
Salivary Proteome Changes in Response to Acute Psychological Stress Due to an Oral Exam Simulation in University Students: Effect of an Olfactory Stimulus


 , ,
, ,  ,
,
Abstract
1. Introduction
2. Results
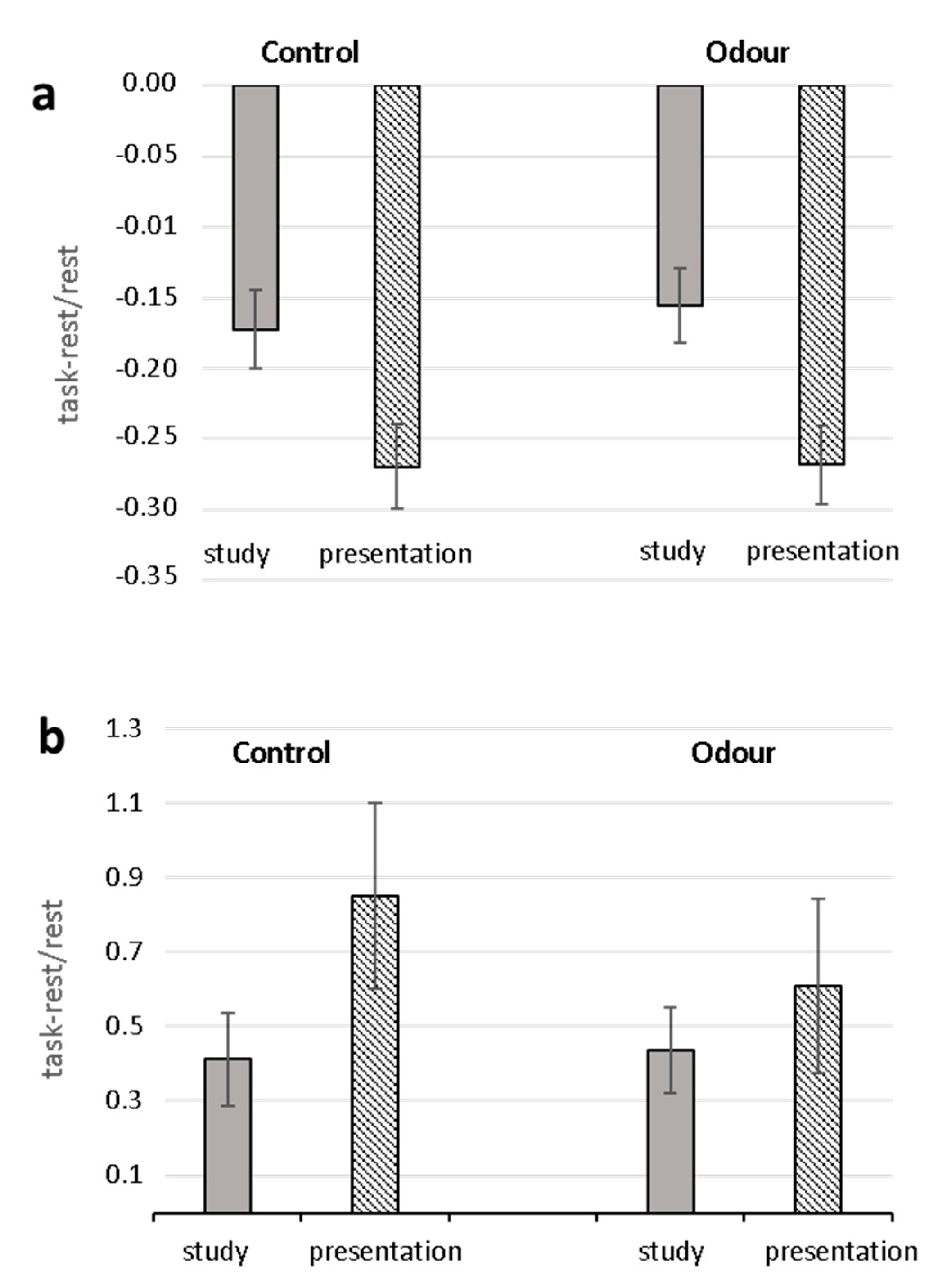
2.1. Questionnaires and Heart Rate Results
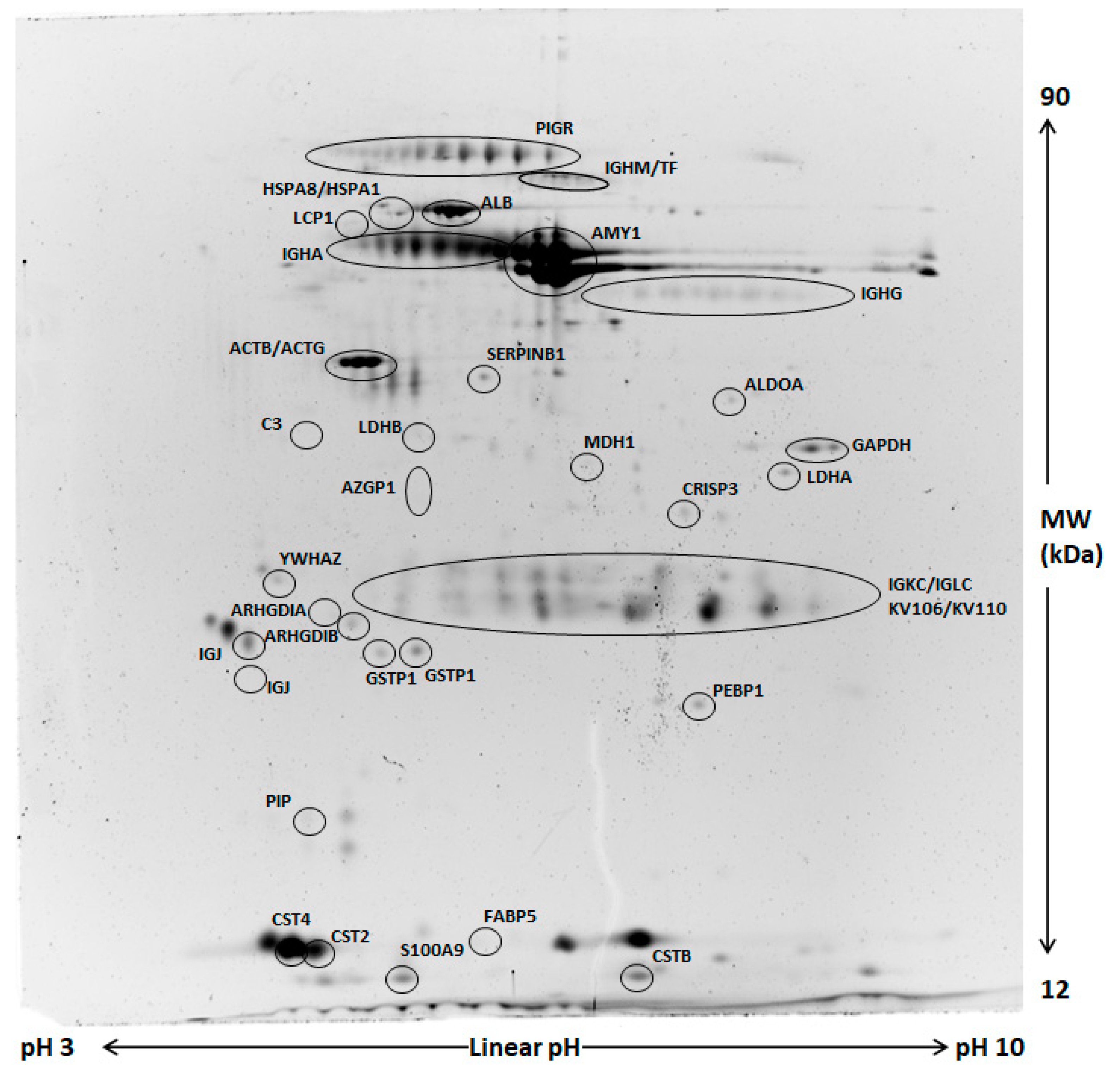
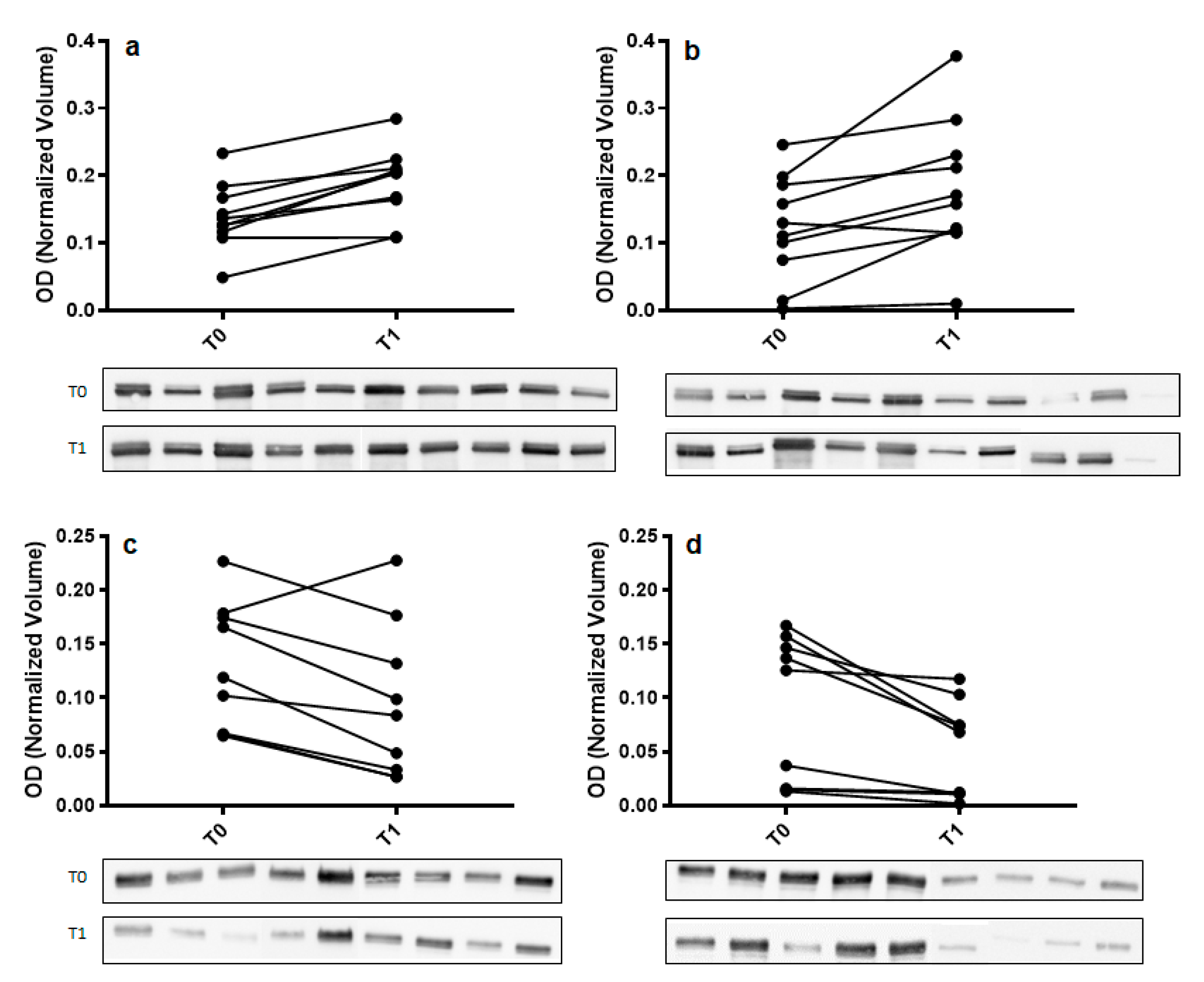
2.2. Comparative Proteomic Analysis and Validation
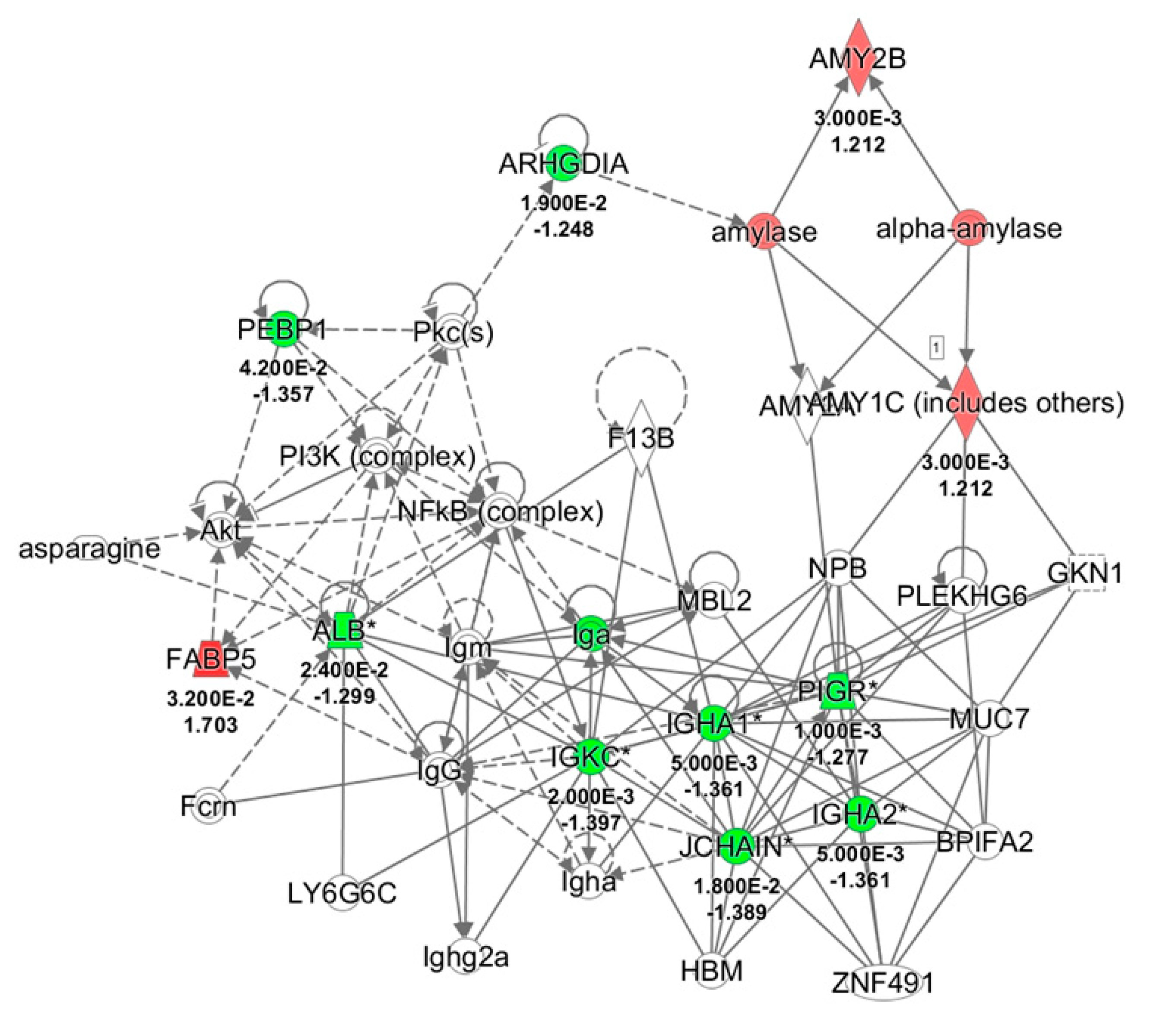
2.3. Ingenuity Pathways Analysis (IPA)
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Physiological Parameters
4.3. Experimental Protocol
4.4. Questionnaires and Heart Analysis
4.5. Salivary Samples
4.6. Proteomic Analysis
4.7. In-Gel Digestion and Mass Spectrometry
4.8. WB Analysis
4.9. IPA
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zeidner, M. Test Anxiety. In The Wiley Handbook of Anxiety Disorders; Emmelkamp, P., Ehring, T., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2014; pp. 581–595. [Google Scholar] [CrossRef]
- Cassady, J.C.; Johnson, R.E. Cognitive test anxiety and academic performance. Contemp. Educ. Psychol. 2002, 27, 270–295. [Google Scholar] [CrossRef]
- Holm-Hadulla, R.M.; Hofmann, F.H.; Sperth, M.; Funke, J. Psychological complaints and mental disorders of students. Psychotherapeut 2009, 54, 346–356. [Google Scholar] [CrossRef]
- Khosravi, M.; Bigdeli, I. Relationships between Characteristics of Personality and Exam Anxiety among Students. J. Behav. Sci. 2008, 2, 13–24. [Google Scholar]
- Eum, K.; Rice, K.G. Test anxiety, perfectionism, goal orientation, and academic performance. Anxiety Stress Coping 2011, 24, 167–178. [Google Scholar] [CrossRef]
- Pekrun, R.; Stephens, E.J. Test Anxiety and Academic Achievement. In International Encyclopedia of the Social & Behavioral Sciences, 2nd ed.; Wright, J.D., Ed.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 244–249. [Google Scholar]
- Fink, G. Stress: Concepts, definition and history. In Reference Module Neuroscience and Biobehavioral Psychology; Stein, J.P., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–9. [Google Scholar]
- Rohleder, N.; Wolf, J.M.; Maldonado, E.F.; Kirschbaum, C. The psychosocial stress-induced increase in salivary alpha-amylase is independent of saliva flow rate. Psychophysiology 2006, 43, 645–652. [Google Scholar] [CrossRef]
- van Stegeren, A.H.; Wolf, O.T.; Kindt, M. Salivary alpha amylase and cortisol responses to different stress tasks: Impact of sex. Int. J. Psychophysiol. 2008, 69, 33–40. [Google Scholar] [CrossRef]
- Nater, U.M.; Rohleder, N. Salivary alpha-amylase as a non-invasive biomarker for the sympathetic nervous system: Current state of research. Psychoneuroendocrinology 2009, 34, 486–496. [Google Scholar] [CrossRef]
- Obayashi, K. Salivary mental stress proteins. Clin. Chim. Acta 2013, 425, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Ballanger, B.; Bath, K.G.; Mandairon, N. Odorants: A tool to provide nonpharmacological intervention to reduce anxiety during normal and pathological aging. Neurobiol. Aging 2019, 82, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Sayed, A.M.; Morsy, S.; Tawfik, G.M.; Naveed, S.; Minh-Duc, N.T.; Hieu, T.H.; Ali, Z.A.; Shinkar, A.; Doheim, M.F.; Hashan, M.R.; et al. The best route of administration of lavender for anxiety: A systematic review and network meta-analysis. Gen. Hosp. Psychiatry 2020, 64, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Aguilar, M.D.; Vialaret, J.; de Périère, D.D.; Escribano, D.; Lehmann, S.; Tecles, F.; Cerón, J.J.; Hirtz, C. Variation of human salivary alpha-amylase proteoforms in three stimulation models. Clin. Oral Investig. 2020, 24, 475–486. [Google Scholar] [CrossRef]
