
Stem Cell Therapy for Modulating Neuroinflammation in Neuropathic Pain

 ,
,  , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Chronic NP
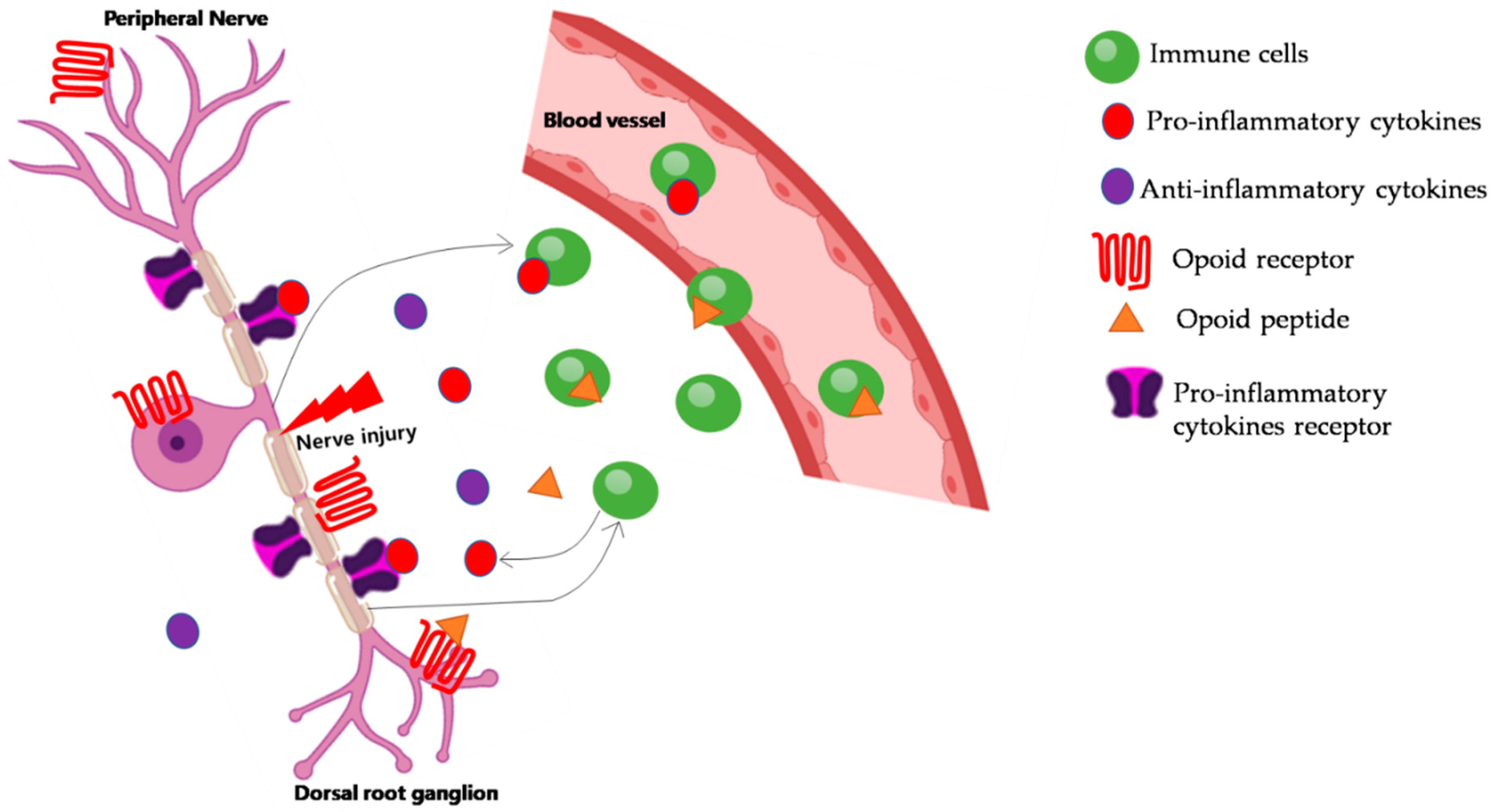
3. Role of Immune Cells in NP
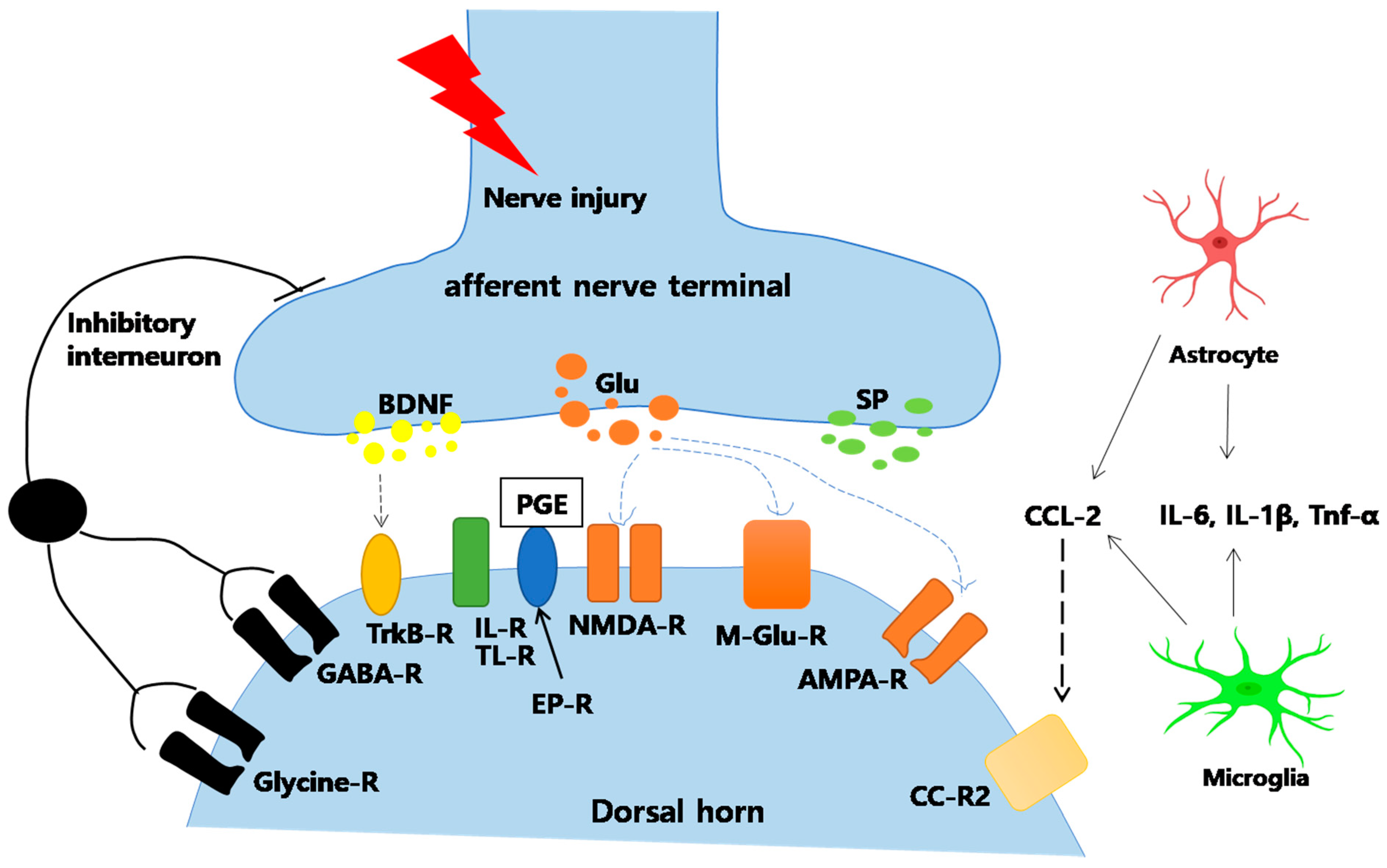
4. Inflammation and Pain
5. The Alternative/Nonpharmacological Treatment of Neuropathic Pain: Spinal Cord Stimulation (SCS)
6. Stem Cell Therapy
6.1. Stem Cell Mechanism of Action in the Pain Recovery Process
6.1.1. Effect of Stem Cells in Peripheral NP
Anti-Inflammatory Regulation
6.1.2. Effect of Stem Cells in Spinal NP
Attenuation of Central Sensitization
Inhibition of Glial Cell Activation
6.2. Viral Vector-Mediated Gene Transfer into Stem Cells
Factors Limiting Gene Transfer into Stem Cells
7. Potential Shortcomings
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMPAR | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor |
| NP | neuropathic pain |
| IASP | International Association for the Study of Pain |
| ICD | International Classification of Diseases |
| SCI | spinal cord injury |
| CCI | chronic constriction injury |
| DRG | dorsal root ganglia |
| TNF-α | tumor necrosis factor-α |
| IL-1β | interleukin-1β |
| cAMP | cyclic adenosine monophosphate |
| PKC | protein kinase C |
| DAG | diacylglycerol |
| cGMP | cyclic guanosine monophosphate |
| TRPA1 | Transient receptor potential cation channel subfamily A member 1 |
| TRPV1 | transient receptor potential cation channel subfamily V member 1, |
| TRPM8 | Transient receptor potential cation channel subfamily M (melastatin) member 8 |
| PKA | protein kinase A |
| PKC | protein kinase C |
| SCS | spinal cord stimulation |
| FBSS | failed back surgery syndrome |
| CRPS | complex regional pain syndrome |
| TCAs | Tricyclic antidepressants |
| CSF | cerebrospinal fluid |
| SVZ | subventricular zone |
| Glu-R | glutamate receptor |
| GFAP | Glial fibrillary acidic protein |
| IBA-1 | Ionized calcium-binding adaptor protein-1 |
| MAPK | mitogen-activated protein kinase |
| hMSCs | human mesenchymal stem cells |
| SNI | spared nerve injury |
| NSCs | neural stem cells |
| MSCs | mesenchymal stem cells |
| BM-MSCs | bone marrow-derived MSC |
| hBMSC | human bone marrow-derived MSC |
| AD-MSCs | adipose tissue-derived MSC |
| UCB-MSC | umbilical cord blood-derived stem cell |
| hUB-MSC | human umbilical cord blood-derived mesenchymal stem cell |
| WJ-MSCs | Wharton’s jelly-derived MSCs |
| SLNC | single ligature nerve constriction |
| DPSC | dental pulp stem cell |
| GDNF | glial-derived neurotrophic factor, |
| VEGF | vascular endothelial factor |
| NGF | nerve growth factor |
| TGF-β1 | transforming growth factor-β1 |
| Glu-R | glutamate receptor |
| NMDAR | N-methyl-d-aspartate (NMDA) receptor |
| ALS | Amyotrophic lateral sclerosis |
References
- Aiyer, R.; Mehta, N.; Gungor, S.; Gulati, A. A Systematic Review of NMDA Receptor Antagonists for Treatment of Neuropathic Pain in Clinical Practice. Clin. J. Pain 2018, 34, 450–467. [Google Scholar] [CrossRef] [PubMed]
- Hylands-White, N.; Duarte, R.V.; Raphael, J.H. An overview of treatment approaches for chronic pain management. Rheumatol. Int. 2017, 37, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Varshney, V.; Osborn, J.; Chaturvedi, R.; Shah, V.; Chakravarthy, K. Advances in the interventional management of neuropathic pain. Ann. Transl. Med. 2021, 9, 187. [Google Scholar] [CrossRef] [PubMed]
- Scholz, J.; Finnerup, N.B.; Attal, N.; Aziz, Q.; Baron, R.; Bennett, M.I.; Benoliel, R.; Cohen, M.; Cruccu, G.; Davis, K.D.; et al. The IASP classification of chronic pain for ICD-11: Chronic neuropathic pain. Pain 2019, 160, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Rosenberger, D.C.; Blechschmidt, V.; Timmerman, H.; Wolff, A.; Treede, R.-D. Challenges of neuropathic pain: Focus on diabetic neuropathy. J. Neural Transm. 2020, 127, 589–624. [Google Scholar] [CrossRef] [PubMed]
- Szok, D.; Tajti, J.; Nyári, A.; Vécsei, L.; Trojano, L. Therapeutic Approaches for Peripheral and Central Neuropathic Pain. Behav. Neurol. 2019, 2019, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Safieh-Garabedian, B.; Nomikos, M.; Saadé, N. Targeting inflammatory components in neuropathic pain: The analgesic effect of thymulin related peptide. Neurosci. Lett. 2019, 702, 61–65. [Google Scholar] [CrossRef]
- Kaur, G.; Singh, N.; Jaggi, A.S. Mast cells in neuropathic pain: An increasing spectrum of their involvement in pathophysiology. Rev. Neurosci. 2017, 28, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Ilkhanizadeh, B.; Zarei, L.; Farhad, N.; Bahrami-Bukani, M.; Mohammadi, R. Mast cells improve functional recovery of transected peripheral nerve: A novel preliminary study. Injury 2017, 48, 1480–1485. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Hore, Z.; Harley, P.; Stanley, F.U.; Michrowska, A.; Dahiya, M.; La Russa, F.; Jager, S.E.; Villa-Hernandez, S.; Denk, F. A transcriptional toolbox for exploring peripheral neuro-immune interactions. Pain 2020, 161, 2089–2106. [Google Scholar] [CrossRef]
