
Exploring the Crosstalk between Hydrostatic Pressure and Adipokines: An In Vitro Study on Human Osteoarthritic Chondrocytes

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
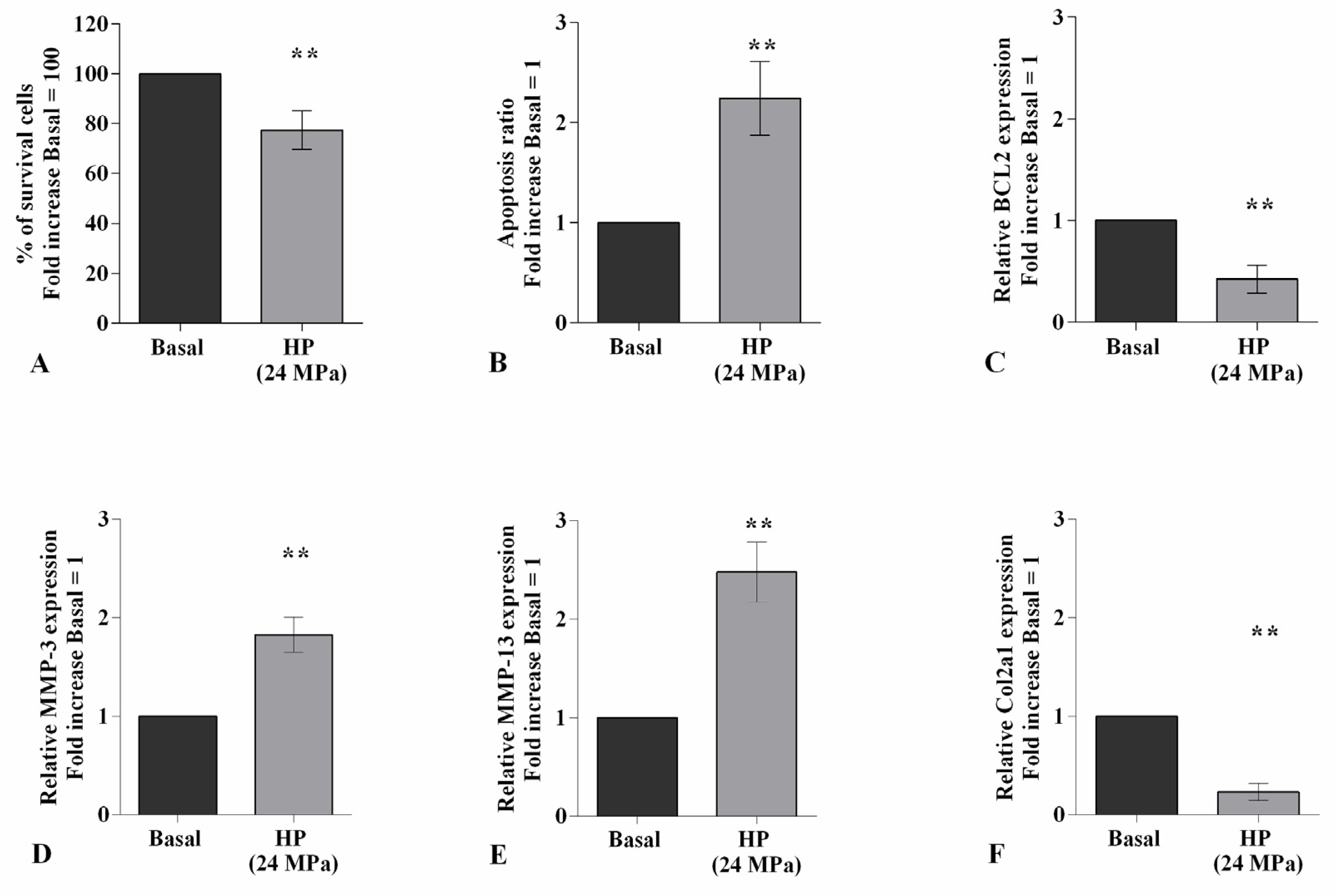
2.1. HP Regulates Cellular Apoptosis and Cartilage Turnover
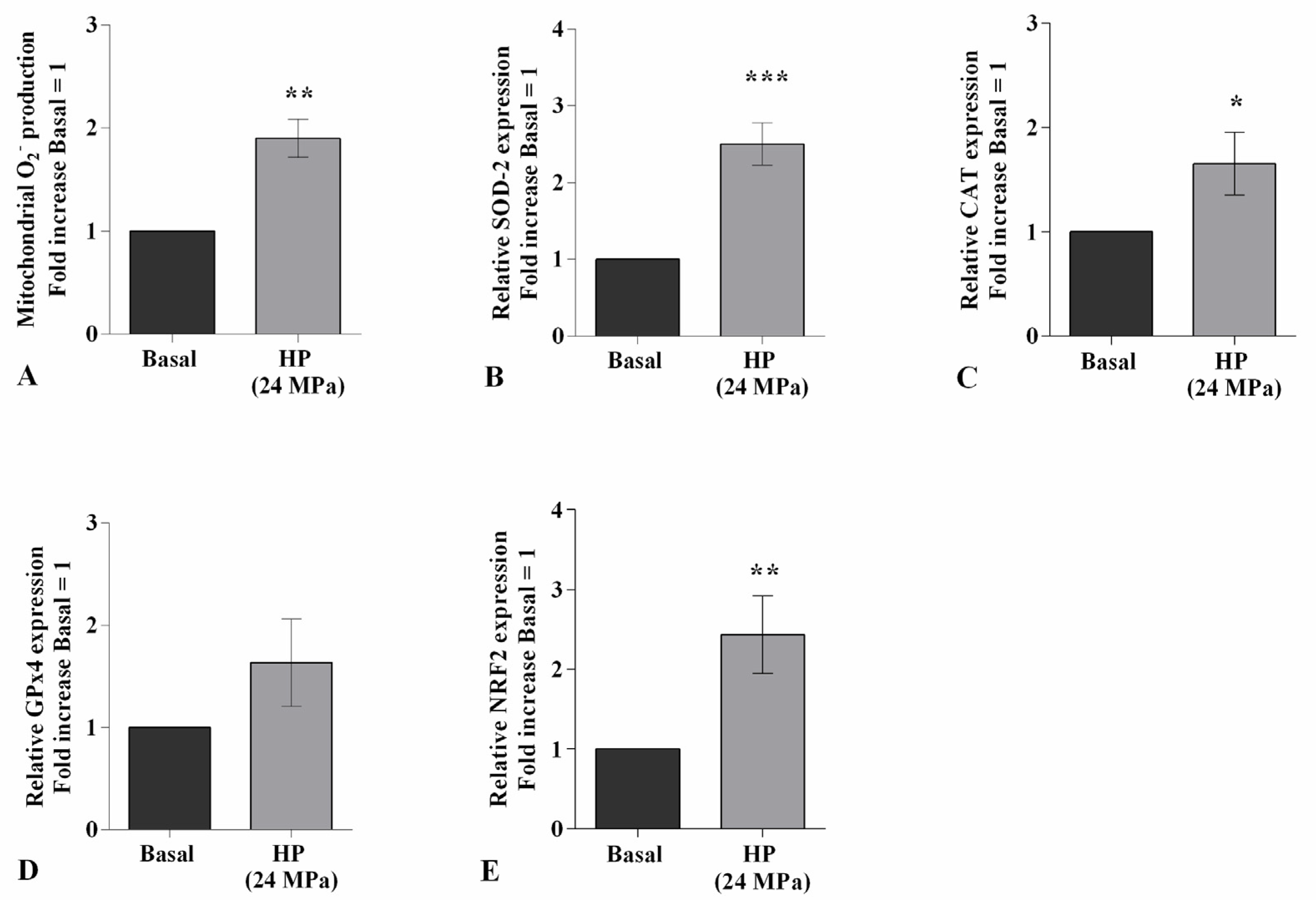
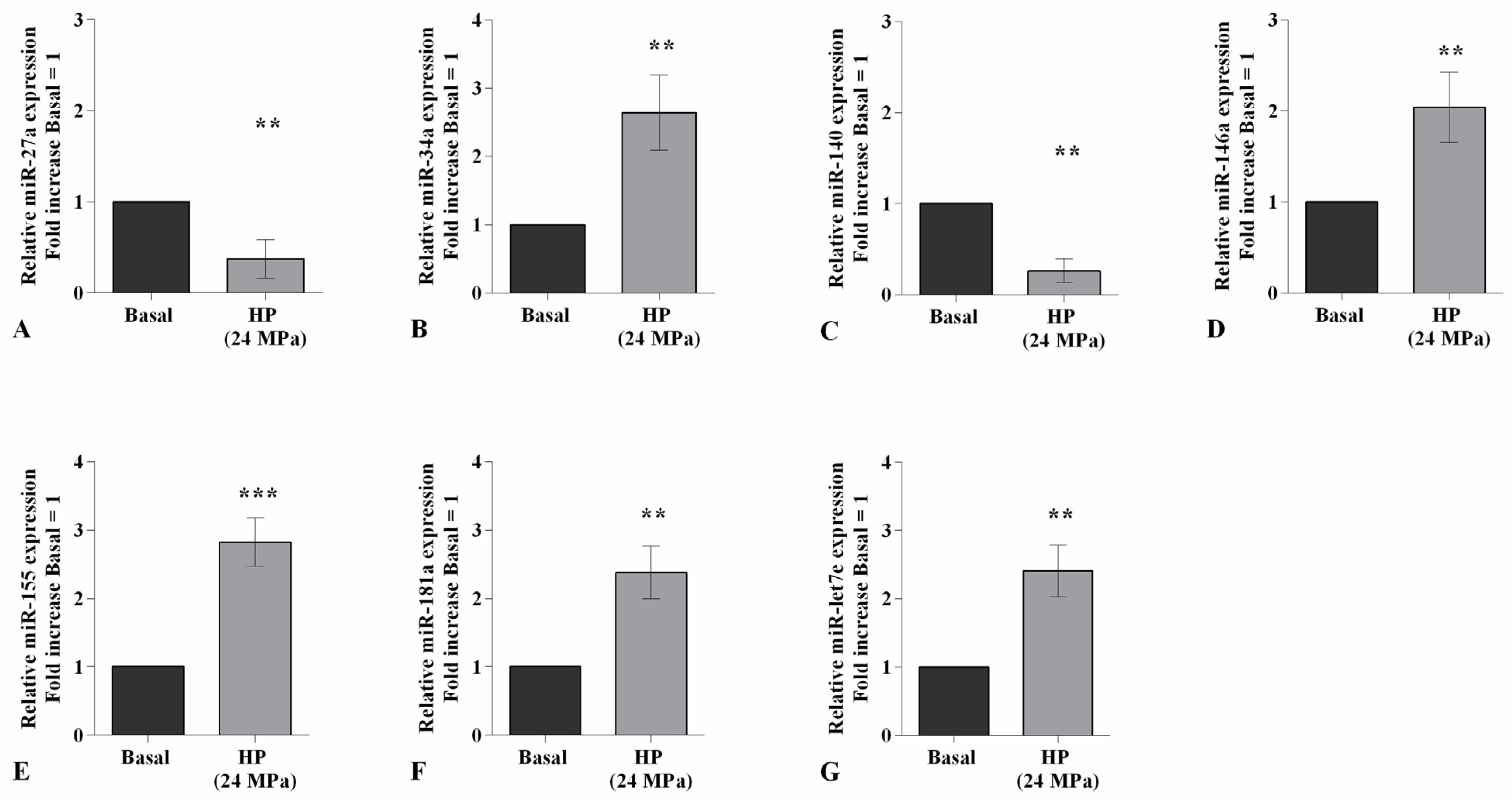
2.2. HP Influences Oxidative Stress Balance and miRNA Expression Profile
2.3. Visfatin Induces Cellular Apoptosis and Regulates Cartilage Turnover
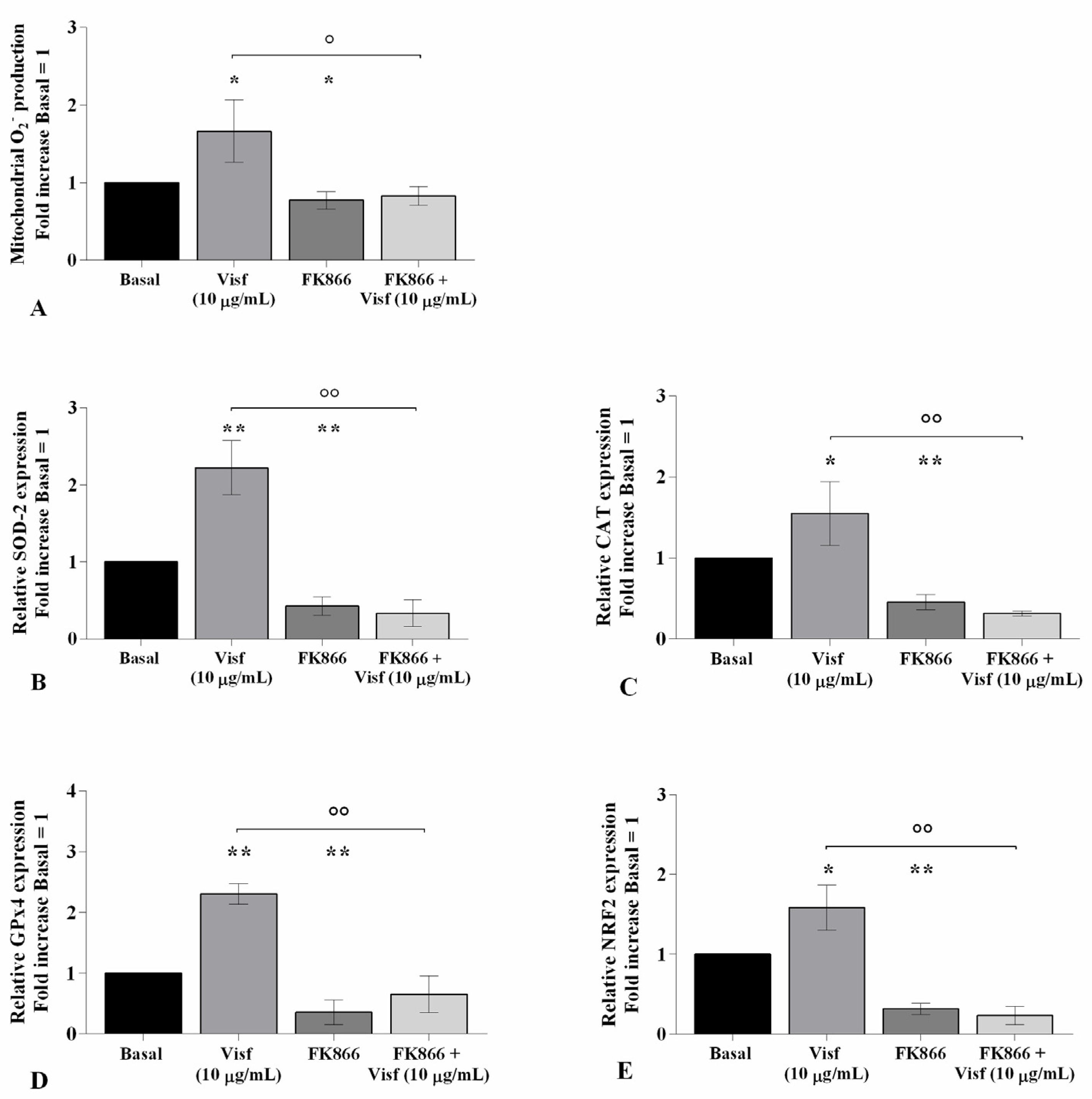
2.4. Visfatin Modulates Oxidant/Antioxidant System and miRNA Expression Profile
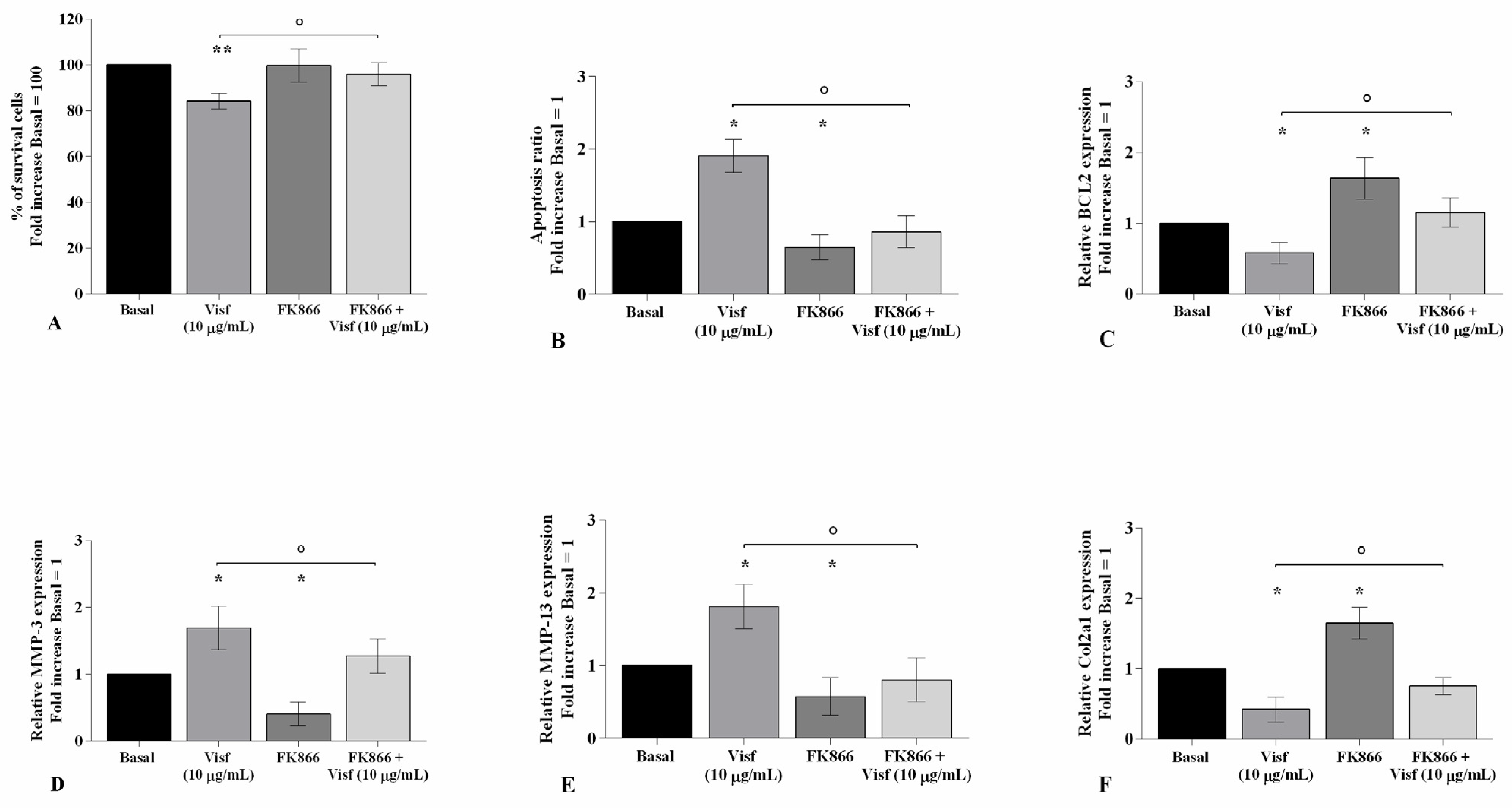
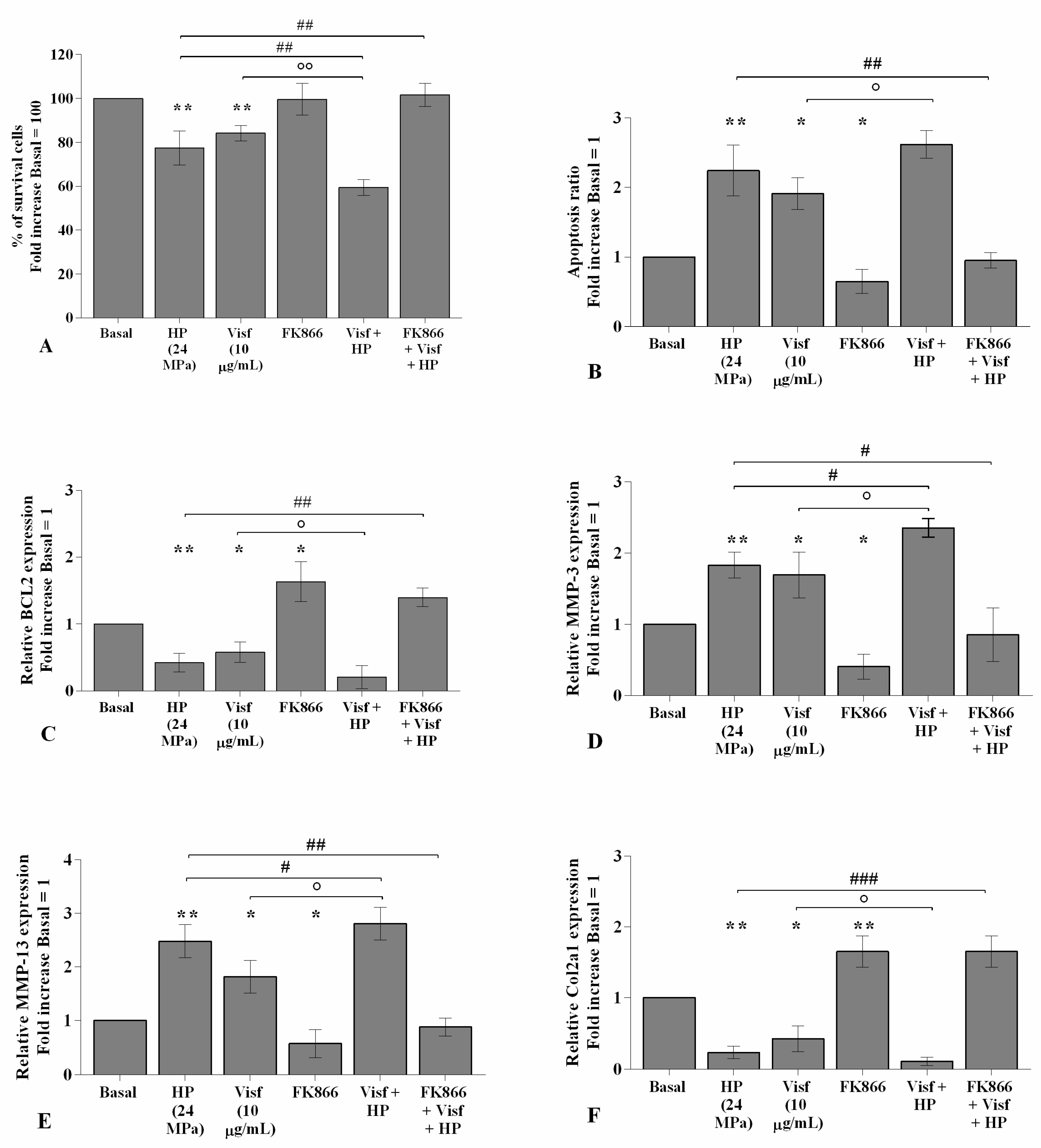
2.5. HP Increases Cellular Apoptosis and Cartilage Damage Caused by Visfatin
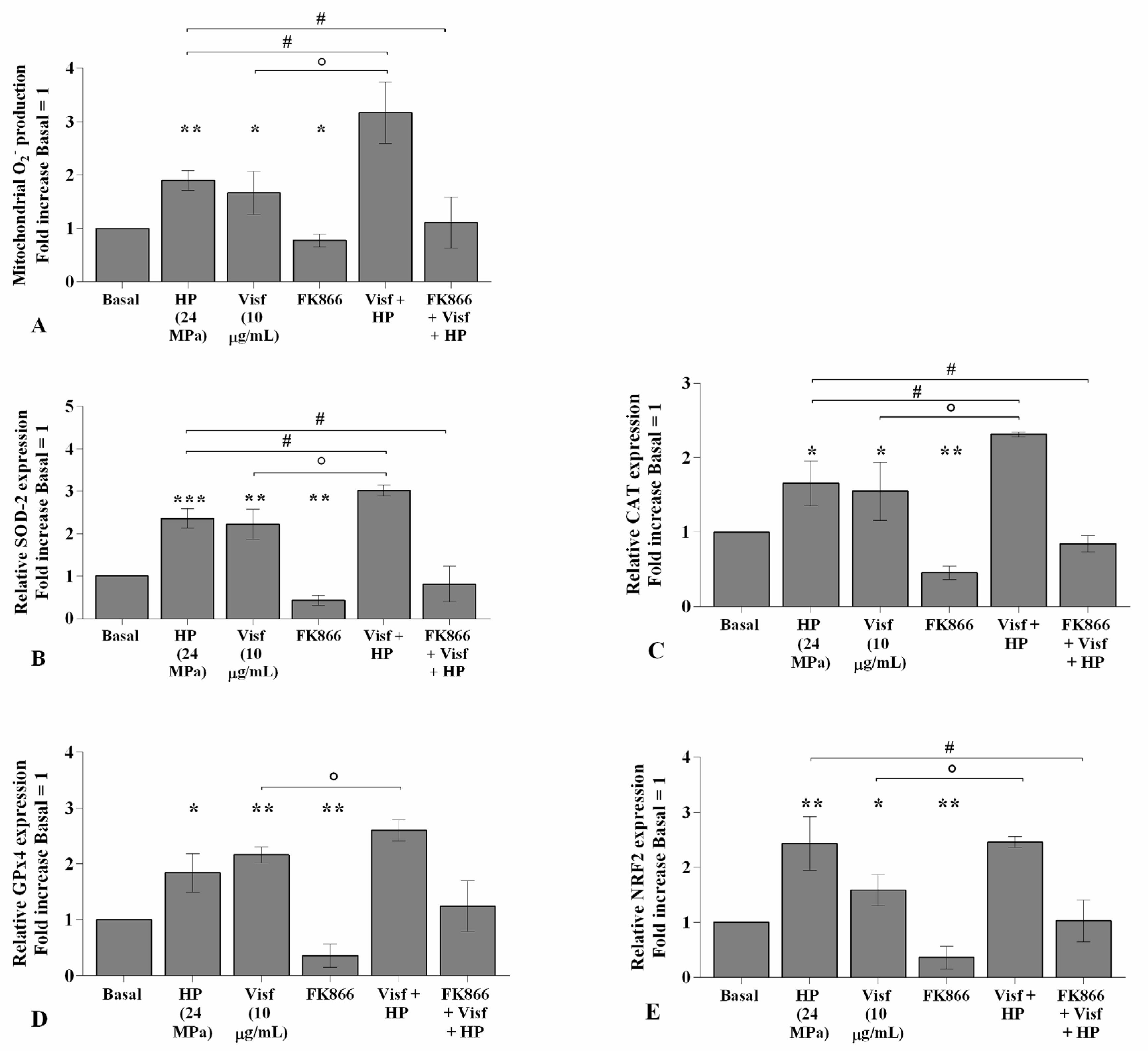
2.6. HP Exacerbates Oxidative Stress Balance Caused by Visfatin
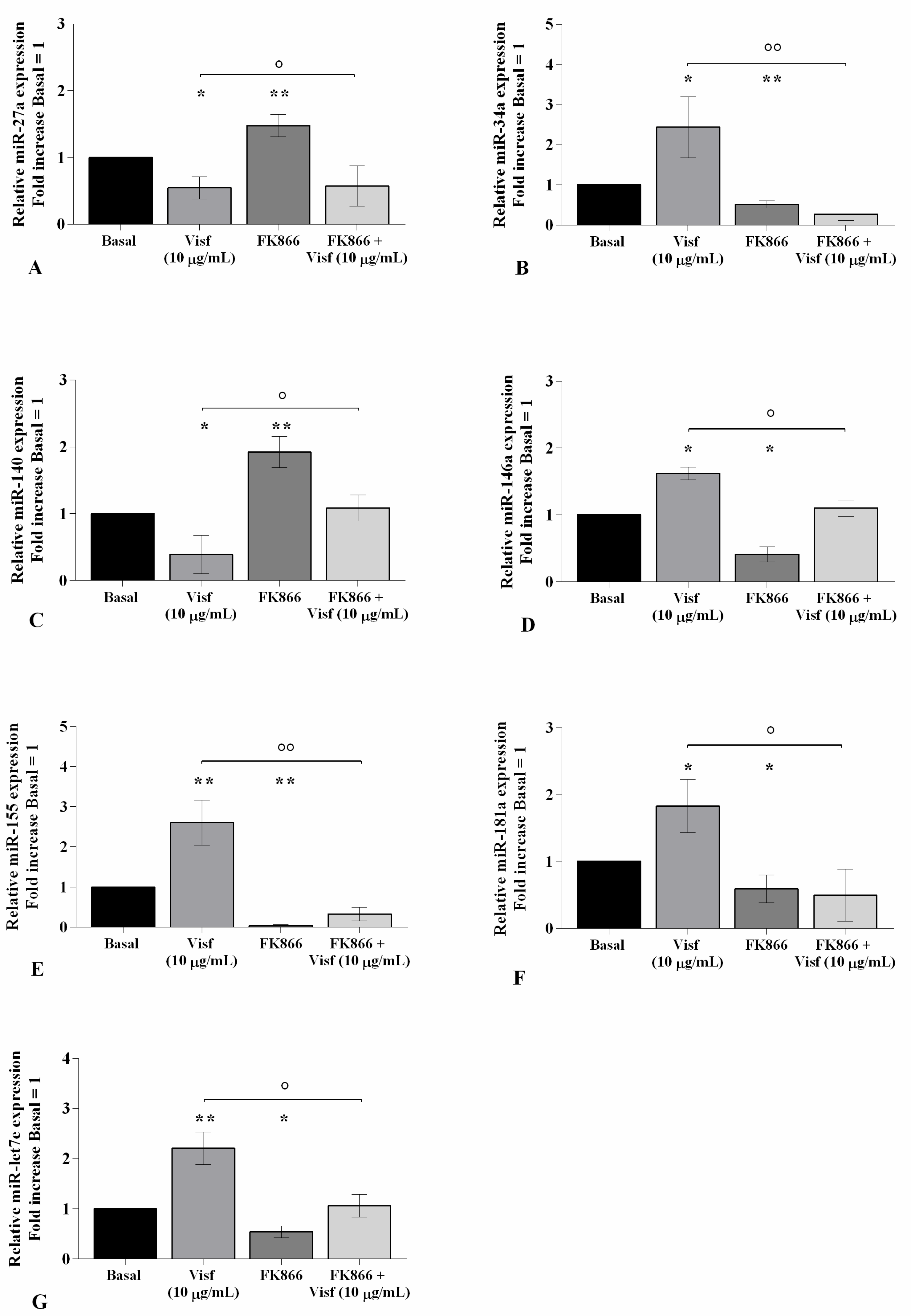
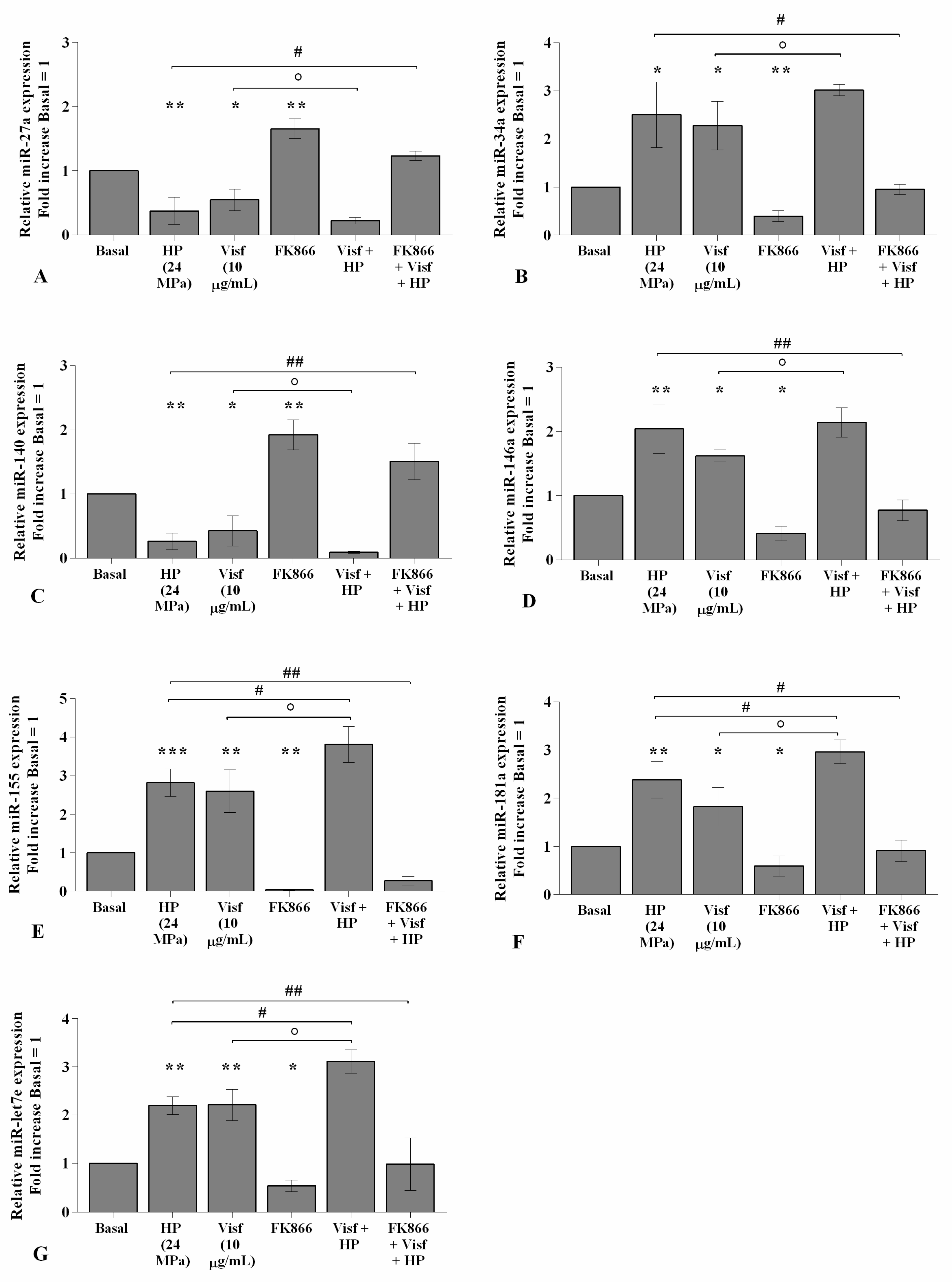
2.7. HP Enhances Visfatin Effect on miRNA Gene Expression Profile
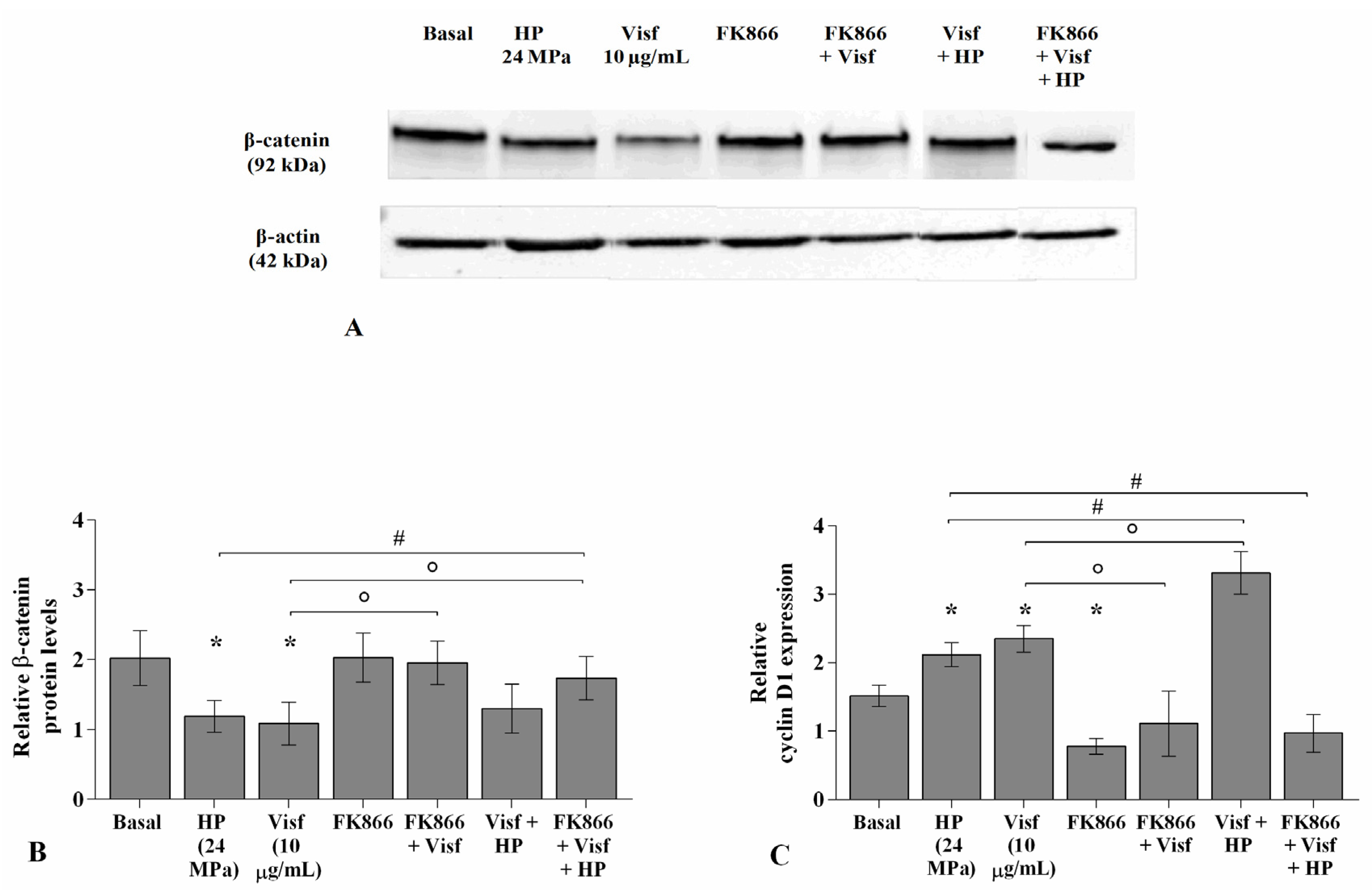
2.8. HP Influences the Regulation of the Wnt/β-Catenin Pathway Induced by Visfatin
3. Discussion
4. Materials and Methods
4.1. Isolation and Culture of Human OA Chondrocytes
4.2. OA Chondrocytes Exposure to HP
4.3. OA Chondrocytes Treatment
4.4. Cell Viability
4.5. Apoptosis Detection
4.6. Mitochondrial Superoxide Anion (·O2-) Assessment
4.7. RNA Isolation and Quantitative Real-Time PCR
4.8. Western Blot
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Hunter, D.J.; Bierma-Zeinstra, S. Osteoarthritis. Lancet 2019, 393, 1745–1759. [Google Scholar] [CrossRef]
- Issa, R.I.; Griffin, T.M. Pathobiology of Obesity and Osteoarthritis: Integrating Biomechanics and Inflammation. Pathobiol. Aging Age Relat. Dis. 2012, 2. [Google Scholar] [CrossRef]
- Francisco, V.; Pérez, T.; Pino, J.; López, V.; Franco, E.; Alonso, A.; Gonzalez-Gay, M.A.; Mera, A.; Lago, F.; Gómez, R.; et al. Biomechanics, Obesity, and Osteoarthritis. The role of Adipokines: When the Levee Breaks. J. Orthop. Res. 2018, 36, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Herger, S.; Vach, W.; Liphardt, A.M.; Egloff, C.; Nüesch, C.; Mündermann, A. Dose-response Relationship Between Ambulatory Load Magnitude and Load-induced Changes in COMP in Young Healthy Adults. Osteoarthr. Cartil. 2019, 27, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Hunt, M.A.; Charlton, J.M.; Esculier, J.F. Osteoarthritis Year in Review 2019: Mechanics. Osteoarthr. Cartil. 2020, 28, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Fioravanti, A.; Collodel, G.; Petraglia, A.; Nerucci, F.; Moretti, E.; Galeazzi, M. Effect of Hydrostatic Pressure of Various Magnitudes on Osteoarthritic Chondrocytes Exposed to IL-1beta. Indian J. Med. Res. 2010, 132, 209–217. [Google Scholar] [PubMed]
