
Abundant Synthesis of Netrin-1 in Satellite Cell-Derived Myoblasts Isolated from EDL Rather Than Soleus Muscle Regulates Fast-Type Myotube Formation

 ,
,
Abstract
1. Introduction
2. Results
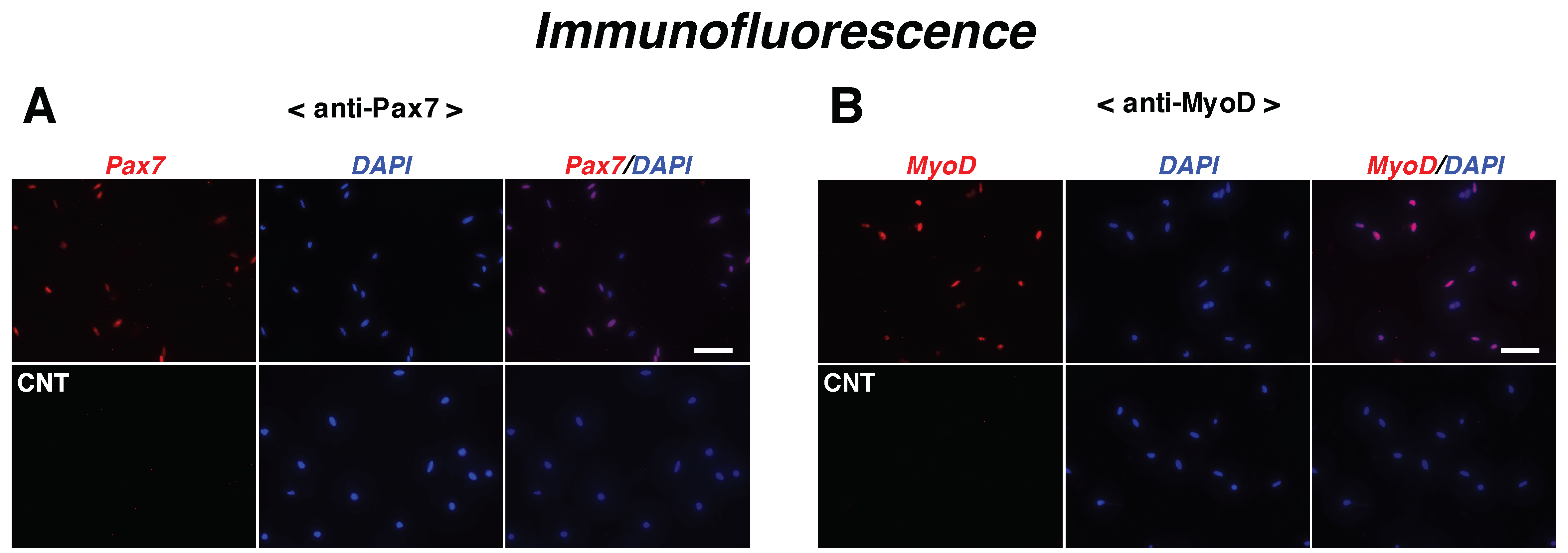
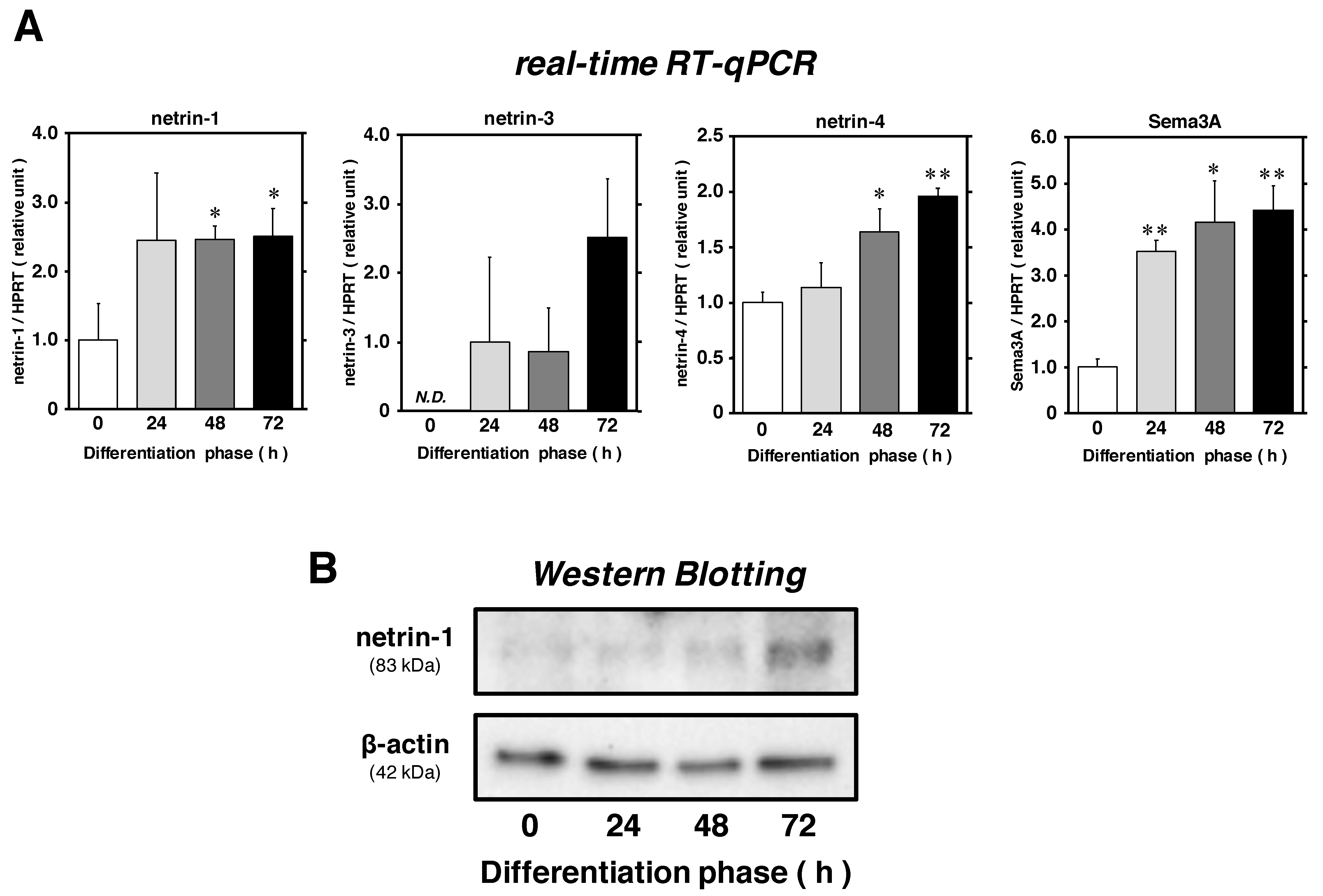
2.1. Netrin-1 and -4 Expression Levels Are Upregulated in Satellite Cells during Myogenic Differentiation
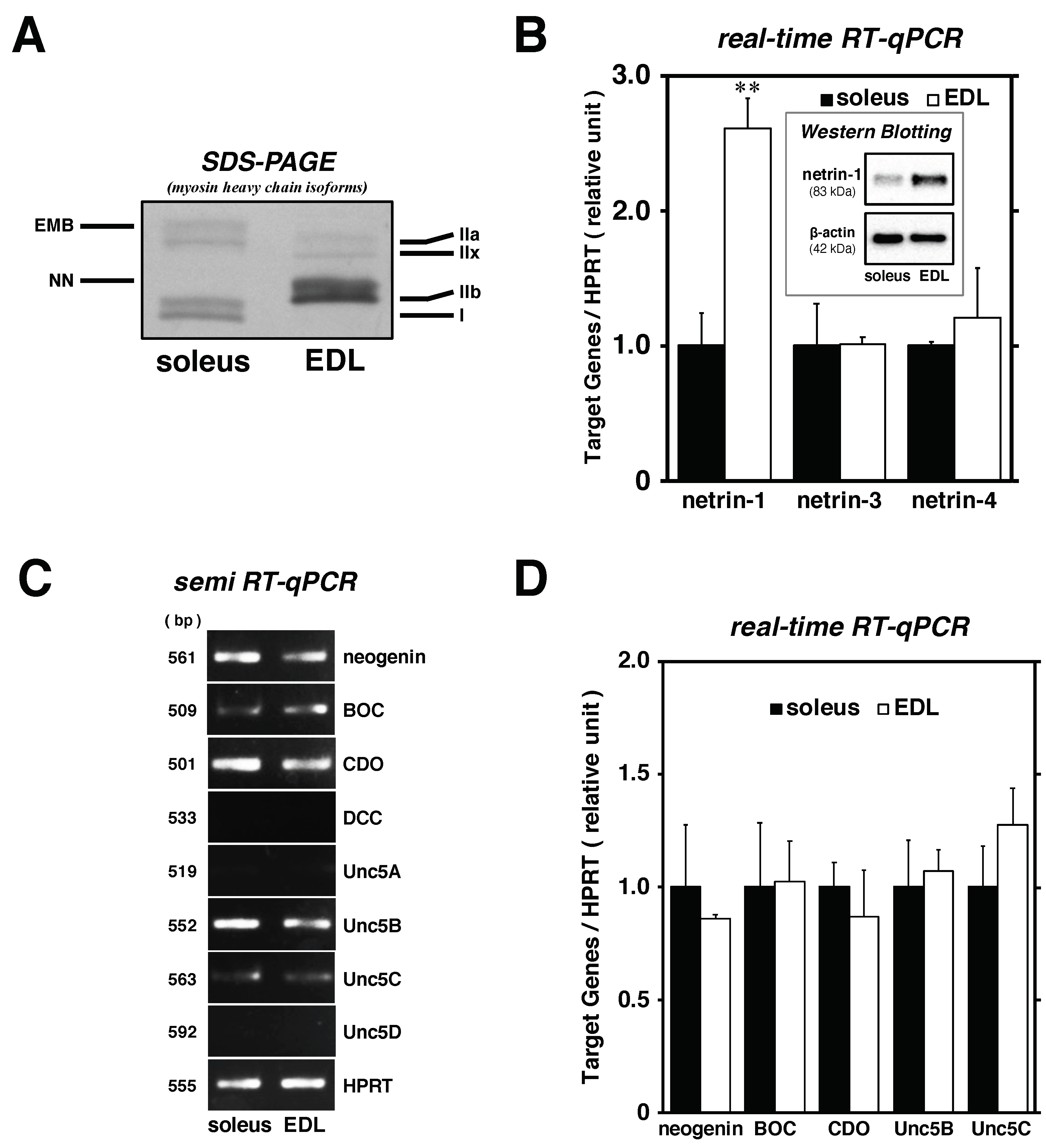
2.2. Satellite Cells Isolated from EDL Synthesize Higher Levels of Netrin-1 Than Those from Soleus
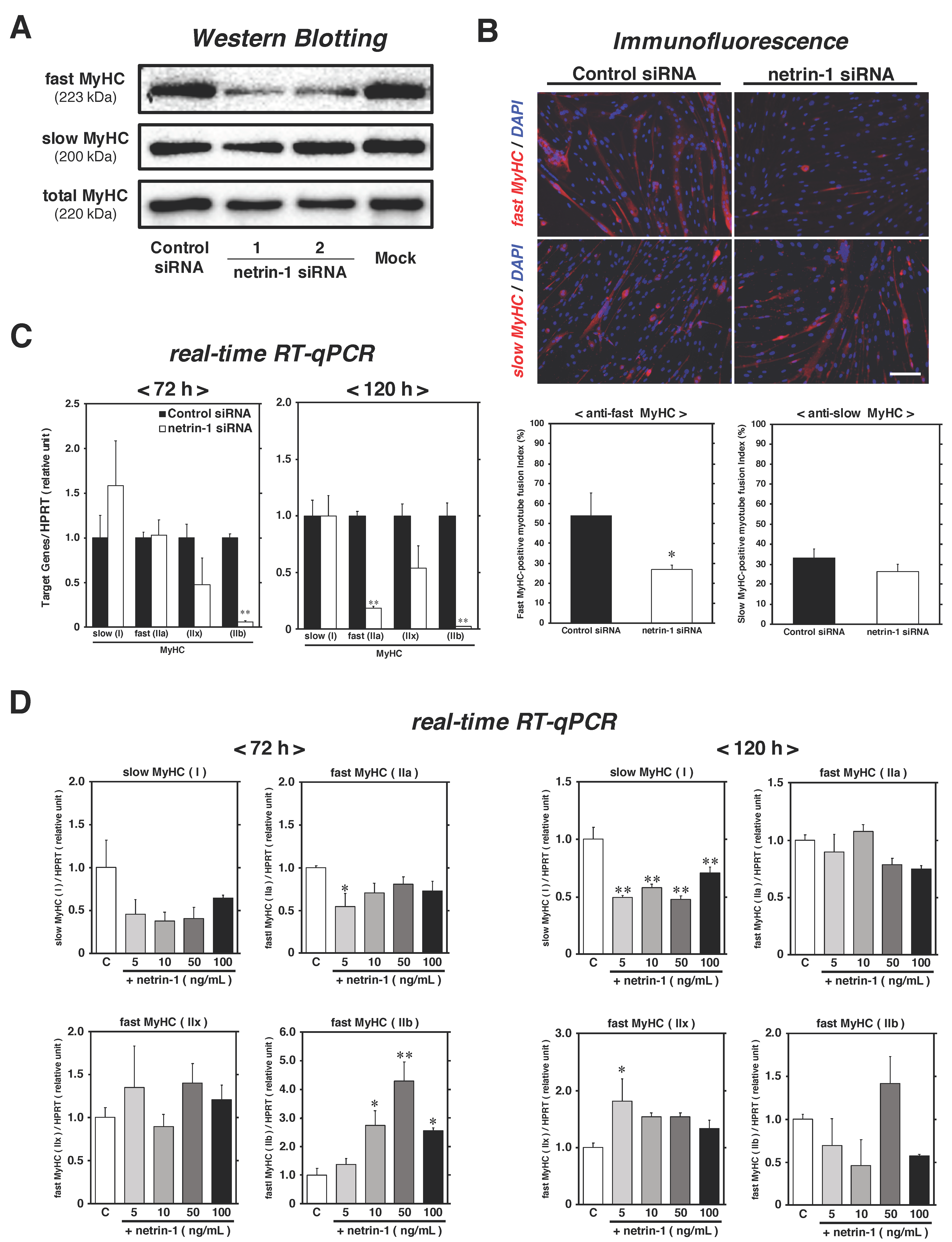
2.3. Netrin-1 Specifically Affects the Expression of Fast-MyHC during Myotube Formation Process
3. Discussion
4. Materials and Methods
4.1. Animal Care and Use
4.2. Satellite Cell Isolation and Primary Cultures
4.3. Knockdown of Netrin-1 Expression by siRNA Transfection in Myoblasts
4.4. Immunocytochemistry of Satellite Cells and Myotubes
4.5. SDS-PAGE for MyHC Isoform Composition
4.6. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.7. Enhanced Chemiluminescence (ECL) and Western Blotting
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EDL | extensor digitorum longus |
| PPAR-delta | peroxisome proliferator-activated receptor-delta |
| NFAT | nuclear factor of activated T-cells |
| miRNA | micro RNA |
| Sema3A | semaphorin 3A |
| DCC | deleted in colorectal cancer |
| CDO | cell adhesion molecule-related/down-regulated by oncogenes |
| BOC | bioregional cell adhesion molecule-related/down-regulated by oncogenes binding protein |
| MyHC | myosin heavy chain |
References
- Schiaffino, S.; Reggiani, C. Fiber types in Mammalian skeletal muscles. Physiol. Rev. 2011, 91, 1447–1531. [Google Scholar] [CrossRef] [PubMed]