- Gong, W.R.-Y.; Abdelhamid, E.; Carvalho, C.S.; Sluka, K.A. Resident Macrophages in Muscle Contribute to Development of Hyperalgesia in a Mouse Model of Noninflammatory Muscle Pain. J. Pain 2016, 17, 1081–1094. [Google Scholar] [CrossRef] [PubMed]
- Montague, K.; Malcangio, M.J.F. The therapeutic potential of monocyte/macrophage manipulation in the treatment of chemotherapy-induced painful neuropathy. Front. Mol. Neurosci. 2017, 10, 397. [Google Scholar] [CrossRef]
- Wang, Y.-R.; Mao, X.-F.; Wu, H.-Y.; Wang, Y.-X.J.B.; Communications, B.R. Liposome-encapsulated clodronate specifically depletes spinal microglia and reduces initial neuropathic pain. Biochem. Biophys. Res. Commun. 2018, 499, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Austin, P.J.; Berglund, A.M.; Siu, S.; Fiore, N.T.; Gerke-Duncan, M.B.; Ollerenshaw, S.L.; Leigh, S.-J.; Kunjan, P.A.; Kang, J.W.M.; Keay, K.A. Evidence for a distinct neuro-immune signature in rats that develop behavioural disability after nerve injury. J. NeuroInflamm. 2015, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Alam, A.; Chen, Q.; Eusman, M.A.; Pal, A.; Eguchi, S.; Wu, L.; Ma, D. The role of microglia in the pathobiology of neuropathic pain development: What do we know? BJA Br. J. Anaesth. 2017, 118, 504–516. [Google Scholar] [CrossRef]
- Tsuda, M. Microglia in the spinal cord and neuropathic pain. J. Diabetes Investig. 2016, 7, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chen, X.; Zhang, C.; Zhang, Y.; Yao, W. An update on reactive astrocytes in chronic pain. J. Neuroinflamm. 2019, 16, 1–13. [Google Scholar] [CrossRef]
- Joshi, H.P.; Kim, S.B.; Kim, S.; Kumar, H.; Jo, M.-J.; Choi, H.; Kim, J.; Kyung, J.W.; Sohn, S.; Kim, K.-T.; et al. Nanocarrier-mediated Delivery of CORM-2 Enhances Anti-allodynic and Anti-hyperalgesic Effects of CORM-2. Mol. Neurobiol. 2019, 56, 5539–5554. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Yamashita, T. Microglia in central nervous system repair after injury. J. Biochem. 2016, 159, 491–496. [Google Scholar] [CrossRef]
- Davies, A.J.; Kim, H.W.; Gonzalez-Cano, R.; Choi, J.; Back, S.K.; Roh, S.E.; Johnson, E.; Gabriac, M.; Kim, M.-S.; Lee, J.; et al. Natural Killer Cells Degenerate Intact Sensory Afferents following Nerve Injury. Cell 2019, 176, 716–728. [Google Scholar] [CrossRef]
- Gadani, S.P.; Walsh, J.T.; Lukens, J.R.; Kipnis, J. Dealing with Danger in the CNS: The Response of the Immune System to Injury. Neuron 2015, 87, 47–62. [Google Scholar] [CrossRef]
- Gattlen, C.; Clarke, C.B.; Piller, N.; Kirschmann, G.; Pertin, M.; Decosterd, I.; Gosselin, R.-D.; Suter, M.R. Spinal Cord T-Cell Infiltration in the Rat Spared Nerve Injury Model: A Time Course Study. Int. J. Mol. Sci. 2016, 17, 352. [Google Scholar] [CrossRef] [PubMed]
- Austin, P.J.; Kim, C.F.; Perera, C.J.; Moalem-Taylor, G. Regulatory T cells attenuate neuropathic pain following peripheral nerve injury and experimental autoimmune neuritis. Pain 2012, 153, 1916–1931. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.; Dawes, J.M.; Bennett, D.L. The role of the immune system in the generation of neuropathic pain. Lancet Neurol. 2012, 11, 629–642. [Google Scholar] [CrossRef]
- Jurjus, A.; Eid, A.; Al Kattar, S.; Zeenny, M.N.; Gerges-Geagea, A.; Haydar, H.; Hilal, A.; Oueidat, D.; Matar, M.; Tawilah, J.; et al. Inflammatory bowel disease, colorectal cancer and type 2 diabetes mellitus: The links. BBA Clin. 2016, 5, 16–24. [Google Scholar] [CrossRef]
- Matsuda, M.; Huh, Y.; Ji, R.-R. Roles of inflammation, neurogenic inflammation, and neuroinflammation in pain. J. Anesth. 2019, 33, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Yam, M.F.; Loh, Y.C.; Tan, C.S.; Adam, S.K.; Manan, N.A.; Basir, R. General Pathways of Pain Sensation and the Major Neurotransmitters Involved in Pain Regulation. Int. J. Mol. Sci. 2018, 19, 2164. [Google Scholar] [CrossRef]
- Park, C.-K.; Kim, M.S.; Fang, Z.; Li, H.Y.; Jung, S.J.; Choi, S.-Y.; Lee, S.J.; Park, K.; Kim, J.S.; Oh, S.B. Functional expression of thermo-transient receptor potential channels in dental primary afferent neurons implication for tooth pain. J. Biol. Chem. 2006, 281, 17304–17311. [Google Scholar] [CrossRef]
- Roh, J.; Hwang, S.-M.; Lee, S.-H.; Lee, K.; Kim, Y.H.; Park, C.-K. Functional Expression of Piezo1 in Dorsal Root Ganglion (DRG) Neurons. Int. J. Mol. Sci. 2020, 21, 3834. [Google Scholar] [CrossRef]
- Rogers, M.; Tang, L.; Madge, D.J.; Stevens, E.B. The role of sodium channels in neuropathic pain. Semin. Cell Dev. Biol. 2006, 17, 571–581. [Google Scholar] [CrossRef]