- Pascarelli, N.A.; Collodel, G.; Moretti, E.; Cheleschi, S.; Fioravanti, A. Changes in Ultrastructure and Cytoskeletal Aspects of Human Normal and Osteoarthritic Chondrocytes Exposed to Interleukin-1β and Cyclical Hydrostatic Pressure. Int. J. Mol. Sci. 2015, 16, 26019–26034. [Google Scholar] [CrossRef] [PubMed]
- Cheleschi, S.; De Palma, A.; Pecorelli, A.; Pascarelli, N.A.; Valacchi, G.; Belmonte, G.; Carta, S.; Galeazzi, M.; Fioravanti, A. Hydrostatic Pressure Regulates MicroRNA Expression Levels in Osteoarthritic Chondrocyte Cultures via the Wnt/β-Catenin Pathway. Int. J. Mol. Sci. 2017, 18, 133. [Google Scholar] [CrossRef] [PubMed]
- Montagne, K.; Onuma, Y.; Ito, Y.; Aiki, Y.; Furukawa, K.S.; Ushida, T. High Hydrostatic Pressure Induces Pro-osteoarthritic Changes in Cartilage Precursor Cells: A Transcriptome Analysis. PLoS ONE 2017, 12, e0183226. [Google Scholar] [CrossRef]
- De Palma, A.; Cheleschi, S.; Pascarelli, N.A.; Giannotti, S.; Galeazzi, M.; Fioravanti, A. Hydrostatic Pressure as Epigenetic Modulator in Chondrocyte Cultures: A Study on miRNA-155, miRNA-181a and miRNA-223 Expression Levels. J. Biomech. 2018, 66, 165–169. [Google Scholar] [CrossRef]
- Rieder, B.; Weihs, A.M.; Weidinger, A.; Szwarc, D.; Nürnberger, S.; Redl, H.; Rünzler, D.; Huber-Gries, C.; Teuschl, A.H. Hydrostatic Pressure-generated Reactive Oxygen Species Induce Osteoarthritic Conditions in Cartilage Pellet Cultures. Sci. Rep. 2018, 8, 17010. [Google Scholar] [CrossRef] [PubMed]
- Tworkoski, E.; Glucksberg, M.R.; Johnson, M. The Effect of the Rate of Hydrostatic Pressure Depressurization on Cells in Culture. PLoS ONE 2018, 13, e0189890. [Google Scholar] [CrossRef] [PubMed]
- Cheleschi, S.; Barbarino, M.; Gallo, I.; Tenti, S.; Bottaro, M.; Frati, E.; Giannotti, S.; Fioravanti, A. Hydrostatic Pressure Regulates Oxidative Stress through microRNA in Human Osteoarthritic Chondrocytes. Int. J. Mol. Sci. 2020, 21, 3653. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Chen, R.; Gui, C.; Tao, T.; Ge, Y.; Zhao, X.; Qin, R.; Yao, W.; Gu, S.; Jiang, Y.; et al. Eliminating Senescent Chondrogenic Progenitor Cells Enhances Chondrogenesis under Intermittent Hydrostatic Pressure for the Treatment of OA. Stem Cell Res. Ther. 2020, 11, 199. [Google Scholar] [CrossRef] [PubMed]
- Zeddou, M. Osteoarthritis Is a Low-Grade Inflammatory Disease: Obesity’s Involvement and Herbal Treatment. Evid. Based Complement. Alternat. Med. 2019, 2019, 2037484. [Google Scholar] [CrossRef]
- Silvestre, M.P.; Rodrigues, A.M.; Canhão, H.; Marques, C.; Teixeira, D.; Calhau, C.; Branco, J. Cross-Talk between Diet-Associated Dysbiosis and Hand Osteoarthritis. Nutrients 2020, 12, 3469. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Chen, Q. Adipokines: New Therapeutic Target for Osteoarthritis? Curr. Rheumatol. Rep. 2019, 21, 71. [Google Scholar] [CrossRef] [PubMed]
