
Particle Size and Biological Fate of ZnO Do Not Cause Acute Toxicity, but Affect Toxicokinetics and Gene Expression Profiles in the Rat Livers after Oral Administration



Abstract

1. Introduction
2. Results
2.1. Characterization of B-ZnO and N-ZnO
2.2. In Vivo Dissolution Properties of B-ZnO and N-ZnO
2.3. Toxicokinetics
2.4. Oral Toxicity and Tissue Distribution
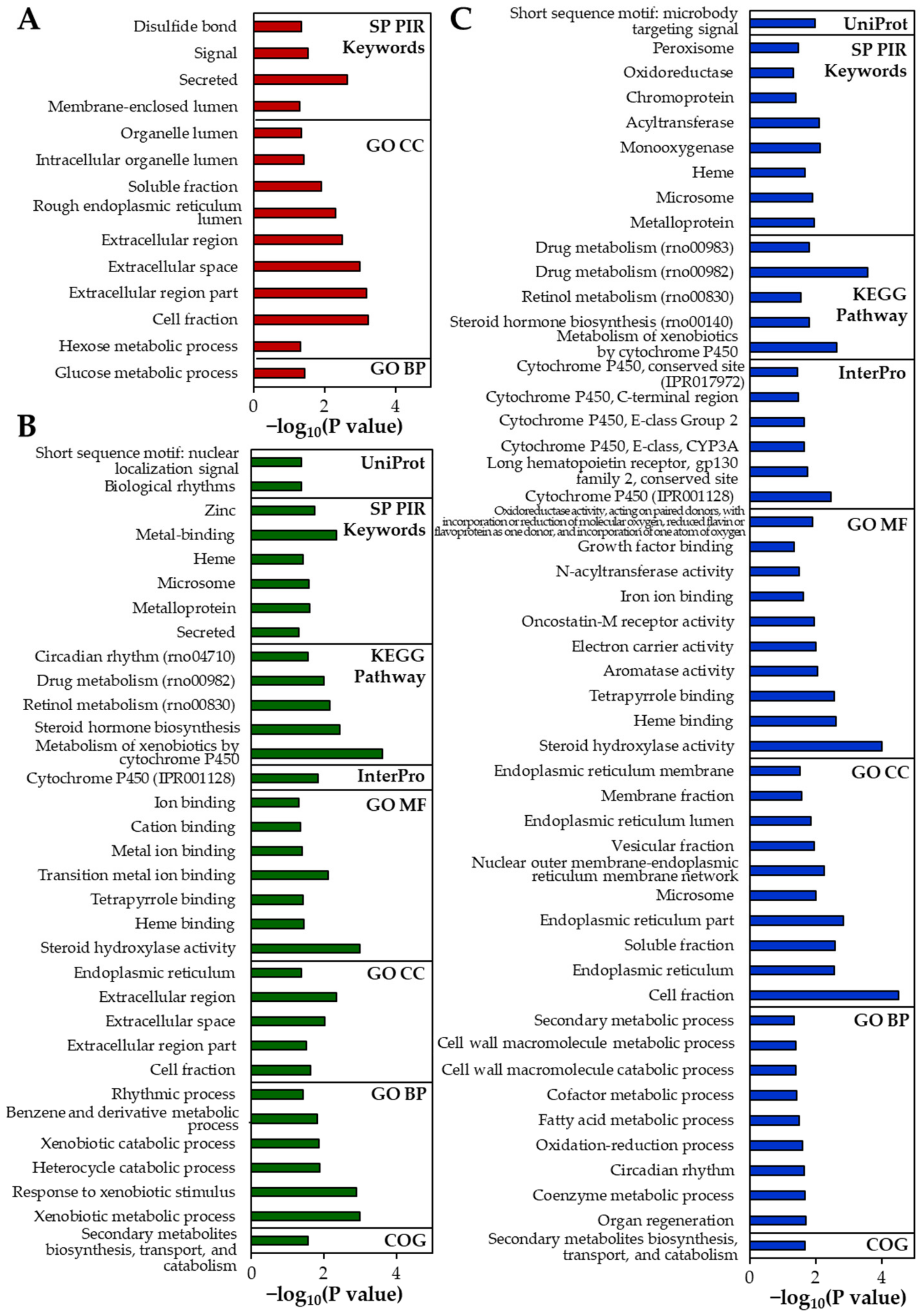
2.5. Gene Expression Profiles in the Rat Livers Following 14-Day Repeated Oral Administration
2.6. Identification of DEGs and Their Effect on Biological Functions
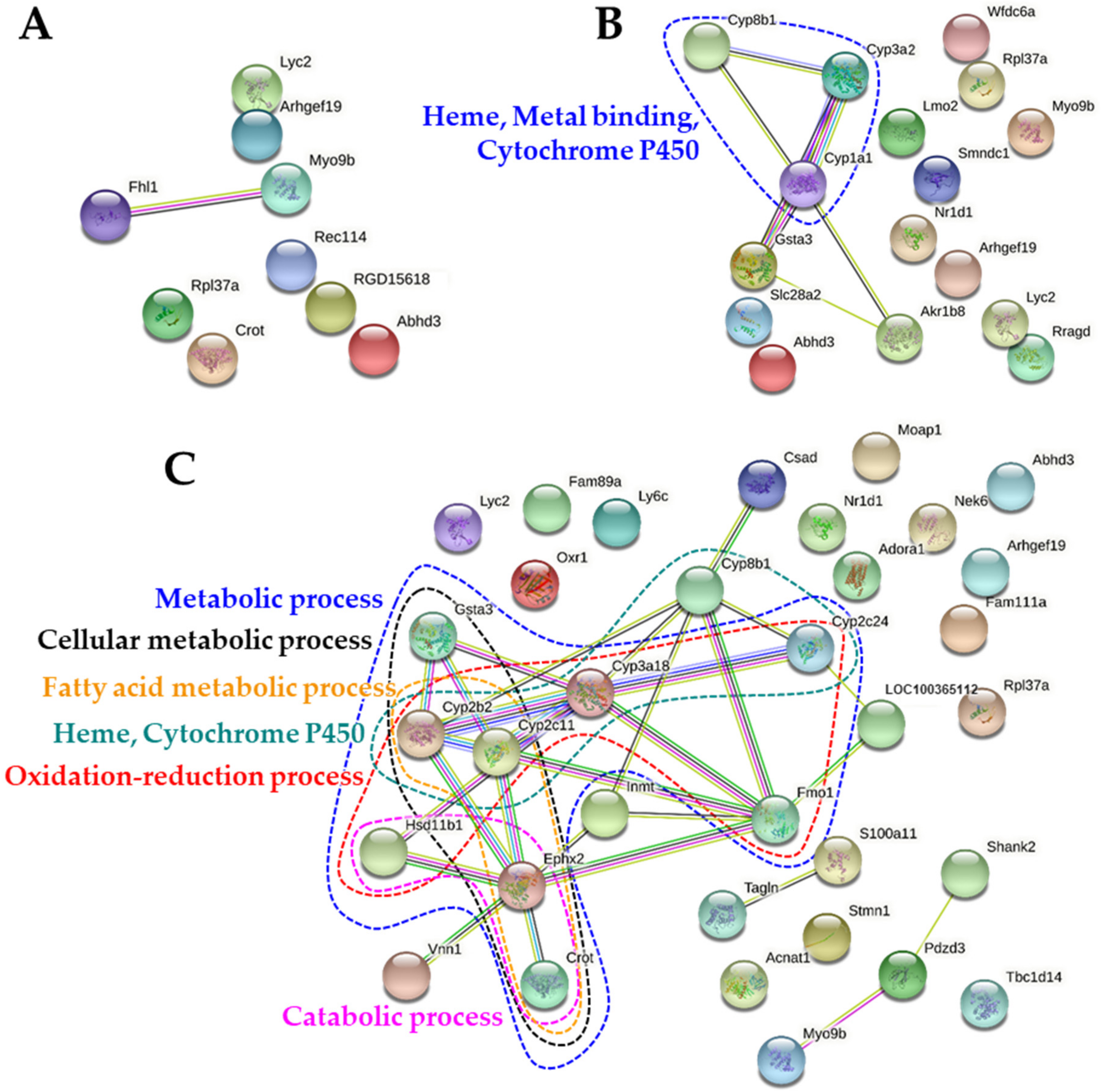
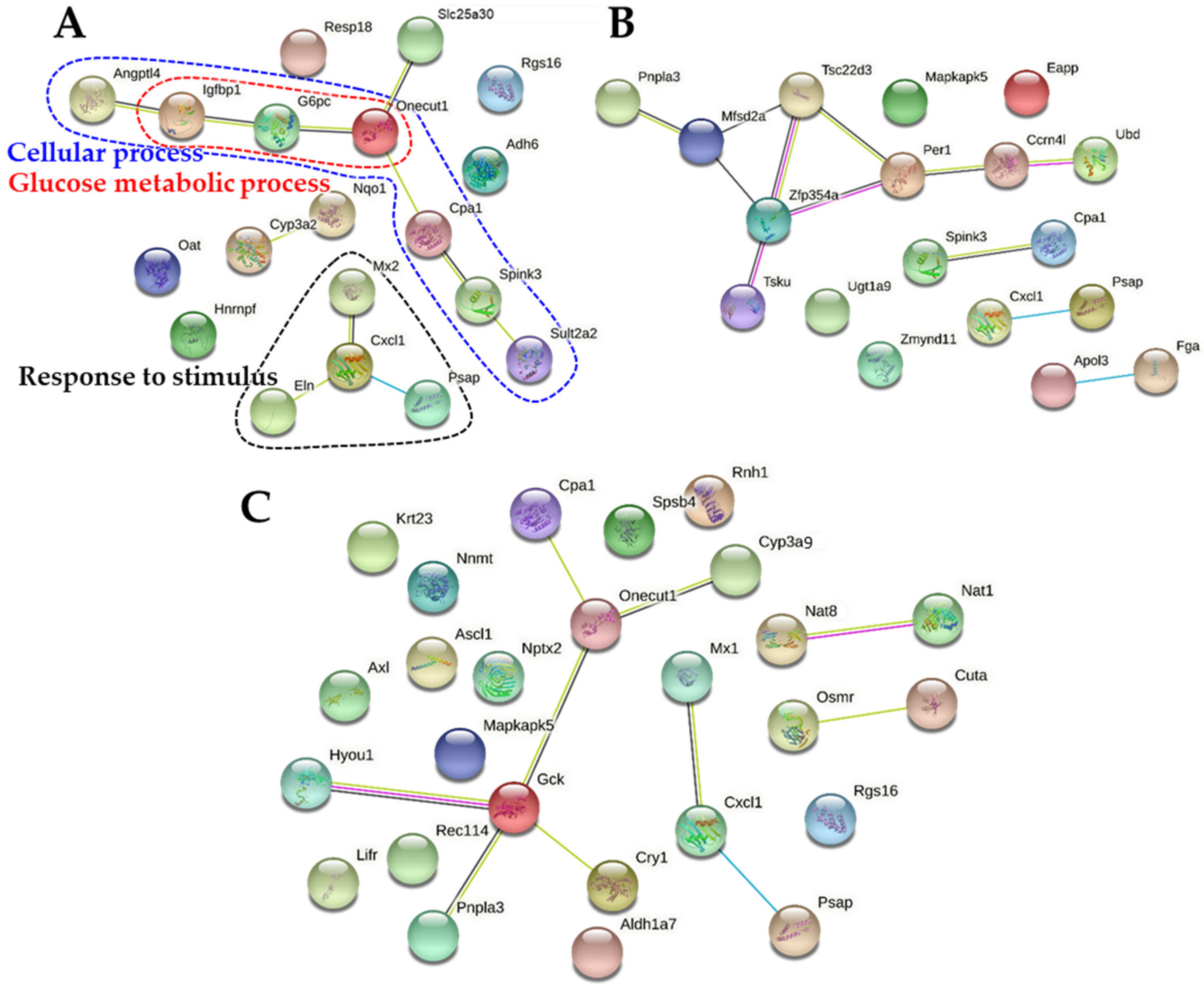
2.7. Protein-Protein Interaction Networks
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Characterization
4.3. Animals
4.4. In Vivo Dissolution Properties of ZnO Particles
4.5. Toxicokinetic Study
4.6. 14-Day Repeated Oral Toxicity and Tissue Accumulation
4.7. ICP-AES Analysis
4.8. RNA Extraction for Sequencing
4.9. RNA Sequencing and Data Analysis
4.10. Gene-Set Enrichment Analysis and PPI Networks RNA Sequencing and Data Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ANOVA | analysis of variance |
| AUC | area under the plasma concentration-time curve |
| BP | Biological Process |
| B-ZnO | bulk-sized zinc oxide |
| CC | Cellular Component |
| Cmax | maximum concentration |
| COG | Cluster of Orthologous Groups |