- Carreira, L.; Castelo, P.M.; Simões, C.; Silva, F.C.E.; Viegas, C.; Lamy, E. Changes in Salivary Proteome in Response to Bread Odour. Nutrients 2020, 12, 1002. [Google Scholar] [CrossRef]
- Baevsky, R.M.; Chernikova, A.G. Heart rate variability analysis: Physiological foundations and main methods. Cardiometry 2017, X, 66–76. [Google Scholar] [CrossRef]
- Bosch, J.A.; Ring, C.; de Geus, E.J. Stress and secretory immunity. Int. Rev. Neurobiol. 2002, 52, 213–253. [Google Scholar]
- Gallacher, D.V.; Petersen, O.H. Stimulus-secretion coupling in mammalian salivary glands. Int. Rev. Physiol. 1983, 28, 1–52. [Google Scholar]
- Speirs, R.L.; Herring, J.; Cooper, W.D.; Hardy, C.C.; Hind, C.R. The influence of sympathetic activity and isoprenaline on the secretion of amylase from the human parotid gland Arch. Oral Biol. 1974, 19, 747–752. [Google Scholar] [CrossRef]
- Nederfors, T.; Dahlof, C. Effects of the beta-adrenoceptor antagonists atenolol and propranolol on human whole saliva flow rate and composition. Arch. Oral Biol. 1992, 37, 579–584. [Google Scholar] [CrossRef]
- Nederfors, T.; Ericsson, T.; Twetman, S.; Dahlof, C. Effects of the beta-adrenoceptor antagonists atenolol and propranolol on human parotid and submandibular-sublingual salivary secretion. J. Dent. Res. 1994, 73, 5–10. [Google Scholar] [CrossRef] [PubMed]
- van Stegeren, A.; Rohleder, N.; Everaerd, W.; Wolf, O.T. Salivary alpha amylase as marker for adrenergic activity during stress: Effect of betablockade. Psychoneuroendocrinology 2006, 31, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Groer, M.; Murphy, R.; Bunnell, W.; Salomon, K.; Van Eepoel, J.; Rankin, B.; White, K.; Bykowski, C. Salivary measures of stress and immunity in police officers engaged in simulated critical incident scenarios. J. Occup. Environ. Med. 2010, 52, 595–602. [Google Scholar] [CrossRef]
- McGraw, L.K.; Out, D.; Hammermeister, J.J.; Ohlson, C.J.; Pickering, M.A.; Granger, D.A. Nature, correlates, and consequences of stress-related biological reactivity and regulation in Army nurses during combat casualty simulation. Psychoneuroendocrinology 2013, 38, 135–144. [Google Scholar] [CrossRef]
- Strahler, J.; Ziegert, T. Psychobiological stress response to a simulated school shooting in police officers. Psychoneuroendocrinology 2015, 51, 80–91. [Google Scholar] [CrossRef]
- Trueba, A.F.; Mizrachi, D.; Auchus, R.J.; Vogel, P.D.; Ritz, T. Effects of psychosocial stress on the pattern of salivary protein release. Physiol. Behav. 2012, 105, 841–849. [Google Scholar] [CrossRef]
- Valentin, B.; Grottke, O.; Skorning, M.; Bergrath, S.; Fischermann, H.; Rörtgen, D.; Mennig, M.T.; Fitzner, C.; Müller, M.P.; Kirschbaum, C.; et al. Cortisol and alpha-amylase as stress response indicators during pre-hospital emergency medicine training with repetitive high-fidelity simulation and scenarios with standardized patients. Scand. J. Trauma Resusc. Emerg. Med. 2015, 23, 31. [Google Scholar] [CrossRef] [PubMed]
- Campisi, J.; Bravo, Y.; Cole, J.; Gobeil, K. Acute psychosocial stress differentially influences salivary endocrine and immune measures in undergraduate students. Physiol. Behav. 2012, 107, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Bosch, J.A.; de Geus, E.J.; Veerman, E.C.; Hoogstraten, J.; Nieuw Amerongen, A.V. Innate secretory immunity in response to laboratory stressors that evoke distinct patterns of cardiac autonomic activity. Psychosom. Med. 2003, 65, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Ring, C.; Carroll, D.; Willemsen, G.; Cooke, J.; Ferraro, A.; Drayson, M. Secretory immunoglobulin A and cardiovascular activity dur-ing mental arithmetic and paced breathing. Psychophysiology 1999, 36, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Willemsen, G.; Ring, C.; Carroll, D.; Evans, P.; Clow, A.; Hucklebridge, F. Secretory immunoglobulin A and cardiovascular reactions tomental arithmetic and cold pressor. Psychophysiology 1998, 35, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Takatsuji, K.; Sugimoto, Y.; Ishizaki, S.; Ozaki, Y.; Matsuyama, E.; Yamaguchi, Y. The effects of examination stress on salivary cortisol, immunoglobulin A, and chromogranina in nursing students. Biomed. Res. 2008, 29, 221–224. [Google Scholar] [CrossRef]