- Recinella, L.; Orlando, G.; Ferrante, C.; Chiavaroli, A.; Brunetti, L.; Leone, S. Adipokines: New Potential Therapeutic Target for Obesity and Metabolic, Rheumatic, and Cardiovascular Diseases. Front. Physiol. 2020, 11, 578966. [Google Scholar] [CrossRef] [PubMed]
- Neumann, E.; Hasseli, R.; Ohl, S.; Lange, U.; Frommer, K.W.; Müller-Ladner, U. Adipokines and Autoimmunity in Inflammatory Arthritis. Cells 2021, 10, 216. [Google Scholar] [CrossRef] [PubMed]
- Fioravanti, A.; Cheleschi, S.; De Palma, A.; Addimanda, O.; Mancarella, L.; Pignotti, E.; Pulsatelli, L.; Galeazzi, M.; Meliconi, R. Can Adipokines Serum Levels be Used as Biomarkers of Hand Osteoarthritis? Biomarkers 2018, 23, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Fioravanti, A.; Tenti, S.; Bacarelli, M.R.; Damiani, A.; Li Gobbi, F.; Bandinelli, F.; Cheleschi, S.; Galeazzi, M.; Benucci, M. Tocilizumab Modulates Serum Levels of Adiponectin and Chemerin in Patients with Rheumatoid Arthritis: Potential Cardiovascular Protective Role of IL-6 Inhibition. Clin. Exp. Rheumatol. 2019, 37, 293–300. [Google Scholar] [PubMed]
- Carrión, M.; Frommer, K.W.; Pérez-García, S.; Müller-Ladner, U.; Gomariz, R.P.; Neumann, E. The Adipokine Network in Rheumatic Joint Diseases. Int. J. Mol. Sci. 2019, 20, 4091. [Google Scholar] [CrossRef]
- MacDonald, I.J.; Liu, S.C.; Huang, C.C.; Kuo, S.J.; Tsai, C.H.; Tang, C.H. Associations between Adipokines in Arthritic Disease and Implications for Obesity. Int. J. Mol. Sci. 2019, 20, 1505. [Google Scholar] [CrossRef]
- Francisco, V.; Ruiz-Fernández, C.; Pino, J.; Mera, A.; González-Gay, M.A.; Gómez, R.; Lago, F.; Mobasheri, A.; Gualillo, O. Adipokines: Linking Metabolic Syndrome, the Immune System, and Arthritic Diseases. Biochem. Pharmacol. 2019, 165, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Han, D.F.; Li, Y.; Xu, H.Y.; Li, R.H.; Zhao, D. An Update on the Emerging Role of Visfatin in the Pathogenesis of Osteoarthritis and Pharmacological Intervention. Evid. Based Complement. Alternat. Med. 2020, 2020, 8303570. [Google Scholar] [CrossRef] [PubMed]
- Fioravanti, A.; Giannitti, C.; Cheleschi, S.; Simpatico, A.; Pascarelli, N.A.; Galeazzi, M. Circulating Levels of Adiponectin, Resistin, and Visfatin after Mud-bath Therapy in Patients with Bilateral Knee Osteoarthritis. Int. J. Biometeorol. 2015, 59, 1691–1700. [Google Scholar] [CrossRef] [PubMed]