| CYP450 | cytochrome P450 |
| DAVID | Database for Annotation, Visualization, and Integrated Discovery |
| DEG | differentially expressed gene |
| DLS | dynamic light scattering |
| ELS | electrophoretic light scattering |
| FPKM | fragments per kilobase of transcript, per million mapped reads |
| GI | gastrointestinal |
| GO | Gene Ontology |
| H2O2 | hydrogen peroxide |
| HNO3 | nitric acid |
| ICP-AES | inductively coupled plasma–atomic emission spectroscopy |
| KEGG | Kyoto encyclopedia of genes and genomes |
| LOAEL | lowest-observed-adverse-effect level |
| MF | Molecular Function |
| MRT | mean residence time |
| NP | nanoparticle |
| N-ZnO | zinc oxide nanoparticle |
| PPI | protein-protein interaction |
| SD | Sprague Dawley |
| SEM | scanning electron microscopy |
| SP PIR | Swiss-Prot and Protein Information Resource |
| STRING | Search Tool for the Retrieval of Interacting Genes/Proteins |
| T1/2 | half-life |
| Tmax | time to maximum concentration |
| Zn | zinc |
| ZnO | zinc oxide |
References
- Ameta, S.K.; Rai, A.K.; Hiran, D.; Ameta, R.; Ameta, S.C. Use of nanomaterials in food science. In Biogenic Nano-Particles and their Use in Agro-Ecosystems, 1st ed.; Ghorbanpour, M., Bhargava, P., Varma, A., Choudhary, D., Eds.; Springer: Singapore, 2020; pp. 457–488. [Google Scholar]
