
Revisiting Steroidogenic Pathways in the Human Placenta and Primary Human Trophoblast Cells

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Results
2.1. Clinical Characteristics of Mothers and Newborns Included in the Study
2.2. PCR Analysis of Placental Steroidogenic Enzyme Signature during Gestation
2.3. Expression Profile of Steroidogenic Enzymes in Primary Cells Isolated from Human Term Placenta and Choriocarcinoma-Derived Cell Lines
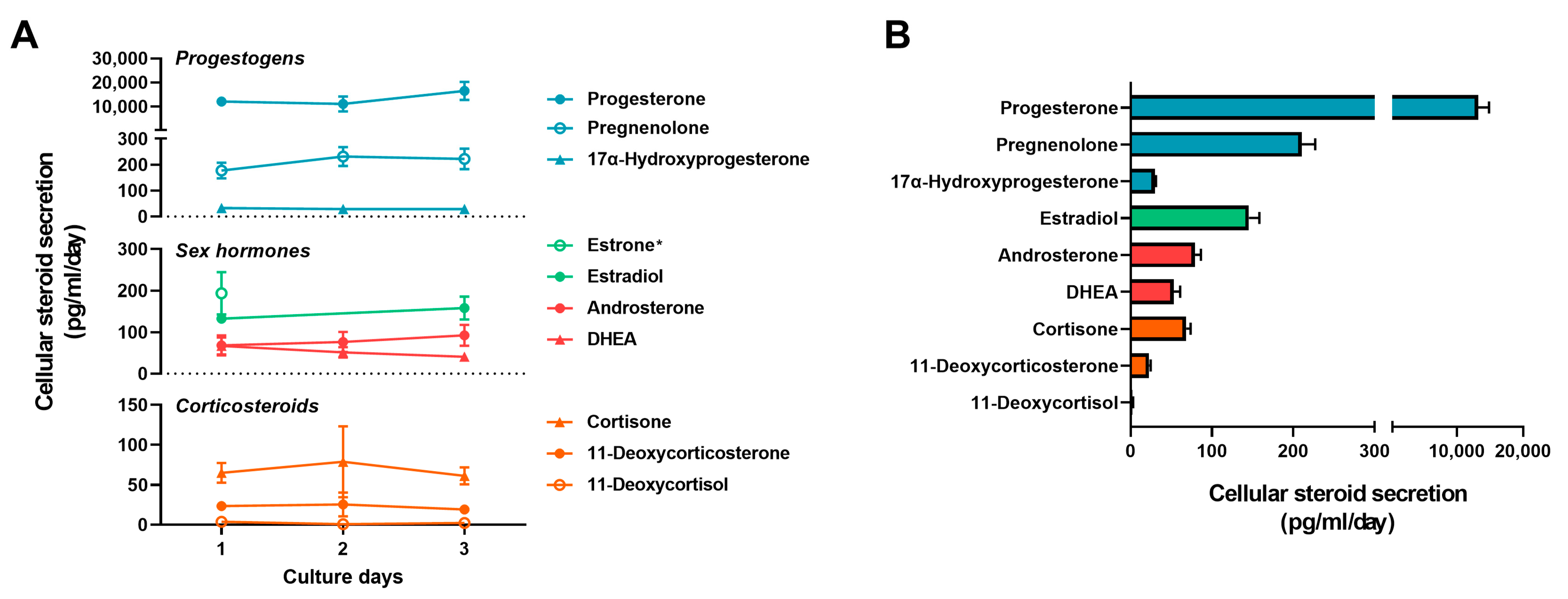
2.4. Secretion of Steroids from Primary Trophoblast Cells in Basal State
3. Discussion and Conclusions
4. Materials and Methods
4.1. Human Placenta Collection
4.2. Primary Cell Isolation and Analysis of Cell Purity
4.3. Cells
4.4. RNA Isolation, Reverse Transcription
4.5. PCR Analysis
4.5.1. Quantitative PCR Analysis
4.5.2. Droplet Digital PCR Analysis
4.6. Extraction of Steroids from Cell Culture Supernatants
4.7. Measurement of Steroids in Cell Supernatant
4.8. Statistical Analysis and Graphical Presentation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AKR | aldo-keto reductase |
| CTB | cytotrophoblast |
| CYP | cytochrome P450 |
| DHEA | dehydroepiandrosterone |
| HSD | hydroxysteroid dehydrogenase |
| SPE | solid phase extraction |
| SRD | steroid 5α-reductase |
| STB | syncytiotrophoblast |
References
- Knöfler, M.; Haider, S.; Saleh, L.; Pollheimer, J.; Gamage, T.K.; James, J. Human placenta and trophoblast development: Key molecular mechanisms and model systems. Cell. Mol. Life Sci. 2019, 76, 3479–3496. [Google Scholar] [CrossRef]
- Napso, T.; Yong, H.E.J.; Lopez-Tello, J.; Sferruzzi-Perri, A.N. The Role of Placental Hormones in Mediating Maternal Adaptations to Support Pregnancy and Lactation. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Staud, F.; Mazancova, K.; Miksik, I.; Pavek, P.; Fendrich, Z.; Pacha, J. Corticosterone transfer and metabolism in the dually perfused rat placenta: Effect of 11beta-hydroxysteroid dehydrogenase type 2. Placenta 2006, 27, 171–180. [Google Scholar] [CrossRef]
- Stirrat, L.I.; Sengers, B.G.; Norman, J.E.; Homer, N.Z.M.; Andrew, R.; Lewis, R.M.; Reynolds, R.M. Transfer and Metabolism of Cortisol by the Isolated Perfused Human Placenta. J. Clin. Endocrinol. Metab. 2018, 103, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Benediktsson, R.; Calder, A.A.; Edwards, C.R.; Seckl, J.R. Placental 11 beta-hydroxysteroid dehydrogenase: A key regulator of fetal glucocorticoid exposure. Clin. Endocrinol. 1997, 46, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Chatuphonprasert, W.; Jarukamjorn, K.; Ellinger, I. Physiology and Pathophysiology of Steroid Biosynthesis, Transport and Metabolism in the Human Placenta. Front Pharm. 2018, 9, 1027. [Google Scholar] [CrossRef] [PubMed]
- Noyola-Martínez, N.; Halhali, A.; Barrera, D. Steroid hormones and pregnancy. Gynecol. Endocrinol. 2019, 35, 376–384. [Google Scholar] [CrossRef]
