
LC3/FtMt Colocalization Patterns Reveal the Progression of FtMt Accumulation in Nigral Neurons of Patients with Progressive Supranuclear Palsy

 ,
, 
Abstract
:1. Introduction
2. Results
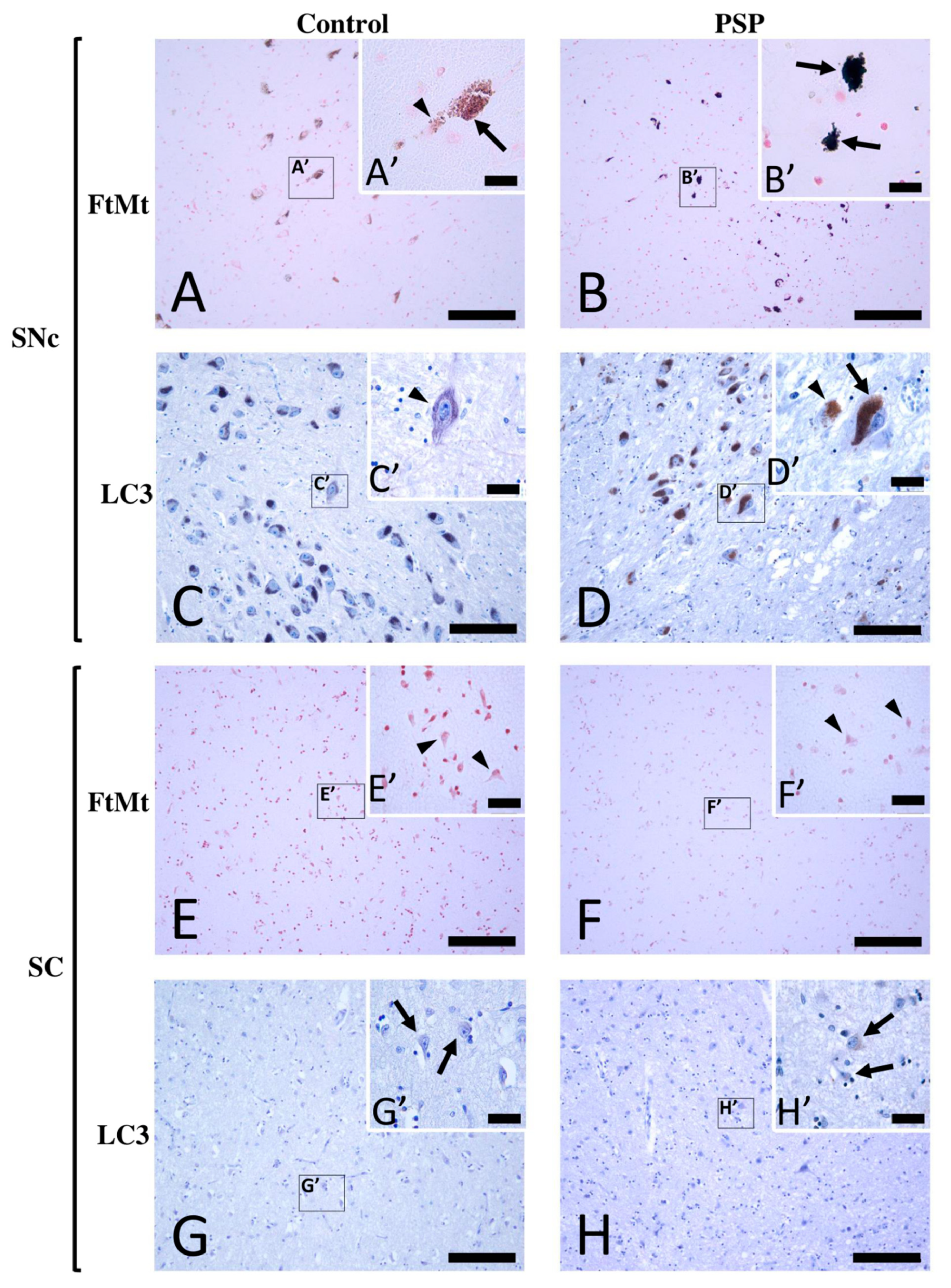
2.1. FtMt and LC3 Immunoreactivity in the Midbrain of Control and PSP Cases
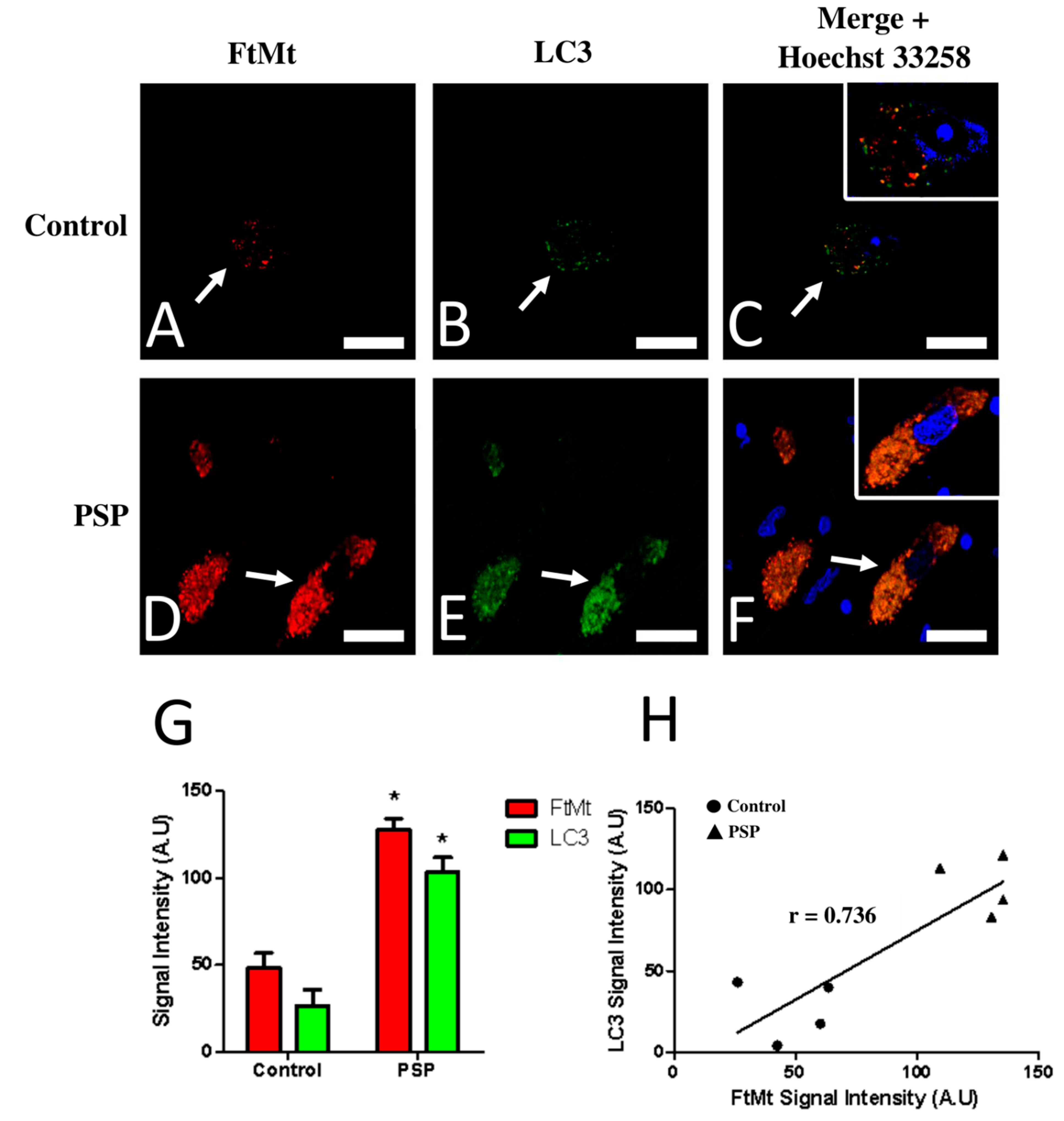
2.2. FtMt and LC3 Colocalization in the Substantia Nigra of Control and PSP Cases
