 2020, 115, 111117. [Google Scholar] [CrossRef]
- Ma, Y.; He, H.; Xia, F.; Li, Y.; Lu, Y.; Chen, D.; Qi, J.; Lu, Y.; Zhang, W.; Wu, W. In vivo fate of lipid-silybin conjugate nanoparticles: Implications on enhanced oral bioavailability. Nanomedicine 2017, 13, 2643–2654. [Google Scholar] [CrossRef]
- Pathomthongtaweechai, N.; Muanprasat, C. Potential Applications of Chitosan-Based Nanomaterials to Surpass the Gastrointestinal Physiological Obstacles and Enhance the Intestinal Drug Absorption. Pharmaceutics 2021, 13, 887. [Google Scholar] [CrossRef] [PubMed]
- Ragelle, H.; Vandermeulen, G.; Préat, V. Chitosan-based siRNA delivery systems. J. Control. Release 2013, 172, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahhab, M.A.; Aljawish, A.; El-Nekeety, A.A.; Abdel-Aiezm, S.H.; Abdel-Kader, H.A.; Rihn, B.H.; Joubert, O. Chitosan nanoparticles and quercetin modulate gene expression and prevent the genotoxicity of aflatoxin B1 in rat liver. Toxicol. Rep. 2015, 2, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Chesnutt, B.M.; Haggard, W.O.; Bumgardner, J.D. Deacetylation of chitosan: Material characterization and in vitro evaluation via albumin adsorption and pre-osteoblastic cell cultures. Materials 2011, 4, 1399–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammed, M.A.; Syeda, J.T.M.; Wasan, K.M.; Wasan, E.K. An overview of chitosan nanoparticles and its application in non-parenteral drug delivery. Pharmaceutics 2017, 9, 53. [Google Scholar] [CrossRef] [Green Version]
- Mikušová, V.; Mikuš, P. Advances in chitosan-based nanoparticles for drug delivery. Int. J. Mol. Sci. 2021, 22, 9652. [Google Scholar] [CrossRef]
- Lang, X.; Wang, T.; Sun, M.; Chen, X.; Liu, Y. Advances and applications of chitosan-based nanomaterials as oral delivery carriers: A review. Int. J. Biol. Macromol. 2020, 154, 433–445. [Google Scholar] [CrossRef]
- Bernkop-Schnürch, A.; Dünnhaupt, S. Chitosan-based drug delivery systems. Eur. J. Pharm. Biopharm. 2012, 81, 463–469. [Google Scholar] [CrossRef]
- Ways, T.M.M.; Lau, W.M.; Khutoryanskiy, V.V. Chitosan and its derivatives for application in mucoadhesive drug delivery systems. Polymers 2018, 10, 267. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Vimal, A.; Kumar, A. Why Chitosan? From properties to perspective of mucosal drug delivery. Int. J. Biol. Macromol. 2016, 91, 615–622. [Google Scholar] [CrossRef]
- Ponnuraj, R.; Janakiraman, K.; Gopalakrishnan, S.; Senthilnathan, K.; Meganathan, V.; Saravanan, P. Formulation and characterization of epigallocatechin gallate nanoparticles. Indo. Am. J. Pharm. Res. 2015, 5, 387–399. [Google Scholar]
- Liang, J.; Yan, H.; Puligundla, P.; Gao, X.; Zhou, Y.; Wan, X. Applications of chitosan nanoparticles to enhance absorption and bioavailability of tea polyphenols: A review. Food Hydrocoll. 2017, 69, 286–292. [Google Scholar] [CrossRef]