- Song, X.-J.; Wang, Z.-B.; Gan, Q.; Walters, E.T. cAMP and cGMP Contribute to Sensory Neuron Hyperexcitability and Hyperalgesia in Rats With Dorsal Root Ganglia Compression. J. Neurophysiol. 2006, 95, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.K.; Ma, Y.; Xie, H. TRPV1 and spinal astrocyte activation contribute to remifentanil-induced hyperalgesia in rats. NeuroReport 2019, 30, 1095–1101. [Google Scholar] [CrossRef] [PubMed]
- Dhukhwa, A.; Bhatta, P.; Sheth, S.; Korrapati, K.; Tieu, C.; Mamillapalli, C.K.; Ramkumar, V.; Mukherjea, D. Targeting Inflammatory Processes Mediated by TRPV1 and TNF-α for Treating Noise-Induced Hearing Loss. Front. Cell. Neurosci. 2019, 13, 444. [Google Scholar] [CrossRef]
- Binshtok, A.M.; Wang, H.; Zimmermann, K.; Amaya, F.; Vardeh, D.; Shi, L.; Brenner, G.J.; Ji, R.R.; Bean, B.P.; Woolf, C.J.; et al. Nociceptors are interleukin-1beta sensors. J. Neurosci. 2008, 28, 14062–14073. [Google Scholar] [CrossRef] [PubMed]
- Dones, I.; Levi, V. Spinal Cord Stimulation for Neuropathic Pain: Current Trends and Future Applications. Brain Sci. 2018, 8, 138. [Google Scholar] [CrossRef] [PubMed]
- Caylor, J.; Reddy, R.; Yin, S.; Cui, C.; Huang, M.; Huang, C.; Rao, R.; Baker, D.G.; Simmons, A.; Souza, D.; et al. Spinal cord stimulation in chronic pain: Evidence and theory for mechanisms of action. Bioelectron. Med. 2019, 5, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Joosten, E.A.; Franken, G. Spinal cord stimulation in chronic neuropathic pain: Mechanisms of action, new locations, new paradigms. Pain 2020, 161, S104–S113. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.J.; Lempka, S.F.; Machado, A.G. Percutaneous spinal cord stimulation for chronic pain: Indications and patient selection. Neurosurg. Clin. 2014, 25, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Fornasari, D. Pharmacotherapy for Neuropathic Pain: A Review. Pain Ther. 2017, 6, 25–33. [Google Scholar] [CrossRef]
- Kremer, M.; Salvat, E.; Muller, A.; Yalcin, I.; Barrot, M. Antidepressants and gabapentinoids in neuropathic pain: Mechanistic insights. Neuroscience 2016, 338, 183–206. [Google Scholar] [CrossRef]
- Stahl, S.M.; Porreca, F.; Taylor, C.P.; Cheung, R.; Thorpe, A.J.; Clair, A. The diverse therapeutic actions of pregabalin: Is a single mechanism responsible for several pharmacological activities? Trends Pharmacol. Sci. 2013, 34, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Baftiu, A.; Landmark, C.J.; Rusten, I.R.; Feet, S.A.; Johannessen, S.I.; Larsson, P.G. Changes in utilisation of antiepileptic drugs in epilepsy and non-epilepsy disorders—a pharmacoepidemiological study and clinical implications. Eur. J. Clin. Pharmacol. 2016, 72, 1245–1254. [Google Scholar] [CrossRef] [PubMed]
- Finnerup, N.B.; Attal, N.; Haroutounian, S.; McNicol, E.; Baron, R.; Dworkin, R.H.; Gilron, I.; Haanpää, M.; Hansson, P.; Jensen, T.S.; et al. Pharmacotherapy for neuropathic pain in adults: A systematic review and meta-analysis. Lancet Neurol. 2015, 14, 162–173. [Google Scholar] [CrossRef]
- Crawford, P.F., III; Xu, Y. Topical capsaicin for treatment of chronic neuropathic pain in adults. Am. Fam. Physician 2017, 96. [Google Scholar] [CrossRef]
- Deng, Y.; Luo, L.; Hu, Y.; Fang, K.; Liu, J. Clinical practice guidelines for the management of neuropathic pain: A systematic review. BMC Anesthesiol. 2015, 16, 12. [Google Scholar] [CrossRef] [PubMed]
- Mensah-Nyagan, A.G.; Meyer, L.; Schaeffer, V.; Kibaly, C.; Patte-Mensah, C. Evidence for a key role of steroids in the modulation of pain. Psychoneuroendocrinology 2009, 34, S169–S177. [Google Scholar] [CrossRef]
- Vyvey, M. Steroids as pain relief adjuvants. Can. Fam. Physician Med. Fam. Can. 2010, 56, 1295–1297. [Google Scholar]
- Otis, J.A.D.; Macone, A. Neuropathic Pain. Semin. Neurol. 2018, 38, 644–653. [Google Scholar] [CrossRef]
- Moisset, X.; De Andrade, D.; Bouhassira, D. From pulses to pain relief: An update on the mechanisms of rTMS-induced analgesic effects. Eur. J. Pain 2016, 20, 689–700. [Google Scholar] [CrossRef]
- Siniscalco, D.; Rossi, F.; Maione, S. Molecular approaches for neuropathic pain treatment. Curr. Med. Chem. 2007, 14, 1783–1787. [Google Scholar] [CrossRef]
- Siniscalco, D. Transplantation of human mesenchymal stem cells in the study of neuropathic pain. In Analgesia; Springer: Berlin/Heidelberg, Germany, 2010; pp. 337–345. [Google Scholar]