- Raghupathy, R.; Al Mutawa, E.; Makhseed, M.; Azizieh, F.; Szekeres-Bartho, J. Modulation of cytokine production by dydrogesterone in lymphocytes from women with recurrent miscarriage. BJOG 2005, 112, 1096–1101. [Google Scholar] [CrossRef]
- Brar, A.K.; Frank, G.R.; Kessler, C.A.; Cedars, M.I.; Handwerger, S. Progesterone-dependent decidualization of the human endometrium is mediated by cAMP. Endocrine 1997, 6, 301–307. [Google Scholar] [CrossRef]
- Chen, J.Z.; Wong, M.H.; Brennecke, S.P.; Keogh, R.J. The effects of human chorionic gonadotrophin, progesterone and oestradiol on trophoblast function. Mol. Cell Endocrinol. 2011, 342, 73–80. [Google Scholar] [CrossRef]
- O’Shaughnessy, P.J.; Antignac, J.P.; Le Bizec, B.; Morvan, M.L.; Svechnikov, K.; Söder, O.; Savchuk, I.; Monteiro, A.; Soffientini, U.; Johnston, Z.C.; et al. Alternative (backdoor) androgen production and masculinization in the human fetus. PLoS Biol. 2019, 17, e3000002. [Google Scholar] [CrossRef]
- Albrecht, E.D.; Pepe, G.J. Estrogen regulation of placental angiogenesis and fetal ovarian development during primate pregnancy. Int. J. Dev. Biol. 2010, 54, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Michael, A.E.; Papageorghiou, A.T. Potential significance of physiological and pharmacological glucocorticoids in early pregnancy. Hum. Reprod. Update 2008, 14, 497–517. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.; Gluckman, P.D.; Godfrey, K.M.; Harding, J.E.; Owens, J.A.; Robinson, J.S. Fetal nutrition and cardiovascular disease in adult life. Lancet 1993, 341, 938–941. [Google Scholar] [CrossRef]
- Edwards, C.R.; Benediktsson, R.; Lindsay, R.S.; Seckl, J.R. Dysfunction of placental glucocorticoid barrier: Link between fetal environment and adult hypertension? Lancet 1993, 341, 355–357. [Google Scholar] [CrossRef]
- Payne, A.H.; Hales, D.B. Overview of steroidogenic enzymes in the pathway from cholesterol to active steroid hormones. Endocr. Rev. 2004, 25, 947–970. [Google Scholar] [CrossRef] [PubMed]
- Benirschke, K.; Kaufmann, P.; Baergen, R. Pathology of the Human Placenta; Springer: New York, NY, USA, 2016. [Google Scholar] [CrossRef]
- Kolahi, K.S.; Valent, A.M.; Thornburg, K.L. Cytotrophoblast, Not Syncytiotrophoblast, Dominates Glycolysis and Oxidative Phosphorylation in Human Term Placenta. Sci. Rep. 2017, 7, 42941. [Google Scholar] [CrossRef]
- Bilban, M.; Tauber, S.; Haslinger, P.; Pollheimer, J.; Saleh, L.; Pehamberger, H.; Wagner, O.; Knöfler, M. Trophoblast invasion: Assessment of cellular models using gene expression signatures. Placenta 2010, 31, 989–996. [Google Scholar] [CrossRef]
- Kallol, S.; Moser-Haessig, R.; Ontsouka, C.E.; Albrecht, C. Comparative expression patterns of selected membrane transporters in differentiated BeWo and human primary trophoblast cells. Placenta 2018, 72, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Karahoda, R.; Abad, C.; Horackova, H.; Kastner, P.; Zaugg, J.; Cerveny, L.; Kucera, R.; Albrecht, C.; Staud, F. Dynamics of Tryptophan Metabolic Pathways in Human Placenta and Placental-Derived Cells: Effect of Gestation Age and Trophoblast Differentiation. Front. Cell Dev. Biol. 2020, 8. [Google Scholar] [CrossRef]