- Safer, A.-M.; Leporatti, S.; Jose, J.; Soliman, M.S. Conjugation of EGCG and chitosan NPS as a novel nano-drug delivery system. Int. J. Nanomed. 2019, 14, 8033. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, N.; Ahmad, R.; Naqvi, A.A.; Alam, M.A.; Ashafaq, M.; Samim, M.; Iqbal, Z.; Ahmad, F.J. Rutin-encapsulated chitosan nanoparticles targeted to the brain in the treatment of Cerebral Ischemia. Int. J. Biol. Macromol. 2016, 91, 640–655. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.-S. Nanoparticles of biodegradable polymers for new-concept chemotherapy. Expert Rev. Med. Devices 2004, 1, 115–125. [Google Scholar] [CrossRef]
- Li, F.-Q.; Su, H.; Chen, X.; Qin, X.-J.; Liu, J.-Y.; Zhu, Q.-G.; Hu, J.-H. Mannose 6-phosphate-modified bovine serum albumin nanoparticles for controlled and targeted delivery of sodium ferulate for treatment of hepatic fibrosis. J. Pharm. Pharmacol. 2009, 61, 1155–1161. [Google Scholar] [CrossRef]
- Alipour, M.; Reza Bigdeli, M.; Aligholi, H.; Rasoulian, B.; Khaksarian, M. Sustained release of silibinin-loaded chitosan nanoparticle induced apoptosis in glioma cells. J. Biomed. Mater. Res. Part A 2020, 108, 458–469. [Google Scholar] [CrossRef]
- Gohulkumar, M.; Gurushankar, K.; Prasad, N.R.; Krishnakumar, N. Enhanced cytotoxicity and apoptosis-induced anticancer effect of silibinin-loaded nanoparticles in oral carcinoma (KB) cells. Mater. Sci. Eng. C 2014, 41, 274–282. [Google Scholar] [CrossRef]
- Pooja, D.; Bikkina, D.J.B.; Kulhari, H.; Nikhila, N.; Chinde, S.; Raghavendra, Y.; Sreedhar, B.; Tiwari, A.K. Fabrication, characterization and bioevaluation of silibinin loaded chitosan nanoparticles. Int. J. Biol. Macromol. 2014, 69, 267–273. [Google Scholar] [CrossRef]
- Kumar, S.; Randhawa, J.K. Preparation and characterization of Paliperidone loaded solid lipid nanoparticles. Colloids Surf. B Biointerfaces 2013, 102, 562–568. [Google Scholar] [CrossRef]
- Bhattacharjee, S. DLS and zeta potential—What they are and what they are not? J. Control Release 2016, 235, 337–351. [Google Scholar] [CrossRef] [PubMed]
- Netto, G.; Jose, J. Development, characterization, and evaluation of sunscreen cream containing solid lipid nanoparticles of silymarin. J. Cosmet. Dermatol. 2018, 17, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.F.; Zhu, Y.L.; Sun, B.; Hu, F.H.; Tian, T.; Li, S.C.; Xiao, Z.D. PLGA-based gene delivering nanoparticle enhance suppression effect of miRNA in HePG2 cells. Nanoscale Res. Lett. 2011, 6, 447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Wang, H.; Yu, M.; Liao, Z.; Wang, X.; Zhang, F.; Ji, W.; Wu, B.; Han, J.; Zhang, H.; et al. Paclitaxel loaded folic acid targeted nanoparticles of mixed lipid-shell and polymer-core: In vitro and in vivo evaluation. Eur. J. Pharm. Biopharm. 2012, 81, 248–256. [Google Scholar] [CrossRef]
- Moghaddam, A.H.; Mokhtari Sangdehi, S.R.; Ranjbar, M.; Hasantabar, V. Preventive effect of silymarin-loaded chitosan nanoparticles against global cerebral ischemia/reperfusion injury in rats. Eur. J. Pharmacol. 2020, 877, 173066. [Google Scholar] [CrossRef]