- Bosch, J.A.; Brand, H.S.; Ligtenberg, T.J.; Bermond, B.; Hoogstraten, J.; Amerongen, A.V.N. Psychological stress as a determinant of protein levels and salivary-induced aggregation of Streptococcus gordonii in human whole saliva. Psychosom. Med. 1996, 58, 374–382. [Google Scholar] [CrossRef]
- Huwe, S.; Hennig, J.; Netter, P. Biological, emotional, behavioral, and coping reactions to examination stress in high and low state anxious subjects, Anxiety. Stress Coping 1998, 11, 47–65. [Google Scholar] [CrossRef]
- McClelland, D.C.; Ross, G.; Patel, V. The Effect of an Academic Examination on Salivary Norepinephrine and Immunoglobulin Levels. J. Hum. Stress 1985, 11, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Spangler, G. Psychological and physiological responses during an exam and their relation to personality characteristics. Psychoneuroendocrinology 1997, 22, 423–441. [Google Scholar] [CrossRef]
- Winzer, A.; Ring, C.; Carroll, D.; Willemsen, G.; Drayson, M.; Kendall, M. Secretory immunoglobulin A and cardiovascular reactionsto mental arithmetic, cold pressor, and exercise: Effects of beta-adrenergic blockade. Psychophysiology 1999, 36, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Marvin, R.K.; Saepoo, M.B.; Ye, S.; White, D.B.; Liu, R.; Hensley, K.; Rega, P.; Kazan, V.; Giovannucci, D.R.; Isailovic, D. Salivary protein changes in response to acute stress in medical residents performing advanced clinical simulations: A pilot proteomics study. Biomarkers 2017, 22, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Bosch, J.A.; de Geus, E.J.; Kelder, A.; Veerman, E.C.; Hoogstraten, J.; Amerongen, A.V. Differential effects of active versus passive coping on secretory immunity. Psychophysiology 2001, 38, 836–846. [Google Scholar] [CrossRef]
- Ring, C.; Harrison, L.K.; Winzer, A.; Carroll, D.; Drayson, M.; Kendall, M. Secretory immunoglobulin A and cardiovascular reactions to mental arithmetic, cold pressor, and exercise: Effects of alpha-adrenergic blockade. Psychophysiology 2000, 37, 634–643. [Google Scholar] [CrossRef]
- Kugler, J.; Reintjes, F.; Tewes, V.; Schedlowski, M. Competition Stress in soccer coaches increases salivary immunoglobulin A and salivary cortisol concentrations. J. Sports Med. Phys. Fit. 1996, 36, 117–120. [Google Scholar]
- Brandtzaeg, P. Synthesis and secretion of human salivary immunoglobulins. In Glandular Mechanisms of Salivary Secretion; Garrett, J.R., Ekström, J., Anderson, L.C., Eds.; Karger: London, UK, 1998; Volume 10, pp. 167–199. [Google Scholar]
- Proctor, G.B.; Carpenter, G.H. Regulation of salivary gland function by autonomic nerves. Auton. Neurosci. 2007, 133, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Rossoni, R.B.; Machado, A.B.; Machado, C.R. A histochemical study of catecholamines and cholinesterases in the autonomic nerves of the human minor salivary glands. Histochem. J. 1979, 11, 661–668. [Google Scholar] [CrossRef]
- Carpenter, G.H.; Garrett, J.R.; Hartley, R.H.; Proctor, G.B. The influence of nerves on the secretion of immunoglobulin A into sub-mandibular saliva in rats. J. Physiol. 1998, 512, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, G.H.; Proctor, G.B.; Anderson, L.C.; Zhang, X.S.; Garrett, J.R. Immunoglobulin A secretion into saliva during dual sympathetic and parasympathetic nerve stimulation of rat submandibular glands. Exp. Physiol. 2000, 85, 281–286. [Google Scholar] [CrossRef]
