
Effects of Supplemental Lighting on Potassium Transport and Fruit Coloring of Tomatoes Grown in Hydroponics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
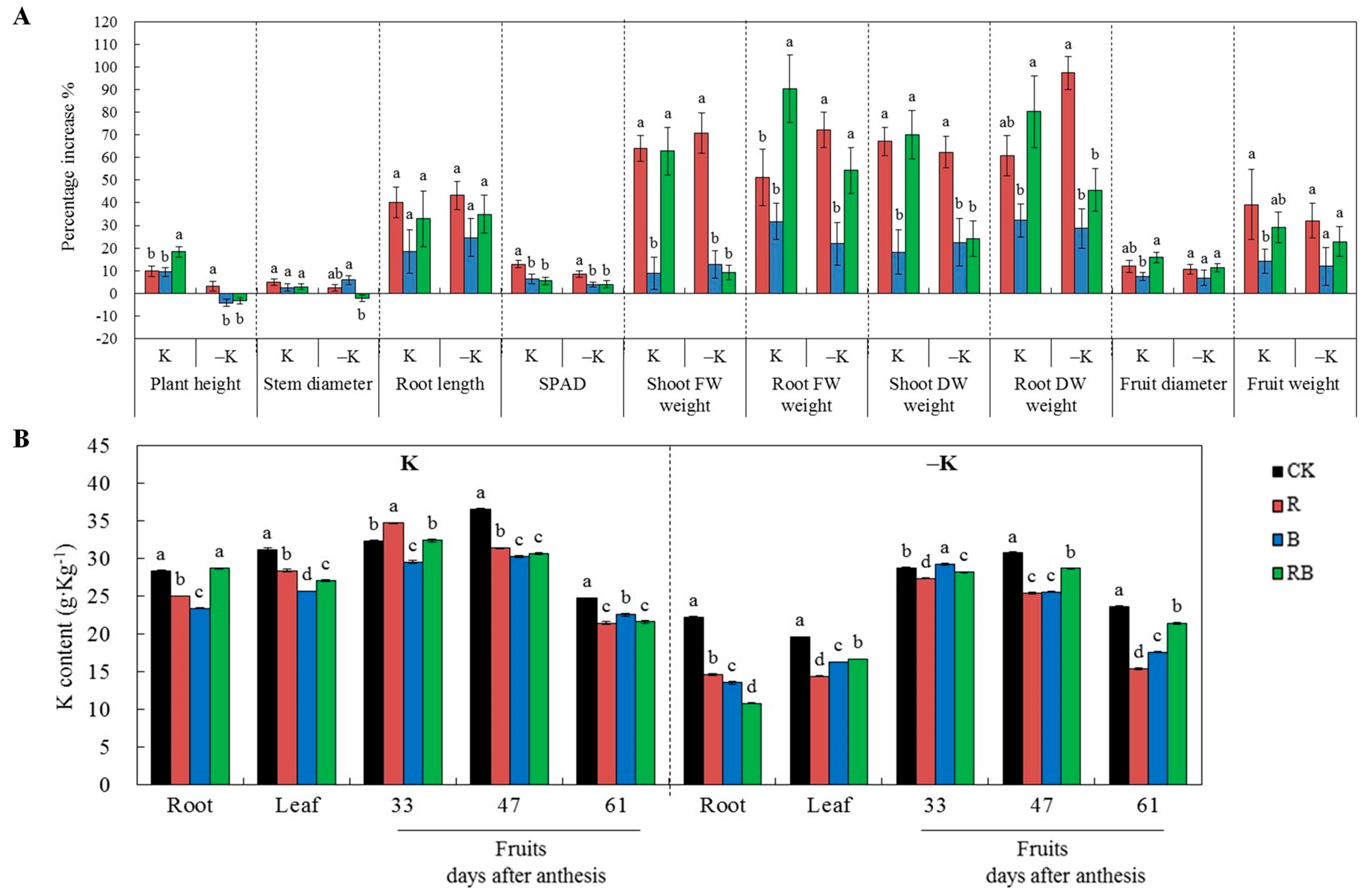
2.1. Characteristics of Plant Growth and K Content in Different Tissues
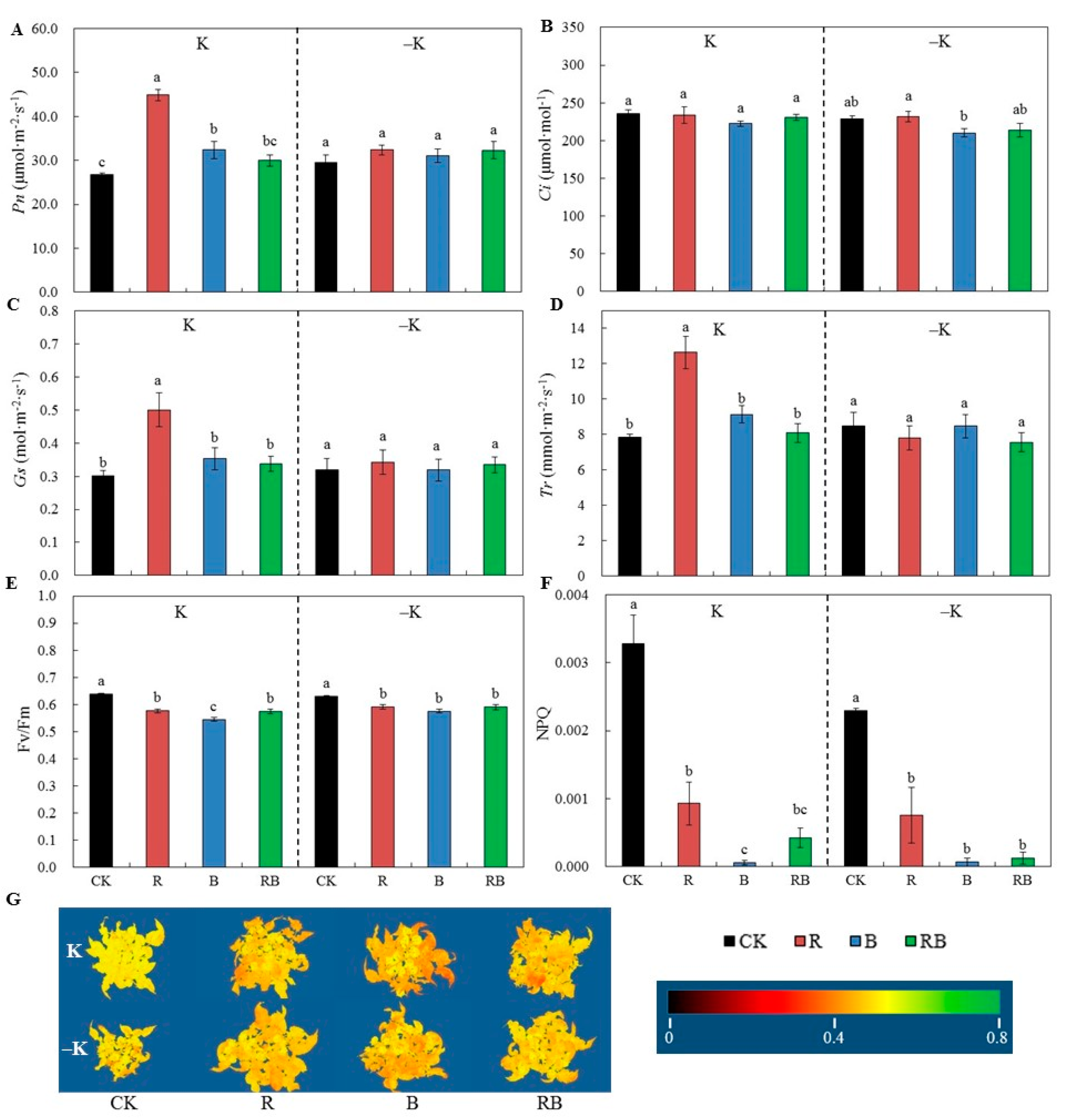
2.2. Characteristics of Photosynthesis, Maximum Quantum Efficiency of Photosystem Ⅱ, and Non-photochemical Quenching.
2.3. K Uptake Rate and Fruit Coloring
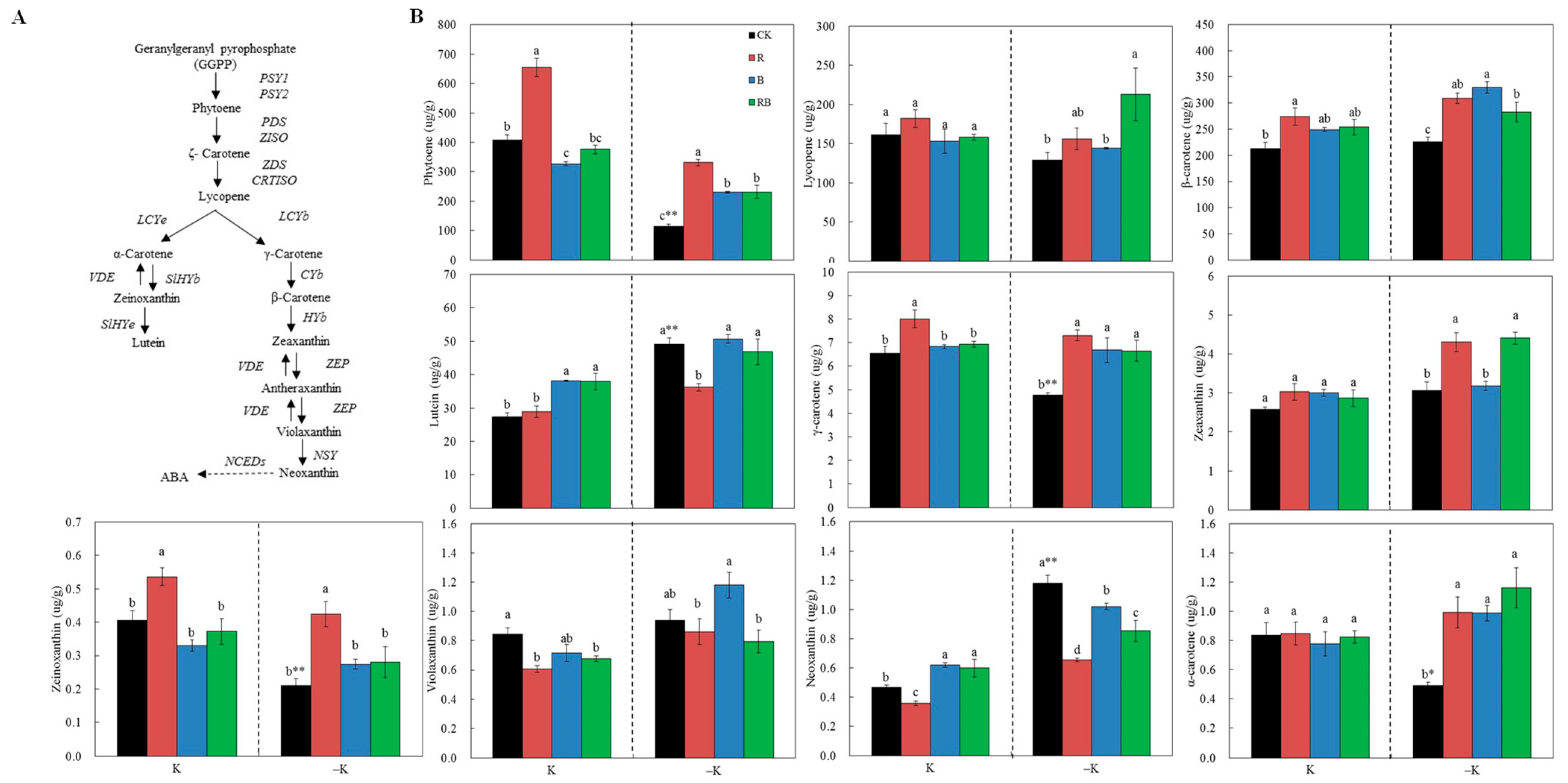
2.4. Fruit Carotenoid Content
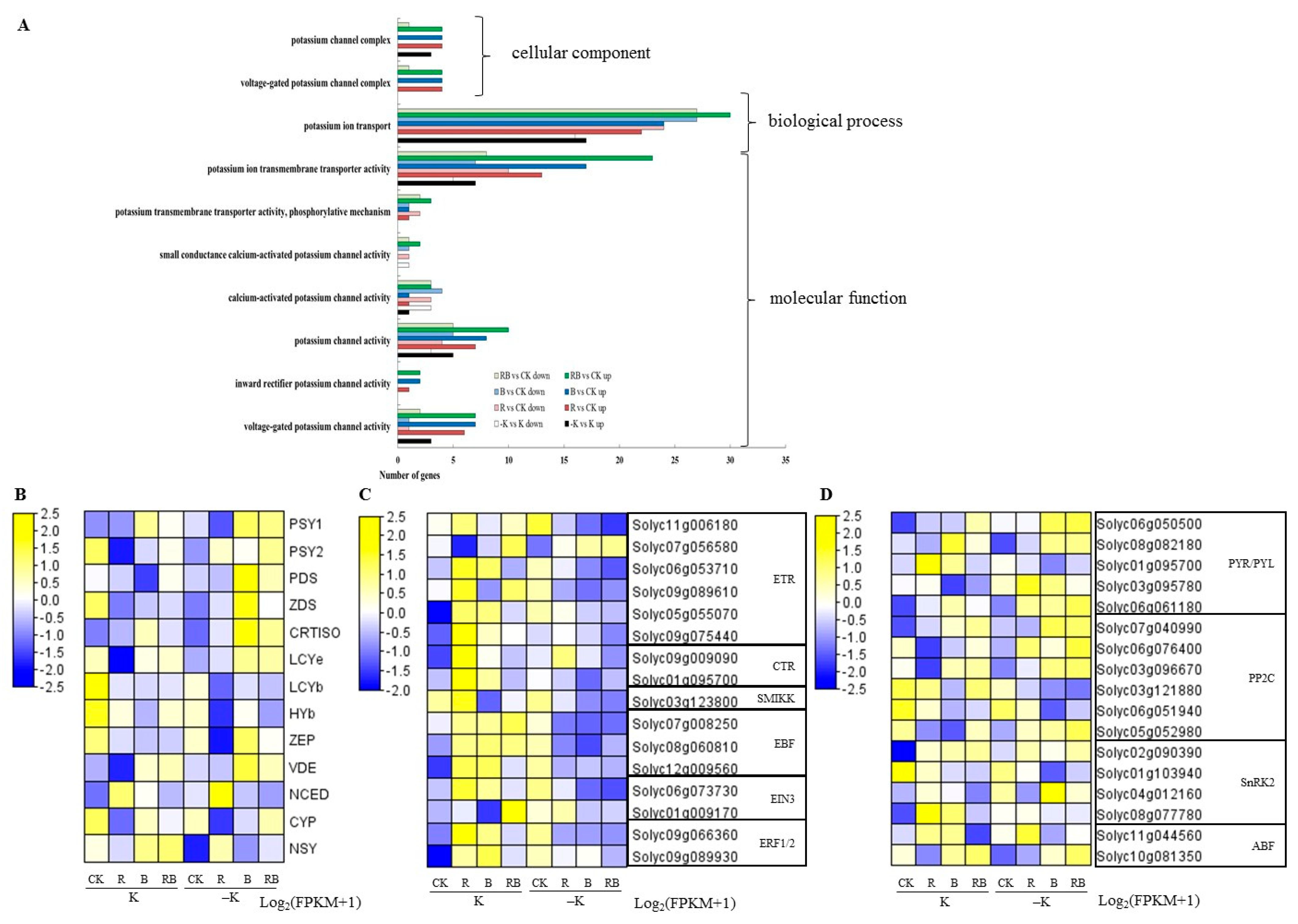
2.5. Expression Analysis of Potassium Transporter and Channel Genes
2.6. Expression Analysis of Genes Related to the Light Signaling Pathway
2.7. Transcriptome Analysis
3. Discussion
3.1. Supplemental Lighting Increased K Uptake and Accumulation by Increasing Photosynthesis, Plant Growth, and Fruit Weight
3.2. Supplemental Lighting Induced Carotenoid and Accelerated Fruit Coloring of Tomato Fruits
3.3. Potassium Transporter and Channel Genes in Tomato Fruits Were Upregulated by Blue Supplemental Lighting
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Potassium and Light Treatments
4.3. Measurement and Statistical Analyses
4.3.1. Growth Characteristics
4.3.2. K Content
4.3.3. Photosynthetic Characteristics
4.3.4. Maximum Quantum Efficiency of PSII and Non-photochemical Quenching
4.3.5. Root K Uptake Rate
4.3.6. Fruit Coloring Measurement
4.3.7. Detection of Carotenoids
4.3.8. Total Ribosenucleic Acid (RNA) Isolation, Complementary Deoxyribonucleic Acid (cDNA) Synthesis, and Real-time PCR Analysis
4.3.9. RNA-Seq Analysis
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Breksa, A.P.; Robertson, L.D.; Labate, J.A.; King, B.A.; King, D.E. Physicochemical and morphological analysis of ten tomato varieties identifies quality traits more readily manipulated through breeding and traditional selection methods. J. Food Compos. Anal. 2015, 42, 16–25. [Google Scholar] [CrossRef]