- He, X.; Deng, H.; Hwang, H.M. The current application of nanotechnology in food and agriculture. J. Food Drug Anal. 2019, 27, 1–21. [Google Scholar] [CrossRef]
- Sekhon, B.S. Food nanotechnology—An overview. Nanotechnol. Sci. Appl. 2010, 3, 1–15. [Google Scholar] [PubMed]
- Jiang, J.; Pi, J.; Cai, J. The advancing of zinc oxide nanoparticles for biomedical applications. Bioinorg. Chem. Appl. 2018, 2018, 1062562. [Google Scholar] [CrossRef]
- Raya, S.D.H.A.; Hassan, M.I.; Farroh, K.Y.; Hashim, S.A.; Salaheldin, T.A. Zinc oxide nanoparticles fortified biscuits as a nutritional supplement for zinc deficient rats. J. Nanomed. Res. 2016, 4, 1–7. [Google Scholar]
- Pei, X.; Xiao, Z.; Liu, L.; Wang, G.; Tao, W.; Wang, M.; Zou, J.; Leng, D. Effects of dietary zinc oxide nanoparticles supplementation on growth performance, zinc status, intestinal morphology, microflora population, and immune response in weaned pigs. J. Sci. Food Agric. 2019, 99, 1366–1374. [Google Scholar] [CrossRef]
- Zhao, C.Y.; Tan, S.X.; Xiao, X.Y.; Qiu, X.S.; Pan, J.Q.; Tang, Z.X. Effects of dietary zinc oxide nanoparticles on growth performance and antioxidative status in broilers. Biol. Trace Elem. Res. 2014, 160, 361–367. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration (FDA). GRAS Notice. Available online: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfCFR/CFRSearch.cfm?fr=182.8991 (accessed on 1 April 2019).