- Bossio, C.; Mastrangelo, R.; Morini, R.; Tonna, N.; Coco, S.; Verderio, C.; Matteoli, M.; Bianco, F. A Simple Method to Generate Adipose Stem Cell-Derived Neurons for Screening Purposes. J. Mol. Neurosci. 2013, 51, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Kingham, P.J.; Kalbermatten, D.F.; Mahay, D.; Armstrong, S.J.; Wiberg, M.; Terenghi, G. Adipose-derived stem cells differentiate into a Schwann cell phenotype and promote neurite outgrowth in vitro. Exp. Neurol. 2007, 207, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Franchi, S.; Castelli, M.; Amodeo, G.; Niada, S.; Ferrari, D.; Vescovi, A.L.; Brini, A.T.; Panerai, A.E.; Sacerdote, P. Adult Stem Cell as New Advanced Therapy for Experimental Neuropathic Pain Treatment. Biomed Res. Int. 2014, 2014, 1–10. [Google Scholar] [CrossRef]
- Hu, C.-D.; Wang, Y.; Wang, C.; Lu, C.-F.; Peng, J. Roles of neural stem cells in the repair of peripheral nerve injury. Neural Regen. Res. 2017, 12, 2106–2112. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.-C.; Du, X.-J.; Chen, Y.-X.; Wang, N.; Wang, X.-Y.; Kong, F.-E. Neural stem cell transplantation inhibits glial cell proliferation and P2X receptor-mediated neuropathic pain in spinal cord injury rats. Neural Regen. Res. 2019, 14, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, D.; Binda, E.; De Filippis, L.; Vescovi, A.L. Isolation of Neural Stem Cells from Neural Tissues Using the Neurosphere Technique. Curr. Protoc. Stem Cell Biol. 2010, 15, 2D.6.1–2D.6.18. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.-G.; Sun, X.-L.; Li, P.; Liu, H.-L.; Wu, H.-L.; Xi, Z.-Q.; Zheng, Z.-H. Neural stem cells transplantation alleviate the hyperalgesia of spinal cord injured (SCI) associated with down-regulation of BDNF. Int. J. Clin. Exp. Med. 2015, 8, 404–412. [Google Scholar]
- Nardi, N.B.; Meirelles, L.D.S. Mesenchymal stem cells: Isolation, in vitro expansion and characterization. Handb. Exp. Pharmacol. 2006, 174, 249–282. [Google Scholar]
- Sethe, S.; Scutt, A.; Stolzing, A. Aging of mesenchymal stem cells. Ageing Res. Rev. 2006, 5, 91–116. [Google Scholar] [CrossRef] [PubMed]
- Musina, R.A.; Bekchanova, E.S.; Sukhikh, G.T. Comparison of Mesenchymal Stem Cells Obtained from Different Human Tissues. Bull. Exp. Biol. Med. 2005, 139, 504–509. [Google Scholar] [CrossRef]
- Alizadeh, R.; Bagher, Z.; Kamrava, S.K.; Falah, M.; Hamidabadi, H.G.; Boroujeni, M.E.; Mohammadi, F.; Khodaverdi, S.; Zare-Sadeghi, A.; Olya, A.; et al. Differentiation of human mesenchymal stem cells (MSC) to dopaminergic neurons: A comparison between Wharton’s Jelly and olfactory mucosa as sources of MSCs. J. Chem. Neuroanat. 2019, 96, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Giordano, A.; Galderisi, U.; Marino, I.R. From the laboratory bench to the patient’s bedside: An update on clinical trials with mesenchymal stem cells. J. Cell. Physiol. 2007, 211, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Pittenger, M. Mesenchymal stem cells: Progress toward promise. Cytotherapy 2005, 7, 36–45. [Google Scholar] [CrossRef]
- Musolino, P.L.; Coronel, M.F.; Hökfelt, T.; Villar, M.J. Bone marrow stromal cells induce changes in pain behavior after sciatic nerve constriction. Neurosci. Lett. 2007, 418, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Magistretti, P.J.; Allaman, I. A Cellular Perspective on Brain Energy Metabolism and Functional Imaging. Neuron 2015, 86, 883–901. [Google Scholar] [CrossRef] [PubMed]
- Henriques, D.; Moreira, R.; Schwamborn, J.; De Almeida, L.P.; Mendonça, L.S. Successes and Hurdles in Stem Cells Application and Production for Brain Transplantation. Front. Neurosci. 2019, 13, 1194. [Google Scholar] [CrossRef]
- Phinney, D.G.; Pittenger, M.F. Concise review: MSC-derived exosomes for cell-free therapy. Stem Cells 2017, 35, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Eirin, A.; Riester, S.M.; Zhu, X.-Y.; Tang, H.; Evans, J.M.; O’Brien, D.; van Wijnen, A.J.; Lerman, L.O. MicroRNA and mRNA cargo of extracellular vesicles from porcine adipose tissue-derived mesenchymal stem cells. Gene 2014, 551, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chopp, M.; Liu, X.S.; Katakowski, M.; Wang, X.; Tian, X.; Wu, D.; Zhang, Z.G. Exosomes Derived from Mesenchymal Stromal Cells Promote Axonal Growth of Cortical Neurons. Mol. Neurobiol. 2017, 54, 2659–2673. [Google Scholar] [CrossRef]
- Zhao, W.; Beers, D.R.; Appel, S.H. Immune-mediated Mechanisms in the Pathoprogression of Amyotrophic Lateral Sclerosis. J. Neuroimmune Pharmacol. 2013, 8, 888–899. [Google Scholar] [CrossRef] [PubMed]
- Coronel, M.; Musolino, P.; Brumovsky, P.; Hokfelt, T.; Villar, M. Bone marrow stromal cells attenuate injury-induced changes in galanin, NPY and NPY Y1-receptor expression after a sciatic nerve constriction. Neuropeptides 2009, 43, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Siniscalco, D.; Giordano, C.; Galderisi, U.; Luongo, L.; De Novellis, V.; Rossi, F.; Maione, S. Long-Lasting Effects of Human Mesenchymal Stem Cell Systemic Administration on Pain-Like Behaviors, Cellular, and Biomolecular Modifications in Neuropathic Mice. Front. Integr. Neurosci. 2011, 5, 79. [Google Scholar] [CrossRef] [PubMed]