- Gomez, C.; Mitchell, C.A. Physiological and productivity responses of high-wire tomato as affected by supplemental light source and distribution within the canopy. J. Am. Soc. Hortic. Sci. 2016, 141, 196–208. [Google Scholar] [CrossRef]
- Tewolde, F.T.; Shiina, K.; Maruo, T.; Takagaki, M.; Kozai, T.; Yamori, W. Supplemental LED inter-lighting compensates for a shortage of light for plant growth and yield under the lack of sunshine. PLoS ONE 2018, 13, e0206592. [Google Scholar] [CrossRef] [PubMed]
- Paponov, M.; Kechasov, D.; Lacek, J.; Verheul, M.J.; Paponov, I.A. Supplemental Light-Emitting Diod inter-lighting increases tomato fruit growth through enhanced photosynthetic light use efficiency and modulated root activity. Front. Plant Sci. 2020, 10, 1656. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Lin, M.Y.; Mitchell, C.A. Light spectral and thermal properties govern biomass allocation in tomato through morphological and physiological changes. Environ. Exp. Bot. 2019, 157, 228–240. [Google Scholar] [CrossRef]
- Kim, H.J.; Yang, T.; Choi, S.H.; Wang, Y.J.; Lin, M.Y.; Liceaga, A.M. Supplemental intracanopy far-red radiation to red light improves fruit quality attributes of greenhouse tomatoes. Sci. Hortic. 2020, 261, 108985. [Google Scholar] [CrossRef]
- Xie, B.X.; Liu, H.C.; Song, S.S.; Sun, G.W.; Chen, R.Y. Effects of light quality on the quality formation of tomato fruits. Adv. Biol. Sci. Res. 2016, 3, 11–15. [Google Scholar]
- Xie, B.X.; Wei, J.J.; Zhang, Y.T.; Song, S.W.; Su, W.; Sun, G.W.; Hao, Y.W.; Liu, H.C. Supplemental blue and red light promote lycopene synthesis in tomato fruits. J. Integr. Agric. 2019, 18, 590–598. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Zhang, Y.T.; Song, S.W.; Su, W.; Hao, Y.W.; Liu, H.C. Supplementary red light results in the earlier ripening of tomato fruit depending on ethylene production. Environ. Exp. Bot. 2020, 175, 104044. [Google Scholar] [CrossRef]