- Kelleher, R.S.; Hann, L.E.; Edwards, J.A.; Sullivan, D.A. Endocrine, neural, and immune control of secretory component output by lacrimal gland acinar cells. J. Immunol. 1991, 146, 3405–3412. [Google Scholar] [PubMed]
- McGee, D.; Eran, M.; McGhee, J.R.; Freier, S. Substance P Accelerates Secretory Component-Mediated Transcytosis of IgA in the Rat Intestine. In Advances in Mucosal Immunology. Advances in Experimental Medicine and Biology; Mestecky, J., Russell, M.W., Jackson, S., Michalek, S.M., Tlaskalová-Hogenová, H., Šterzl, J., Eds.; Springer: Boston, MA, USA, 1995; p. 371. [Google Scholar] [CrossRef]
- Schmidt, P.T.; Eriksen, L.; Loftager, M.; Rasmussen, T.N.; Holst, J.J. Fast acting nervous regulation of immunoglobulin A secretion from isolated perfused porcine ileum. Gut 1999, 45, 679–685. [Google Scholar] [CrossRef]
- Wilson, I.D.; Soltis, R.D.; Olson, R.E.; Erlandsen, S.L. Cholinergic stimulation of immunoglobulin a secretion in rat intestine. Gastroenterology 1982, 83, 881–888. [Google Scholar] [CrossRef]
- Castagnola, M.; Cabras, T.; Iavarone, F.; Vincenzoni, F.; Vitali, A.; Pisano, E.; Nemolato, S.; Scarano, E.; Fiorita, A.; Vento, G.; et al. Top-down platform for deciphering the human salivary proteome. J. Matern. Fetal Neonatal Med. 2012, 25, 27–43. [Google Scholar] [CrossRef]
- Hopsu-Havu, V.K.; Joronen, I.A.; Järvinen, M.; Rinne, A.; Aalto, M. Cysteine proteinase inhibitors produced by mononuclear phagocytes. Cell Tissue Res. 1984, 236, 161–164. [Google Scholar] [CrossRef]
- Suzuki, T.; Hashimoto, S.; Toyoda, N.; Nagai, S.; Yamazaki, N.; Dong, H.Y.; Sakai, J.; Yamashita, T.; Nukiwa, T.; Matsushima, K. Comprehensive gene expression profile of LPS stimulated human monocytes by SAGE. Blood 2000, 96, 2584–2591. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Tay, K.H.; Dong, L.; Thorne, R.F.; Jiang, C.C.; Yang, E.; Tseng, H.Y.; Liu, H.; Christopherson, R.; Hersey, P.; et al. Cystatin B inhibition of TRAIL-induced apoptosis is associated with the protection of FLIP(L) from degradation by the E3 ligase itch in human melanoma cells. Cell Death Differ. 2010, 17, 1354–1367. [Google Scholar] [CrossRef] [PubMed]
- Ceru, S.; Konjar, S.; Maher, K.; Repnik, U.; Krizaj, I.; Bencina, M.; Renko, M.; Nepveu, A.; Žerovnik, E.; Turk, B.; et al. Stefin B interacts with histones and cathepsin L in the nucleus. J. Biol. Chem. 2010, 285, 10078–10086. [Google Scholar] [CrossRef]
- Maher, K.; Kokelj, B.J.; Butinar, M.; Mikhaylov, G.; Manček-Keber, M.; Stoka, V.; Vasiljeva, O.; Turk, B.; Grigoryev, S.A.; Kopitar-Jerala, N. A role for stefin B (cystatin B) in inflammation and endotoxemia. J. Biol. Chem. 2014, 289, 31736–31750. [Google Scholar] [CrossRef] [PubMed]
- Lehtinen, M.K.; Tegelberg, S.; Schipper, H.; Su, H.; Zukor, H.; Manninen, O.; Kopra, O.; Joensuu, T.; Hakala, P.; Bonni, A.; et al. Cystatin B deficiency sensitizes neurons to oxidative stress in progressive myoclonus epilepsy, EPM1. J. Neurosci. 2009, 29, 5910–5915. [Google Scholar] [CrossRef] [PubMed]
- Contini, C.; Firinu, D.; Serrao, S.; Manconi, B.; Olianas, A.; Cinetto, F.; Cossu, F.; Castagnola, M.; Messana, I.; Del Giacco, S.; et al. RP-HPLC-ESI-IT Mass Spectrometry Reveals Significant Variations of the Human Salivary Protein Profile Associated with Predominantly Antibody Deficiencies. J. Clin. Immunol. 2020, 40, 329–339. [Google Scholar] [CrossRef]
