
Evaluation of Selective COX-2 Inhibition and In Silico Study of Kuwanon Derivatives Isolated from Morus alba

 , ,
, , 
Abstract
1. Introduction
2. Results
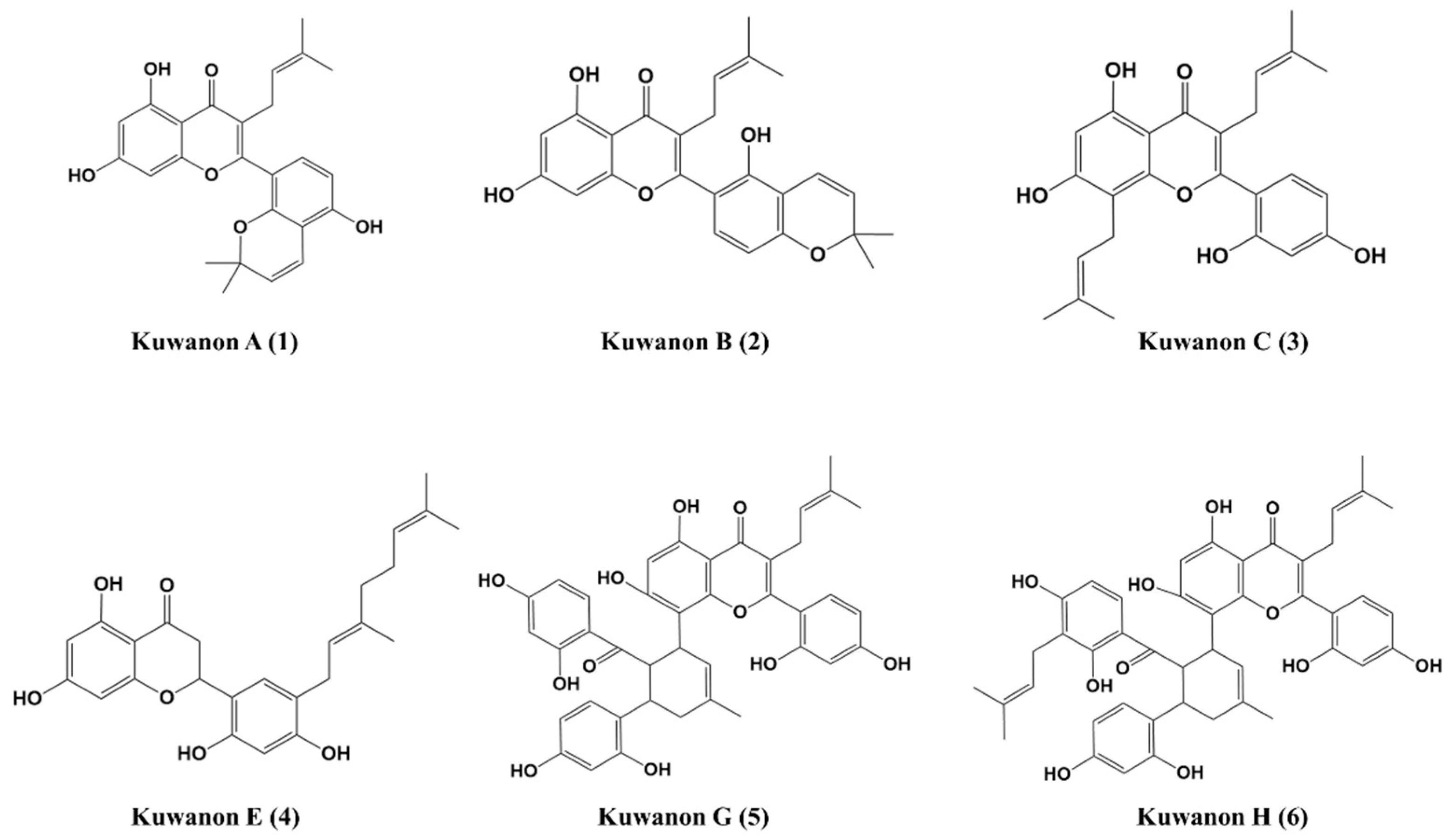
2.1. Isolation and Identification of Compounds
2.2. Analysis of COX-1/2 Enzyme Inhibitory Activity
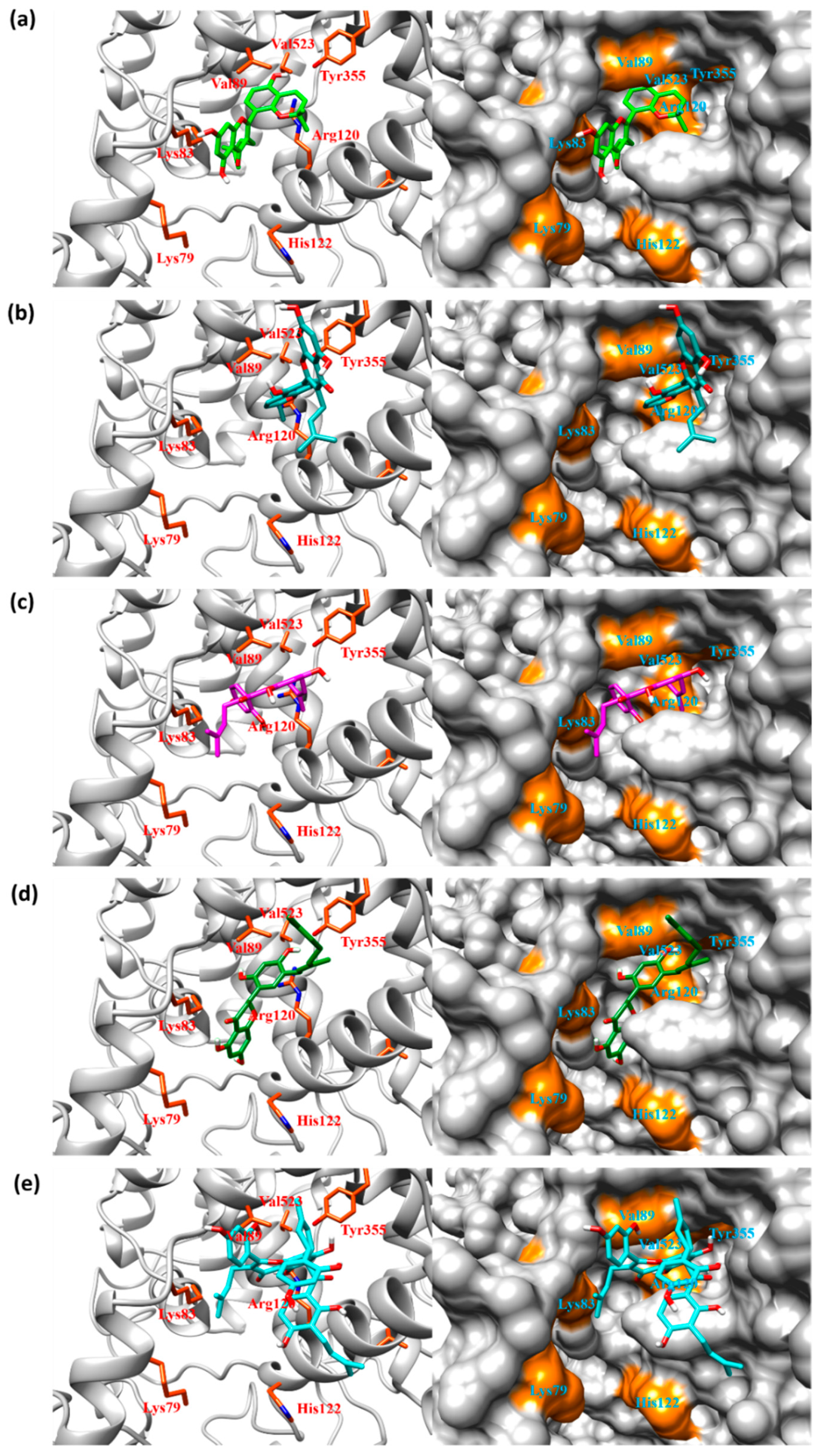
2.3. Docking Simulation
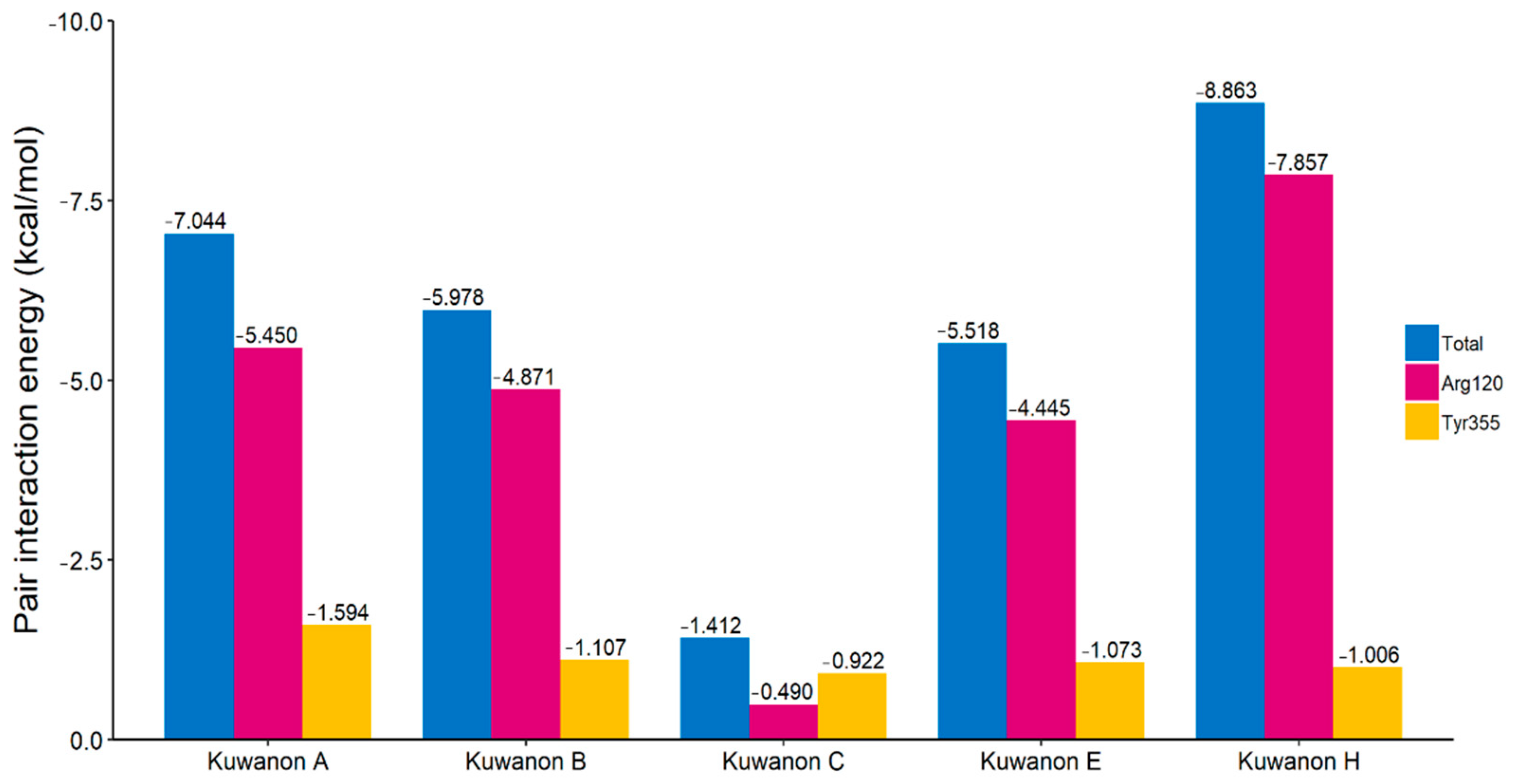
2.4. Interaction Energy in the Gate of COX Active Site
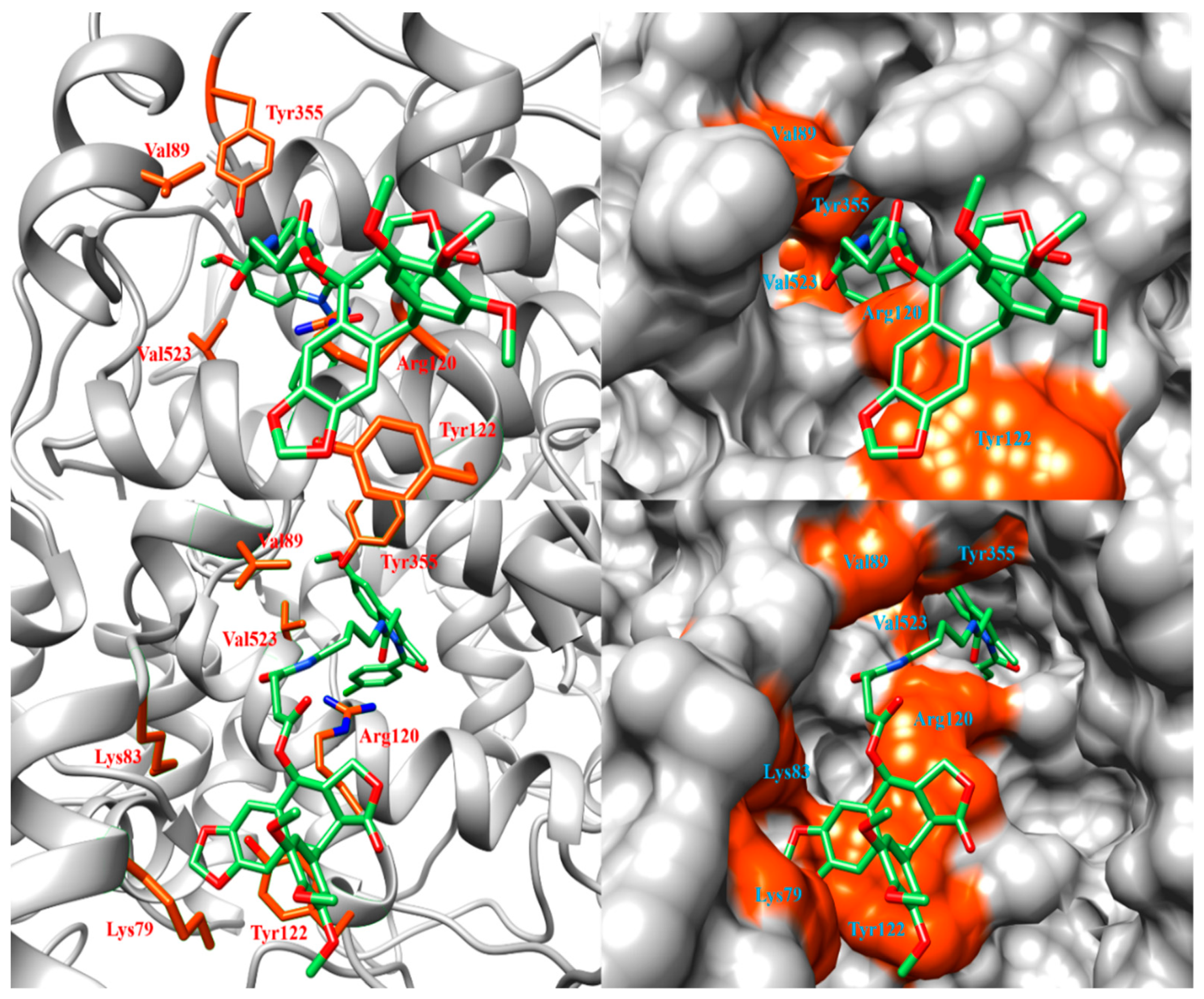
2.5. Interaction Energy in the Membrane-Binding Domain and the Entrance/Exit Routes of COX Active Sites
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Extraction and Isolation
4.3. Assay Materials
4.4. In Vitro COX-1/COX-2 Inhibition Assay
4.5. Docking Simulation
4.6. FMO Calculation and PIEDA
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alam, M.J.; Alam, O.; Khan, S.A.; Naim, M.J.; Islamuddin, M.; Deora, G.S. Synthesis, anti-inflammatory, analgesic, COX1/2-inhibitory activity, and molecular docking studies of hybrid pyrazole analogues. Drug Des. Devel. Ther. 2016, 10, 3529–3543. [Google Scholar] [CrossRef]
- Zidar, N.; Odar, K.; Glavac, D.; Jerse, M.; Zupanc, T.; Stajer, D. Cyclooxygenase in normal human tissues—Is COX-1 really a constitutive isoform, and COX-2 an inducible isoform? J. Cell. Mol. Med. 2009, 13, 3753–3763. [Google Scholar] [CrossRef]
- Choi, S.-H.; Aid, S.; Bosetti, F. The distinct roles of cyclooxygenase-1 and-2 in neuroinflammation: Implications for translational research. Trends Pharmacol. Sci. 2009, 30, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Willoughby, D.A.; Moore, A.R.; Colville-Nash, P.R. COX-1, COX-2, and COX-3 and the future treatment of chronic inflammatory disease. Lancet 2000, 355, 646–648. [Google Scholar] [CrossRef]