- Staud, F.; Karahoda, R. Trophoblast: The central unit of fetal growth, protection and programming. Int. J. Biochem. Cell Biol. 2018, 105, 35–40. [Google Scholar] [CrossRef]
- Costa, M.A. The endocrine function of human placenta: An overview. Reprod. Biomed. Online 2016, 32, 14–43. [Google Scholar] [CrossRef] [PubMed]
- Voutilainen, R.; Miller, W.L. Developmental expression of genes for the stereoidogenic enzymes P450scc (20,22-desmolase), P450c17 (17 alpha-hydroxylase/17,20-lyase), and P450c21 (21-hydroxylase) in the human fetus. J. Clin. Endocrinol. Metab. 1986, 63, 1145–1150. [Google Scholar] [CrossRef]
- Siiteri, P.K.; MacDonald, P.C. Placental estrogen biosynthesis during human pregnancy. J. Clin. Endocrinol. Metab. 1966, 26, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Little, B.; Shaw, A. The conversion of progesterone to 17 alpha-hydroxyprogesterone by human placenta in vitro. Acta Endocrinol. 1961, 36, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Escobar, J.C.; Patel, S.S.; Beshay, V.E.; Suzuki, T.; Carr, B.R. The human placenta expresses CYP17 and generates androgens de novo. J. Clin. Endocrinol. Metab. 2011, 96, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Durkee, T.J.; McLean, M.P.; Hales, D.B.; Payne, A.H.; Waterman, M.R.; Khan, I.; Gibori, G. P450(17 alpha) and P450SCC gene expression and regulation in the rat placenta. Endocrinology 1992, 130, 1309–1317. [Google Scholar] [CrossRef]
- Tulchinsky, D.; Simmer, H.H. Sources of plasma 17alpha-hydroxyprogesterone in human pregnancy. J. Clin. Endocrinol. Metab. 1972, 35, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Baronio, F.; Ortolano, R.; Menabò, S.; Cassio, A.; Baldazzi, L.; Di Natale, V.; Tonti, G.; Vestrucci, B.; Balsamo, A. 46,XX DSD due to Androgen Excess in Monogenic Disorders of Steroidogenesis: Genetic, Biochemical, and Clinical Features. Int. J. Mol. Sci. 2019, 20, 4605. [Google Scholar] [CrossRef]
- Mathur, R.S.; Landgrebe, S.; Moody, L.O.; Powell, S.; Williamson, H.O. Plasma steroid concentrations in maternal and umbilical circulation after spontaneous onset of labor. J. Clin. Endocrinol. Metab. 1980, 51, 1235–1238. [Google Scholar] [CrossRef] [PubMed]
- Rhéaume, E.; Lachance, Y.; Zhao, H.F.; Breton, N.; Dumont, M.; de Launoit, Y.; Trudel, C.; Luu-The, V.; Simard, J.; Labrie, F. Structure and expression of a new complementary DNA encoding the almost exclusive 3 beta-hydroxysteroid dehydrogenase/delta 5-delta 4-isomerase in human adrenals and gonads. Mol. Endocrinol. 1991, 5, 1147–1157. [Google Scholar] [CrossRef] [PubMed]
- Flück, C.E.; Meyer-Böni, M.; Pandey, A.V.; Kempná, P.; Miller, W.L.; Schoenle, E.J.; Biason-Lauber, A. Why boys will be boys: Two pathways of fetal testicular androgen biosynthesis are needed for male sexual differentiation. Am. J. Hum. Genet. 2011, 89, 201–218. [Google Scholar] [CrossRef]