- Garnett, R.A.; O’Donovan, M.J.; Stephens, J.A.; Taylor, A. Motor unit organization of human medial gastrocnemius. J. Physiol. 1979, 287, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Handschin, C.; Spiegelman, B.M. Metabolic control through the PGC-1 family of transcription coactivators. Cell Metab. 2005, 1, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Mauro, A. Satellite cell of skeletal muscle fibers. J. Biophys. Biochem. Cytol. 1961, 9, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Price, F.; Rudnicki, M.A. Satellite cells and the muscle stem cell niche. Physiol. Rev. 2013, 93, 23–67. [Google Scholar] [CrossRef] [PubMed]
- Lagord, C.; Soulet, L.; Bonavaud, S.; Bassaglia, Y.; Rey, C.; Barlovatz-Meimon, G.; Gautron, J.; Martelly, I. Differential myogenicity of satellite cells isolated from extensor digitorum longus (EDL) and soleus rat muscles revealed in vitro. Cell Tissue Res. 1998, 291, 455–468. [Google Scholar] [CrossRef]
- Collins, C.A.; Olsen, I.; Zammit, P.S.; Heslop, L.; Petrie, A.; Partridge, T.A.; Morgan, J.E. Stem cell function, self-renewal, and behavioral heterogeneity of cells from the adult muscle satellite cell niche. Cell 2005, 122, 289–301. [Google Scholar] [CrossRef]
- Ono, Y.; Boldrin, L.; Knopp, P.; Morgan, J.E.; Zammit, P.S. Muscle satellite cells are a functionally heterogeneous population in both somite-derived and branchiomeric muscles. Dev. Biol. 2010, 337, 29–41. [Google Scholar] [CrossRef]
- Motohashi, N.; Uezumi, A.; Asakura, A.; Ikemoto-Uezumi, M.; Mori, S.; Mizunoe, Y.; Takashima, R.; Miyagoe-Suzuki, Y.; Takeda, S.; Shigemoto, K. Tbx1 regulates inherited metabolic and myogenic abilities of progenitor cells derived from slow- and fast-type muscle. Cell Death Differ. 2019, 26, 1024–1036. [Google Scholar] [CrossRef]
- Khodabukus, A.; Baar, K. Contractile and metabolic properties of engineered skeletal muscle derived from slow and fast phenotype mouse muscle. J. Cell. Physiol. 2015, 230, 1750–1757. [Google Scholar] [CrossRef]
- Tatsumi, R.; Sankoda, Y.; Anderson, J.E.; Sato, Y.; Mizunoya, W.; Shimizu, N.; Suzuki, T.; Yamada, M.; Rhoads, R.P.; Ikeuchi, Y.; et al. Possible implication of satellite cells in regenerative motoneuritogenesis: HGF upregulates neural chemorepellent Sema3A during myogenic differentiation. Am. J. Physiol. Cell Physiol. 2009, 297, 238–252. [Google Scholar] [CrossRef]