- Passam, H.C.; Karapanos, I.C.; Bebeli, P.J.; Savvas, D. A review of recent research on tomato nutrition, breeding and post-harvest technology with reference to fruit quality. Eur. J. Plant Sci. Biotechnol. 2007, 1, 1–21. [Google Scholar]
- Hartz, T.K.; Miyao, G.; Mullen, R.J.; Cahn, M.D.; Valencia, J.; Brittan, K.L. Potassium requirements for maximum yield and fruit quality of processing tomato. J. Am. Soc. Hortic. Sci. 1999, 124, 199–204. [Google Scholar] [CrossRef]
- Chapagain, B.P.; Wiesman, Z. Effect of potassium magnesium chloride in the fertigation solution as partial source of potassium on growth, yield and quality of greenhouse tomato. Sci. Hortic. 2004, 99, 279–288. [Google Scholar] [CrossRef]
- Picha, D.H. Physiological factors associated with yellow shoulder expression in tomato fruit. J. Am. Soc. Hortic. Sci. 1987, 112, 798–801. [Google Scholar]
- Hou, G.X.; Wu, Z.H.; Liu, M.C.; Liang, H.; Ji, Y.H.; Zhao, M. Effects of zinc and potassium nutrition on physiology and quality of soilless cultivated greenhouse tomato. China Veg. 2018, 9, 34–37. (In English) [Google Scholar]
- Zhang, Y.T.; Suzuki, K.; Liu, H.C.; Nukaya, A.; Kiriiwa, Y. Fruit yellow-shoulder disorder as related to mineral element uptake of tomatoes grown in high temperature. Sci. Hortic. 2018, 242, 25–29. [Google Scholar] [CrossRef]
- Serio, F.; Leo, L.; Parente, A.; Santamaria, P. Potassium nutrition increases the lycopene content of tomato fruit. J. Hortic. Sci. Biotechnol. 2007, 82, 941–945. [Google Scholar] [CrossRef]
- Chen, X.B.; Yao, Q.F.; Gao, X.H.; Jiang, C.F.; Harberd, N.P.; Fu, X.D. Shoot-to-root mobile transcription factor HY5 coordinates plant carbon and nitrogen acquisition. Curr. Biol. 2016, 26, 640–646. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Kanno, S.; Mabuchi, A.; Monda, K.; Iba, K.; Yanagisawa, S.A. A phytochrome-B-mediated regulatory mechanism of phosphorus acquisition. Nat Plants 2018, 4, 1089–1101. [Google Scholar] [CrossRef]
