
Neurogenic Inflammation in the Context of Endometriosis—What Do We Know?



{kind=link}
Abstract
1. Introduction
2. Patient Presentation, Diagnosis and Current Treatments
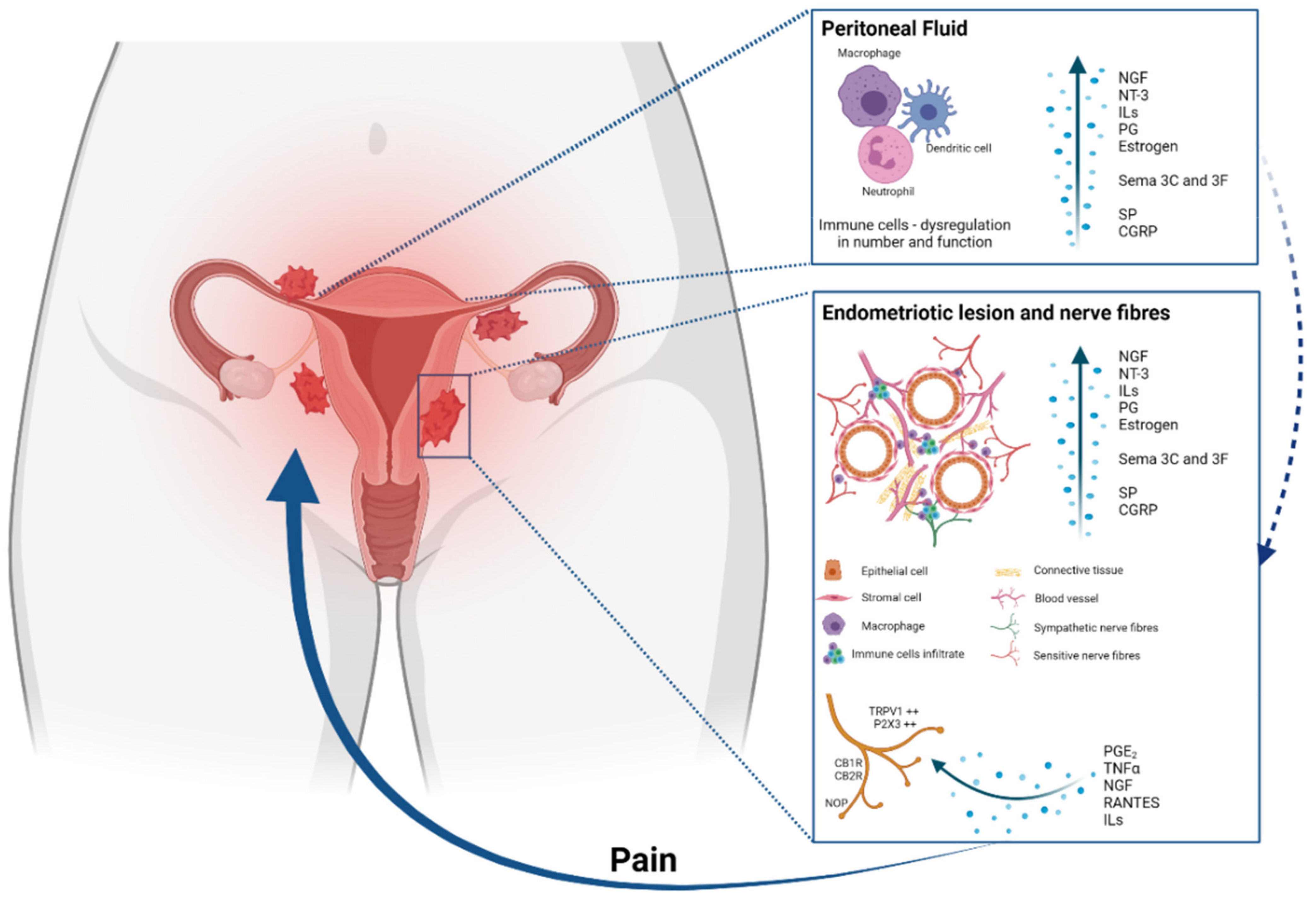
3. Pathophysiology of Endometriosis Related Pain
4. Neurogenic Inflammation
5. Neuroimmunomodulation in Endometriosis
5.1. Neuro Fibres and Neurotransmitters
5.2. Semaphorins and Neuromodulation
6. Pain Receptors in Endometriosis-Associated Nerve Fibres
6.1. Purinergic Receptors
6.2. Opioid Receptors
6.3. Endocannabinoid Receptors
7. Endometriosis and Inflammation
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Olive, D.L.; Pritts, E. Treatment of endometriosis. N. Engl. J. Med. 2001, 345, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Macer, M.L.; Taylor, H.S. Endometriosis and infertility. A review of the pathogenesis and treatment of endometriosis-associated infertility. Obstet. Gynecol. Clin. N. Am. 2012, 39, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Liang, Y.; Lin, H.; Dai, Y.; Yao, S. Autonomic nervous system and inflammation interaction in endometriosis-associated pain. J. Neuroinflam. 2020, 17, 80. [Google Scholar] [CrossRef]
- Simoens, S.; Hummelshoj, L.; D’Hooghe, T. Endometriosis: Cost estimates and methodological perspective. Hum. Reprod. Update 2007, 13, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Nnoaham, K.E.; Hummelshoj, L.; Webster, P.; d’Hooghe, T.; de Cicco Nardone, F.; de Cicco Nardone, C.; Jenkinson, C.; Kennedy, S.H.; Zondervan, K.T.; World Endometriosis Research Foundation Global Study of Women’s Health consortium. Impact of endometriosis on quality of life and work productivity: A multicenter study across ten countries. Fertil. Steril. 2011, 96, 366.e8–373.e8. [Google Scholar] [CrossRef]
- Maddern, J.; Grundy, L.; Castro, J.; Brierley, S.M. Pain in endometriosis. Front. Cell. Neurosci. 2020, 14, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Cramer, D.W.; Missmer, S.A. The epidemiology of endometriosis. Obstet. Gynecol. Clin. N. Am. 2003, 30, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Milingos, S.; Protopapas, A.; Drakakis, P.; Liapi, A.; Loutradis, D.; Kallipolitis, G.; Milingos, D.; Michalas, S. Laparoscopic management of patients with endometriosis and chronic pelvic pain. Ann. N. Y. Acad. Sci. 2003, 997, 269–273. [Google Scholar] [CrossRef]
- Abesadze, E.; Sehouli, J.; Mechsner, S.; Chiantera, V. Possible role of the posterior compartment peritonectomy, as a part of the complex surgery, regarding recurrence rate, improvement of symptoms and fertility rate in patients with endometriosis, long-term follow-up. J. Minim. Invasive Gynecol. 2019, 27, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Leyendecker, G.; Herbertz, M.; Kunz, G.; Mall, G. Endometriosis results from the dislocation of basal endometrium. Hum. Reprod. 2002, 17, 2725–2736. [Google Scholar] [CrossRef]