- Cheleschi, S.; Giordano, N.; Volpi, N.; Tenti, S.; Gallo, I.; Di Meglio, M.; Giannotti, S.; Fioravanti, A. A Complex Relationship between Visfatin and Resistin and microRNA: An In Vitro Study on Human Chondrocyte Cultures. Int. J. Mol. Sci. 2018, 19, 3909. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.H.; Tsai, C.H.; Huang, Y.L.; Fong, Y.C.; Tang, C.H. Visfatin Promotes IL-6 and TNF-α Production in Human Synovial Fibroblasts by Repressing miR-199a-5p through ERK, p38 and JNK Signaling Pathways. Int. J. Mol. Sci. 2018, 19, 190. [Google Scholar] [CrossRef] [PubMed]
- Cheleschi, S.; Tenti, S.; Mondanelli, N.; Corallo, C.; Barbarino, M.; Giannotti, S.; Gallo, I.; Giordano, A.; Fioravanti, A. MicroRNA-34a and MicroRNA-181a Mediate Visfatin-Induced Apoptosis and Oxidative Stress via NF-κB Pathway in Human Osteoarthritic Chondrocytes. Cells 2019, 8, 874. [Google Scholar] [CrossRef] [PubMed]
- Cheleschi, S.; Gallo, I.; Barbarino, M.; Giannotti, S.; Mondanelli, N.; Giordano, A.; Tenti, S.; Fioravanti, A. MicroRNA Mediate Visfatin and Resistin Induction of Oxidative Stress in Human Osteoarthritic Synovial Fibroblasts Via NF-κB Pathway. Int. J. Mol. Sci. 2019, 20, 5200. [Google Scholar] [CrossRef] [PubMed]
- Adyshev, D.M.; Elangovan, V.R.; Moldobaeva, N.; Mapes, B.; Sun, X.; Garcia, J.G. Mechanical Stress Induces Pre-B-cell Colony-enhancing Factor/NAMPT Expression via Epigenetic Regulation by miR-374a and miR-568 in Human Lung Endothelium. Am. J. Respir. Cell Mol. Biol. 2014, 50, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.P.; Chen, C.N.; Chang, H.I.; Huang, K.C.; Cheng, C.C.; Chiu, F.Y.; Lee, K.C.; Lo, C.M.; Chang, S.F. Low Shear Stress Attenuates COX-2 Expression Induced by Resistin in Human Osteoarthritic Chondrocytes. J. Cell Physiol. 2017, 232, 1448–1457. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; He, Z.; Xu, L.; Lu, L.; Feng, H.; Leong, D.J.; Kim, S.J.; Hirsh, D.M.; Majeska, R.J.; Goldring, M.B.; et al. CITED2 Mediates the Mechanical Loading-induced Suppression of Adipokines in the Infrapatellar Fat Pad. Ann. N. Y. Acad. Sci. 2019, 1442, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Zhou, X.; Jin, M.; Nie, J.; Li, X. Molecular Mechanisms of Mechanical Load-induced Osteoarthritis. Int. Orthop. 2021. [Google Scholar] [CrossRef]
- Nerucci, F.; Fioravanti, A.; Cicero, M.R.; Marcolongo, R.; Spinelli, G. Preparation of a Pressurization System to Study the Effect of Hydrostatic Pressure on Chondrocyte Cultures. In Vitro Cell. Dev. Biol. Anim. 1998, 34, 9–10. [Google Scholar] [CrossRef] [PubMed]
- Buckwalter, J.A.; Anderson, D.D.; Brown, T.D.; Tochigi, Y.; Martin, J.A. The Roles of Mechanical Stresses in the Pathogenesis of Osteoarthritis: Implications for Treatment of Joint Injuries. Cartilage 2013, 4, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, R.C.; Mellor, L.F.; Krause, A.R.; Donahue, H.J.; Loboa, E.G. LRP Receptors in Chondrocytes are Modulated by Simulated Microgravity and Cyclic Hydrostatic Pressure. PLoS ONE 2019, 14, e0223245. [Google Scholar] [CrossRef] [PubMed]