- Do Mai-Khoi, Q.; Sato, Y.; Shimizu, N.; Suzuki, T.; Shono, J.I.; Mizunoya, W.; Nakamura, M.; Ikeuchi, Y.; Anderson, J.E.; Tatsumi, R. Growth factor regulation of neural chemorepellent sema3A expression in satellite cell cultures. Am. J. Physiol. Cell Physiol. 2011, 301, 1270–1279. [Google Scholar]
- Do, M.K.Q.; Suzuki, T.; Gerelt, B.; Sato, Y.; Mizunoya, W.; Nakamura, M.; Ikeuchi, Y.; Anderson, J.E.; Tatsumi, R. Time-coordinated prevalence of extracellular HGF, FGF2 and TGF-β3 in crush-injured skeletal muscle. Anim. Sci. J. 2012, 83, 712–717. [Google Scholar] [CrossRef]
- Sato, Y.; Do, M.K.Q.; Suzuki, T.; Ohtsubo, H.; Mizunoya, W.; Nakamura, M.; Furuse, M.; Ikeuchi, Y.; Tatsumi, R. Satellite cells produce neural chemorepellent semaphorin 3A upon muscle injury. Anim. Sci. J. 2013, 84, 185–189. [Google Scholar] [CrossRef]
- Suzuki, T.; Do, M.K.Q.; Sato, Y.; Ojima, K.; Hara, M.; Mizunoya, W.; Nakamura, M.; Furuse, M.; Ikeuchi, Y.; Anderson, J.E.; et al. Comparative analysis of semaphorin 3A in soleus and EDL muscle satellite cells in vitro toward understanding its role in modulating myogenin expression. Int. J. Biochem. Cell Biol. 2013, 45, 476–482. [Google Scholar] [CrossRef]
- Tatsumi, R.; Suzuki, T.; Do, M.K.Q.; Ohya, Y.; Anderson, J.E.; Shibata, A.; Kawaguchi, M.; Ohya, S.; Ohtsubo, H.; Mizunoya, W.; et al. Slow-Myofiber Commitment by Semaphorin 3A Secreted from Myogenic Stem Cells. Stem Cells 2017, 35, 1815–1834. [Google Scholar] [CrossRef]
- Kang, J.S.; Yi, M.J.; Zhang, W.; Feinleib, J.L.; Cole, F.; Krauss, R.S. Netrins and neogenin promote myotube formation. J. Cell Biol. 2004, 167, 493–504. [Google Scholar] [CrossRef]
- Siegel, A.L.; Atchison, K.; Fisher, K.E.; Davis, G.E.; Cornelison, D.D.W. 3D Timelapse Analysis of Muscle Satellite Cell Motility. Stem Cells 2009, 27, 2527–2538. [Google Scholar] [CrossRef]