- Aillon, K.L.; Xie, Y.; El-Gendy, N.; Berkland, C.J.; Forrest, M.L. Effects of nanomaterial physicochemical properties on in vivo toxicity. Adv. Drug Deliver. Rev. 2009, 61, 457–466. [Google Scholar] [CrossRef]
- Hsiao, I.L.; Huang, Y.J. Effects of serum on cytotoxicity of nano- and micro-sized ZnO particles. J. Nanopart. Res. 2013, 15, 1829. [Google Scholar] [CrossRef]
- Moos, P.J.; Chung, K.; Woessner, D.; Honeggar, M.; Cutler, N.S.; Veranth, J.M. ZnO particulate matter requires cell contact for toxicity in human colon cancer cells. Chem. Res. Toxicol. 2010, 23, 733–739. [Google Scholar] [CrossRef]
- Nair, S.; Sasidharan, A.; Rani, V.V.D.; Menon, D.; Nair, S.; Manzoor, K.; Raina, S. Role of size scale of ZnO nanoparticles and microparticles on toxicity toward bacteria and osteoblast cancer cells. J. Mater. Sci. Mater. Med. 2009, 20, S235–S241. [Google Scholar] [CrossRef]
- Sahu, D.; Kannan, G.M.; Vijayaraghavan, R. Size-dependent effect of zinc oxide on toxicity and inflammatory potential of human monocytes. J. Toxicol. Environ. A 2014, 77, 177–191. [Google Scholar] [CrossRef]
- Corbo, C.; Molinaro, R.; Parodi, A.; Furman, N.E.T.; Salvatore, F.; Tasciotti, E. The impact of nanoparticle protein corona on cytotoxicity, immunotoxicity and target drug delivery. Nanomedicine 2016, 11, 81–100. [Google Scholar] [CrossRef]
- Jo, M.R.; Yu, J.; Kim, H.J.; Song, J.H.; Kim, K.M.; Oh, J.M.; Choi, S.J. Titanium dioxide nanoparticle-biomolecule interactions influence oral absorption. Nanomaterials 2016, 6, 225. [Google Scholar] [CrossRef]
- McClements, D.J.; Xiao, H. Is nano safe in foods? Establishing the factors impacting the gastrointestinal fate and toxicity of organic and inorganic food-grade nanoparticles. NPJ Sci. Food 2017, 1, 1–13. [Google Scholar] [CrossRef]
- Yu, J.; Kim, H.J.; Go, M.R.; Bae, S.H.; Choi, S.J. ZnO interactions with biomatrices: Effect of particle size on ZnO-protein corona. Nanomaterials 2017, 7, 377. [Google Scholar] [CrossRef]
- Yu, J. Effects of Interactions Between ZnO Nanoparticles and Biological/Food Matrices on Toxicological Responses. Ph.D. Thesis, Seoul Women’s University, Seoul, Korea, 21 August 2020. [Google Scholar]
- Bae, S.H.; Yu, J.; Lee, T.G.; Choi, S.J. Protein food matrix–ZnO nanoparticle interactions affect protein conformation, but may not be biological responses. Int. J. Mol. Sci. 2018, 19, 3926. [Google Scholar] [CrossRef]
- Go, M.R.; Yu, J.; Bae, S.H.; Kim, H.J.; Choi, S.J. Effects of interactions between ZnO nanoparticles and saccharides on biological responses. Int. J. Mol. Sci. 2018, 19, 486. [Google Scholar] [CrossRef]
- Avramescu, M.L.; Rasmussen, P.E.; Chenier, M.; Gardner, H.D. Influence of pH, particle size and crystal form on dissolution behaviour of engineered nanomaterials. Environ. Sci. Pollut. Res. 2017, 24, 1553–1564. [Google Scholar] [CrossRef]
- Choi, S.J.; Choy, J.H. Biokinetics of zinc oxide nanoparticles: Toxicokinetics, biological fates, and protein interaction. Int. J. Nanomed. 2014, 9, 261–269. [Google Scholar] [CrossRef]
- Sirelkhatim, A.; Mahmud, S.; Seeni, A.; Kaus, N.H.M.; Ann, L.C.; Bakhori, S.K.M.; Hasan, H.; Mohamad, D. Review on zinc oxide nanoparticles: Antibacterial activity and toxicity mechanism. Nano Micro Lett. 2015, 7, 219–242. [Google Scholar] [CrossRef]
- Vandebriel, R.J.; Jong, W.H.D. A review of mammalian toxicity of ZnO nanoparticles. Nanotechnol. Sci. Appl. 2012, 5, 61–71. [Google Scholar] [CrossRef]
- Park, H.S.; Shin, S.S.; Meang, E.H.; Hong, J.S.; Park, J.I.; Kim, S.H.; Koh, S.B.; Lee, S.Y.; Jang, D.H.; Lee, J.Y.; et al. A 90-day study of subchronic oral toxicity of 20 nm, negatively charged zinc oxide nanoparticles in Sprague Dawley rats. Int. J. Nanomed. 2014, 9, 79–92. [Google Scholar]