- Laiguillon, M.C.; Houard, X.; Bougault, C.; Gosset, M.; Nourissat, G.; Sautet, A.; Jacques, C.; Berenbaum, F.; Sellam, J. Expression and Function of Visfatin (Nampt), an Adipokine-enzyme Involved in Inflammatory Pathways of Osteoarthritis. Arthritis Res. Ther. 2014, 16, R38. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xing, X.; Hensley, G.; Chang, L.W.; Liao, W.; Abu-Amer, Y.; Sandell, L.J. Resistin Induces Expression of Proinflammatory Cytokines and Chemokines in Human Articular Chondrocytes via Transcription and Messenger RNA Stabilization. Arthritis Rheum. 2010, 62, 1993–2003. [Google Scholar] [CrossRef] [PubMed]
- Correia, C.; Pereira, A.L.; Duarte, A.R.; Frias, A.M.; Pedro, A.J.; Oliveira, J.T.; Sousa, R.A.; Reis, R.L. Dynamic Culturing of Cartilage Tissue: The Significance of Hydrostatic Pressure. Tissue Eng. Part A 2012, 18, 1979–1991. [Google Scholar] [CrossRef] [PubMed]
- Pérez-García, S.; Gutiérrez-Cañas, I.; Seoane, I.V.; Fernández, J.; Mellado, M.; Leceta, J.; Tío, L.; Villanueva-Romero, R.; Juarranz, Y.; Gomariz, R.P. Healthy and Osteoarthritic Synovial Fibroblasts Produce a Disintegrin and Metalloproteinase with Thrombospondin Motifs 4, 5, 7, and 12: Induction by IL-1β and Fibronectin and Contribution to Cartilage Damage. Am. J. Pathol. 2016, 186, 2449–2461. [Google Scholar] [CrossRef] [PubMed]
- Nazempour, A.; Quisenberry, C.R.; Abu-Lail, N.I.; Van Wie, B.J. Combined Effects of Oscillating Hydrostatic Pressure, Perfusion and Encapsulation in a Novel Bioreactor for Enhancing Extracellular Matrix Synthesis by Bovine Chondrocytes. Cell Tissue Res. 2017, 370, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Machado, C.R.L.; Resende, G.G.; Macedo, R.B.V.; do Nascimento, V.C.; Branco, A.S.; Kakehasi, A.M.; Andrade, M.V. Fibroblast-like Synoviocytes from Fluid and Synovial Membrane from Primary Osteoarthritis Demonstrate Similar Production of Interleukin 6, and Metalloproteinases 1 and 3. Clin. Exp. Rheumatol. 2019, 37, 306–309. [Google Scholar] [PubMed]
- D‘Lima, D.D.; Hashimoto, S.; Chen, P.C.; Colwell, C.W., Jr.; Lotz, M.K. Human Chondrocyte Apoptosis in Response to Mechanical Injury. Osteoarthr. Cartil. 2001, 9, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Loening, A.M.; James, I.E.; Levenston, M.E.; Badger, A.M.; Frank, E.H.; Kurz, B.; Nuttall, M.E.; Hung, H.H.; Blake, S.M.; Grodzinsky, A.J.; et al. Injurious Mechanical Compression of Bovine Articular Cartilage Induces Chondrocyte Apoptosis. Arch. Biochem. Biophys. 2000, 381, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Zhao, J.; Jing, W.; Yan, S.; Wang, X.; Xiao, C.; Ma, B. Role of miR-146a in Human Chondrocyte Apoptosis in Response to Mechanical Pressure Injury in Vitro. Int. J. Mol. Med. 2014, 34, 451–463. [Google Scholar] [CrossRef]