- Pairet, M.; Van Ryn, J.; Mauz, A.; Schierok, H.; Diederen, W.; Türck, D.; Engelhardt, G. Differential inhibition of COX-1 and COX-2 by NSAIDs: A summary of results obtained using various test systems. In Selective COX-2 Inhibitors; Springer: Berlin/Heidelberg, Germany, 1998; pp. 27–46. [Google Scholar]
- Cryer, B.; Feldman, M. Cyclooxygenase-1 and cyclooxygenase-2 selectivity of widely used nonsteroidal anti-inflammatory drugs. Am. J. Med. 1998, 104, 413–421. [Google Scholar] [CrossRef]
- Bagachi, A.; Semwal, A.; Bharadwaj, A. Traditional uses, phytochemistry and pharmacology of Morus alba Linn.: A review. J. Med. Plants Res. 2013, 7, 461–469. [Google Scholar]
- Ahmad, A.; Gupta, G.; Afzal, M.; Kazmi, I.; Anwar, F. Antiulcer and antioxidant activities of a new steroid from Morus alba. Life Sci. 2013, 92, 202–210. [Google Scholar] [CrossRef]
- Ali, A.; Ali, M. New triterpenoids from Morus alba L. stem bark. Nat. Prod. Res. 2013, 27, 524–531. [Google Scholar] [CrossRef]
- Asano, N.; Yamashita, T.; Yasuda, K.; Ikeda, K.; Kizu, H.; Kameda, Y.; Kato, A.; Nash, R.J.; Lee, H.S.; Ryu, K.S. Polyhydroxylated alkaloids isolated from mulberry trees (Morus alba L.) and silkworms (Bombyx mori L.). J. Agric. Food Chem. 2001, 49, 4208–4213. [Google Scholar] [CrossRef]
- Qiu, F.; Komatsu, K.; Kawasaki, K.; Saito, K.; Yao, X.; Kano, Y. A novel stilbene glucoside, oxyresveratrol 3′-O-β-glucopyranoside, from the root bark of Morus alba. Planta Med. 1996, 62, 559–561. [Google Scholar] [CrossRef]