- Sacerdote, P.; Niada, S.; Franchi, S.; Arrigoni, E.; Rossi, A.; Yenagi, V.; De Girolamo, L.; Panerai, A.E.; Brini, A.T. Systemic Administration of Human Adipose-Derived Stem Cells Reverts Nociceptive Hypersensitivity in an Experimental Model of Neuropathy. Stem Cells Dev. 2013, 22, 1252–1263. [Google Scholar] [CrossRef] [PubMed]
- Franchi, S.; Valsecchi, A.E.; Borsani, E.; Procacci, P.; Ferrari, D.; Zaffa, C.; Sartori, P.; Rodella, L.F.; Vescovi, A.; Maione, S.; et al. Intravenous neural stem cells abolish nociceptive hypersensitivity and trigger nerve regeneration in experimental neuropathy. Pain 2012, 153, 850–861. [Google Scholar] [CrossRef]
- Choi, J.I.; Cho, H.T.; Jee, M.K.; Kang, S.K. Core-shell nanoparticle controlled hATSCs neurogenesis for neuropathic pain therapy. Biomaterials 2013, 34, 4956–4970. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Park, E.-M.; Kim, B.J.; Kim, J.-S.; Choi, B.; Lee, S.-H.; Han, I. Transplantation of human Wharton’s jelly-derived mesenchymal stem cells highly expressing TGFβ receptors in a rabbit model of disc degeneration. Stem Cell Res. Ther. 2015, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Klass, M.; Gavrikov, V.; Drury, D.; Stewart, B.; Hunter, S.; Denson, D.D.; Hord, A.; Csete, M. Intravenous Mononuclear Marrow Cells Reverse Neuropathic Pain from Experimental Mononeuropathy. Anesth. Analg. 2007, 104, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Naruse, K.; Sato, J.; Funakubo, M.; Hata, M.; Nakamura, N.; Kobayashi, Y.; Kamiya, H.; Shibata, T.; Kondo, M.; Himeno, T.; et al. Transplantation of Bone Marrow-Derived Mononuclear Cells Improves Mechanical Hyperalgesia, Cold Allodynia and Nerve Function in Diabetic Neuropathy. PLoS ONE 2011, 6, e27458. [Google Scholar] [CrossRef]
- Meacham, K.; Shepherd, A.; Mohapatra, D.P.; Haroutounian, S.J.C. Neuropathic pain: Central vs. peripheral mechanisms. Curr. Pain Headache Rep. 2017, 21, 28. [Google Scholar] [CrossRef]
- Sommer, C.; Leinders, M.; Üçeyler, N. Inflammation in the pathophysiology of neuropathic pain. Pain 2018, 159, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Harrell, C.R.; Jankovic, M.G.; Fellabaum, C.; Volarevic, A.; Djonov, V.; Arsenijevic, A.; Volarevic, V. Molecular mechanisms responsible for anti-inflammatory and immunosuppressive effects of mesenchymal stem cell-derived factors. In Tissue Engineering and Regenerative Medicine; Springer: Berlin/Heidelberg, Germany, 2018; pp. 187–206. [Google Scholar]
- Takizawa, N.; Okubo, N.; Kamo, M.; Chosa, N.; Mikami, T.; Suzuki, K.; Yokota, S.; Ibi, M.; Ohtsuka, M.; Taira, M.; et al. Bone marrow-derived mesenchymal stem cells propagate immunosuppressive/anti-inflammatory macrophages in cell-to-cell contact-independent and -dependent manners under hypoxic culture. Exp. Cell Res. 2017, 358, 411–420. [Google Scholar] [CrossRef]
- Mert, T.; Kurt, A.H.; Altun, İ.; Celik, A.; Baran, F.; Gunay, I. Pulsed magnetic field enhances therapeutic efficiency of mesenchymal stem cells in chronic neuropathic pain model. Bioelectromagnetics 2017, 38, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Omi, M.; Hata, M.; Nakamura, N.; Miyabe, M.; Kobayashi, Y.; Kamiya, H.; Nakamura, J.; Ozawa, S.; Tanaka, Y.; Takebe, J.; et al. Transplantation of dental pulp stem cells suppressed inflammation in sciatic nerves by promoting macrophage polarization towards anti-inflammation phenotypes and ameliorated diabetic polyneuropathy. J. Diabetes Investig. 2016, 7, 485–496. [Google Scholar] [CrossRef]
- Liu, M.; Li, K.; Wang, Y.; Zhao, G.; Jiang, J. Stem Cells in the Treatment of Neuropathic Pain: Research Progress of Mechanism. Stem Cells Int. 2020, 2020, 1–13. [Google Scholar] [CrossRef]
- Xie, J.; Ren, J.; Liu, N.; Wu, C.; Xiao, D.; Luo, H.; Du, J. Pretreatment with AM1241 Enhances the Analgesic Effect of Intrathecally Administrated Mesenchymal Stem Cells. Stem Cells Int. 2019, 2019, 7025473-17. [Google Scholar] [CrossRef] [PubMed]
- Kamp, J.; Van Velzen, M.; Olofsen, E.; Boon, M.; Dahan, A.; Niesters, M. Pharmacokinetic and pharmacodynamic considerations for NMDA-receptor antagonist ketamine in the treatment of chronic neuropathic pain: An update of the most recent literature. Expert Opin. Drug Metab. Toxicol. 2019, 15, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, C.; Peng, Y.; Wang, Y.; Xu, H.; Liu, C. ROR2 modulates neuropathic pain via phosphorylation of NMDA receptor subunit GluN2B in rats. Br. J. Anaesth. 2019, 123, e239–e248. [Google Scholar] [CrossRef]