- Lee, B.; Koprivova, A.; Kopriva, S. The key enzyme of sulfate assimilation, adenosine 5’-phosphosulfate reductase, is regulated by HY5 in Arabidopsis. Plant J. 2011, 67, 1042–1054. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Zhao, X.; Li, J.F.; Cai, H.Q.; Deng, W.X.; Li, L. MicroRNA408 is critical for the HY5-SPL7 gene network that mediates the coordinated response to light and copper. Plant Cell. 2014, 26, 4933–4953. [Google Scholar] [CrossRef]
- Davies, C.; Shin, R.; Liu, W.H.; Thomas, M.R.; Schachtman, D.P. Transporters expressed during grape berry (Vitis vinifera L.) development are associated with an increase in berry size and berry potassium accumulation. J. Exp. Bot. 2006, 57, 3209–3216. [Google Scholar] [CrossRef]
- Song, Z.Z.; Ma, R.J.; Yu, M.L. Genome-wide analysis and identification of KT/HAK/KUP potassium transporter gene family in peach (Prunus persica). Genet. Mol. Res. 2015, 14, 774–787. [Google Scholar] [CrossRef]
- Song, Z.Z.; Guo, S.L.; Zhang, C.H.; Zhang, B.B.; Ma, R.J.; Korir, N.K.; Yu, M.L. KT/HAK/KUP potassium transporter genes differentially expressed during fruit development, ripening, and postharvest shelf-life of ‘Xiahui6’ peaches. Acta Physiol. Plant 2015, 37, 131. [Google Scholar] [CrossRef]
- Song, M.Y.; Wang, S.F.; Chai, L.; Zhang, S.H.; Shen, Y.Y. Characterization of an ABA-induced and K+ channel gene FaKAT1 that regulates strawberry fruit ripening. J. Plant Growth Regul. 2016. [Google Scholar] [CrossRef]
- Hyun, T.K.; Rim, Y.; Kim, E.; Kim, J.S. Genome-wide and molecular evolution analyses of the KT/HAK/KUP family in tomato (Solanum lycopersicum L.). Genes Genom. 2014, 36, 365–374. [Google Scholar] [CrossRef]