- Oh, H.; Ko, E.-K.; Jun, J.-Y.; Oh, M.-H.; Park, S.-U.; Kang, K.-H.; Lee, H.-S.; Kim, Y.-C. Hepatoprotective and free radical scavenging activities of prenylflavonoids, coumarin, and stilbene from Morus alba. Planta Med. 2002, 68, 932–934. [Google Scholar] [CrossRef]
- Nomura, T.; Fukal, T.; Katayanagi, M. Studies on the constituents of the cultivated mulberry tree. III. Isolation of four new flavones, kuwanon A, B, C and oxydihydromorusin from the root bark of Morus alba L. Chem. Pharm. Bull. 1978, 26, 1453–1458. [Google Scholar] [CrossRef]
- Nomura, T. Hypotensive constituent, kuwanon H, a new flavone derivative from the root bark of the cultivated mulberry tree (Morus alba L.). Heterocycles 1980, 14, 1943–4951. [Google Scholar] [CrossRef]
- Nomura, T.; Fukai, T. Kuwanon G, a new flavone derivative from the root barks of the cultivated mulberry tree (Morus alba L.). Chem. Pharm. Bull. 1980, 28, 2548–2552. [Google Scholar] [CrossRef]
- Woo, H.; Lee, J.; Park, D.; Jung, E. Protective effect of mulberry (Morus alba L.) extract against benzo [a] pyrene induced skin damage through inhibition of aryl hydrocarbon receptor signaling. J. Agric. Food Chem. 2017, 65, 10925–10932. [Google Scholar] [CrossRef] [PubMed]
- Rayam, S.; Kudagi, B.L.; Buchineni, M.; Pathapati, R.M.; Immidisetty, M.R. Assessment of Morus alba (mulberry) leaves extract for anti-psychotic effect in rats. Int. J. Basic Clin. Pharmacol. 2019, 8, 2130–2133. [Google Scholar] [CrossRef]
- Wei, H.; Zhu, J.-J.; Liu, X.-Q.; Feng, W.-H.; Wang, Z.-M.; Yan, L.-H. Review of bioactive compounds from root barks of Morus plants (Sang-Bai-Pi) and their pharmacological effects. Cogent Chem. 2016, 2, 1212320. [Google Scholar] [CrossRef]