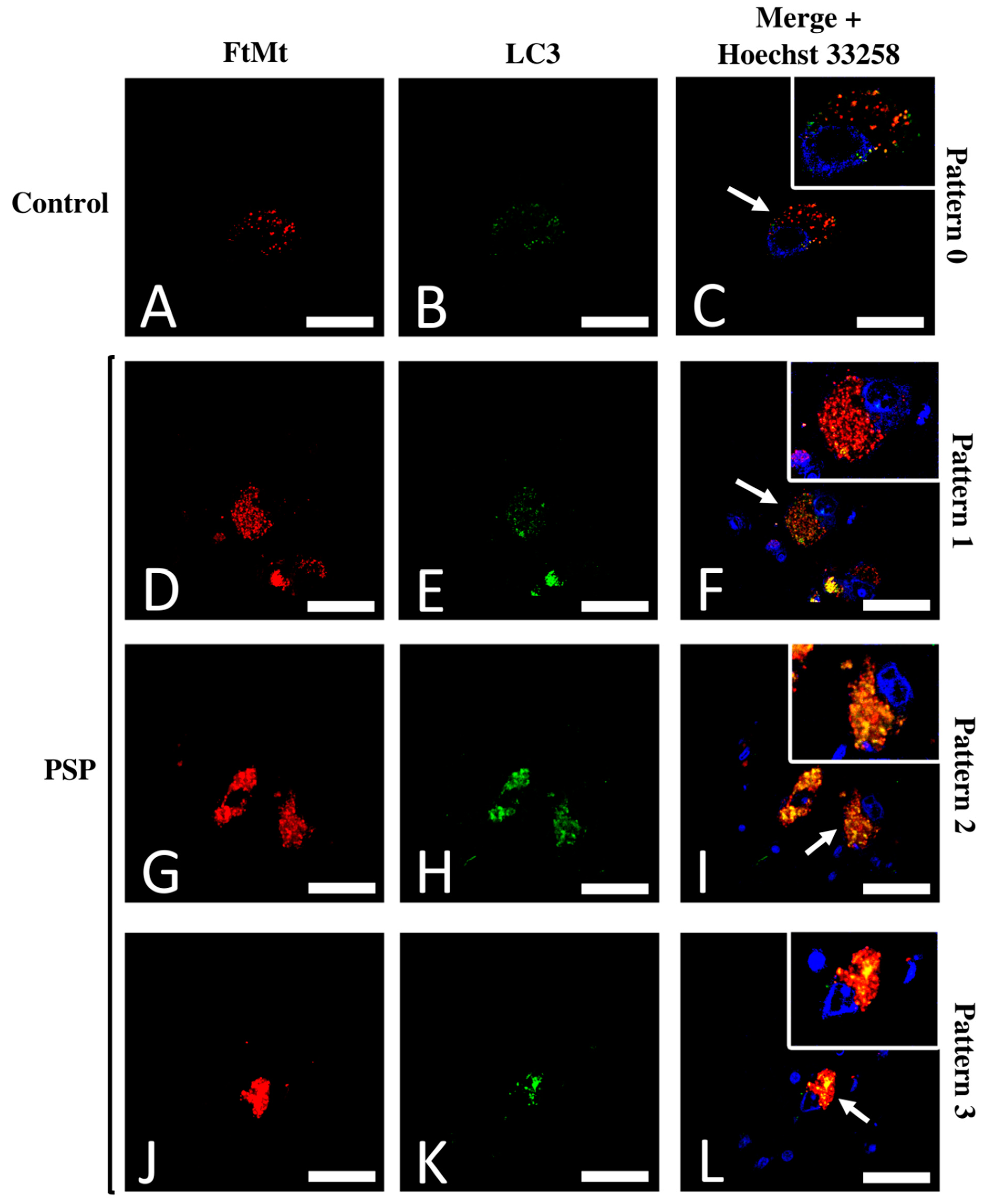
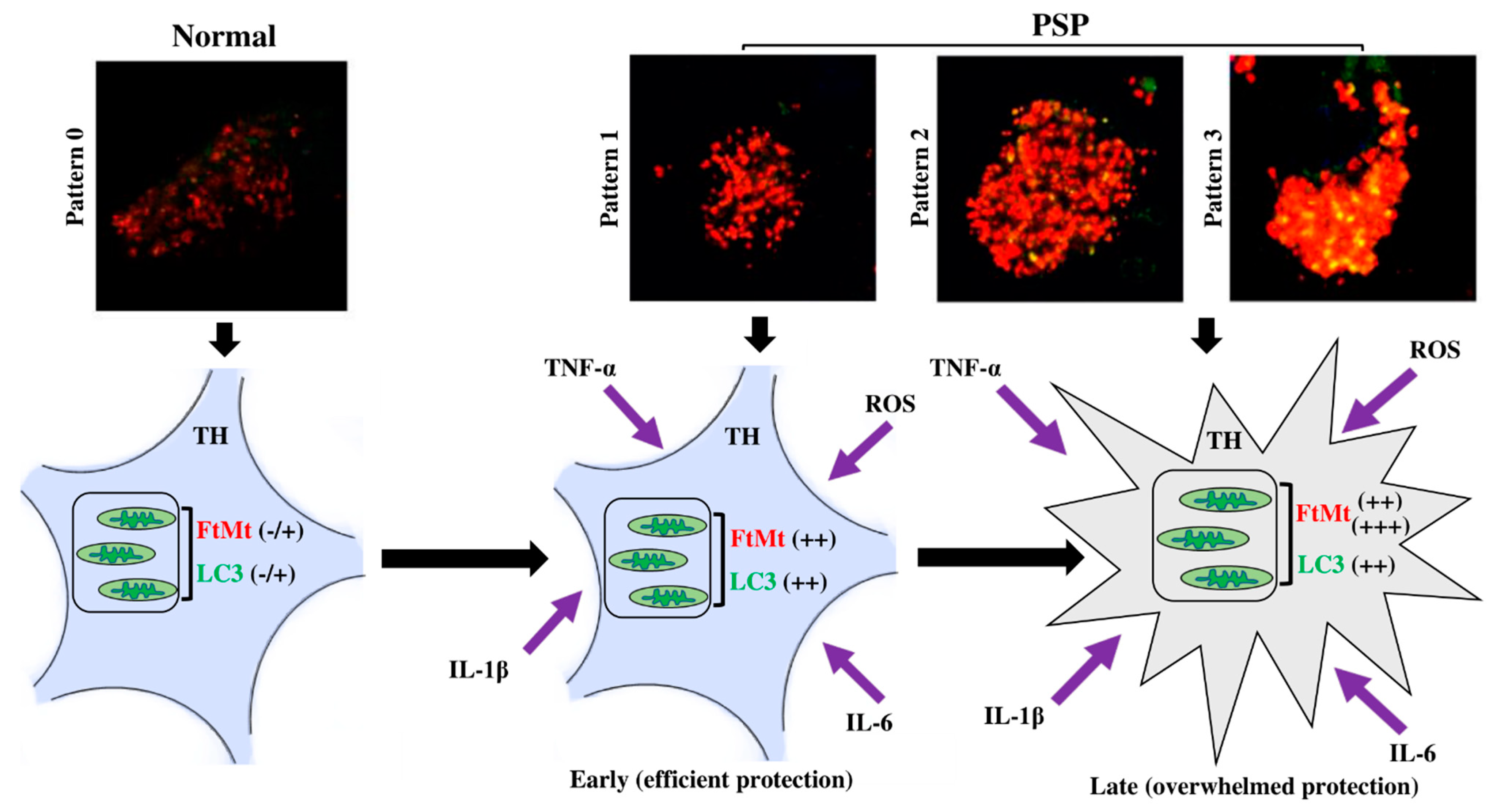
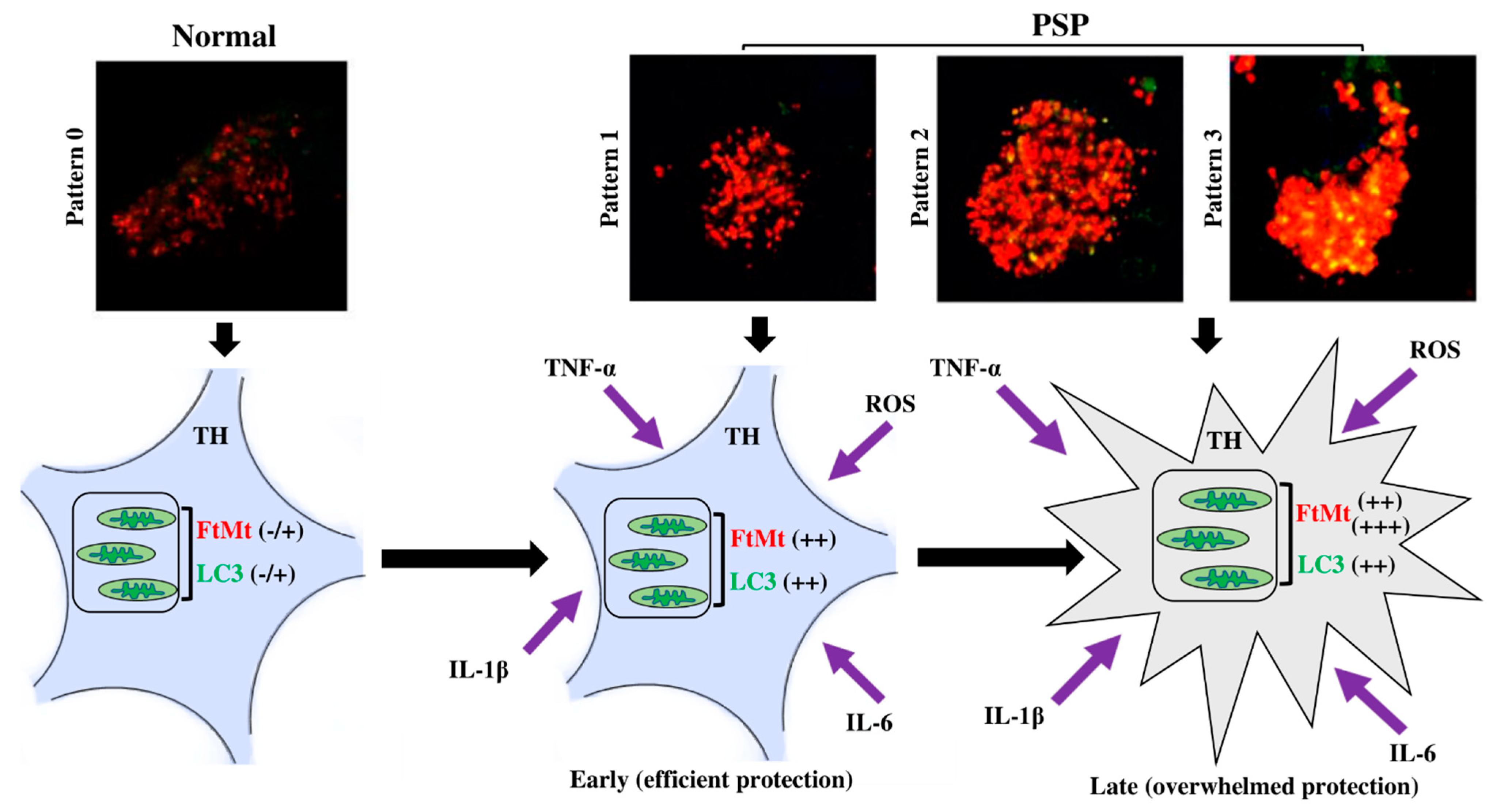
2.3. Subpopulations of FtMt-IR Neurons in SNcs of PSP Patients
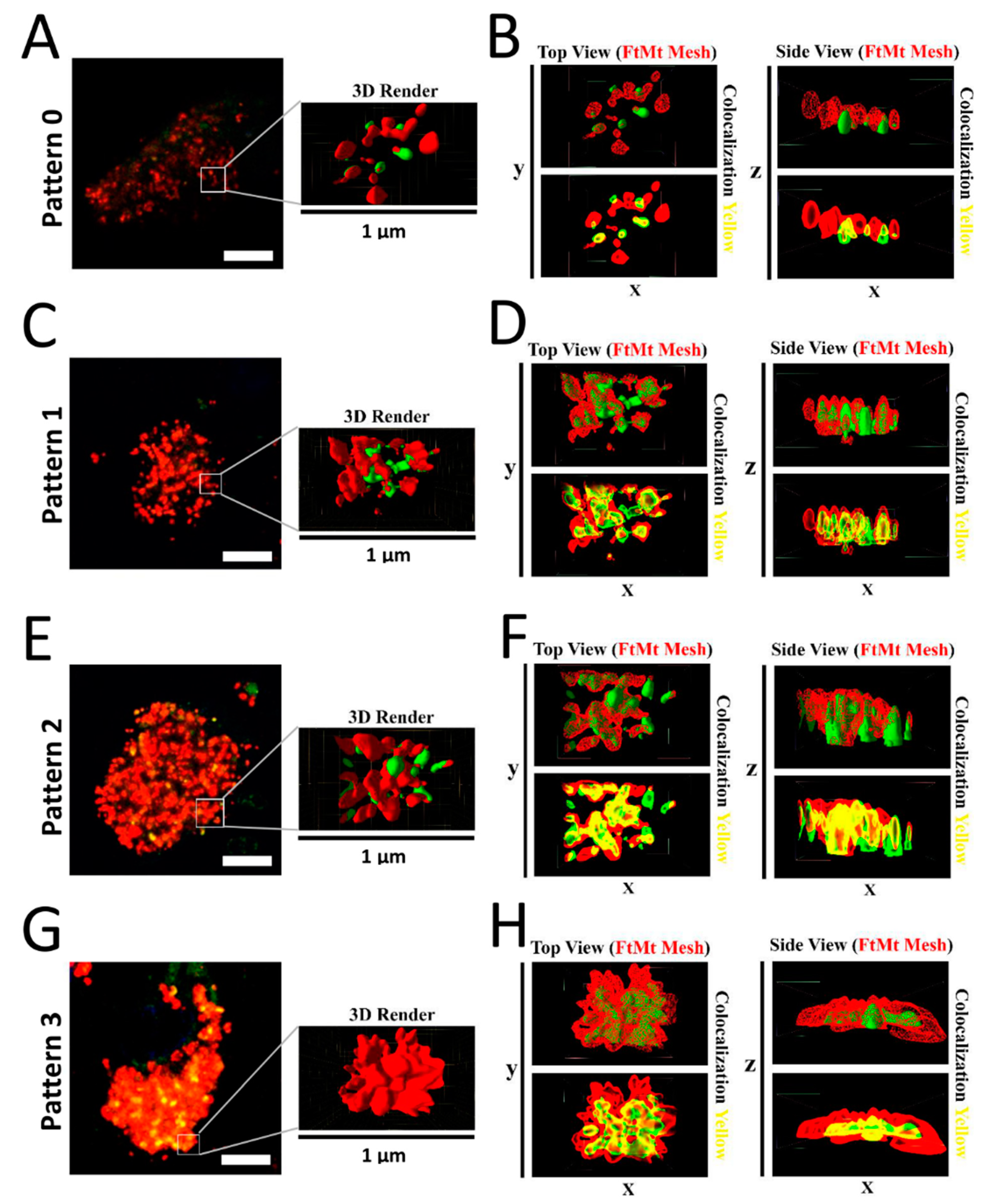
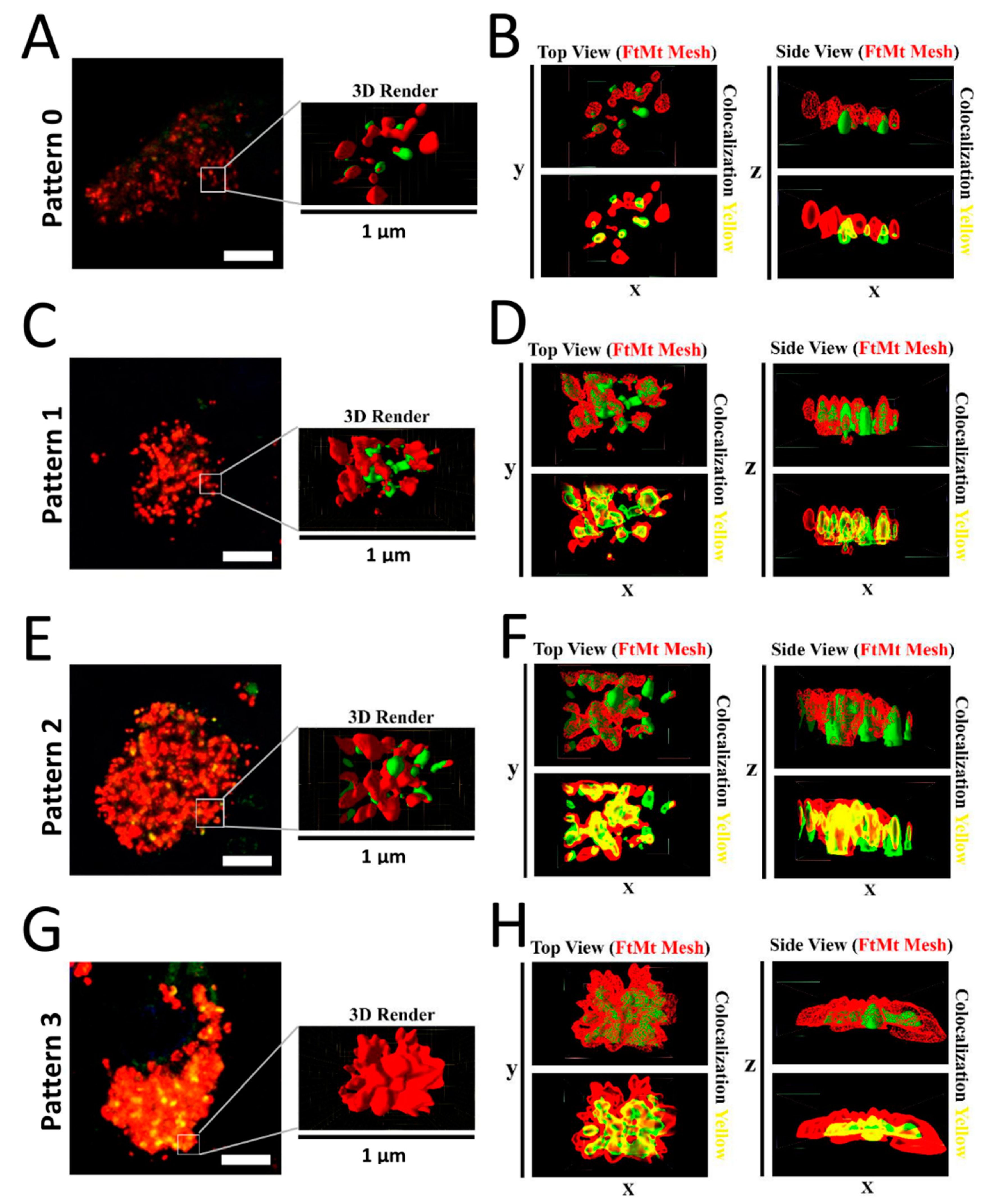
2.4. Assessment of FtMt-IR and LC3-IR Subcellular Arrangement Using 3D Imaging
3. Discussion