- Faser, P.D.; Truesdale, M.R.; Bird, C.R.; Schuch, W.; Bramley, P.M. Carotenoid biosynthesis during tomato fruit development (Evidence for tissue-specific gene expression). Plant Physiol. 1994, 105, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Yanagi, T.; Okamoto, K.; Takita, S. Effect of blue, red, and blue/red lights of two different PPF levels on growth and morphogenesis of lettuce plants. Acta Hortic. 1996, 440, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Ménard, C.; Dorais, M.; Hovi, T.; Gosselin, A. Developmental and physiological responses of tomato and cucumber to additional blue light. Acta Hortic. 2006, 711, 291–296. [Google Scholar] [CrossRef]
- Chen, X.L.; Guo, W.Z.; Xue, X.Z.; Wang, L.C.; Qiao, X.J. Growth and quality responses of ‘Green Oak Leaf’ lettuce as affected by monochromic or mixed radiation provided by fluorescent lamp (FL) and light-emitting diode (LED). Sci. Hortic. 2014, 172, 168–175. [Google Scholar] [CrossRef]
- Trouwborst, G.; Oosterkamp, J.; Hogewoning, S.W.; Harbinson, J.; Ieperen, W.V. The responses of light interception, photosynthesis and fruit yield of cucumber to LED-lighting within the canopy. Physiol. Plant. 2010, 138, 289–300. [Google Scholar] [CrossRef]
- Shimada, H.; Iba, K.; Takamiya, K.I. Blue-light irradiation reduces the expression of puf and puc operons of Rhodobacter sphaeroides under semi-aerobic conditions. Plant Cell Physiol. 1992, 33, 471–475. [Google Scholar]
- Hoffmann, A.M.; Georg, N.; Mauricio, H. High blue light improves acclimation and photosynthetic recovery of pepper plants exposed to UV stress. Environ. Exp. Bot. 2015, 109, 254–263. [Google Scholar] [CrossRef]
- Walker, R.R.; Clingeleffer, P.R.; Keeridge, G.H.; Rűhl, E.H.; Nicholas, P.R.; Blackmore, D.H. Effects of the rootstock Ramsey (Vitis champini) on ion and organic acid composition of grapes and wine, and on wine spectral characteristics. Aust. J. Grape Wine Res. 1998, 4, 100–110. [Google Scholar] [CrossRef]
- Zhou, M.; Mao, X.; Chen, H.; Bai, M.; Liu, K.; Yang, G.S. Research advances on potassium nutrition and berry accumulation in grapevines. J. Fruit Sci. 2017, 34, 752–761. (In English) [Google Scholar]