- Chen, J.; Li, L.; Chen, S.-R.; Chen, H.; Xie, J.-D.; Sirrieh, R.E.; MacLean, D.M.; Zhang, Y.; Zhou, M.-H.; Jayaraman, V.; et al. The α2δ-1-NMDA Receptor Complex Is Critically Involved in Neuropathic Pain Development and Gabapentin Therapeutic Actions. Cell Rep. 2018, 22, 2307–2321. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.P.; Mao, J. Neuropathic pain: Mechanisms and their clinical implications. BMJ 2014, 348, f7656. [Google Scholar] [CrossRef]
- Guo, W.; Chu, Y.-X.; Imai, S.; Yang, J.-L.; Zou, S.; Mohammad, Z.; Wei, F.; Dubner, R.; Ren, K. Further observations on the behavioral and neural effects of bone marrow stromal cells in rodent pain models. Mol. Pain 2016, 12, 1744806916658043. [Google Scholar] [CrossRef]
- Chen, N.-F.; Huang, S.-Y.; Chen, W.-F.; Chen, C.-H.; Lu, C.-H.; Yang, S.-N.; Wang, H.-M.; Wen, Z.-H.; Chen, C.-L. TGF-β1 Attenuates Spinal Neuroinflammation and the Excitatory Amino Acid System in Rats with Neuropathic Pain. J. Pain 2013, 14, 1671–1685. [Google Scholar] [CrossRef]
- Chen, G.; Park, C.-K.; Xie, R.-G.; Ji, R.-R. Intrathecal bone marrow stromal cells inhibit neuropathic pain via TGF-β secretion. J. Clin. Investig. 2015, 125, 3226–3240. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M. Modulation of pain and itch by spinal glia. Neurosci. Bull. 2018, 34, 178–185. [Google Scholar] [CrossRef]
- Mika, J.; Zychowska, M.; Popiolek-Barczyk, K.; Rojewska, E.; Przewlocka, B. Importance of glial activation in neuropathic pain. Eur. J. Pharmacol. 2013, 716, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Forouzanfar, F.; Amin, B.; Ghorbani, A.; Ghazavi, H.; Ghasemi, F.; Sadri, K.; Mehri, S.; Sadeghnia, H.; Hosseinzadeh, H. New approach for the treatment of neuropathic pain: Fibroblast growth factor 1 gene-transfected adipose-derived mesenchymal stem cells. Eur. J. Pain 2018, 22, 295–310. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wang, W.; Liu, X.; Xi, Y.; Yu, J.; Yang, X.; Ye, X. Bone mesenchymal stem cells attenuate radicular pain by inhibiting microglial activation in a rat noncompressive disk herniation model. Cell Tissue Res. 2018, 374, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.-R.; Gereau, R.W., IV; Malcangio, M.; Strichartz, G. MAP kinase and pain. Brain Res. Rev. 2009, 60, 135–148. [Google Scholar] [CrossRef]
- Watanabe, S.; Uchida, K.; Nakajima, H.; Matsuo, H.; Sugita, D.; Yoshida, A.; Honjoh, K.; Johnson, W.E.; Baba, H. Early Transplantation of Mesenchymal Stem Cells After Spinal Cord Injury Relieves Pain Hypersensitivity Through Suppression of Pain-Related Signaling Cascades and Reduced Inflammatory Cell Recruitment. Stem Cells 2015, 33, 1902–1914. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Deng, G.; Wang, H.; Yang, M.; Yang, R.; Li, X.; Zhang, X.; Yuan, H. Interleukin-1β pre-treated bone marrow stromal cells alleviate neuropathic pain through CCL7-mediated inhibition of microglial activation in the spinal cord. Sci. Rep. 2017, 7, 42260. [Google Scholar] [CrossRef] [PubMed]
- Kanao-Kanda, M.; Kanda, H.; Liu, M.S.; Roy, S.; Toborek, M.; Hao, S. Viral Vector–Mediated Gene Transfer of Glutamic Acid Decarboxylase for Chronic Pain Treatment: A Literature Review. Hum. Gene Ther. 2020, 31, 405–414. [Google Scholar] [CrossRef]
- Chattopadhyay, M.; Mata, M.; Finkl, D.J. Vector-mediated release of GABA attenuates pain-related behaviors and reduces NaV 1.7 in DRG neurons. Eur. J. Pain 2011, 15, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Asahara, T.; Kalka, C.; Isner, J.M. Stem cell therapy and gene transfer for regeneration. Gene Ther. 2000, 7, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.G.A.; Adam, M.; Miller, A.D. Gene transfer by retrovirus vectors occurs only in cells that are actively replicating at the time of infection. Mol. Cell. Biol. 1990, 10, 4239–4242. [Google Scholar] [CrossRef]
- Naldini, L.; Blömer, U.; Gallay, P.; Ory, D.; Mulligan, R.; Gage, F.H.; Verma, I.M.; Trono, D. In Vivo Gene Delivery and Stable Transduction of Nondividing Cells by a Lentiviral Vector. Science 1996, 272, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Goodman, S.; Xiao, X.; Donahue, R.E.; Moulton, A.; Miller, J.; Walsh, C.; Young, N.S.; Samulski, R.J.; Nienhuis, A.W. Recombinant adeno-associated virus-mediated gene transfer into hematopoietic progenitor cells. Blood 1994, 84, 1492–1500, Erratum in 1995, 85, 862. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Kuszynski, C.; Ino, K.; Heimann, D.; Shepard, H.; Yasui, Y.; Maneval, D.; Talmadge, J. Gene transfer into human bone marrow hematopoietic cells mediated by adenovirus vectors [see comments]. Blood 1996, 87, 5032–5039. [Google Scholar] [CrossRef] [PubMed]