4. Materials and Methods
4.1. Human Brain Sample
4.2. Immunohistochemical Localization of FtMt- and LC3-Immunoreactivities
4.3. Double Immunofluorescence Histochemistry for FtMt and LC3
4.4. Characterization of FtMt and LC3 Colocalization
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murphy, K.E.; Karaconji, T.; Hardman, C.D.; Halliday, G.M. Excessive dopamine neuron loss in progressive supranuclear palsy. Mov. Disord. 2008, 23, 607–610. [Google Scholar] [CrossRef]
- Mastaglia, F.L.; Grainger, K.; Kee, F.; Sadka, M.; Lefroy, R. Progressive supranuclear palsy (the Steele–Richardson–Olszewski syndrome) clinical and electrophysiological observations in eleven cases. Proc. Aust. Assoc. Neurol. 1973, 10, 35–44. [Google Scholar]
- Alster, P.; Madetko, N.; Koziorowski, D.; Friedman, A. Progressive supranuclear palsy—Parkinsonism predominant (PSP-P)—A clinical challenge at the boundaries of PSP and Parkinson’s disease (PD). Front. Neurol. 2020, 11, 180. [Google Scholar] [CrossRef]
- DelveInsight. Progressive supranuclear palsy (PSP)—Epidemiology forecast. Available online: https://www.researchandmarkets.com/reports/5025131/progressive-supranuclear-palsy-psp (accessed on 3 October 2021).