- Nieves-Cordones, M.; Alemán, F.; Fon, M.; Martínez, V.; Rubio, F. K+ nutrition, uptake, and its role in environmental stress in plants. Environ. Adapt. Stress Toler. Plants Era Clim. Chang. 2012, 4, 5–112. [Google Scholar]
- Lu, N.; Marou, T.; Jahkan, M.; Hohjo, M.; Tsukagoshi, S.; Ito, Y.; Ichimura, T.; Shinohara, Y. Effects of supplemental lighting within the canopy at different developing stages on tomato yield and quality of single-truss tomato plants grown at high density. Environ. Control Biol. 2012, 50, 1–11. [Google Scholar] [CrossRef]
- Pinho, P.; Jokinen, K.; Halonen, L. The influence of the LED light spectrum on the growth and nutrient uptake of hydroponically grown lettuce. Light. Res. Technol. 2017, 49, 866–881. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Sams, C.E. Increases in shoot tissue pigments, glucosinolates, and mineral elements in sprouting broccoli after exposure to short-duration blue light from light emitting diodes. J. Am. Soc. Hortic. Sci. 2013, 138, 31–37. [Google Scholar] [CrossRef]
- Babourina, O.; Newman, I.; Shabala, S. Blue light-induced kinetics of H+ and Ca2+ fluxes in etiolated wild-type and phototropin-mutant Arabidopsis seedlings. Proc. Natl. Acad. Sci. USA 2002, 99, 2433–2438. [Google Scholar] [CrossRef]
- Alexander, L.; Grierson, D. Ethylene biosynthesis and action in tomato: A model for climacteric fruit ripening. J. Exp. Bot. 2002, 53, 2039–2055. [Google Scholar] [CrossRef]
- Bramely, P.M. Regulation of carotenoid formation during tomato fruit ripening and development. J. Exp. Bot. 2002, 53, 2107–2113. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, W.H. Potassium transport and signaling in higher plants. Annu. Rev. Plant Biol. 2013, 64, 451–476. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Y.F.; Wu, W.H. Potassium and phosphorus transport and signaling in plants. J. Integr. Plant Biol. 2020, 63, 34–52. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; Aleman, F.; Martinez, V.; Rubio, F. K+ uptake in plant roots. The systems involved, their regulation and parallels in other organisms. J. Plant Physiol. 2014, 171, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Golldack, D.; Zhao, C.S.; Bohnert, H.J. The expression of HAK-type K+ transporters is regulated in response to salinity stress in common ice plant. Plant Physiol. 2002, 129, 1482–1493. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.J.; Shin, R.; Schachtman, D.P. Expression of KT/KUP genes in Arabidopsis and the role of root hairs in K+ uptake. Plant Physiol. 2004, 134, 1135–1145. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, J.; Chen, Y.; Li, R.; Wang, H.; Wei, J. Genome-wide analysis and identification of HAK potassium transporter gene family in maize (Zea mays L.). Mol. Biol. Rep. 2012, 39, 8465–8473. [Google Scholar] [CrossRef] [PubMed]
- Gierth, M.; Mäser, P. Potassium transporters in plants-involvement in K+ acquisition, redistribution and homeostasis. FEBS Lett. 2007, 581, 2348–2356. [Google Scholar] [CrossRef] [PubMed]
- Okasakabe, Y.; Arinaga, N.; Umezawa, T.; Katsura, S.; Nagasihma, K.; Tanaka, H.; Ohiraki, H.; Seo, S.U.; Abo, M.; Yoshimura, E.; et al. Osmotic stress responses and plant growth controlled by potassium transporters in Arabidopsis. Plant Cell. 2013, 25, 609–624. [Google Scholar] [CrossRef] [PubMed]
- Blum, D.E.; Elaenga, T.M.; Linnemeyer, P.A.; Volkenburgh, E.V. Stimulation of growth and ion uptake in bean leaves by red and blue light. Plant Physiol. 1992, 100, 1968–1975. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.J.; Moran, N.; Lee, Y.S. Blue light activates potassium-efflux channels in flexor cells from samanea saman motor organs via two Mechanisms. Plant Physiol. 2000, 123, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Spalding, E.P.; Cosgrove, D.J. Large plasma-membrane depolarization precedes rapid blue-light-induced growth growth inhibition in cucumber. Planta 1989, 178, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Min, J.K.; Ruzicka, D.; Shin, R.; Schachtman, D.P. The Arabidopsis AP2/ERF transcription factor RAP2.11 modulates plant response to low-potassium conditions. Mol. Plant 2012, 5, 1042–1057. [Google Scholar]