- Cui, Y.; Liu, H.; Ze, Y.; Zhang, Z.; Hu, Y.; Cheng, Z.; Cheng, J.; Hu, R.; Gao, G.; Wang, L.; et al. Gene expression in liver injury caused by long-term exposure to titanium dioxide nanoparticles in mice. Toxicol. Sci. 2012, 128, 171–185. [Google Scholar] [CrossRef]
- Duescher, R.J.; Lawton, M.P.; Philpot, R.M.; Elfarra, A.A. Flavin-containing monooxygenase (FMO)-dependent metabolism of methionine and evidence for FMO3 being the major FMO involved in methionine sulfoxidation in rabbit liver and kidney microsomes. J. Biol. Chem. 1994, 269, 17525–17530. [Google Scholar] [CrossRef]
- Karmouty-Quintana, H.; Xia, Y.; Blackburn, M.R. Adenosine signaling during acute and chronic disease states. J. Mol. Med. 2013, 91, 173–181. [Google Scholar] [CrossRef]
- Escriva, L.; Jennen, D.; Caiment, F.; Manyes, L. Transcriptomic study of the toxic mechanism triggered by beauvericin in Jurkat cells. Toxicol. Lett. 2018, 284, 213–221. [Google Scholar] [CrossRef]
- Huang, C.C.; Aronstam, R.S.; Chen, D.R.; Huang, Y.W. Oxidative stress, calcium homeostasis, and altered gene expression in human lung epithelial cells exposed to ZnO nanoparticles. Toxicol. Vitr. 2010, 24, 45–55. [Google Scholar] [CrossRef]
- Cho, H.Y.; Kleeberger, S.R. Association of Nrf2 with airway pathogenesis: Lessons learned from genetic mouse models. Arch. Toxicol. 2015, 89, 1931–1957. [Google Scholar] [CrossRef]
- Falkowski, P.; Bednarek, P.; Danelska, A.; Mizerski, T.; Szafran, M. Application of monosaccharides derivatives in colloidal processing of aluminum oxide. J. Eur. Ceram. Soc. 2010, 30, 2805–2811. [Google Scholar] [CrossRef]
- Kim, M.K.; Lee, J.A.; Jo, M.R.; Choi, S.J. Bioavailability of silica, titanium dioxide, and zinc oxide nanoparticles in rats. J. Nanosci. Nanotechnol. 2016, 15, 6580–6586. [Google Scholar] [CrossRef]
- Turner, P.V.; Pekow, C.; Vasbinder, M.A.; Brabb, T. Administration of substances to laboratory animals: Equipment considerations, vehicle selection, and solute preparation. J. Am. Assoc. Lab. Anim. 2011, 50, 614–627. [Google Scholar]
- Li, X.; Yoneda, M.; Shimada, Y.; Matsui, Y. Effect of surfactants on the aggregation and sedimentation of zinc oxide nanomaterial in natural water matrices. Sci. Total Environ. 2017, 581, 649–656. [Google Scholar] [CrossRef]
- European Commission. Commission recommendation of 18 October 2011 on the definition of nanomaterial (2011/696/EU). Off. J. Eur. Union 2011, L375, 38–40. [Google Scholar]
- McConnell, E.L.; Basit, A.W.; Murdan, S. Measurements of rat and mouse gastrointestinal pH, fluid and lymphoid tissue, and implications for in-vivo experiments. J. Pharm. Pharmacol. 2008, 60, 63–70. [Google Scholar] [CrossRef]
- van der Zande, M.; Vandebriel, R.J.; Groot, M.J.; Kramer, E.; Rivera, Z.E.H.; Rasmussen, K.; Ossenkoppele, J.S.; Tromp, P.; Gremmer, E.R.; Peters, R.J.B.; et al. Sub-chronic toxicity study in rats orally exposed to nanostructured silica. Part. Fibre Toxicol. 2014, 11, 8. [Google Scholar] [CrossRef]
- Yan, X.; Rong, R.; Zhu, S.; Guo, M.; Gao, S.; Wang, S.; Xu, X. Effects of ZnO nanoparticles on dimethoate-induced toxicity in mice. J. Agric. Food Chem. 2015, 63, 8292–8298. [Google Scholar] [CrossRef]
- Paek, H.J.; Lee, Y.J.; Chung, H.E.; Yoo, N.H.; Lee, J.A.; Kim, M.K.; Lee, J.K.; Jeong, J.; Choi, S.J. Modulation of the pharmacokinetics of zinc oxide nanoparticles and their fates in vivo. Nanoscale 2013, 5, 11416–11427. [Google Scholar] [CrossRef]
- Lammel, T.; Mackevica, A.; Johansson, B.R.; Sturve, J. Endocytosis, intracellular fate, accumulation, and agglomeration of titanium dioxide (TiO2) nanoparticles in the rainbow trout liver cell line RTL-W1. Environ. Sci. Pollut. Res. 2019, 26, 15354–15372. [Google Scholar] [CrossRef]
- Banerjee, A.; Qi, J.; Gogoi, R.; Wong, J.; Mitragotri, S. Role of nanoparticle size, shape and surface chemistry in oral drug delivery. J. Control. Release 2016, 238, 176–185. [Google Scholar] [CrossRef]