- Jana, U.; Mohanty, A.K.; Manna, P.K.; Mohanta, G.P. Preparation and characterization of nebivolol nanoparticles using Eudragit® RS100. Colloids Surf. B Biointerfaces 2014, 113, 269–275. [Google Scholar] [CrossRef]
- Leena, R.; Vairamani, M.; Selvamurugan, N. Alginate/Gelatin scaffolds incorporated with Silibinin-loaded Chitosan nanoparticles for bone formation in vitro. Colloids Surf. B Biointerfaces 2017, 158, 308–318. [Google Scholar] [CrossRef]
- Radu, I.-C.; Hudita, A.; Zaharia, C.; Stanescu, P.O.; Vasile, E.; Iovu, H.; Stan, M.; Ginghina, O.; Galateanu, B.; Costache, M.; et al. Poly (hydroxybutyrate-co-hydroxyvalerate)(PHBHV) nanocarriers for silymarin release as adjuvant therapy in colo-rectal cancer. Front. Pharmacol. 2017, 8, 508. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yang, X.; Yang, J.; Wang, Y.; Chen, R.; Wu, J.; Liu, Y.; Zhang, N. Self-assembled nanoparticles of methotrexate conjugated O-carboxymethyl chitosan: Preparation, characterization and drug release behavior in vitro. Carbohydr. Polym. 2011, 86, 1665–1670. [Google Scholar] [CrossRef]
- Aboshanab, M.H.; El-Nabarawi, M.A.; Teaima, M.H.; El-Nekeety, A.A.; Abdel-Aziem, S.H.; Hassan, N.S.; Abdel-Wahhab, M.A. Fabrication, characterization and biological evaluation of silymarin nanoparticles against carbon tetrachloride-induced oxidative stress and genotoxicity in rats. Int. J. Pharm. 2020, 587, 119639. [Google Scholar] [CrossRef]
- Javed, S.; Kohli, K.; Ali, M. Reassessing bioavailability of silymarin. Altern. Med. Rev. 2011, 16, 239. [Google Scholar] [PubMed]
- Igartua, M.; Hernandez, R.M.; Esquisabel, A.; Gascon, A.R.; Calvo, M.B.; Pedraz, J.J. Influence of formulation variables on the in-vitro release of albumin from biodegradable microparticulate systems. J. Microencapsul. 1997, 14, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Rafati, H.; Coombes, A.; Adler, J.; Holland, J.; Davis, S.S. Protein-loaded poly (DL-lactide-co-glycolide) microparticles for oral administration: Formulation, structural and release characteristics. J. Control. Release 1997, 43, 89–102. [Google Scholar] [CrossRef]
- Gupta, S.; Singh, S.K.; Girotra, P. Targeting silymarin for improved hepatoprotective activity through chitosan nanoparticles. Int. J. Pharm. Investig. 2014, 4, 156. [Google Scholar]
- Bodmeier, R.; McGinity, J. Solvent selection in the preparation of poly (DL-lactide) microspheres prepared by the solvent evaporation method. Int. J. Pharm. 1988, 43, 179–186. [Google Scholar] [CrossRef]
- Hsu, Y.-J.; Wang, C.-Y.; Lee, M.-C.; Huang, C.-C. Hepatoprotection by Traditional Essence of Ginseng against Carbon Tetrachloride—Induced Liver Damage. Nutrients 2020, 12, 3214. [Google Scholar] [CrossRef]
- Kabir, N.; Ali, H.; Ateeq, M.; Bertino, M.F.; Shah, M.R.; Franzel, L. Silymarin coated gold nanoparticles ameliorates CCl4-induced hepatic injury and cirrhosis through down regulation of hepatic stellate cells and attenuation of Kupffer cells. RSC Adv. 2014, 4, 9012–9020. [Google Scholar] [CrossRef]