- Sun, K.L.W.; Correia, J.P.; Kennedy, T.E. Netrins: Versatile extracellular cues with diverse functions. Development 2011, 138, 2153–2169. [Google Scholar] [CrossRef]
- Huber, A.B.; Kolodkin, A.L.; Ginty, D.D.; Cloutier, J.-F. Signaling at the g rowth cone: Ligand-Receptor Complexes and the Control of Axon Growth and Guidance. Annu. Rev. Neurosci. 2003, 26, 509–563. [Google Scholar] [CrossRef]
- Mediero, A.; Ramkhelawon, B.; Perez-Aso, M.; Moore, K.J.; Cronstein, B.N. Netrin-1 is a critical autocrine/paracrine factor for osteoclast differentiation. J. Bone Miner. Res. 2015, 30, 837–854. [Google Scholar] [CrossRef]
- Hayashi, M.; Nakashima, T.; Taniguchi, M.; Kodama, T.; Kumanogoh, A.; Takayanagi, H. Osteoprotection by semaphorin 3A. Nature 2012, 485, 69–74. [Google Scholar] [CrossRef]
- Kang, J.S.; Mulieri, P.J.; Miller, C.; Sassoon, D.A.; Krauss, R.S. CDO, a Robo-related cell surface protein that mediates myogenic differentiation. J. Cell Biol. 1998, 143, 403–413. [Google Scholar] [CrossRef]
- Kang, J.S.; Mulieri, P.J.; Hu, Y.; Taliana, L.; Krauss, R.S. BOC, an Ig superfamily member, associates with CDO to positively regulate myogenic differentiation. EMBO J. 2002, 21, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Mizunoya, W.; Wakamatsu, J.-i.; Tatsumi, R.; Ikeuchi, Y. Protocol for high-resolution separation of rodent myosin heavy chain isoforms in a mini-gel electrophoresis system. Anal. Biochem. 2008, 377, 111–113. [Google Scholar] [CrossRef] [PubMed]
- Agbulut, O.; Noirez, P.; Beaumont, F.; Butler-Browne, G. Myosin heavy chain isoforms in postnatal muscle development of mice. Biol. Cell 2003, 95, 399–406. [Google Scholar] [CrossRef]
- Cole, F.; Zhang, W.; Geyra, A.; Kang, J.S.; Krauss, R.S. Positive regulation of myogenic bHLH factors and skeletal muscle development by the cell surface receptor CDO. Dev. Cell 2004, 7, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Bae, G.-U.; Yang, Y.-J.; Jiang, G.; Hong, M.; Lee, H.-J.; Tessier-Lavigne, M.; Kang, J.-S.; Krauss, R.S. Neogenin Regulates Skeletal Myofiber Size and Focal Adhesion Kinase and Extracellular Signal-regulated Kinase Activities In Vivo and In Vitro. Mol. Biol. Cell 2009, 20, 4920–4931. [Google Scholar] [CrossRef]