- Persons, D.A.; Allay, J.A.; Riberdy, J.M.; Wersto, R.P.; Donahue, R.E.; Sorrentino, B.P.; Nienhuis, A.W. Use of the green fluorescent protein as a marker to identify and track genetically modified hematopoietic cells. Nat. Med. 1998, 4, 1201–1205. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, B.; Brandt, S.; Bodine, D.; Gottesman, M.; Pastan, I.; Cline, A.; Nienhuis, A. Selection of drug-resistant bone marrow cells in vivo after retroviral transfer of human MDR1. Science 1992, 257, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Cova, L.; Armentero, M.-T.; Zennaro, E.; Calzarossa, C.; Bossolasco, P.; Busca, G.; Deliliers, G.L.; Polli, E.; Nappi, G.; Silani, V. Multiple neurogenic and neurorescue effects of human mesenchymal stem cell after transplantation in an experimental model of Parkinson’s disease. Brain Res. 2010, 1311, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Lee, P.H.; Bang, O.Y.; Lee, G.; Ahn, Y.H. Mesenchymal stem cells therapy exerts neuroprotection in a progressive animal model of Parkinson’s disease. J. Neurochem. 2008, 107, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yasuhara, T.; Shingo, T.; Kameda, M.; Tajiri, N.; Yuan, W.J.; Kondo, A.; Kadota, T.; Baba, T.; Tayra, J.T.; et al. Intravenous administration of mesenchymal stem cells exerts therapeutic effects on parkinsonian model of rats: Focusing on neuroprotective effects of stromal cell-derived factor-1α. BMC Neurosci. 2010, 11, 52. [Google Scholar] [CrossRef] [PubMed]
- Levi, A.D.; Okonkwo, D.O.; Park, P.; Jenkins, A.L., 3rd; Kurpad, S.N.; Parr, A.M.; Ganju, A.; Aarabi, B.; Kim, D.; Casha, S.; et al. Emerging Safety of Intramedullary Transplantation of Human Neural Stem Cells in Chronic Cervical and Thoracic Spinal Cord Injury. Neurosurgery 2018, 82, 562–575. [Google Scholar] [CrossRef] [PubMed]
- Van Pham, P. Clinical trials for stem cell transplantation: When are they needed? Stem Cell Res. Ther. 2016, 7, 65. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, M.V.P.; LaRocca, T.F.; Souza, B.S.D.F.; Villarreal, C.F.; Silva, L.F.M.; Matos, A.C.; Novaes, M.A.; Bahia, C.M.P.; Martinez, A.C.D.O.M.; Kaneto, C.M.; et al. Safety and neurological assessments after autologous transplantation of bone marrow mesenchymal stem cells in subjects with chronic spinal cord injury. Stem Cell Res. Ther. 2014, 5, 126. [Google Scholar] [CrossRef]
- Vaquero, J.; Zurita, M.; Rico, M.; Aguayo, C.; Fernández, C.; Gutiérrez, R.; Rodríguez-Boto, G.; Saab, A.; Hassan, R.; Ortega, C. Intrathecal administration of autologous bone marrow stromal cells improves neuropathic pain in patients with spinal cord injury. Neurosci. Lett. 2018, 670, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Vickers, E.R.; Karsten, E.; Lilischkis, R.; Flood, J. A preliminary report on stem cell therapy for neuropathic pain in humans. J. Pain Res. 2014, 7, 255–263. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Top (1st) Level Diagnosis | Chronicneuropathic Pain | |
|---|---|---|
| 2nd level diagnosis | Chronic peripheralneuropathic pain | Chronic central |
| neuropathic pain | ||
| 3rd level diagnosis | Trigeminal neuralgia Painful polyneuropathy | Chronic central neuropathic pain associated with spinal cord injury |
| Chronic neuropathic pain after peripheral nerve injury | Chronic central | |
| neuropathic pain associated with brain injury | ||
| Postherpetic neuralgia | Chronic central poststroke pain | |
| Painful radiculopathy | Chronic central | |
| neuropathic pain associated with multiple sclerosis | ||
| Multiple parents | Chronic posttraumatic pain | Chronic posttraumatic pain |
| Chronic secondary headaches and oro-facial pain | ||
| Etiology | Typical Syndrome (Example) |
|---|---|
| Mechanical | Carpal tunnel syndrome, Postsurgical pain, |
| Painful radiculopathy, Cancer pain, Phantom limb pain | |
| Metabolic/ischemic | Diabetic polyneuropathy, Vitamin B12 deficiency |
| Inflammatory | Postherpetic neuralgia, HIV neuropathy, |
| Leprosy, Guillain-Barré Syndrome, | |
| Critical illness polyneuropathy | |
| Neurotoxic | Chemotherapy-induced, peripheral neuropathy, Alcoholic neuropathy |
| Radiation | Postradiation neuropathy |
| Hereditary | Charcot-Marie-Tooth disease, Fabry disease |
| Classical Pharmacological Treatment of Neuropathic Pain | |||||
|---|---|---|---|---|---|
| Drug Class | Types of Neuropathic Pain | Effects | Side Effects | References | |
| Antidepressants | Tricyclic antidepressants (TCAs): | Diabetic neuropathy | Inhibition of serotonin and noradrenaline reuptake at synapses between nociceptors and spinothalamic neurons | Sedation | [39,40] |
| amitriptyline, nortriptyline, | Postherpetic neuralgia | Constipation | |||
| desipramine, imipramine; | Poststroke pain | Weight gain | |||
| Serotonin-norepinephrine | Painful polyneuropathy | Dry mouth | |||
| reuptake inhibitors (SNRIs): | Lower back pain | Nausea | |||
| duloxetine, venlafaxine | |||||