- Gosset, M.; Berenbaum, F.; Salvat, C.; Sautet, A.; Pigenet, A.; Tahiri, K.; Jacques, C. Crucial Role of Visfatin/Pre-B cell Colony-enhancing Factor in Matrix Degradation and Prostaglandin E2 Synthesis in Chondrocytes: Possible Influence on Osteoarthritis. Arthritis Rheum. 2008, 58, 1399–1409. [Google Scholar] [CrossRef] [PubMed]
- Zahan, O.M.; Serban, O.; Gherman, C.; Fodor, D. The Evaluation of Oxidative Stress in Osteoarthritis. Med. Pharm. Rep. 2020, 93, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Young, I.C.; Chuang, S.T.; Gefen, A.; Kuo, W.T.; Yang, C.T.; Hsu, C.H.; Lin, F.H. A Novel Compressive Stress-based Osteoarthritis-like Chondrocyte System. Exp. Biol. Med. 2017, 242, 1062–1071. [Google Scholar] [CrossRef]
- De Palma, A.; Cheleschi, S.; Pascarelli, N.A.; Tenti, S.; Galeazzi, M.; Fioravanti, A. Do MicroRNAs Have a Key Epigenetic Role in Osteoarthritis and in Mechanotransduction? Clin. Exp. Rheumatol. 2017, 35, 518–526. [Google Scholar]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy. Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Liu, Y.L.; Chen, X.Y.; Li, Q.; Zhong, J.; Dai, B.Y.; Shao, X.F.; Wu, G.B. Role of MicroRNA, LncRNA, and Exosomes in the Progression of Osteoarthritis: A Review of Recent Literature. Orthop. Surg. 2020, 12, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Guan, Y.; Tian, S.; Wang, Y.; Sun, K.; Chen, Q. Mechanical and IL-1β Responsive miR-365 Contributes to Osteoarthritis Development by Targeting Histone Deacetylase 4. Int. J. Mol. Sci. 2016, 17, 436. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Liu, M.; Fan, X.; Yang, M.; Liu, H.; Zhou, Y. Intermittent Cyclic Mechanical Tension Altered the microRNA Expression Profile of Human Cartilage Endplate Chondrocytes. Mol. Med. Rep. 2018, 17, 5238–5246. [Google Scholar] [CrossRef] [PubMed]
- Stadnik, P.S.; Gilbert, S.J.; Tarn, J.; Charlton, S.; Skelton, A.J.; Barter, M.J.; Duance, V.C.; Young, D.A.; Blain, E.J. Regulation of microRNA-221, -222, -21 and -27 in Articular Cartilage Subjected to Abnormal Compressive Forces. J. Physiol. 2021, 599, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Lories, R.J.; Monteagudo, S. Review Article: Is Wnt Signaling an Attractive Target for the Treatment of Osteoarthritis? Rheumatol. Ther. 2020, 7, 259–270. [Google Scholar] [CrossRef]
- Zhou, X.; Cao, H.; Yuan, Y.; Wu, W. Biochemical Signals Mediate the Crosstalk between Cartilage and Bone in Osteoarthritis. Biomed. Res. Int. 2020, 2020, 5720360. [Google Scholar] [CrossRef]