- Omar, N.A.; Elbakrya, K.A.; Nasra, S.M.; Elemshatyb, H.M. Immunohistopathological Studies on Rats Injected with CCl4 and Treated with Propolis and Honey Bee. Egypt. J. Hosp. Med. 2015, 61, 425–436. [Google Scholar] [CrossRef]
- El-Demerdash, E.; Abdel-Sattar, S.A.; El-Bakly, W.M.; Mohamed, E.A. Antifibrotic effects of carvedilol and impact of liver fibrosis on carvedilol pharmacokinetics in a rat model. Eur. J. Drug Metab. Pharmacokinet. 2017, 42, 767–779. [Google Scholar] [CrossRef]
- Adhikari, A.; Polley, N.; Darbar, S.; Bagchi, D.; Pal, S.K. Citrate functionalized Mn3O4 in nanotherapy of hepatic fibrosis by oral administration. Future Sci. OA 2016, 2, FSO146. [Google Scholar] [CrossRef] [Green Version]
- Wasmuth, H.; Weiskirchen, R. Pathogenesis of liver fibrosis: Modulation of stellate cells by chemokines. Z. Gastroenterol. 2010, 48, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Chakraborty, A.K.; Dey, P.; Kar, P.; Guha, P.; Sen, S.; Kumar, A.; Sen, A.; Chaudhuri, T.K. Amelioration of CCl4 induced liver injury in swiss albino mice by antioxidant rich leaf extract of Croton bonplandianus Baill. PLoS ONE 2018, 13, e0196411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naz, I.; Khan, M.R.; Zai, J.A.; Batool, R.; Zahra, Z.; Tahir, A. Pilea umbrosa ameliorate CCl4 induced hepatic injuries by regulating endoplasmic reticulum stress, pro-inflammatory and fibrosis genes in rat. Environ. Health Prev. Med. 2020, 25, 53. [Google Scholar] [CrossRef] [PubMed]
- Devaraj, E.; Roy, A.; Veeraragavan, G.R.; Magesh, A.; Sleeba, A.V.; Arivarasu, L.; Parasuraman, B.M. β-Sitosterol attenuates carbon tetrachloride–induced oxidative stress and chronic liver injury in rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Elsawy, H.; Badr, G.M.; Sedky, A.; Abdallah, B.M.; Alzahrani, A.M.; Abdel-Moneim, A.M. Rutin ameliorates carbon tetrachloride (CCl4)-induced hepatorenal toxicity and hypogonadism in male rats. PeerJ 2019, 7, e7011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Zhang, H.; Cao, Y.; Li, Y.; Sun, S.; Zhang, J.; Zhang, G. Schisandrin B attenuates CCl4-induced liver fibrosis in rats by regulation of Nrf2-ARE and TGF-β/Smad signaling pathways. Drug Des. Dev. Ther. 2017, 11, 2179. [Google Scholar] [CrossRef] [Green Version]
- Köhler, U.A.; Böhm, F.; Rolfs, F.; Egger, M.; Hornemann, T.; Pasparakis, M.; Weber, A.; Werner, S. NF-κB/RelA and Nrf2 cooperate to maintain hepatocyte integrity and to prevent development of hepatocellular adenoma. J. Hepatol. 2016, 64, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Shimozono, R.; Asaoka, Y.; Yoshizawa, Y.; Aoki, T.; Noda, H.; Yamada, M.; Kaino, M.; Mochizuki, H. Nrf2 activators attenuate the progression of nonalcoholic steatohepatitis–related fibrosis in a dietary rat model. Mol. Pharmacol. 2013, 84, 62–70. [Google Scholar] [CrossRef]
- Niu, L.; Cui, X.; Qi, Y.; Xie, D.; Wu, Q.; Chen, X.; Ge, J.; Liu, Z. Involvement of TGF-β1/Smad3 signaling in carbon tetrachloride-induced acute liver injury in mice. PLoS ONE 2016, 11, e0156090. [Google Scholar] [CrossRef]