- Kim, J.-M.; Baek, J.-M.; Kim, H.-S.; Choe, M. Antioxidative and anti-asthma effect of Morus bark water extracts. J. Korean Soc. Food Sci. Nutr. 2010, 39, 1263–1269. [Google Scholar] [CrossRef]
- Čulenová, M.; Sychrová, A.; Hassan, S.T.S.; Berchová-Bímová, K.; Svobodová, P.; Helclová, A.; Michnová, H.; Hošek, J.; Vasilev, H.; Suchý, P. Multiple In vitro biological effects of phenolic compounds from Morus alba root bark. J. Ethnopharmacol. 2020, 248, 112296. [Google Scholar] [CrossRef] [PubMed]
- Blobaum, A.L.; Xu, S.; Rowlinson, S.W.; Duggan, K.C.; Banerjee, S.; Kudalkar, S.N.; Birmingham, W.R.; Ghebreselasie, K.; Marnett, L.J. Action at a Distance: Action at a Distance: Mutations of Peripheral Residues Transform Rapid Reversible Inhibitors to Slow, Tight Binders of Cyclooxygenase-2. J. Biol. Chem. 2015, 290, 12793–12803. [Google Scholar] [CrossRef]
- Knights, K.M.; Mangoni, A.A.; Miners, J.O. Defining the COX inhibitor selectivity of NSAIDs: Implications for understanding toxicity. Expert Rev. Clin. Pharmacol. 2010, 3, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Simmons, D.L.; Botting, R.M.; Hla, T. Cyclooxygenase Isozymes: The Biology of Prostaglandin Synthesis and Inhibition. Pharmacol. Rev. 2004, 56, 387–437. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.J.; Crews, B.C.; Xu, S.; Ghebreselasie, K.; Daniel, C.K.; Kingsley, P.J.; Banerjee, S.; Marnett, L.J. Antitumor Activity of Cytotoxic Cyclooxygenase-2 Inhibitors. ACS Chem. Biol. 2016, 11, 3052–3060. [Google Scholar] [CrossRef] [PubMed]
- Kitaura, K.; Ikeo, E.; Asada, T.; Nakano, T.; Uebayasi, M. Fragment molecular orbital method: An approximate computational method for large molecules. Chem. Phys. Lett. 1999, 313, 701–706. [Google Scholar] [CrossRef]