- Niu, Q.; Li, F.; Zhang, L.; Xu, X.; Liu, Y.; Gao, J.; Feng, X. Role of the Wnt/β-catenin Signaling Pathway in the Response of Chondrocytes to Mechanical Loading. Int. J. Mol. Med. 2016, 37, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.G.; Zheng, Q.; Song, J.X.; Li, J.; Wang, H.; Liu, P.; Wang, J.; Wang, C.D.; Zhang, X.L. Intermittent Cyclic Mechanical Tension Promotes Endplate Cartilage Degeneration via Canonical Wnt Signaling Pathway and E-cadherin/β-catenin Complex Cross-talk. Osteoarthr. Cartil. 2016, 24, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Ohba, S.; Lanigan, T.M.; Roessler, B.J. Leptin Receptor JAK2/STAT3 Signaling Modulates Expression of Frizzled Receptors in Articular Chondrocytes. Osteoarthr. Cartil. 2010, 18, 1620–1629. [Google Scholar] [CrossRef] [PubMed]
- Philp, A.M.; Collier, R.L.; Grover, L.M.; Davis, E.T.; Jones, S.W. Resistin Promotes the Abnormal Type I Collagen Phenotype of Subchondral Bone in Obese Patients with End Stage Hip Osteoarthritis. Sci. Rep. 2017, 7, 4042. [Google Scholar] [CrossRef] [PubMed]
- Denysenko, T.; Annovazzi, L.; Cassoni, P.; Melcarne, A.; Mellai, M.; Schiffer, D. WNT/β-catenin Signaling Pathway and Downstream Modulators in Low- and High-grade Glioma. Cancer Genom. Proteom. 2016, 13, 31–45. [Google Scholar]
- Sun, Y.; Wang, F.; Sun, X.; Wang, X.; Zhang, L.; Li, Y. CX3CR1 Regulates Osteoarthrosis Chondrocyte Proliferation and Apoptosis via Wnt/β-catenin Signaling. Biomed. Pharmacother. 2017, 96, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Bougault, C.; Priam, S.; Houard, X.; Pigenet, A.; Sudre, L.; Lories, R.J.; Jacques, C.; Berenbaum, F. Protective Role of Frizzled-related Protein B on Matrix Metalloproteinase Induction in Mouse Chondrocytes. Arthritis Res. Ther. 2014, 16, R137. [Google Scholar] [CrossRef]
- Robinson, J.A.; Chatterjee-Kishore, M.; Yaworsky, P.J.; Cullen, D.M.; Zhao, W.; Li, C.; Kharode, Y.; Sauter, L.; Babij, P.; Brown, E.L.; et al. Wnt/beta-catenin Signaling is a Normal Physiological Response to Mechanical Loading in Bone. J. Biol. Chem. 2006, 281, 31720–31728. [Google Scholar] [CrossRef]
- Li, F.F.; Zhang, B.; Cui, J.H.; Chen, F.L.; Ding, Y.; Feng, X. Alterations in β-catenin/E-cadherin Complex Formation During the Mechanotransduction of Saos-2 Osteoblastic Cells. Mol. Med. Rep. 2018, 18, 1495–1503. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, C.; Zhang, Y.; Gao, J.; Teng, F.; Tian, W.; Yang, W.; Yan, Y.; Xue, F. Visfatin Stimulates Endometrial cancer cell proliferation via activation of PI3K/Akt and MAPK/ERK1/2 signalling pathways. Gynecol. Oncol. 2016, 143, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Altman, R.; Alarcón, G.; Appelrouth, D.; Bloch, D.; Borenstein, D.; Brandt, K.; Brown, C.; Cooke, T.D.; Daniel, W.; Feldman, D.; et al. The American College of Rheumatology Criteria for the Classification and Reporting of Osteoarthritis of the Hip. Arthritis Rheum. 1991, 34, 505–514. [Google Scholar] [CrossRef]