- Lee, H.J.; Bae, G.U.; Leem, Y.E.; Choi, H.K.; Kang, T.M.; Cho, H.; Kim, S.T.; Kang, J.S. Phosphorylation of Stim1 at serine 575 via netrin-2/Cdo-activated ERK1/2 is critical for the promyogenic function of Stim1. Mol. Biol. Cell 2012, 23, 1376–1387. [Google Scholar] [CrossRef]
- Ojima, K.; Uezumi, A.; Miyoshi, H.; Masuda, S.; Morita, Y.; Fukase, A.; Hattori, A.; Nakauchi, H.; Miyagoe-Suzuki, Y.; Takeda, S. Mac-1 low early myeloid cells in the bone marrow-derived SP fraction migrate into injured skeletal muscle and participate in muscle regeneration. Biochem. Biophys. Res. Commun. 2004, 321, 1050–1061. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, L.; Zhang, L.; Liu, X.; Hou, X.; Gao, H.; Yan, H.; Zhao, F.; Wang, L. NTN1 Affects Porcine Intramuscular Fat Content by Affecting the Expression of Myogenic Regulatory Factors. Animals 2019, 9, 609. [Google Scholar] [CrossRef]
- Graef, I.A.; Wang, F.; Charron, F.; Chen, L.; Neilson, J.; Tessier-Lavigne, M.; Crabtree, G.R. Neurotrophins and netrins require calcineurin/NFAT signaling to stimulate outgrowth of embryonic axons. Cell 2003, 113, 657–670. [Google Scholar] [CrossRef]
- Tierney, M.T.; Sacco, A. Satellite Cell Heterogeneity in Skeletal Muscle Homeostasis. Trends Cell Biol. 2016, 26, 434–444. [Google Scholar] [CrossRef]
- Feige, P.; Brun, C.E.; Ritso, M.; Rudnicki, M.A. Orienting Muscle Stem Cells for Regeneration in Homeostasis, Aging, and Disease. Cell Stem Cell 2018, 23, 653–664. [Google Scholar] [CrossRef]
- Rosenblatt, J.D.; Lunt, A.I.; Parry, D.J.; Partridge, T.A. Culturing satellite cells from living single muscle fiber explants. Vitr. Cell. Dev. Biol. Anim. 1995, 31, 773–779. [Google Scholar] [CrossRef]