- de Bruin, W.C.C.; Wagenmans, M.J.M.; Peters, W.H.M. Expression of glutathione s-transferase alpha, p1-1 and t1-1 in the human gastrointestinal tract. Cancer Sci. 2000, 91, 310–316. [Google Scholar]
- Kang, J.H.; Hwang, S.M.; Chung, I.Y. S100A8, S100A9 and S100A12 activate airway epithelial cells to produce MUC5AC via extracellular signal-regulated kinase and nuclear factor-κB pathways. Immunology 2015, 144, 79–90. [Google Scholar] [CrossRef]
- Cho, S.; Yong, H.; Park, H.; Ryoo, K.; Kang, K.; Park, J.; Eom, S.J.; Kim, M.J.; Chang, T.S.; Choi, S.Y.; et al. Glutathione S-transferase mu modulates the stress-activated signals by suppressing apoptosis signal-regulating kinase 1. J. Biol. Chem. 2001, 276, 12749–12755. [Google Scholar] [CrossRef]
- Hassan, M.I.; Waheed, A.; Yadav, S.; Singh, T.P.; Ahmad, F. Review: Prolactin inducible protein in cancer, fertility and immunoregulation: Structure, function and its clinical implications. Cell Mol. Life Sci. 2009, 66, 447–459. [Google Scholar] [CrossRef]
- Mounayar, R.; Morzel, M.; Brignot, H.; Tremblay-Franco, M.; Canlet, C.; Lucchi, G.; Ducoroy, P.; Feron, G.; Neyraud, E. Salivary markers of taste sensitivity to oleic acid: A combined proteomics and metabolomics approach. Metabolomics 2014, 10, 688–696. [Google Scholar] [CrossRef]
- Szabo, Y.Z.; Slavish, D.C.; Graham-Engeland, J.E. The effect of acute stress on salivary markers of inflammation: A systematic review and meta-analysis. Brain Behav. Immun. 2020, 88, 887–900. [Google Scholar] [CrossRef]
- Rusconi, F.; Grillo, B.; Ponzoni, L.; Bassani, S.; Toffolo, E.; Paganini, L.; Mallei, A.; Braida, D.; Passafaro, M.; Popoli, M.; et al. LSD1 modulates stress-evoked transcription of immediate early genes and emotional behavior. Proc. Natl. Acad. Sci. USA 2016, 113, 3651–3656. [Google Scholar] [CrossRef]
- Rusconi, F.; Grillo, B.; Toffolo, E.; Mattevi, A.; Battaglioli, E. NeuroLSD1: Splicing-Generated Epigenetic Enhancer of Neuroplasticity. Trends Neurosci. 2017, 40, 28–38. [Google Scholar] [CrossRef]
- Sela, L.; Sobel, N. Human olfaction: A constant state of change-blindness. Exp. Brain Res. 2010, 205, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Baroni, D.; Caccico, L.; Fiesoli, A.; Grassi, E.; Lauretta, F.; Marsigli, N.; Rosadoni, M.; Chiorri, C. Italian validation of the Self-Report Liebowitz Social Anxiety Scale. In Proceedings of the 9th World Congress of Behavioural and Cognitive Therapies, Berlin, Germany, 17–20 July 2019. Unpublished. [Google Scholar] [CrossRef]
- Smeets, M.A.; Schifferstein, H.N.; Boelema, S.R.; Lensvelt-Mulders, G. The Odor Awareness Scale: A new scale for measuring positive and negative odor awareness. Chem. Senses 2008, 33, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Sica, C.; Chiri, L.R.; Musoni, I.; Bisi, B. Social Phobia Scale (SPS) and Social Interaction Anxiety Scale (SIAS): Their psycometric properties on italian population. Boll. Psicol. Appl. 2007, 252, 59–71. [Google Scholar]
- Gori, A.; Giannini, M.; Socci, S.; Luca, M.; Dewey, D.E.; Schuldberg, D.; Craparo, G. Assessing social anxiety disorder: Psychometric properties of the Italian social phobia inventory (I-SPIN). Clin. Neuropsychiatry 2013, 10, 37. [Google Scholar]
- Davidson, J.R.T.; Potts, N.L.S.; Richichi, E.A.; Ford, S.M.; Krishnan, K.R.R.; Smith, R.D.; Wilson, W. Brief Social Phobia Scale (BSPS) [Database record]. APA PsycTests 1991. [Google Scholar] [CrossRef]
- Howard, S.; Hughes, B.M. Expectancies, not aroma, explain impact of lavender aromatherapy on psychophysiological indices of relaxation in young healthy women. Br. J. Health Psychol. 2008, 13, 603–617. [Google Scholar] [CrossRef]
- Ciregia, F.; Giacomelli, C.; Giusti, L.; Boldrini, C.; Piga, I.; Pepe, P.; Consensi, A.; Gori, S.; Lucacchini, A.; Mazzoni, M.R.; et al. Putative salivary biomarkers useful to differentiate patients with fibromyalgia. J. Proteomics. 2019, 190, 44–54. [Google Scholar] [CrossRef]