- Bulun, S.E.; Yilmaz, B.D.; Sison, C.; Miyazaki, K.; Bernardi, L.; Liu, S.; Kohlmeier, A.; Yin, P.; Milad, M.; Wei, J. Endometriosis. Endocr. Rev. 2019, 40, 1048–1079. [Google Scholar] [CrossRef]
- Ibrahim, M.G.; Sillem, M.; Plendl, J.; Taube, E.T.; Schüring, A.; Götte, M.; Chiantera, V.; Sehouli, J.; Mechsner, S. Arrangement of myofibroblastic and smooth muscle-like cells in superficial peritoneal endometriosis and a possible role of transforming growth factor beta 1 (TGFβ1) in myofibroblastic metaplasia. Arch. Gynecol. Obstet. 2019, 299, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Xie, H.; Wu, J.; Liu, D.; Yao, S. Villainous role of estrogen in macrophage-nerve interaction in endometriosis. Reprod. Biol. Endocrinol. 2018, 16, 122. [Google Scholar] [CrossRef] [PubMed]
- Gruber, T.M.; Mechsner, S. Pathogenesis of endometriosis: The origin of pain and subfertility. Cells 2021, 10, 1381. [Google Scholar] [CrossRef] [PubMed]
- Greene, R.; Stratton, P.; Cleary, S.D.; Ballweg, M.L.; Sinaii, N. Diagnostic experience among 4334 women reporting surgically diagnosed endometriosis. Fertil. Steril. 2009, 91, 32–39. [Google Scholar] [CrossRef]
- Dückelmann, A.M.; Taube, E.; Abesadze, E.; Chiantera, V.; Sehouli, J.; Mechsner, S. When and how should peritoneal endometriosis be operated on in order to improve fertility rates and symptoms? The experience and outcomes of nearly 100 cases. Arch. Gynecol. Obstet. 2021, 304, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Ballweg, M.L. Impact of endometriosis on women’s health: Comparative historical data show that the earlier the onset, the more severe the disease. Best Pract. Res. Clin. Obstet. Gynaecol. 2004, 18, 201–218. [Google Scholar] [CrossRef]
- Carlyle, D.; Khader, T.; Lam, D.; Vadivelu, N.; Shiwlochan, D.; Yonghee, C. Endometriosis pain management: A review. Curr. Pain Headache Rep. 2020, 24, 49. [Google Scholar] [CrossRef]
- Bazot, M.; Daraï, E. Diagnosis of deep endometriosis: Clinical examination, ultrasonography, magnetic resonance imaging, and other techniques. Fertil. Steril. 2017, 108, 886–894. [Google Scholar] [CrossRef]
- Hogg, C.; Horne, A.W.; Greaves, E. Endometriosis-associated macrophages: Origin, phenotype, and function. Front. Endocrinol. 2020, 11, 7. [Google Scholar] [CrossRef] [PubMed]