- Hughes, S.M.; Taylor, J.M.; Tapscott, S.J.; Gurley, C.M.; Carter, W.J.; Peterson, C.A. Selective accumulation of MyoD and myogenin mRNAs in fast and slow adult skeletal muscle is controlled by innervation and hormones. Development 1993, 118, 1137–1147. [Google Scholar] [CrossRef]
- Hughes, S.M.; Koishi, K.; Rudnicki, M.; Maggs, A.M. MyoD protein is differentially accumulated in fast and slow skeletal muscle fibres and required for normal fibre type balance in rodents. Mech. Dev. 1997, 61, 151–163. [Google Scholar] [CrossRef]
- Maves, L.; Waskiewicz, A.J.; Paul, B.; Cao, Y.; Tyler, A.; Moens, C.B.; Tapscott, S.J. Pbx homeodomain proteins direct Myod activity to promote fast-muscle differentiation. Development 2007, 134, 3371–3382. [Google Scholar] [CrossRef]
- Stark, D.A.; Karvas, R.M.; Siege, A.L.; Cornelison, D.D.W. Eph/ephrin interactions modulate muscle satellite cell motility and patterning. Development 2011, 138, 5279–5289. [Google Scholar] [CrossRef]
- Stark, D.A.; Coffey, N.J.; Pancoast, H.R.; Arnold, L.L.; Walker, J.P.D.; Vallée, J.; Robitaille, R.; Garcia, M.L.; Cornelison, D.D.W. Ephrin-A3 promotes and maintains slow muscle fiber identity during postnatal development and reinnervation. 2015, 211, 1077–1091. J. Cell Biol. 2015, 211, 1077–1091. [Google Scholar] [CrossRef]
- Allen, R.E.; Temm-Grove, C.J.; Sheehan, S.M.; Rice, G. Skeletal muscle satellite cell cultures. Methods Cell Biol. 1997, 155–176. [Google Scholar]
- Tatsumi, R.; Yamada, M.; Katsuki, Y.; Okamoto, S.; Ishizaki, J.; Mizunoya, W.; Ikeuchi, Y.; Hattori, A.; Shimokawa, H.; Allen, R.E. Low-pH preparation of skeletal muscle satellite cells can be used to study activation in vitro. Int. J. Biochem. Cell Biol. 2006, 38, 1678–1685. [Google Scholar] [CrossRef]
- Seo, K.; Suzuki, T.; Kobayashi, K.; Nishimura, T. Adipocytes suppress differentiation of muscle cells in a co-culture system. Anim. Sci. J. 2019, 90, 423–434. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software “EZR” for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| siRNA | Sense (5′ to 3′) | Antisense (5′ to 3′) |