- Supriono, S.; Nugraheni, A.; Kalim, H.; Eko, M.H. The Effect of Curcumin on Regression of Liver Fibrosis through Decreased Expression of Transforming Growth Factor-β1 (TGF-β1). Indones. Biomed. J. 2019, 11, 52–58. [Google Scholar] [CrossRef]
- Mohseni, R.; Karimi, J.; Tavilani, H.; Khodadadi, I.; Hashemnia, M. Carvacrol ameliorates the progression of liver fibrosis through targeting of Hippo and TGF-β signaling pathways in carbon tetrachloride (CCl4)-induced liver fibrosis in rats. Immunopharmacol. Immunotoxicol. 2019, 41, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, J.; Ma, Y.; Wang, J.; Zhu, J.; Liu, J.; Zhang, J. Integration of high-throughput data of microRNA and mRNA expression profiles reveals novel insights into the mechanism of liver fibrosis. Mol. Med. Rep. 2019, 19, 115–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gjorgjieva, M.; Sobolewski, C.; Ay, A.-S.; Abegg, D.; Correia de Sousa, M.; Portius, D.; Berthou, F.; Fournier, M.; Maeder, C.; Rantakari, P. Genetic ablation of MiR-22 fosters diet-induced obesity and NAFLD development. J. Pers. Med. 2020, 10, 170. [Google Scholar] [CrossRef]
- Roulot, D.; Sevcsik, A.M.; Coste, T.; Strosberg, A.D.; Marullo, S. Role of transforming growth factor β type II receptor in hepatic fibrosis: Studies of human chronic hepatitis C and experimental fibrosis in rats. Hepatology 1999, 29, 1730–1738. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Cao, H.; Wang, Q.; Ye, J.; Sui, L.; Feng, J.; Cai, X.; Song, H.; Zhang, X.; Chen, X. MiR-22 may suppress fibrogenesis by targeting TGFβR I in cardiac fibroblasts. Cell. Physiol. Biochem. 2016, 40, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, T.; Hamada, Y.; Yamada, H.; Horii, I. Changes of micro-RNA expression in rat liver treated by acetaminophen or carbon tetrachloride− regulating role of micro-rna for RNA expression. J. Toxicol. Sci. 2007, 32, 401–409. [Google Scholar] [CrossRef] [Green Version]
- Robinson, T.F.; Cohen-Gould, L.; Factor, S.M.; Eghbali, M.; Blumenfeld, O.O. Structure and function of connective tissue in cardiac muscle: Collagen types I and III in endomysial struts and pericellular fibers. Scanning Microsc. 1988, 2, 1005–1015. [Google Scholar]
- Hafez, M.M.; Hamed, S.S.; El-Khadragy, M.F.; Hassan, Z.K.; Al Rejaie, S.S.; Sayed-Ahmed, M.M.; Al-Harbi, N.O.; Al-Hosaini, K.A.; Al-Harbi, M.M.; Alhoshani, A.R.; et al. Effect of ginseng extract on the TGF-β1 signaling pathway in CCl4-induced liver fibrosis in rats. BMC Complement. Altern. Med. 2017, 17, 45. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Wang, Q.; Wang, Q.; Zhao, X.; Zhao, P.; Geng, T.; Gong, D. Role of miR29c in goose fatty liver is mediated by its target genes that are involved in energy homeostasis and cell growth. BMC Vet. Res. 2018, 14, 325. [Google Scholar] [CrossRef] [Green Version]
- Bansal, M.B.; Chamroonkul, N. Antifibrotics in liver disease: Are we getting closer to clinical use? Hepatol. Int. 2019, 13, 25–39. [Google Scholar] [CrossRef]