- Lee, J.A.; Kim, M.K.; Song, J.H.; Jo, M.R.; Yu, J.; Kim, K.M.; Kim, Y.R.; Oh, J.M.; Choi, S.J. Biokinetics of food additive silica nanoparticles and their interactions with food components. Colloid. Surface B 2017, 150, 384–392. [Google Scholar] [CrossRef]
- Sigfridsson, K.; Nordmark, A.; Theilig, S.; Lindahl, A. A formulation comparison between micro- and nanosuspensions: The importance of particle size for absorption of a model compound, following repeated oral administration to rats during early development. Drug Dev. Ind. Pharm. 2011, 37, 185–192. [Google Scholar] [CrossRef]
- Srivastav, A.K.; Kumar, M.; Ansari, N.G.; Jain, A.K.; Shankar, J.; Arjaria, N.; Jagdale, P.; Singh, D. A comprehensive toxicity study of zinc oxide nanoparticles versus their bulk in Wistar rats: Toxicity study of zinc oxide nanoparticles. Hum. Exp. Toxicol. 2016, 35, 1286–1304. [Google Scholar] [CrossRef]
- Dekic, R.; Savic, N.; Manojlovic, M.; Golub, D.; Pavlicevic, J. Condition factor and organosomatic indices of rainbow trout (Oncorhynchus mykiss, Wal.) from different broodstock. Biotechnol. Anim. Husb. 2016, 32, 229–237. [Google Scholar] [CrossRef]
- McDonnell, A.M.; Dang, C.H. Basic review of the cytochrome P450 system. J. Adv. Pract. Oncol. 2013, 4, 263–268. [Google Scholar] [CrossRef]
- Nebert, D.W.; Russell, D.W. Clinical importance of the cytochromes P450. Lancet 2002, 360, 1155–1162. [Google Scholar] [CrossRef]
- Dunkelberger, J.R.; Song, W.C. Complement and its role in innate and adaptive immune responses. Cell Res. 2010, 20, 34–50. [Google Scholar] [CrossRef]
- Kyoreva, M.; Li, Y.; Hoosenally, M.; Hardman-Smart, J.; Morrison, K.; Tosi, I.; Tolaini, M.; Barinaga, G.; Stockinger, B.; Mrowietz, U.; et al. CYP1A1 enzymatic activity influences skin inflammation via regulation of the AHR pathway. J. Investig. Dermatol. 2020, in press. [Google Scholar] [CrossRef]
- Tian, L.X.; Tang, X.; Zhu, J.Y.; Luo, L.; Ma, X.Y.; Cheng, S.W.; Zhang, W.; Tang, W.Q.; Ma, W.; Yang, X.; et al. Cytochrome P450 1A1 enhances inflammatory responses and impedes phagocytosis of bacteria in macrophages during sepsis. Cell Commun. Signal. 2020, 18, 1–16. [Google Scholar] [CrossRef]
- Bell, S.G.; Vallee, B.L. The metallothionein/thionein system: An oxidoreductive metabolic zinc link. ChemBioChem 2009, 10, 55–62. [Google Scholar] [CrossRef]
- Starcher, B.C.; Glauber, J.G.; Madaras, J.G. Zinc absorption and its relationship to intestinal metallothionein. J. Nutr. 1980, 110, 1391–1397. [Google Scholar] [CrossRef]
- Baek, M.; Chung, H.E.; Yu, J.; Lee, J.A.; Kim, T.H.; Oh, J.M.; Lee, W.J.; Paek, S.M.; Lee, J.K.; Jeong, J.; et al. Pharmacokinetics, tissue distribution, and excretion of zinc oxide nanoparticles. Int. J. Nanomed. 2012, 7, 3081–3097. [Google Scholar] [CrossRef]
- Alexeyenko, A.; Lee, W.; Pernemalm, M.; Guegan, J.; Dessen, P.; Lazar, V.; Lehtio, J.; Pawitan, Y. Network enrichment analysis: Extension of gene-set enrichment analysis to gene networks. BMC Bioinform. 2012, 13, 226. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ZnO Particle | Fraction (Number %) | Fraction (Mass %) | Z-Average Diameter (nm) | Zeta Potential (mV) | ||||
|---|---|---|---|---|---|---|---|---|
| <100 nm | 100–200 nm | >200 nm | <100 nm | 100–200 nm | >200 nm | |||
| B-ZnO | ND 1 | ND 1 | 100.0 ± 0.0 | ND 1 | ND 1 | 100.0 ± 0.0 | 603.6 ± 1.9 a | 9.5 ± 0.4 a |
| N-ZnO | ND 1 | 9.3 ± 5.1 | 90.7 ± 5.1 | ND 1 | 1.4 ± 0.6 | 98.6 ± 0.6 | 401.1 ± 4.2 b | 12.8 ± 0.2 b |
| Toxicokinetic Parameter 1 | B-ZnO | N-ZnO | Zn Ions |
|---|---|---|---|
| Cmax (µg/mL) | 16.6 ± 2.8 a | 44.8 ± 8.5 b | 132.0 ± 37.6 c |
| Tmax (h) | 2.8 ± 1.1 a | 3.0 ± 1.2 a | 1.6 ± 0.7 b |
| AUC (h × µg/mL) | 70.8 ± 2.7 a | 212.4 ± 24.0 b | 450.6 ± 77.7 c |
| T1/2 (h) | 3.2 ± 1.1 a | 3.3 ± 1.4 a | 3.3 ± 0.5 a |