|---|---|---|
| Stealth_708 (netrin siRNA 1) | GAGGUGACCUAUGUGAGCCUGCAAU | AUUGCAGGCUCACAUAGGUCACCUC |
| Stealth_1463 (netrin siRNA 2) | UAUCACCCACCGGAAGGCUUGCAAA | UUUGCAAGCCUUCCGGUGGGUGAUA |
| Primer Name | Primer Sequence (5′ To 3′) Forward | Primer Sequence (5′ to 3′) Reverse | Probe No. |
|---|---|---|---|
| netrin-1 | CCCCTTGCATCAAGATTCC | GGCCTTGCAATAGGAGTCAC | 71 |
| netrin-3 | CCCAGGTTTCCAGCAGAG | GGTCCAGGGACAGGAGTCTTA | 2 |
| netrin-4 | CAGCACTGTGCACCATTGTA | TGACGTCGAAGTGACAGGTG | 81 |
| Sema3A | ATCAGTGGGTGCCTTACCAA | TCCGCCAAATGTTTTACTGG | 72 |
| neogenin | GGGGAATGAGACCAAAAATG | GGATGGGCACTAATCACAGG | 83 |
| BOC | GGACACCCATCCTGTACTACG | GAATGCCAGAAATGGTCCAG | 82 |
| CDO | GATTCCACAGAAGACACAGCAG | CCTCTGCTTCAGAATGACCAC | 94 |
| Unc5B | TTCCAGCTGCACACAACG | GCAGAGCAGAGAGCATCCA | 40 |
| Unc5C | GCACAGACCCCAGAATGAAT | CCCACGTAGAGAGCCACATC | 68 |
| embryonic MyHC | AGCCTCCAGCTTCAGAAGAA | GGCCCTCTCTGCCTCTATCT | 17 |
| perinatal MyHC | CCTGCAACTCCAGGTTCAAT | CTCTTTGATTTTGGCCTCCA | 73 |
| slow MyHC (I) | CAAGGTCAATACTCTGACCAAGG | CCATGCGCACCTTCTTCT | 78 |
| fast MyHC (IIa) | AGAGGGTTCTTGGCGAGAGT | AAGGCGCGGATGTTGTACT | 22 |
| fast MyHC (IIx) | TCTGCAGACGGAGTCAGGT | TTGAGTGAATGCCTGTTTGC | 33 |
| fast MyHC (IIb) | TGATGCAGGAGAAAAATGACC | ATCAGCTGGTCGCACCTTT | 1 |
| HPRT | CCTCCTCAGACCGCTTTTT | AACCTGGTTCATCATCGCTAA | 95 |
| Primer Name | Primer Sequence (5′ to 3′) Forward | Primer Sequence (5′ to 3′) Reverse | Amplicon (bp) |
|---|---|---|---|
| neogenin | TGAATTGTGAAGTTAATGCAGATTTGGTCC | TGTCCATGGATTCGTGAGCATATATGTTAG | 561 |
| BOC | GCCGAAGCTAATATCCCAGAAAGACTATAT | ACAGAGATATGTTGCCACCGATTATTGATT | 509 |
| CDO | GGAAGGCTTTTCATACAAACGTGTGAAATA | CAAGTCAAGATGTAAGGCTACAAACTACCT | 501 |
| DCC | GTATCTACGGCTTGAAACCAGCTGAATATA | CACATAGTTCCTCGAAAGTACTTCCAGAAA | 533 |
| Unc5A | GAGTCGCCCTCTCATCTCTACTACTG | GTTAAAGTTGATGTTGAAGCTCTGTCCATC | 519 |