- Ebrahimi, H.; Naderian, M.; Sohrabpour, A.A. New concepts on reversibility and targeting of liver fibrosis; A review article. Middle East J. Dig. Dis. 2018, 10, 133. [Google Scholar] [CrossRef] [PubMed]
- Gillessen, A.; Schmidt, H.H.-J. Silymarin as supportive treatment in liver diseases: A narrative review. Adv. Ther. 2020, 37, 1279–1301. [Google Scholar] [CrossRef] [Green Version]
- De Avelar, C.R.; Pereira, E.M.; De Farias Costa, P.R.; De Jesus, R.P.; De Oliveira, L.P. Effect of silymarin on biochemical indicators in patients with liver disease: Systematic review with meta-analysis. World J. Gastroenterol. 2017, 23, 5004. [Google Scholar] [CrossRef]
- Abenavoli, L.; Aviello, G.; Capasso, R.; Milic, N.; Capasso, F. Milk thistle for treatment of nonalcoholic fatty liver disease. Hepat. Mon. 2011, 11, 173–177. [Google Scholar]
- Pérez-Sánchez, A.; Cuyàs, E.; Ruiz-Torres, V.; Agulló-Chazarra, L.; Verdura, S.; González-Álvarez, I.; Bermejo, M.; Joven, J.; Micol, V.; Bosch-Barrera, J.; et al. Intestinal permeability study of clinically relevant formulations of silibinin in Caco-2 cell monolayers. Int. J. Mol. Sci. 2019, 20, 1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saller, R.; Meier, R.; Brignoli, R. The use of silymarin in the treatment of liver diseases. Drugs 2001, 61, 2035–2063. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Manautou, J.E. Molecular mechanisms underlying chemical liver injury. Expert Rev. Mol. Med. 2012, 14, e4. [Google Scholar] [CrossRef] [Green Version]
- Bonferoni, M.C.; Gavini, E.; Rassu, G.; Maestri, M.; Giunchedi, P. Chitosan Nanoparticles for Therapy and Theranostics of Hepatocellular Carcinoma (HCC) and Liver-Targeting. Nanomaterials 2020, 10, 870. [Google Scholar] [CrossRef]
- Jeon, T.I.; Hwang, S.G.; Park, N.G.; Jung, Y.R.; Im Shin, S.; Choi, S.D.; Park, D.K. Antioxidative effect of chitosan on chronic carbon tetrachloride induced hepatic injury in rats. Toxicology 2003, 187, 67–73. [Google Scholar] [CrossRef]
- Teksoy, O.; Sahinturk, V.; Cengiz, M.; İnal, B.; Ayhancı, A. The Protective Effects of Silymarin on Thioacetamide-Induced Liver Damage: Measurement of miR-122, miR-192, and miR-194 Levels. Appl. Biochem. Biotechnol. 2019, 191, 528–539. [Google Scholar] [CrossRef]
- Kim, M.; Yang, S.-G.; Kim, J.M.; Lee, J.-W.; Kim, Y.S.; Lee, J.I. Silymarin suppresses hepatic stellate cell activation in a dietary rat model of non-alcoholic steatohepatitis: Analysis of isolated hepatic stellate cells. Int. J. Mol. Med. 2012, 30, 473–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, S.; Yang, F.; Wang, Y.; Qin, Y.; Xian, H.; Che, H.; Wang, L. Silymarin ameliorates diabetic cardiomyopathy via inhibiting TGF-β1/Smad signaling. Cell Biol. Int. 2019, 43, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Lu, H.; Yang, H. Chitosan inhibits fibroblasts growth in Achilles tendon via TGF-β1/Smad3 pathway by miR-29b. Int. J. Clin. Exp. Pathol. 2014, 7, 8462. [Google Scholar] [PubMed]
- Potdar, P.D.; Shetti, A.U. Evaluation of anti-metastatic effect of chitosan nanoparticles on esophageal cancer-associated fibroblasts. J. Cancer Metastasis Treat. 2016, 2, 259–267. [Google Scholar] [CrossRef]