- Ciregia, F.; Giusti, L.; Da Valle, Y.; Donadio, E.; Consensi, A.; Giacomelli, C.; Sernissi, F.; Scarpellini, P.; Maggi, F.; Lucacchini, A.; et al. A multidisciplinary approach to study a couple of monozygotic twins discordant for the chronic fatigue syndrome: A focus on potential salivary biomarkers. J. Transl. Med. 2013, 11, 243. [Google Scholar] [CrossRef]
- Lacerenza, S.; Ciregia, F.; Giusti, L.; Bonotti, A.; Greco, V.; Giannaccini, G.; D’Antongiovanni, V.; Fallahi, P.; Pieroni, L.; Cristaudo, A.; et al. Putative Biomarkers for Malignant Pleural Mesothelioma Suggested by Proteomic Analysis of Cell Secretome. Cancer Genom. Proteom. 2020, 17, 225–236. [Google Scholar] [CrossRef]
- Giusti, L.; Angeloni, C.; Barbalace, M.C.; Lacerenza, S.; Ciregia, F.; Ronci, M.; Urbani, A.; Manera, C.; Digiacomo, M.; Macchia, M.; et al. A Proteomic Approach to Uncover Neuroprotective Mechanisms of Oleocanthal against Oxidative Stress. Int. J. Mol. Sci. 2018, 19, 2329. [Google Scholar] [CrossRef] [PubMed]
- Ciregia, F.; Giusti, L.; Ronci, M.; Bugliani, M.; Piga, I.; Pieroni, L.; Rossi, C.; Marchetti, P.; Urbani, A.; Lucacchini, A. Glucagon-like peptide 1 protects INS-1E mitochondria against palmitate-mediated beta-cell dysfunction: A protomi study. Mol. Biosyst. 2015, 11, 1696–1707. [Google Scholar] [CrossRef]
- Ciregia, F.; Bugliani, M.; Ronci, M.; Giusti, L.; Boldrini, C.; Mazzoni, M.R.; Mossuto, S.; Grano, F.; Cnop, M.; Marselli, L.; et al. Palmitate-induced lipotoxicity alters acetylation of multiple proteins in clonal β cells and human pancreatic islets. Sci. Rep. 2017, 7, 13445. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scales | Control | Odour |
|---|---|---|
| mean ± SD | mean ± SD | |
| Liebowitz | 41.89 ± 16.76 | 38.89 ± 16.66 |
| Westside | 2.81 ± 0.76 | 2.76 ± 0.59 |
| I-SPIN | 21.11 ± 12.12 | 20.83 ± 8.26 |
| BSPS-Fear | 6.21 ± 3.94 | 7.06 ± 2.76 |
| BSPS-Avoidance | 6.21 ± 3.58 | 6.28 ± 3.16 |
| BSPS-Physiology | 4.53 ± 2.61 | 4.89 ± 3.28 |
| STAI-Y2 | 50.32 ± 11.94 | 48.50 ± 9.71 |
| SPS | 19.21 ± 13.16 | 20.50 ± 8.57 |
| SIAS | 25.84 ± 13.89 | 23.94 ± 10.09 |
| Parameters | Phases | Control | Odour |
|---|---|---|---|
| mean ± SD | mean ± SD | ||
| relaxation | 866.22 ± 88.12 | 910.07 ± 98.38 | |
| RR (msec) | study | 718.11 ± 119.49 | 767.92 ± 129.23 |
| presentation | 631.52 ± 96.10 | 671.09 ± 123.21 | |
| relaxation | 51.86 ± 23.69 | 51.65 ± 27.83 | |
| RMSSD (msec) | study | 41.95 ± 18.85 | 38.24 ± 23.67 |
| presentation | 37.32 ± 22.31 | 36.38 ± 26.31 | |
| relaxation | 8.15 ± 2.34 | 8.88 ± 3.08 | |
| Stress Index | study | 11.23 ± 3.43 | 12.13 ± 4.12 |
| presentation | 14.38 ± 7.27 | 13.09 ± 5.58 |
| PREP | Control | Odour |
|---|---|---|
| mean ± SD | mean ± SD | |
| confidence | 2.43 ± 1.02 | 2.35 ± 1.05 |
| nervousness | 2.86 ± 0.77 | 2.88 ± 1.05 |
| calmness | 2.42 ± 0.94 | 2.82 ± 1.07 |
| preparedness | 2.43 ± 0.75 | 2.23 ± 1.03 |
| how good | 2.14 ± 0.53 | 1.94 ± 0.82 |
| # | ID | Protein | Gene | Cov | Pep | Unic | MW (th) | pI (th) | p-Value (min–max) | Ratio (T1/T0) (min–max) |
|---|---|---|---|---|---|---|---|---|---|---|
| 67, 69 | P01833 | Polymeric immunoglobulin receptor | PIGR | 24 | 18 | 18 | 83,284 | 5.59 | 0.030–0.044 | 0.80–0.86 |
| 98, 100, 102, 105 | P01871 | Ig µ chain C region | IGHM | 33 | 15 | 15 | 49,307 | 6.35 | 0.008–0.046 | 0.75–0.80 |
| 98, 100, 102, 105 | P02787 | Serotransferrin | TF | 16 | 12 | 12 | 77,064 | 6.7 | 0.008–0.046 | 0.75–0.80 |
| 126, 128 | P11142 | Heat shock cognate 71 kDa protein | HSPA8 | 45 | 30 | 28 | 70,898 | 5.37 | 0.003–0.006 | 0.69-0.77 |