| Unc5B | AACTAAGTAGACAGCTGTATACCCACTCTT | CACAATCAACTTGTCTCTACTCGATTCTCA | 552 |
| Unc5C | CTATTCAGAGATATGTGCTGGTGTAAGTCC | GAGCTTAAGTGTCAGCTTTTTAGAAACAGG | 563 |
| Unc5D | TCAAGAAGGGAGATAGACCTCAAACTAGTT | CTTCCTTGGAGACATTTATTCACTCACTCA | 592 |
| HPRT | CCGAGGATTTGGAAAAAGTGTTTATTCCTC | CTTTTCCAGTTTCACTAATGACACAAACGT | 555 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, T.; Mori, A.; Maeno, T.; Arimatsu, R.; Ichimura, E.; Nishi, Y.; Hisaeda, K.; Yamaya, Y.; Kobayashi, K.; Nakamura, M.; et al. Abundant Synthesis of Netrin-1 in Satellite Cell-Derived Myoblasts Isolated from EDL Rather Than Soleus Muscle Regulates Fast-Type Myotube Formation. Int. J. Mol. Sci. 2021, 22, 4499. https://doi.org/10.3390/ijms22094499
Suzuki T, Mori A, Maeno T, Arimatsu R, Ichimura E, Nishi Y, Hisaeda K, Yamaya Y, Kobayashi K, Nakamura M, et al. Abundant Synthesis of Netrin-1 in Satellite Cell-Derived Myoblasts Isolated from EDL Rather Than Soleus Muscle Regulates Fast-Type Myotube Formation. International Journal of Molecular Sciences. 2021; 22(9):4499. https://doi.org/10.3390/ijms22094499
Chicago/Turabian StyleSuzuki, Takahiro, Aika Mori, Takahiro Maeno, Rio Arimatsu, Emi Ichimura, Yuriko Nishi, Kouga Hisaeda, Yuki Yamaya, Ken Kobayashi, Mako Nakamura, and et al. 2021. "Abundant Synthesis of Netrin-1 in Satellite Cell-Derived Myoblasts Isolated from EDL Rather Than Soleus Muscle Regulates Fast-Type Myotube Formation" International Journal of Molecular Sciences 22, no. 9: 4499. https://doi.org/10.3390/ijms22094499
APA StyleSuzuki, T., Mori, A., Maeno, T., Arimatsu, R., Ichimura, E., Nishi, Y., Hisaeda, K., Yamaya, Y., Kobayashi, K., Nakamura, M., Tatsumi, R., Ojima, K., & Nishimura, T. (2021). Abundant Synthesis of Netrin-1 in Satellite Cell-Derived Myoblasts Isolated from EDL Rather Than Soleus Muscle Regulates Fast-Type Myotube Formation. International Journal of Molecular Sciences, 22(9), 4499. https://doi.org/10.3390/ijms22094499