- Fedorov, D.G.; Kitaura, K. Pair interaction energy decomposition analysis. J. Comput. Chem. 2007, 28, 222–237. [Google Scholar] [CrossRef] [PubMed]
- Gaus, M.; Cui, Q.; Elstner, M. DFTB3: Extension of the Self-Consistent-Charge Density-Functional Tight-Binding Method (SCC-DFTB). J. Chem. Theory Comput. 2011, 7, 931–948. [Google Scholar] [CrossRef] [PubMed]
- Ryu, T.-B.; Ha, T.-J.; Curtis-Long, M.-J.; Ryu, H.-W.; Gap, S.-W.; Park, K.-H. Inhibitory effects on mushroom tyrosinase by flavones from the stem barks of Morus lhou (S.) Koidz. J. Enz. Inhibi. Medici. Chem. 2008, 23, 922–930. [Google Scholar] [CrossRef]
- Nomura, T.; Fukai, T.; Katayanagi, M. Kuwanon A, B, C and oxydihydromorusin, four new flavones from the root bark of the cultivated mulberry tree (Morus alba L.). Chem. Pharm. Bull. 1977, 25, 529–532. [Google Scholar] [CrossRef]
- Tang, F.; Wang, Y.; Hou, A.-J. Total syntheses of Nigrasin I and Kuwanon C. Tetrahedron 2014, 70, 3963–3970. [Google Scholar] [CrossRef]
- Nomura, T.; Fukai, T.; Hano, Y.; Nemoto, K.; Terada, S.; Kuramochi, T. Constituents of cultivated mulberry tree. Planta Med. 1983, 47, 151–156. [Google Scholar] [CrossRef]
- Nomura, T.; Fukai, T.; Narita, T.; Terada, S.; Uzawa, J.; Iitaka, Y.; Takasugi, M.; Ishikawa, S.; Nagao, S.; Masamune, T. Confirmation of the structures of kuwanons G and H (albanins F and G) by partial synthesis. Tetrahedron Lett. 1981, 22, 2195–2198. [Google Scholar] [CrossRef]
- Al-Saeed, A. Gastrointestinal and cardiovascular risk of nonsteroidal anti-inflammatory drugs. Oman Med. J. 2011, 26, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Furse, K.E.; Pratt, D.A.; Porter, N.A.; Lybrand, T.P. Molecular Dynamics Simulations of Arachidonic Acid Complexes with COX-1 and COX-2: Insights into Equilibrium Behavior. Biochemistry 2006, 45, 3189–3205. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Landrum, G. RDKit: Open-Source Cheminformatics 2006. Available online: https://www.bibsonomy.org/bibtex/28d01fceeccd6bf2486e47d7c4207b108/salotz (accessed on 15 August 2020).
- Barca, G.M.J.; Bertoni, C.; Carrington, L.; Datta, D.; De Silva, N.; Deustua, J.E.; Fedorov, D.G.; Gour, J.R.; Gunina, A.O.; Guidez, E.; et al. Recent developments in the general atomic and molecular electronic structure system. J. Chem. Phys. 2020, 152, 154102. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | IC50 (μM) 1 | SI 2 | |
|---|---|---|---|
| COX-1 | COX-2 | ||
| Kuwanon A | >100 | 14 | >7.1 |
| Kuwanon B | 36 | 28 | 1.3 |
| Kuwanon C | 67 | 42 | 1.6 |
| Kuwanon E | 46 | 34 | 1.4 |
| Kuwanon G | >100 | >100 | - 3 |
| Kuwanon H | 37 | 7 | 5.0 |
| Celecoxib | >100 | 16 | >6.3 |
| Name | Molecular Weight (g/mol) | Molecular Volume |
|---|---|---|
| Celecoxib | 381.076 | 359.848 |
| Chemcoxib A | 923.303 | 891.808 |
| Indomethacin | 357.077 | 350.384 |
| Podophyllotoxin | 456.142 | 434.096 |
| Kuwanon A | 420.157 | 417.768 |
| Kuwanon B | 420.157 | 413.624 |
| Kuwanon C | 422.173 | 426.152 |
| Kuwanon E | 424.189 | 427.352 |
| Kuwanon G | 692.226 | 662.832 |
| Kuwanon H | 760.288 | 744.432 |
| Compound Name | Amino Acid | Pair Interaction Energy (kcal/mol) | Component of Pair Interaction Energy (kcal/mol) c | ||||
|---|---|---|---|---|---|---|---|
| ES | EX | CT | DI | SL | |||
| Chemocoxib A a | Lys79 | −15.267 | −1.364 | −0.126 | −0.195 | −5.647 | −7.936 |
| Lys83 | −10.241 | −2.342 | 0.303 | −0.257 | −3.839 | −4.105 | |