| 170 187 | P02768 | Serum albumin | ALB | 74 | 61 | 61 | 69,367 | 5.67 | 0.008 | 0.66–0.71 |
| 186 | P0DMV8 P0DMV9 | Heat shock 70 kDa protein 1A, 1B | HSPA1A HSPA1B | 37 | 25 | 20 | 69,921 | 5.48 | 0.0176 | 0.67 |
| 211 | P13796 | Plastin-2 | LCP1 | 29 | 16 | 16 | 70,289 | 5.29 | 0.0185 | 0.566 |
| 271, 339, 348 | P0DUB6 | α-amylase,1A | AMY1A | 76 | 99 | 99 | 57,768 | 6.34 | 0.0001–0.004 | 1.30–1.39 |
| 364, 368, 369, 370, 373, 374, 375, 376, 1148 | P01857 P01859 | Ig γ-1, 2 chain C region | IGHG1, IGHG2 | 21 | 7 | 5 | 36,106 | 8.46 | 0.0001–0.012 | 0.52–0.80 |
| 457, 461, 464 | P60709 P63261 | Actin, cytoplasmic Iso 1, 2 | ACTB, ACTG | 29 | 10 | 9 | 41,737 | 5.29 | 0.007–0.021 | 0.75–0.79 |
| 487 | P30740 | Leukocyte elastase inhibitor | SERPINB1 | 36 | 17 | 17 | 42,742 | 5.90 | 0.024 | 0.69 |
| 539 | P01024 | Complement C3c a-chain fragment | C3 | 4 | 6 | 6 | 39,488 | 4.79 | 0.019 | 0.62 |
| 1155 | P04406 | Glyceraldehyde-3-phosphate dehydrogenase | GAPDH | 31 | 16 | 16 | 35,922 | 8.58 | 0.011 | 0.51 |
| 582 | P40925 | Malate dehydrogenase | MDH1 | 15 | 4 | 4 | 36,426 | 6.89 | 0.049 | 0.75 |
| 585, 612 | P25311 | Zinc-α-2-glycoprotein | AZGP1 | 38 | 11 | 11 | 34,259 | 5.58 | 0.019–0.024 | 0.57–0.63 |
| 1158 | P00338 | L-lactate dehydrogenase A chain | LDHA | 30 | 13 | 11 | 36,558 | 8.46 | 0.0498 | 0.56 |
| 636 | P54108 | Cysteine-rich secretory protein 3 | CRISP3 | 4 | 1 | 1 | 27,630 | 8,11 | 0.003 | 0.62 |
| 692 | P63104 | 14-3-3 protein ζ/δ | YWHAZ | 28 | 7 | 5 | 27,745 | 4.73 | 0.013 | 0.61 |
| 733 | P52565 | Rho GDP-dissociation inhibitor 1 | ARHGDIA | 29 | 6 | 6 | 23,207 | 5.01 | 0.0007 | 0.65 |
| 745 | P52566 | Rho GDP-dissociation inhibitor 2 | ARHGDIB | 44 | 11 | 11 | 22,988 | 5.08 | 0.007 | 0.53 |
| 756 | P01591 | Immunoglobulin J chain | IGJ | 45 | 10 | 10 | 18,099 | 5.09 | 0.025 | 0.78 |
| 770, 771 | P09211 | Glutathione S-transferase P | GSTP1 | 39 | 7 | 6 | 23,356 | 5.44 | 0.013–0.026 | 0.65–0.67 |
| 1159 | P12273 | Prolactin inducible protein | PIP | 69 | 10 | 10 | 13,523 | 5.40 | 0.032 | 0.80 |
| 1160 | P06702 | Calgranulin B | S100A9 | 32 | 3 | 3 | 13,241 | 5.71 | 0.027 | 0.70 |
| 1018 | P09228 | Cystatin SA | CST2 | 70 | 27 | 17 | 14,350 | 4.85 | 0.047 | 1.43 |
| 1020 | P01036 | Cystatin S | CST4 | 59 | 13 | 2 | 14,189 | 4.83 | 0.044 | 1.30 |
| 1162,1090,1163, 1164,1165,709, 737, 689,1166, 741, 1167 | P01834 | Ig κ chain C region | IGKC | 80 | 8 | 8 | 11,609 | 6.11 | 0.0009–0.014 | 0.67–0.78 |
| 1162,1090,1163, 1164,1165,709, 737, 689,1166, 741, 1167 | P0CG05, P0CG06 | Ig λ chain C regions, Iso 2, 3 | IGLC2, IGLC3 | 24 | 2 | 2 | 11,294 11,237 | 6.91 6.91 | 0.0009–0.014 | 0.67–0.78 |
| 1166, 741 | P01598, P01602 | Ig κ chain V-I region EU, HK102 (Fragment) | KV106 KV110 | 32 | 3 | 2 | 11,788 | 8.49 | 0.0009–0.004 | 0.78 |
| 239, 243,246 | P01876, P01877 | Ig α chain C, Iso 1, 2 | IGHA1 IGHA2 | 20 | 6 | 4 | 37,655 | 6.08 | 0.05 | 0.74–0.86 |
| # | ID | Protein | Gene | Cov | Pep | Unic | MW (th) | pI (th) | p-Value (min–max) | Ratio (T1/T0) (min–max) |
|---|---|---|---|---|---|---|---|---|---|---|
| 75, 78, 81 | P01833 | Polymeric immunoglobulin receptor | PIGR | 32 | 26 | 26 | 83,284 | 5.59 | 0.001–0.040 | 0.78–0.86 |
| 170, 192 | P02768 | Serum albumin | ALB | 63 | 56 | 56 | 69,367 | 5.67 | 0.024–0.027 | 0.77–0.83 |
| 216, 246, 247, 250 | P01876, P01877 | Ig α chain C, Iso 1, 2 | IGHA1, IGHA2 | 16 | 6 | 5 | 37,655 | 6.08 | 0.001–0.031 | 0.74–0.86 |
| 271, 348 | P0DUB6 | α-amylase, 1A | AMY1A | 55 | 38 | 38 | 57,768 | 6.34 | 0.003–0.038 | 1.16–1.21 |
| 733 | P52565 | Rho GDP-dissociation inhibitor 1 | ARHGDIA | 29 | 6 | 6 | 23,207 | 5.01 | 0.019 | 0.80 |
| 756, 827 | P01591 | Immunoglobulin J chain | IGJ | 45 | 10 | 10 | 18,099 | 4.59 | 0.039 | 0.78 |