- Song, J.L.; Huang, H.; Song, S.W.; Zhang, Y.T.; Su, W.; Liu, H.C. Effects of photoperiod interacted with nutrient solution concentration on nutritional quality, antioxidant and mineral content in lettuce. Agronomy 2020, 10, 920. [Google Scholar] [CrossRef]
- Fang, X.F.; Liu, S.; Gao, P.; Liu, H.Y.; Wang, X.Z.; Luan, F.S.; Zhang, Q.; Dai, Z.Y. Expression of ClPAP and ClPSY in watermelon correlates with chromoplast differentiation, carotenoid accumulation, and flesh color formation. Sci. Hortic. 2020, 270, 109437. [Google Scholar] [CrossRef]
- Livak, J.; Schmittgen, D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Liu, D.; Qin, M.; Xie, Z.; Chen, R.; Zhang, Y. Effects of Supplemental Lighting on Potassium Transport and Fruit Coloring of Tomatoes Grown in Hydroponics. Int. J. Mol. Sci. 2021, 22, 2687. https://doi.org/10.3390/ijms22052687
Wang W, Liu D, Qin M, Xie Z, Chen R, Zhang Y. Effects of Supplemental Lighting on Potassium Transport and Fruit Coloring of Tomatoes Grown in Hydroponics. International Journal of Molecular Sciences. 2021; 22(5):2687. https://doi.org/10.3390/ijms22052687
Chicago/Turabian StyleWang, Wei, Danxia Liu, Min Qin, Zhenbin Xie, Riyuan Chen, and Yiting Zhang. 2021. "Effects of Supplemental Lighting on Potassium Transport and Fruit Coloring of Tomatoes Grown in Hydroponics" International Journal of Molecular Sciences 22, no. 5: 2687. https://doi.org/10.3390/ijms22052687
APA StyleWang, W., Liu, D., Qin, M., Xie, Z., Chen, R., & Zhang, Y. (2021). Effects of Supplemental Lighting on Potassium Transport and Fruit Coloring of Tomatoes Grown in Hydroponics. International Journal of Molecular Sciences, 22(5), 2687. https://doi.org/10.3390/ijms22052687