| Tyr122 | −6.049 | 0.502 | 0.104 | 0.000 | −6.321 | −0.334 | |
| Leu117 | −3.464 | −3.101 | 0.001 | 0.000 | −0.565 | 0.201 | |
| Val89 | −3.267 | 0.018 | −0.072 | −0.002 | −3.200 | −0.012 | |
| Kuwanon A b | Lys79 | −1.807 | −2.344 | 0.000 | 0.000 | −0.531 | 1.068 |
| Lys83 | −14.430 | −7.995 | 0.782 | −0.152 | −8.657 | 1.591 | |
| His122 | −0.830 | −0.825 | 0.000 | 0.000 | −0.258 | 0.253 | |
| Leu117 | −0.934 | −1.120 | 0.756 | −0.022 | −0.609 | 0.061 | |
| Val89 | −6.599 | −0.771 | 0.532 | −0.016 | −6.172 | −0.172 | |
| Kuwanon B b | Lys79 | −1.468 | 0.839 | 0.000 | 0.000 | −0.037 | −2.271 |
| Lys83 | −1.721 | 4.021 | 0.000 | 0.000 | −0.503 | −5.239 | |
| His122 | −0.096 | 0.069 | 0.000 | 0.000 | −0.051 | −0.114 | |
| Leu117 | −0.024 | −0.103 | 0.000 | 0.000 | −0.370 | 0.449 | |
| Val89 | −4.665 | 1.035 | 0.217 | −0.011 | −4.866 | −1.040 | |
| Kuwanon C b | Lys79 | −3.164 | 0.158 | −0.003 | 0.000 | −1.035 | −2.285 |
| Lys83 | −14.541 | −7.330 | 3.374 | −0.198 | −7.612 | −2.775 | |
| His122 | −0.830 | −0.825 | 0.000 | 0.000 | −0.258 | 0.253 | |
| Leu117 | −0.934 | −1.120 | 0.756 | −0.022 | −0.609 | 0.061 | |
| Val89 | 4.212 | −0.154 | 8.521 | −0.128 | −3.934 | −0.091 | |
| Kuwanon E b | Lys79 | −2.662 | −5.799 | 0.000 | 0.000 | −0.382 | 3.518 |
| Lys83 | −8.604 | −3.086 | 0.050 | −0.001 | −5.367 | −0.200 | |
| His122 | −1.933 | −1.678 | −0.001 | 0.000 | −0.775 | 0.521 | |
| Leu117 | −0.199 | −0.139 | 0.000 | 0.000 | −0.351 | 0.291 | |
| Val89 | −5.810 | −0.328 | 0.563 | −0.023 | −5.754 | −0.267 | |
| Kuwanon H b | Lys79 | −3.997 | −0.092 | 0.000 | 0.000 | −0.434 | −3.471 |
| Lys83 | −10.573 | −6.387 | 1.361 | −0.033 | −4.870 | −0.645 | |