| 839 | P30086 | Phosphatidylethanolamine-binding protein 1 | PEBP1 | 43 | 9 | 9 | 21,057 | 7.43 | 0.042 | 0.74 |
| 1010 | Q01469 | Fatty acid-binding protein, epidermal | FABP5 | 68 | 17 | 10 | 15,033 | 6.82 | 0.032 | 1.70 |
| 1047 | P04080 | Cystatin B | CSTB | 52 | 6 | 4 | 11,139 | 6.96 | 0.026 | 0.66 |
| 1153 | P04075 | Fructose-bisphosphate aldolase A | ALDOA | 23 | 11 | 9 | 39,420 | 8.39 | 0.021 | 0.69 |
| 706,1161, 1162, 1090,1163,1164, 1166, 741 | P01834 | Ig κ chain C region | IGKC | 52 | 3 | 3 | 11,609 | 6.11 | 0.002–0.046 | 0.02–0.79 |
| 706, 1161, 1162, 1090,1163,1164, 1166, 741 | P0CG05, P0CG06 | Ig λchain C regions, Iso 2, 3 | IGLC2, IGLC3 | 24 | 2 | 2 | 11,294 | 6.91 | 0.002–0.046 | 0.02–0.79 |
| 1166, 741 | P01598, P01602 | Ig κ chain V-I region EU, HK102 (Fragment) | KV106, KV110 | 30 | 3 | 2 | 11,788 | 8.49 | 0.022–0.046 | 0.02–0.04 |
| 1154 | P07195 | L-lactate dehydrogenase B chain | LDHB | 22 | 8 | 6 | 36,639 | 5.72 | 0.0317 | 0.65 |
| Upstream Regulator | Molecule Type | z-Score Control | z-Score Odor | p-Value | Target Molecules Control Group |
|---|---|---|---|---|---|
| methylprednisolone | chemical drug | 1.784 | 2.646 | 1.1 × 10−9 | GSTP1,HSPA1A/HSPA1B,IGHG1,IGHG2,IGHM,IGKC,IGKV1-5,IGLC2,IGLC3,PIGR,SERPINB1 |
| lipopolysaccharide | chemical drug | −3.217 | −1.124 | 5.25 × 10−9 | ALB,AZGP1,C3,GSTP1,HSPA1A/HSPA1B,HSPA8,IGHG1,IGHM,IGKC,JCHAIN,LDHA,MDH1,PIGR,S100A9,SERPINB1,TF |
| TNF | cytokine | −1.61 | −0.656 | 1.26 × 10−6 | ACTB,ALB,ARHGDIB,C3,GSTP1,HSPA1A/HSPA1B,HSPA8,IGKC,LDHA,PIGR,S100A9,SERPINB1,TF |
| IL4 | cytokine | −2.214 | N/A | 1.11 × 10−5 | ACTB,ACTG1,C3,HSPA1A/HSPA1B,IGHG1,IGHG2,JCHAIN,PIGR,S100A9 |
| β-estradiol | chemical endogenous | −1.937 | −1.406 | 3.66 × 10−5 | ACTB,ALB,C3,CST2,CST4,GSTP1,HSPA8,LDHA,PIGR,S100A9,TF,YWHAZ |
| KDM1A | enzyme | 2 | N/A | 0.000244 | ARHGDIB,AZGP1,LCP1,SERPINB1 |
| PRDM1 | transcription regulator | −1.98 | N/A | 0.00056 | IGHG1,IGHM,JCHAIN,S100A9 |
| IL1B | cytokine | −2.363 | N/A | 0.00281 | C3,HSPA1A/HSPA1B,LCP1,LDHA,PIGR,S100A9 |
| CSF2 | cytokine | −1.955 | N/A | 0.00765 | C3,LCP1,LDHA,TF |
| EGF | growth factor | −1.964 | N/A | 0.00826 | GSTP1,LDHA,S100A9,TF |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zallocco, L.; Giusti, L.; Ronci, M.; Mussini, A.; Trerotola, M.; Mazzoni, M.R.; Lucacchini, A.; Sebastiani, L. Salivary Proteome Changes in Response to Acute Psychological Stress Due to an Oral Exam Simulation in University Students: Effect of an Olfactory Stimulus. Int. J. Mol. Sci. 2021, 22, 4295. https://doi.org/10.3390/ijms22094295
Zallocco L, Giusti L, Ronci M, Mussini A, Trerotola M, Mazzoni MR, Lucacchini A, Sebastiani L. Salivary Proteome Changes in Response to Acute Psychological Stress Due to an Oral Exam Simulation in University Students: Effect of an Olfactory Stimulus. International Journal of Molecular Sciences. 2021; 22(9):4295. https://doi.org/10.3390/ijms22094295
Chicago/Turabian StyleZallocco, Lorenzo, Laura Giusti, Maurizio Ronci, Andrea Mussini, Marco Trerotola, Maria Rosa Mazzoni, Antonio Lucacchini, and Laura Sebastiani. 2021. "Salivary Proteome Changes in Response to Acute Psychological Stress Due to an Oral Exam Simulation in University Students: Effect of an Olfactory Stimulus" International Journal of Molecular Sciences 22, no. 9: 4295. https://doi.org/10.3390/ijms22094295
APA StyleZallocco, L., Giusti, L., Ronci, M., Mussini, A., Trerotola, M., Mazzoni, M. R., Lucacchini, A., & Sebastiani, L. (2021). Salivary Proteome Changes in Response to Acute Psychological Stress Due to an Oral Exam Simulation in University Students: Effect of an Olfactory Stimulus. International Journal of Molecular Sciences, 22(9), 4295. https://doi.org/10.3390/ijms22094295