- National Institute of Neurological Disorders and Stroke Progressive Supranuclear Palsy Fact Sheet. Available online: https://www.ninds.nih.gov/Disorders/Patient-Caregiver-Education/Fact-Sheets/Progressive-Supranuclear-Palsy-Fact-Sheet (accessed on 10 October 2021).
- Bakar, Z.H.A.; Kato, T.; Yanagisawa, D.; Bellier, J.-P.; Mukaisho, K.-i.; Tooyama, I. Immunohistochemical study of mitochondrial ferritin in the midbrain of patients with progressive supranuclear palsy. Acta Histochem. Cytochem. 2021, 54, 97–104. [Google Scholar] [CrossRef]
- Nogami, A.; Yamazaki, M.; Saito, Y.; Hatsuta, H.; Sakiyama, Y.; Takao, M.; Kimura, K.; Murayama, S. Early stage of progressive supranuclear palsy: A neuropathological study of 324 consecutive autopsy cases. J. Nippon Med. Sch. 2015, 82, 266–273. [Google Scholar] [CrossRef] [Green Version]
- Albers, D.S.; Beal, M.F. Mitochondrial dysfunction in progressive supranuclear palsy. Neurochem. Int. 2002, 40, 559–564. [Google Scholar] [CrossRef]
- Albers, D.S.; Swerdlow, R.H.; Manfredi, G.; Gajewski, C.; Yang, L.; Parker, W.D., Jr.; Beal, M.F. Further evidence for mitochondrial dysfunction in progressive supranuclear palsy. Exp. Neurol. 2001, 168, 196–198. [Google Scholar] [CrossRef]
- Gao, G.; Chang, Y.Z. Mitochondrial ferritin in the regulation of brain iron homeostasis and neurodegenerative diseases. Front. Pharmacol. 2014, 5, 19. [Google Scholar] [CrossRef]
- Levi, S.; Corsi, B.; Bosisio, M.; Invernizzi, R.; Volz, A.; Sanford, D.; Arosio, P.; Drysdale, J. A human mitochondrial ferritin encoded by an intronless gene. J. Biol. Chem. 2001, 276, 24437–24440. [Google Scholar] [CrossRef] [Green Version]
- Corsi, B.; Cozzi, A.; Arosio, P.; Drysdale, J.; Santambrogio, P.; Campanella, A.; Biasiotto, G.; Albertini, A.; Levi, S. Human mitochondrial ferritin expressed in HeLa cells incorporates iron and affects cellular iron metabolism. J. Biol. Chem. 2002, 277, 22430–22437. [Google Scholar] [CrossRef] [Green Version]
- Nie, G.; Sheftel, A.D.; Kim, S.F.; Ponka, P. Overexpression of mitochondrial ferritin causes cytosolic iron depletion and changes cellular iron homeostasis. Blood 2005, 105, 2161–2167. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Yang, M.; Guan, H.; Liu, Z.; Zhao, S.; Takeuchi, S.; Yanagisawa, D.; Tooyama, I. Mitochondrial ferritin in neurodegenerative diseases. Neurosci. Res. 2013, 77, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Guan, H.; Yang, M.; Liu, Z.; Takeuchi, S.; Yanagisawa, D.; Vincent, S.R.; Zhao, S.; Tooyama, I. Upregulation of mitochondrial ferritin by proinflammatory cytokines: Implications for a role in Alzheimer’s disease. J. Alzheimers Dis. 2015, 45, 797–811. [Google Scholar] [CrossRef]
- Gao, G.; Zhang, N.; Wang, Y.Q.; Wu, Q.; Yu, P.; Shi, Z.H.; Duan, X.L.; Zhao, B.L.; Wu, W.S.; Chang, Y.Z. Mitochondrial ferritin protects hydrogen peroxide-induced neuronal cell damage. Aging Dis. 2017, 8, 458–470. [Google Scholar] [CrossRef] [Green Version]
- Kandimalla, R.; Manczak, M.; Yin, X.; Wang, R.; Reddy, P.H. Hippocampal phosphorylated tau induced cognitive decline, dendritic spine loss and mitochondrial abnormalities in a mouse model of Alzheimer’s disease. Hum. Mol. Genet. 2018, 27, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Reddy, P.H.; Oliver, D.M. Amyloid beta and phosphorylated tau-induced defective autophagy and mitophagy in Alzheimer’s disease. Cells 2019, 8, 488. [Google Scholar] [CrossRef] [Green Version]
- Manczak, M.; Reddy, P.H. Abnormal interaction between the mitochondrial fission protein Drp1 and hyperphosphorylated tau in Alzheimer’s disease neurons: Implications for mitochondrial dysfunction and neuronal damage. Hum. Mol. Genet. 2012, 21, 2538–2547. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Yang, H.; Yanagisawa, D.; Bellier, J.P.; Morino, K.; Zhao, S.; Liu, P.; Vigers, P.; Tooyama, I. Mitochondrial ferritin affects mitochondria by stabilizing HIF-1α in retinal pigment epithelium: Implications for the pathophysiology of age-related macular degeneration. Neurobiol. Aging 2016, 47, 168–179. [Google Scholar] [CrossRef] [Green Version]