| Anticonvulsants | Phenytoin | Lancinating pain and allodynia | Reduction of neuronal excitability and local neuronal discharges, acting through sodium channel blockade or modulation of calcium channels | Dizziness, Skin reaction (e.g., Steven-Johnson syndrome), Leukopenia | [41,42] |
| Gabapentin | Painful diabetic neuropathy | ||||
| Carbamazepine | Trigeminal neuralgia | ||||
| Oxcarbazepine | Postherpetic neuralgia | ||||
| Valproic acid | Painful polyneuropathy | ||||
| Lower back pain | |||||
| Topical agents | Lidocaine | Allodynia | Blockade of voltage-gated sodium channels expressed by nerve fibers, responsible for the propagation of action potential. | Local irritation Possible hypersensitivity | |
| Capsaicin | Postherpetic neuralgia | [43,44] | |||
| Clonidine | Chemotherapy-induced peripheral neuropathy | ||||
| EMLA (eutectic mixture of local anesthetic) | Postsurgical and post-traumatic neuropathic pain | ||||
| Opioids | Morphine | Diabetic peripheral neuropathy | Opioid receptors are coupled to calcium and potassium channels, block synaptic transmission, restricting the number of stimuli | Drowsiness | [39,45] |
| Hydromorphine | Postherpetic neuropathy | Nausea | |||
| Tramadol | Polyneuropathy | Dependence overdoses | |||
| Oxycodone | Phantom limb pain | ||||
| Corticosteroids | Prednisolone | Allodynia | Inhibition of prostaglandin synthesis, reduction of inflammation, vascular permeability, and tissue edema | Gastrointestinal disease | [46,47] |
| Dexamethasone | Spinal cord compression | psychiatric disorders | |||
| Postherpetic neuralgia | electrolyte imbalances | ||||
| Bone demineralization | |||||
| Alternative nonpharmacological therapies | Acupuncture | Chemotherapy-induced peripheral neuropathy | Local inhibition of nociceptive fibers, stimulates blood flow to restore nerve damage | [48,49] | |
| Magnetic insoles | Trigeminal neuralgia | Bruising | |||
| Repetitive transcranial magnetic stimulation (rTMS) | Poststroke pain | Infection | |||
| Postherpetic pain | |||||
| Nonpharmacological/Alternative Treatment of Neuropathic Pain: Spinal Cord Stimulation (SCS) | |||||
| SCS Methods | Types of Neuropathic Pain | SCS Main Contraindications | SCS Common Complications | References | |
| Spinal cord stimulation (SCS) | Tonic spinal cord stimulation and suprasegmental mechanisms New stimulation location: Dorsal root ganglion High-frequency spinal cord stimulation in neuropathic pain Burst spinal cord stimulation in neuropathic pain | Failed back surgery syndrome Complex regional pain syndrome (I and II) Radicular and nerve root pain Postherpetic neuralgia Pain due to peripheral nerve injury Intercostal neuralgia Phantom pain | Infection Coagulopathy Spinal stenosis Psychiatric disorders Substance abuse | More frequent: Hardware-related (lead migration, breakage, connection failure, malfunctioning, pain at the implantable pulse generator site) Hematoma and seroma at implantable pulse generator site Rare: Spinal epidural hematoma, CSF leakNeurological deficit | [35,36,37] |
| Cell Type | Model of NP | Advantages | Limitations | Reference |
|---|---|---|---|---|
| Neural stem cells (NSCs) | CCI | extensive self-renewal capacity | low stem cell transplant efficiency | [55,56] |
| SCI (rats) | ||||
| Mesenchymal stem cells (MSCs) | SNI (mice) | strong immunosuppressive properties; long lasting therapy | impossibility to predict which site the injected MSCs to be trapped | [61,62,66] |
| Bone marrow stromal cells (BMScs) | SLNC (rats) | down regulation of glial and microglial activation and proinflammatory cytokines | some limited analgesic effect | [65,72] |
| SNI (mice) | ||||
| Bone marrow-derived mononuclear cells (BM-MNCs) | CCI (rats) | functional recovery of the peripheral nerve followed by increased nerve blood flow | may cause neuronal apoptosis | [78,79] |
| Diabetic neuropathy (rats) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joshi, H.P.; Jo, H.-J.; Kim, Y.-H.; An, S.-B.; Park, C.-K.; Han, I. Stem Cell Therapy for Modulating Neuroinflammation in Neuropathic Pain. Int. J. Mol. Sci. 2021, 22, 4853. https://doi.org/10.3390/ijms22094853
Joshi HP, Jo H-J, Kim Y-H, An S-B, Park C-K, Han I. Stem Cell Therapy for Modulating Neuroinflammation in Neuropathic Pain. International Journal of Molecular Sciences. 2021; 22(9):4853. https://doi.org/10.3390/ijms22094853
Chicago/Turabian StyleJoshi, Hari Prasad, Hyun-Jung Jo, Yong-Ho Kim, Seong-Bae An, Chul-Kyu Park, and Inbo Han. 2021. "Stem Cell Therapy for Modulating Neuroinflammation in Neuropathic Pain" International Journal of Molecular Sciences 22, no. 9: 4853. https://doi.org/10.3390/ijms22094853
APA StyleJoshi, H. P., Jo, H.-J., Kim, Y.-H., An, S.-B., Park, C.-K., & Han, I. (2021). Stem Cell Therapy for Modulating Neuroinflammation in Neuropathic Pain. International Journal of Molecular Sciences, 22(9), 4853. https://doi.org/10.3390/ijms22094853