- Mankin, H.J.; Dorfman, H.; Lippiello, L.; Zarins, A. Biochemical and Metabolic Abnormalities in Articular Cartilage from Osteo-arthritic Human Hips. II. Correlation of Morphology with Biochemical and Metabolic Data. J. Bone Joint Surg. Am. 1971, 53, 523–537. [Google Scholar] [CrossRef] [PubMed]
- Francin, P.J.; Guillaume, C.; Humbert, A.C.; Pottie, P.; Netter, P.; Mainard, D.; Presle, N. Association between the Chondrocyte Phenotype and the Expression of Adipokines and Their Receptors: Evidence for a Role of Leptin but not Adiponectin in the Expression of Cartilage-specific Markers. J. Cell. Physiol. 2011, 226, 2790–2797. [Google Scholar] [CrossRef]
- Fioravanti, A.; Cantarini, L.; Chellini, F.; Manca, D.; Paccagnini, E.; Marcolongo, R.; Collodel, G. Effect of Hyaluronic Acid (MW 500-730 kDa) on Proteoglycan and Nitric Oxide Production in Human Osteoarthritic Chondrocyte Cultures Exposed to Hydrostatic Pressure. Osteoarthr. Cartil. 2005, 13, 688–696. [Google Scholar] [CrossRef] [PubMed]
- Cheleschi, S.; Calamia, V.; Fernandez-Moreno, M.; Biava, M.; Giordani, A.; Fioravanti, A.; Anzini, M.; Blanco, F. In Vitro Comprehensive Analysis of VA692 a New Chemical Entity for the Treatment of Osteoarthritis. Int. Immunopharmacol. 2018, 64, 86–100. [Google Scholar] [CrossRef] [PubMed]
- Ramakers, C.; Ruijter, J.M.; Deprez, R.H.; Moorman, A.F. Assumption-free Analysis of Quantitative Real-time Polymerase Chain Reaction (PCR) Data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new Mathematical Model for Relative Quantification in Real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheleschi, S.; Tenti, S.; Barbarino, M.; Giannotti, S.; Bellisai, F.; Frati, E.; Fioravanti, A. Exploring the Crosstalk between Hydrostatic Pressure and Adipokines: An In Vitro Study on Human Osteoarthritic Chondrocytes. Int. J. Mol. Sci. 2021, 22, 2745. https://doi.org/10.3390/ijms22052745
Cheleschi S, Tenti S, Barbarino M, Giannotti S, Bellisai F, Frati E, Fioravanti A. Exploring the Crosstalk between Hydrostatic Pressure and Adipokines: An In Vitro Study on Human Osteoarthritic Chondrocytes. International Journal of Molecular Sciences. 2021; 22(5):2745. https://doi.org/10.3390/ijms22052745
Chicago/Turabian StyleCheleschi, Sara, Sara Tenti, Marcella Barbarino, Stefano Giannotti, Francesca Bellisai, Elena Frati, and Antonella Fioravanti. 2021. "Exploring the Crosstalk between Hydrostatic Pressure and Adipokines: An In Vitro Study on Human Osteoarthritic Chondrocytes" International Journal of Molecular Sciences 22, no. 5: 2745. https://doi.org/10.3390/ijms22052745
APA StyleCheleschi, S., Tenti, S., Barbarino, M., Giannotti, S., Bellisai, F., Frati, E., & Fioravanti, A. (2021). Exploring the Crosstalk between Hydrostatic Pressure and Adipokines: An In Vitro Study on Human Osteoarthritic Chondrocytes. International Journal of Molecular Sciences, 22(5), 2745. https://doi.org/10.3390/ijms22052745