- Towers, C.G.; Wodetzki, D.K.; Thorburn, J.; Smith, K.R.; Caino, M.C.; Thorburn, A. Mitochondrial-derived vesicles compensate for loss of LC3-mediated mitophagy. Dev. Cell 2021, 56, 2029–2042.e5. [Google Scholar] [CrossRef]
- Swerdlow, N.S.; Wilkins, H.M. Mitophagy and the brain. Int. J. Mol. Sci. 2020, 21, 9661. [Google Scholar] [CrossRef]
- Piras, A.; Collin, L.; Grüninger, F.; Graff, C.; Rönnbäck, A. Autophagic and lysosomal defects in human tauopathies: Analysis of post-mortem brain from patients with familial Alzheimer disease, corticobasal degeneration and progressive supranuclear palsy. Acta Neuropathol. Commun. 2016, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wang, X.; Fei, X.; Xia, L.; Qin, Z.; Liang, Z. Parkinson’s disease involves autophagy and abnormal distribution of cathepsin L. Neurosci. Lett. 2011, 489, 62–67. [Google Scholar] [CrossRef]
- Liu, B.; Sun, J.; Zhang, J.; Mao, W.; Ma, Y.; Li, S.; Cheng, X.; Lv, C. Autophagy-related protein expression in the substantia nigra and Eldepryl intervention in rat models of Parkinson’s disease. Brain Res. 2015, 1625, 180–188. [Google Scholar] [CrossRef]
- Boman, A. Lysosomal Network Proteins as Biomarkers and Therapeutic Targets in Neurodegenerative Disease. Ph.D. Thesis, Linköping University, Linköping, Sweden, 15 November 2015. [Google Scholar]
- Krüger, U.; Wang, Y.; Kumar, S.; Mandelkow, E.M. Autophagic degradation of tau in primary neurons and its enhancement by trehalose. Neurobiol. Aging 2012, 33, 2291–2305. [Google Scholar] [CrossRef]
- Wang, Y.; Mandelkow, E. Degradation of Tau protein by autophagy and proteasomal pathways. Biochem. Soc. Trans. 2012, 40, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Martinez-Vicente, M.; Krüger, U.; Kaushik, S.; Wong, E.; Mandelkow, E.M.; Cuervo, A.M.; Mandelkow, E. Tau fragmentation, aggregation and clearance: The dual role of lysosomal processing. Hum. Mol. Genet. 2009, 18, 4153–4170. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.J.; Lee, J.H.; Rubinsztein, D.C. Tau degradation: The ubiquitin–proteasome system versus the autophagy-lysosome system. Prog. Neurobiol. 2013, 105, 49–59. [Google Scholar] [CrossRef]
- Hansen, T.E.; Johansen, T. Following autophagy step by step. BMC Biol. 2011, 9, 39. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.M.; Jung, Y.K. A molecular approach to mitophagy and mitochondrial dynamics. Mol. Cells 2018, 41, 18–26. [Google Scholar]
- Sarkar, C.; Zhao, Z.; Aungst, S.; Sabirzhanov, B.; Faden, A.I.; Lipinski, M.M. Impaired autophagy flux is associated with neuronal cell death after traumatic brain injury. Autophagy 2014, 10, 2208–2222. [Google Scholar] [CrossRef] [Green Version]
- Cui, D.; Sun, D.; Wang, X.; Yi, L.; Kulikowicz, E.; Reyes, M.; Zhu, J.; Yang, Z.J.; Jiang, W.; Koehler, R.C. Impaired autophagosome clearance contributes to neuronal death in a piglet model of neonatal hypoxic-ischemic encephalopathy. Cell Death Dis. 2017, 8, e2919. [Google Scholar] [CrossRef] [Green Version]
- Long, Z.; Chen, J.; Zhao, Y.; Zhou, W.; Yao, Q.; Wang, Y.; He, G. Dynamic changes of autophagic flux induced by Abeta in the brain of postmortem Alzheimer’s disease patients, animal models and cell models. Aging 2020, 12, 10912–10930. [Google Scholar] [CrossRef]
- Wang, L.; Yang, H.; Zhao, S.; Sato, H.; Konishi, Y.; Beach, T.G.; Abdelalim, E.M.; Bisem, N.J.; Tooyama, I. Expression and localization of mitochondrial ferritin mRNA in Alzheimer’s disease cerebral cortex. PLoS ONE 2011, 6, e22325. [Google Scholar] [CrossRef]
- Alster, P.; Madetko, N.; Koziorowski, D.; Friedman, A. Microglial activation and inflammation as a factor in the pathogenesis of progressive supranuclear palsy (PSP). Front. Neurosci. 2020, 14, 893. [Google Scholar] [CrossRef]
- Filomeni, G.; De Zio, D.; Cecconi, F. Oxidative stress and autophagy: The clash between damage and metabolic needs. Cell Death Differ. 2015, 22, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Trist, B.G.; Hare, D.J.; Double, K.L. Oxidative stress in the aging substantia nigra and the etiology of Parkinson’s disease. Aging Cell 2019, 18, e13031. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ma, H.; Sun, J.; Zheng, T.; Zhao, P.; Li, H.; Yang, M. Mitochondrial ferritin deficiency promotes osteoblastic ferroptosis via mitophagy in Type 2 diabetic osteoporosis. Biol. Trace Elem. Res. 2021, 200, 298–307. [Google Scholar] [CrossRef]