| His122 | −0.515 | −0.150 | 0.000 | 0.000 | −0.088 | −0.277 | |
| Leu117 | 1.646 | 2.200 | 0.000 | 0.000 | −0.349 | −0.205 | |
| Val89 | −6.257 | 0.117 | 0.334 | −0.002 | −6.521 | −0.185 | |
| Residues | Closest Distance (Å) | ||||
|---|---|---|---|---|---|
| Kuwanon A | Kuwanon B | Kuwanon C | Kuwanon E | Kuwanon H | |
| Val89 | 2.595 | 2.195 | 1.404 | 1.961 | 2.142 |
| His90 | 3.951 | 5.109 | 5.205 | 3.664 | 4.857 |
| Ser119 | 6.440 | 7.132 | 6.301 | 7.665 | 3.778 |
| Average | 4.329 | 4.812 | 4.303 | 4.430 | 3.592 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baek, S.-H.; Hwang, S.; Park, T.; Kwon, Y.-J.; Cho, M.; Park, D. Evaluation of Selective COX-2 Inhibition and In Silico Study of Kuwanon Derivatives Isolated from Morus alba. Int. J. Mol. Sci. 2021, 22, 3659. https://doi.org/10.3390/ijms22073659
Baek S-H, Hwang S, Park T, Kwon Y-J, Cho M, Park D. Evaluation of Selective COX-2 Inhibition and In Silico Study of Kuwanon Derivatives Isolated from Morus alba. International Journal of Molecular Sciences. 2021; 22(7):3659. https://doi.org/10.3390/ijms22073659
Chicago/Turabian StyleBaek, Seung-Hwa, Sungbo Hwang, Tamina Park, Yoon-Ju Kwon, Myounglae Cho, and Daeui Park. 2021. "Evaluation of Selective COX-2 Inhibition and In Silico Study of Kuwanon Derivatives Isolated from Morus alba" International Journal of Molecular Sciences 22, no. 7: 3659. https://doi.org/10.3390/ijms22073659
APA StyleBaek, S.-H., Hwang, S., Park, T., Kwon, Y.-J., Cho, M., & Park, D. (2021). Evaluation of Selective COX-2 Inhibition and In Silico Study of Kuwanon Derivatives Isolated from Morus alba. International Journal of Molecular Sciences, 22(7), 3659. https://doi.org/10.3390/ijms22073659