- Sooklert, K.; Nilyai, S.; Rojanathanes, R.; Jindatip, D.; Sae-Liang, N.; Kitkumthorn, N.; Mutirangura, A.; Sereemaspun, A. N-Acetylcysteine reverses the decrease of DNA methylation status caused by engineered gold, silicon, and chitosan nanoparticles. Int. J. Nanomed. 2019, 14, 4573. [Google Scholar] [CrossRef] [Green Version]
- Calvo, P.; Remuñán-López, C.; Vila-Jato, J.L.; Alonso, M.J. Novel hydrophilic chitosan-polyethylene oxide nanoparticles as protein carriers. J. Appl. Polym. Sci. 1997, 63, 125–132. [Google Scholar] [CrossRef]
- Mitra, A.; Dey, B. Chitosan microspheres in novel drug delivery systems. Indian J. Pharm. Sci. 2011, 73, 355. [Google Scholar]
- Sipoli, C.C.; Santana, N.; Shimojo, A.A.M.; Azzoni, A.; de la Torre, L.G. Scalable production of highly concentrated chitosan/TPP nanoparticles in different pHs and evaluation of the in vitro transfection efficiency. Biochem. Eng. J. 2015, 94, 65–73. [Google Scholar] [CrossRef]
- Othman, N.; Masarudin, M.J.; Kuen, C.Y.; Dasuan, N.A.; Abdullah, L.C.; Md. Jamil, S.N.A. Synthesis and optimization of chitosan nanoparticles loaded with L-ascorbic acid and thymoquinone. Nanomaterials 2018, 8, 920. [Google Scholar] [CrossRef] [Green Version]
- Nagpal, K.; Singh, S.K.; Mishra, D.N. Optimization of brain targeted gallic acid nanoparticles for improved antianxiety-like activity. Int. J. Biol. Macromol. 2013, 57, 83–91. [Google Scholar] [CrossRef]
- Reddy, A.S.; Lakshmi, B.A.; Kim, S.; Kim, J. Synthesis and characterization of acetyl curcumin-loaded core/shell liposome nanoparticles via an electrospray process for drug delivery, and theranostic applications. Eur. J. Pharm. Biopharm. 2019, 142, 518–530. [Google Scholar] [CrossRef] [PubMed]
- Laouar, A.; Klibet, F.; Bourogaa, E.; Benamara, A.; Boumendjel, A.; Chefrour, A.; Messarah, M. Potential antioxidant properties and hepatoprotective effects of Juniperus phoenicea berries against CCl4 induced hepatic damage in rats. Asian Pac. J. Trop. Med. 2017, 10, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Maksoud, H.A.; Magid, A.D.A.; Mostafa, Y.; Elharrif, M.G.; Sorour, R.I.; Sorour, M.I. Ameliorative effect of liquorice extract versus silymarin in experimentally induced chronic hepatitis: A biochemical and genetical study. Clin. Nutr. Exp. 2019, 23, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Draper, H.; Hadley, M. [43] Malondialdehyde determination as index of lipid Peroxidation. Methods Enzymol. 1990, 186, 421–431. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Wu, Y.C.; Huang, H.H.; Wu, Y.J.; Manousakas, I.; Yang, C.C.; Kuo, S.M. Therapeutic and Protective Effects of Liposomal Encapsulation of Astaxanthin in Mice with Alcoholic Liver Fibrosis. Int. J. Mol. Sci. 2019, 20, 4057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Luo, J.; Li, L.; Cheng, M.; Huang, N.; Liu, J.; Waalkes, M.P. The collagenolytic effects of the traditional Chinese medicine preparation, Han-Dan-Gan-Le, contribute to reversal of chemical-induced liver fibrosis in rats. Life Sci. 2003, 72, 1563–1571. [Google Scholar] [CrossRef]