| MRT (h) | 4.1 ± 0.7 a | 4.6 ± 1.3 a | 5.8 ± 0.9 a |
| Absorption (%) | 5.0 ± 0.1 a | 14.5 ± 1.6 b | 31.4 ± 5.4 c |
| Organ | Control | B-ZnO | N-ZnO | Zn Ions |
|---|---|---|---|---|
| Kidney | 1.0 ± 0.1 | 1.1 ± 0.0 | 1.0 ± 0.1 | 1.1 ± 0.0 |
| Liver | 3.8 ± 0.5 | 4.3 ± 0.3 | 4.2 ± 0.5 | 4.1 ± 0.2 |
| Lung | 0.6 ± 0.1 | 0.6 ± 0.0 | 0.6 ± 0.0 | 0.6 ± 0.1 |
| Spleen | 0.2 ± 0.0 | 0.3 ± 0.0 | 0.3 ± 0.0 | 0.3 ± 0.0 |
| Gene | Gene Description | Fold Change | ||
|---|---|---|---|---|
| B-ZnO | N-ZnO | Zn Ions | ||
| Abhd3 | Abhydrolase domain containing 3 | 2.13 | 2.23 | 2.42 |
| Acnat2 | Acyl-coenzyme A amino acid N-acyltransferase 2 | 2.33 | ||
| Adora1 | Adenosine A1 receptor | 2.18 | ||
| Akr1b8 | Aldo-keto reductase family 1, member B8 | 2.09 | ||
| Arhgef19 | Rho guanine nucleotide exchange factor (GEF) 19 | 2.52 | 2.90 | 2.96 |
| Crot | Carnitine O-octanoyltransferase | 3.32 | 2.06 | |
| Csad | Cysteine sulfinic acid decarboxylase | 2.25 | ||
| Cyp1a1 | Cytochrome P450, family 1, subfamily a, polypeptide 1 | 3.02 | ||
| Cyp2b2 | Cytochrome P450, family 2, subfamily b, polypeptide 2 | 2.22 | ||
| Cyp2c11 | Cytochrome P450, subfamily 2, polypeptide 11 | 2.40 | ||
| Cyp2c24 | Cytochrome P450, family 2, subfamily c, polypeptide 24 | 2.60 | ||
| Cyp3a2 | Cytochrome P450, family 3, subfamily a, polypeptide 2 | 2.62 | ||
| Cyp3a18 | Cytochrome P450, family 3, subfamily a, polypeptide 18 | 2.02 | ||
| Cyp8b1 | Cytochrome P450, family 8, subfamily b, polypeptide 1 | 2.55 | 2.18 | |
| Ephx2 | Epoxide hydrolase 2, cytoplasmic | 2.03 | ||
| Fam111a | Family with sequence similarity 111, member A | 2.13 | ||
| Fam89a | Family with sequence similarity 89, member A | 2.36 | ||
| Fhl1 | Four and a half LIM domains 1 | 2.12 | ||
| Fmo1 | Flavin containing monooxygenase 1 | 2.34 | ||
| Gsta3 | Glutathione S-transferase alpha 3 | 2.86 | 2.06 | |
| Hsd11b1 | Hydroxysteroid 11-beta dehydrogenase 1 | 2.00 | ||
| Inmt | Indolethylamine N-methyltransferase | 2.55 | ||
| Lmo2 | LIM domain only 2 | 2.39 | ||
| LOC100365112 | Carboxylesterase-like precursor | 2.47 | ||
| Ly6c | Ly6-C antigen | 2.15 | ||
| Lyc2 | Lysozyme C type 2 | 2.69 | 2.66 | 2.15 |
| Moap1 | Modulator of apoptosis 1 | 3.10 | ||
| Myo9b | Myosin IXb | 3.74 | 2.10 | 2.23 |
| Nek6 | NIMA-related kinase 6 | 3.53 | ||
| Nr1d1 | Nuclear receptor subfamily 1, group D, member 1 | 2.24 | 2.15 | |
| Oxr1 | Oxidation resistance 1 | 2.10 | ||
| Pdzd3 | PDZ domain containing 3 | 2.21 | ||
| Rec114 | REC114 meiotic recombination protein | 2.37 | ||
| RGD15618 | Similar to RIKEN cDNA 3110035E14 | 2.06 | ||
| Rpl37a | Ribosomal protein L37a | 5.95 | 4.01 | 11.54 |
| Rragd | Ras-related GTP binding D | 2.07 | ||
| S100a11 | S100 calcium binding protein A11 | 2.04 | ||
| Shank2 | SH3 and multiple ankyrin repeat domains 2 | 2.03 | ||
| Slc28a2 | Solute carrier family 28 (sodium-coupled nucleoside transporter), member 2 | 2.42 | ||
| Smndc1 | Survival motor neuron domain containing 1 | 2.29 | ||
| Stmn1 | Stathmin 1 | 2.20 | ||
| Tagln | Transgelin | 2.01 | ||
| Tbc1d14 | TBC1 domain family, member 14 | 2.13 | ||
| Vnn1 | Vanin 1 | 2.72 | ||
| Wfdc6a | WAP four-disulfide core domain 6A | 2.10 | ||
| Gene | Gene Description | Fold Change | ||
|---|---|---|---|---|
| B-ZnO | N-ZnO | Zn Ions | ||
| Adh6 | Alcohol dehydrogenase 6 (class V) | −4.20 | ||
| Aldh1a7 | Aldehyde dehydrogenase, cytosolic 1 | −2.46 | ||
| Angptl4 | Angiopoietin-like 4 | −2.59 | ||
| Apol3 | Apolipoprotein L3 | −2.09 | ||
| Ascl1 | Achaete-scute family bHLH transcription factor 1 | −2.24 | ||
| Axl | Axl receptor tyrosine kinase | −2.26 | ||
| Ccrn4l | CCR4 carbon catabolite repression 4-like (S. cerevisiae) | −2.35 | ||
| Cpa1 | Carboxypeptidase A1 (pancreatic) | −2.57 | −3.03 | −2.86 |
| Cry1 | Cryptochrome circadian clock 1 | −2.08 | ||