- Yoshida, N.; Osawa, Y. Purification of human placental aromatase cytochrome P-450 with monoclonal antibody and its characterization. Biochemistry 1991, 30, 3003–3010. [Google Scholar] [CrossRef] [PubMed]
- Engler, J.B.; Kursawe, N.; Solano, M.E.; Patas, K.; Wehrmann, S.; Heckmann, N.; Lühder, F.; Reichardt, H.M.; Arck, P.C.; Gold, S.M.; et al. Glucocorticoid receptor in T cells mediates protection from autoimmunity in pregnancy. Proc. Natl. Acad. Sci. USA 2017, 114, E181–E190. [Google Scholar] [CrossRef] [PubMed]
- Solano, M.E.; Arck, P.C. Steroids, Pregnancy and Fetal Development. Front. Immunol. 2020, 10, 3017. [Google Scholar] [CrossRef]
- Murphy, V.E.; Clifton, V.L. Alterations in human placental 11beta-hydroxysteroid dehydrogenase type 1 and 2 with gestational age and labour. Placenta 2003, 24, 739–744. [Google Scholar] [CrossRef]
- Pasqualini, J.R. Enzymes involved in the formation and transformation of steroid hormones in the fetal and placental compartments. J. Steroid. Biochem. Mol. Biol. 2005, 97, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Yang, K.; Challis, J.R. Differential expression of 11 beta-hydroxysteroid dehydrogenase types 1 and 2 in human placenta and fetal membranes. J. Clin. Endocrinol. Metab. 1997, 82, 300–305. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Michael, A.E.; Thurston, L.M.; Rae, M.T. Glucocorticoid metabolism and reproduction: A tale of two enzymes. Reproduction 2003, 126, 425–441. [Google Scholar] [CrossRef] [PubMed]
- Yinon, Y.; Kingdom, J.C.; Proctor, L.K.; Kelly, E.N.; Salle, J.L.; Wherrett, D.; Keating, S.; Nevo, O.; Chitayat, D. Hypospadias in males with intrauterine growth restriction due to placental insufficiency: The placental role in the embryogenesis of male external genitalia. Am. J. Med. Genet. A 2010, 152a, 75–83. [Google Scholar] [CrossRef]
- Hakim, C.; Padmanabhan, V.; Vyas, A.K. Gestational Hyperandrogenism in Developmental Programming. Endocrinology 2017, 158, 199–212. [Google Scholar] [CrossRef]
- Shimodaira, M.; Nakayama, T.; Sato, I.; Sato, N.; Izawa, N.; Mizutani, Y.; Furuya, K.; Yamamoto, T. Glucocorticoid synthesis-related genes: HSD11B1 and HSD11B2 in hypertensive disorders in pregnancy. Gynecol. Endocrinol. 2013, 29, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Shimodaira, M.; Nakayama, T.; Sato, I.; Sato, N.; Izawa, N.; Mizutani, Y.; Furuya, K.; Yamamoto, T. Estrogen synthesis genes CYP19A1, HSD3B1, and HSD3B2 in hypertensive disorders of pregnancy. Endocrine 2012, 42, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Kliman, H.J.; Nestler, J.E.; Sermasi, E.; Sanger, J.M.; Strauss, J.F., 3rd. Purification, characterization, and in vitro differentiation of cytotrophoblasts from human term placentae. Endocrinology 1986, 118, 1567–1582. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Estrada, J.; Menu, E.; Roques, P.; Barre-Sinoussi, F.; Chaouat, G. Evaluation of Cytokeratin 7 as an accurate intracellular marker with which to assess the purity of human placental villous trophoblast cells by flow cytometry. J. Immunol. Methods 2004, 286, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Schust, D.J. Isolation, purification and in vitro differentiation of cytotrophoblast cells from human term placenta. Reprod. Biol. Endocrinol. 2015, 13, 71. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.0031. [Google Scholar] [CrossRef]
- Karahoda, R.; Horackova, H.; Kastner, P.; Matthios, A.; Cerveny, L.; Kucera, R.; Kacerovsky, M.; Tebbens, J.D.; Bonnin, A.; Abad, C.; et al. Serotonin homeostasis in the materno-fetal interface at term: Role of transporters (SERT/SLC6A4 and OCT3/SLC22A3) and monoamine oxidase A (MAO-A) in uptake and degradation of serotonin by human and rat term placenta. Acta Physiol. 2020. [Google Scholar] [CrossRef]
- Peitzsch, M.; Dekkers, T.; Haase, M.; Sweep, F.C.; Quack, I.; Antoch, G.; Siegert, G.; Lenders, J.W.; Deinum, J.; Willenberg, H.S.; et al. An LC-MS/MS method for steroid profiling during adrenal venous sampling for investigation of primary aldosteronism. J. Steroid. Biochem. Mol. Biol. 2015, 145, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | First Trimester | Term |
|---|---|---|
| Number of individuals | 13 | 25 |
| Maternal age (years) | 27.91 ± 8.04 | 33.16 ± 4.92 |
| Gestational age (weeks) | 9.62 ± 1.19 | 39.44 ± 0.91 *** |
| Smoking (%) | 38 | 0 |
| Pre-pregnancy BMI (kg m−2) | 24.48 ± 3.31 | 24.18 ± 4.27 |
| Delivery BMI (kg m−2) | – | 28.72 ± 4.57 |
| Labor (% caesarian section) | – | 52 |
| Birth weight (kg) | – | 3.32 ± 0.36 |
| Birth height (cm) | – | 50.10 ± 1.62 |
| Fetal sex (% male) | – | 48 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karahoda, R.; Kallol, S.; Groessl, M.; Ontsouka, E.; Anderle, P.; Fluck, C.; Staud, F.; Albrecht, C. Revisiting Steroidogenic Pathways in the Human Placenta and Primary Human Trophoblast Cells. Int. J. Mol. Sci. 2021, 22, 1704. https://doi.org/10.3390/ijms22041704
Karahoda R, Kallol S, Groessl M, Ontsouka E, Anderle P, Fluck C, Staud F, Albrecht C. Revisiting Steroidogenic Pathways in the Human Placenta and Primary Human Trophoblast Cells. International Journal of Molecular Sciences. 2021; 22(4):1704. https://doi.org/10.3390/ijms22041704
Chicago/Turabian StyleKarahoda, Rona, Sampada Kallol, Michael Groessl, Edgar Ontsouka, Pascale Anderle, Christa Fluck, Frantisek Staud, and Christiane Albrecht. 2021. "Revisiting Steroidogenic Pathways in the Human Placenta and Primary Human Trophoblast Cells" International Journal of Molecular Sciences 22, no. 4: 1704. https://doi.org/10.3390/ijms22041704
APA StyleKarahoda, R., Kallol, S., Groessl, M., Ontsouka, E., Anderle, P., Fluck, C., Staud, F., & Albrecht, C. (2021). Revisiting Steroidogenic Pathways in the Human Placenta and Primary Human Trophoblast Cells. International Journal of Molecular Sciences, 22(4), 1704. https://doi.org/10.3390/ijms22041704