- Hsu, S.-M.; Raine, L.; Fanger, H. A comparative study of the peroxidase-antiperoxidase method and an avidin-biotin complex method for studying polypeptide hormones with radioimmunoassay antibodies. Am. J. Clin. Pathol. 1981, 75, 734–738. [Google Scholar] [CrossRef]
- Gabius, H.-J.; Gabius, S. Lectins and Glycobiology; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Parameters | ||||
|---|---|---|---|---|---|
| AST (U/L) | ALT (U/L) | ALP (U/L) | Total Bilirubin (mg/dL) | Albumin (g/dL) | |
| Control | 4.13 a ± 0.21 | 0.41 c ± 0.10 | 103.5 d ± 13.79 | 44.34 d ± 7.42 | 112.6 d ± 12.44 |
| Liver fibrosis | 3.12 c ± 0.13 | 1.16 a ± 0.30 | 253.3 a ± 19.62 | 131.7 a ± 13.70 | 196.4 a ± 17.76 |
| Liver fibrosis + SIL | 3.48 b,c ± 0.24 | 0.74 b ± 0.21 | 147.9 c ± 16.72 | 62.19 c ± 9.02 | 133.9 b–d ± 12.75 |
| Liver fibrosis + CNPs | 3.30 b,c ± 0.21 | 0.83 b ± 0.12 | 152.0 c ± 14.06 | 68.67 c ± 5.79 | 149.8 b,c ± 19.07 |
| Liver fibrosis + SCNPs | 3.62 b ± 0.08 | 0.61 b,c ± 0.10 | 137.7 c ± 14.54 | 53.3 c,d ± 6.62 | 129.7 c,d ± 19.87 |
| Gene | Sequence | |
|---|---|---|
| COL3A1 | Forward | 5′-AAC GGA GCT CCT GGC CCC AT-3′ |
| Reverse | 5′-ATT GCC TCG AGC ACC TGC GG-3′ | |
| TGFβR1 | Forward | 5′-GCT GAC ATC TAT GCA ATG GG-3′ |
| Reverse | 5′-ATA TTT GGC CTT AAC TTC TGT TC-3′ | |
| TGFβR2 | Forward | 5′-CCA GGG CAT CCA GAT CGT GTG-3′ |
| Reverse | 5′-TAG TGT TCA GGG AGC CGT CTT-3′ | |
| GAPDH | Forward | 5′-GGG TGT GAA CCA CGA GAA ATA-3′ |
| Reverse | 5′-AGT TGT CAT GGA TGA CCT T-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdullah, A.S.; Sayed, I.E.T.E.; El-Torgoman, A.M.A.; Kalam, A.; Wageh, S.; Kamel, M.A. Green Synthesis of Silymarin–Chitosan Nanoparticles as a New Nano Formulation with Enhanced Anti-Fibrotic Effects against Liver Fibrosis. Int. J. Mol. Sci. 2022, 23, 5420. https://doi.org/10.3390/ijms23105420
Abdullah AS, Sayed IETE, El-Torgoman AMA, Kalam A, Wageh S, Kamel MA. Green Synthesis of Silymarin–Chitosan Nanoparticles as a New Nano Formulation with Enhanced Anti-Fibrotic Effects against Liver Fibrosis. International Journal of Molecular Sciences. 2022; 23(10):5420. https://doi.org/10.3390/ijms23105420
Chicago/Turabian StyleAbdullah, Abdullah Saad, Ibrahim El Tantawy El Sayed, Abdel Moneim A. El-Torgoman, Abul Kalam, S. Wageh, and Maher A. Kamel. 2022. "Green Synthesis of Silymarin–Chitosan Nanoparticles as a New Nano Formulation with Enhanced Anti-Fibrotic Effects against Liver Fibrosis" International Journal of Molecular Sciences 23, no. 10: 5420. https://doi.org/10.3390/ijms23105420
APA StyleAbdullah, A. S., Sayed, I. E. T. E., El-Torgoman, A. M. A., Kalam, A., Wageh, S., & Kamel, M. A. (2022). Green Synthesis of Silymarin–Chitosan Nanoparticles as a New Nano Formulation with Enhanced Anti-Fibrotic Effects against Liver Fibrosis. International Journal of Molecular Sciences, 23(10), 5420. https://doi.org/10.3390/ijms23105420