| Cuta | CutA divalent cation tolerance homolog (E. coli) | −2.49 | ||
| Cxcl1 | Chemokine (C-X-C motif) ligand 1 | −2.90 | −3.03 | −2.77 |
| Cyp3a2 | Cytochrome P450, family 3, subfamily a, polypeptide 2 | −2.18 | ||
| Cyp3a9 | Cytochrome P450, family 3, subfamily a, polypeptide 9 | −2.38 | ||
| Eapp | E2F-associated phosphoprotein | −2.07 | ||
| Eln | Elastin | −2.17 | ||
| Fga | Fibrinogen alpha chain | −2.16 | ||
| G6pc | Glucose-6-phosphatase, catalytic subunit | −4.34 | ||
| Gck | Glucokinase | −2.08 | ||
| Hnrnpf | Heterogeneous nuclear ribonucleoprotein F | −2.13 | ||
| Hyou1 | Hypoxia up-regulated 1 | −2.42 | ||
| Igfbp1 | Insulin-like growth factor binding protein 1 | −2.17 | ||
| Krt23 | Keratin 23 (histone deacetylase inducible) | −2.82 | ||
| Lifr | Leukemia inhibitory factor receptor alpha | −3.52 | ||
| Mapkapk5 | Mitogen-activated protein kinase-activated protein kinase 5 | −3.10 | −2.19 | |
| Mfsd2a | Major facilitator superfamily domain containing 2A | −2.55 | ||
| Mx1 | Myxovirus (influenza virus) resistance 1 | −2.00 | ||
| Mx2 | Myxovirus (influenza virus) resistance 2 | −2.19 | ||
| Nat1 | N-acetyltransferase 1 (arylamine N-acetyltransferase) | −2.13 | ||
| Nat8 | N-acetyltransferase 8 | −2.07 | ||
| Nnmt | Nicotinamide N-methyltransferase | −2.16 | ||
| Nptx2 | Neuronal pentraxin Ⅱ | −2.40 | ||
| Nqo1 | NAD(P)H dehydrogenase, quinone 1 | −2.14 | ||
| Oat | Ornithine aminotransferase | −2.10 | ||
| Onecut1 | One cut homeobox 1 | −2.06 | −2.28 | |
| Osmr | Oncostatin M receptor | −2.48 | ||
| Per1 | Period circadian clock 1 | −2.38 | ||
| Pnpla3 | Patatin-like phospholipase domain containing 3 | −2.03 | −2.29 | |
| Psap | Prosaposin | −2.04 | −2.04 | −2.04 |
| Rec114 | REC114 meiotic recombination protein | −2.15 | ||
| Resp18 | Regulated endocrine-specific protein 18 | −2.56 | ||
| Rgs16 | Regulator of G-protein signaling 16 | −5.87 | −2.67 | |
| Rnh1 | Ribonuclease/angiogenin inhibitor 1 | −2.02 | ||
| Slc25a30 | Solute carrier family 25, member 30 | −2.15 | ||
| Spink3 | Serine peptidase inhibitor, Kazal type 3 | −2.08 | −2.28 | |
| Spsb4 | SplA/ryanodine receptor domain and SOCS box containing 4 | −2.06 | ||
| Sult2a2 | Sulfotransferase family 2A, dehydroepiandrosterone (DHEA)-preferring, member 2 | −2.11 | ||
| Tsc22d3 | TSC22 domain family, member 3 | −2.02 | ||
| Tsku | Tsukushi, small leucine rich proteoglycan | −2.03 | ||
| Ubd | Ubiquitin D | −2.04 | ||
| Ugt1a9 | UDP glucuronosyltransferase 1 family, polypeptide A9 | −2.35 | ||
| Zfp354a | Zinc finger protein 354A | −3.23 | ||
| Zmynd11 | Zinc finger, MYND-type containing 11 | −3.12 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Choi, S.-J. Particle Size and Biological Fate of ZnO Do Not Cause Acute Toxicity, but Affect Toxicokinetics and Gene Expression Profiles in the Rat Livers after Oral Administration. Int. J. Mol. Sci. 2021, 22, 1698. https://doi.org/10.3390/ijms22041698
Yu J, Choi S-J. Particle Size and Biological Fate of ZnO Do Not Cause Acute Toxicity, but Affect Toxicokinetics and Gene Expression Profiles in the Rat Livers after Oral Administration. International Journal of Molecular Sciences. 2021; 22(4):1698. https://doi.org/10.3390/ijms22041698
Chicago/Turabian StyleYu, Jin, and Soo-Jin Choi. 2021. "Particle Size and Biological Fate of ZnO Do Not Cause Acute Toxicity, but Affect Toxicokinetics and Gene Expression Profiles in the Rat Livers after Oral Administration" International Journal of Molecular Sciences 22, no. 4: 1698. https://doi.org/10.3390/ijms22041698
APA StyleYu, J., & Choi, S.-J. (2021). Particle Size and Biological Fate of ZnO Do Not Cause Acute Toxicity, but Affect Toxicokinetics and Gene Expression Profiles in the Rat Livers after Oral Administration. International Journal of Molecular Sciences, 22(4), 1698. https://doi.org/10.3390/ijms22041698