- Kageyama, Y.; Saito, A.; Pletnikova, O.; Rudow, G.L.; Irie, Y.; An, Y.; Murakami, K.; Irie, K.; Resnick, S.M.; Fowler, D.R. Amyloid β toxic conformer has dynamic localization in the human inferior parietal cortex in absence of amyloid plaques. Sci. Rep. 2018, 8, 1–17. [Google Scholar]
- Abdelalim, E.M.; Tooyama, I. Mapping of NPR-B immunoreactivity in the brainstem of Macaca fascicularis. Brain Struct. Funct. 2011, 216, 387–402. [Google Scholar] [CrossRef]
- Yang, M.; Yang, H.; Guan, H.; Kato, T.; Mukaisho, K.; Sugihara, H.; Ogasawara, K.; Terada, T.; Tooyama, I. Characterization of a novel monoclonal antibody against human mitochondrial ferritin and its immunohistochemical application in human and monkey substantia Nigra. Acta Histochem. Cytochem. 2017, 50, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Stauffer, W.; Sheng, H.; Lim, H.N. EzColocalization: An ImageJ plugin for visualizing and measuring colocalization in cells and organisms. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Meshlab. Available online: https://www.meshlab.net/ (accessed on 20 September 2021).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Pearson Correlation Coefficient (PCC) of the Colocalized Signal | PCC Transformed Value (z-Score) |
|---|---|---|
| Control-1 | 0.539 | −0.175 |
| Control-2 | 0.419 | −0.761 |
| Control-3 | 0.333 | −1.181 |
| Control-4 | 0.339 | −1.151 |
| PSP-1 | 0.655 | 0.391 |
| PSP-2 | 0.629 | 0.264 |
| PSP-3 | 0.815 | 1.172 |
| PSP-4 | 0.87 | 1.440 |
| Pattern | FtMt-IR * | LC3-IR * | Colocalization * | Feature |
|---|---|---|---|---|
| 0 | −/+, scattered, punctiform | −/+, scattered, punctiform | −/+ | Rare colocalization |
| 1 | ++, numerous, punctiform | +, scattered punctiform | + | Overlapping colocalization |
| 2 | ++, numerous, punctiform | ++, numerous, punctiform | ++ | Overlapping colocalization |
| 3 | +++, dense, packed | ++, numerous, punctiform and granular | +++ | LC3-IR enclosed in FtMt-IR |
| Cases | Age (years) | Gender | Post-Mortem Delay (h) | Clinical Diagnosis |
|---|---|---|---|---|
| Control-1 | 60 | M | 5.0 | Pancreatic cancer |
| Control-2 | 52 | M | 10.0 | Malignant lymphoma |
| Control-3 | 83 | F | 6.5 | Malignant lymphoma |
| Control-4 | 64 | M | 12.0 | Prostate cancer/subdural hemorrhage |
| PSP-1 | 47 | M | <12.0 * | PSP |
| PSP-2 | 68 | F | 5.0 | PSP |
| PSP-3 | 76 | M | 10.0 | PSP |
| PSP-4 | 69 | M | 13.0 | PSP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abu Bakar, Z.H.; Bellier, J.-P.; Yanagisawa, D.; Kato, T.; Mukaisho, K.-i.; Tooyama, I. LC3/FtMt Colocalization Patterns Reveal the Progression of FtMt Accumulation in Nigral Neurons of Patients with Progressive Supranuclear Palsy. Int. J. Mol. Sci. 2022, 23, 537. https://doi.org/10.3390/ijms23010537
Abu Bakar ZH, Bellier J-P, Yanagisawa D, Kato T, Mukaisho K-i, Tooyama I. LC3/FtMt Colocalization Patterns Reveal the Progression of FtMt Accumulation in Nigral Neurons of Patients with Progressive Supranuclear Palsy. International Journal of Molecular Sciences. 2022; 23(1):537. https://doi.org/10.3390/ijms23010537
Chicago/Turabian StyleAbu Bakar, Zulzikry Hafiz, Jean-Pierre Bellier, Daijiro Yanagisawa, Tomoko Kato, Ken-ichi Mukaisho, and Ikuo Tooyama. 2022. "LC3/FtMt Colocalization Patterns Reveal the Progression of FtMt Accumulation in Nigral Neurons of Patients with Progressive Supranuclear Palsy" International Journal of Molecular Sciences 23, no. 1: 537. https://doi.org/10.3390/ijms23010537
APA StyleAbu Bakar, Z. H., Bellier, J.-P., Yanagisawa, D., Kato, T., Mukaisho, K.-i., & Tooyama, I. (2022). LC3/FtMt Colocalization Patterns Reveal the Progression of FtMt Accumulation in Nigral Neurons of Patients with Progressive Supranuclear Palsy. International Journal of Molecular Sciences, 23(1), 537. https://doi.org/10.3390/ijms23010537