- Velho, R.V.; Halben, N.; Chekerov, R.; Keye, J.; Plendl, J.; Sehouli, J.; Mechsner, S. Functional changes of immune cells: Signal of immune tolerance of the ectopic lesions in endometriosis? Reprod. Biomed. Online 2021, 43, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Haas, D.; Chvatal, R.; Reichert, B.; Renner, S.; Shebl, O.; Binder, H.; Wurm, P.; Oppelt, P. Endometriosis: A premenopausal disease? Age pattern in 42,079 patients with endometriosis. Arch. Gynecol. Obstet. 2012, 286, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Terman, G.; Bonica, J. Spinal mechanisms and their modulation. In Bonica’s Management of Pain; Loeser, J., Butler, S., Chapman, C., Turk, D., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2001. [Google Scholar]
- Wang, G.; Tokushige, N.; Russell, P.; Dubinovsky, S.; Markham, R.; Fraser, I.S. Hyperinnervation in intestinal deep infiltrating endometriosis. J. Minim. Invasive Gynecol. 2009, 16, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Coxon, L.; Horne, A.W.; Vincent, K. Pathophysiology of endometriosis-associated pain: A review of pelvic and central nervous system mechanisms. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 51, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Bayliss, W.M. On the origin from the spinal cord of the vaso-dilator fibres of the hind-limb, and on the nature of these fibres. J. Physiol. 1901, 26, 173–209. [Google Scholar] [CrossRef]
- Arnold, J.; Vercellino, G.F.; Chiantera, V.; Schneider, A.; Mechsner, S.; de Arellano, M.L.B. Neuroimmunomodulatory alterations in non-lesional peritoneum close to peritoneal endometriosis. Neuroimmunomodulation 2013, 20, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Van der Kleij, H.P.M.; Forsythe, P.; Bienenstock, J. Autonomic neuroimmunology. In Encyclopedia of Neuroscience; Academic Press: London, England, 2009; pp. 1003–1008. [Google Scholar]
- Chiu, I.M.; von Hehn, C.A.; Woolf, C.J. Neurogenic inflammation and the peripheral nervous system in host defense and immunopathology. Nat. Neurosci. 2012, 15, 1063–1067. [Google Scholar] [CrossRef]
- Bacci, M.; Capobianco, A.; Monno, A.; Cottone, L.; Di Puppo, F.; Camisa, B.; Mariani, M.; Brignole, C.; Ponzoni, M.; Ferrari, S.; et al. Macrophages are alternatively activated in patients with endometriosis and required for growth and vascularization of lesions in a mouse model of disease. Am. J. Pathol. 2009, 175, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Anaf, V.; Chapron, C.; El Nakadi, I.; De Moor, V.; Simonart, T.; Noël, J.C. Pain, mast cells, and nerves in peritoneal, ovarian, and deep infiltrating endometriosis. Fertil. Steril. 2006, 86, 1336–1343. [Google Scholar] [CrossRef]
- Kirchhoff, D.; Kaulfuss, S.; Fuhrmann, U.; Maurer, M.; Zollner, T.M. Mast cells in endometriosis: Guilty or innocent bystanders? Expert Opin. Ther. Targets 2012, 16, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Kempuraj, D.; Papadopoulou, N.; Stanford, E.J.; Christodoulou, S.; Madhappan, B.; Sant, G.R.; Solage, K.; Adams, T.; Theoharides, T.C. Increased numbers of activated mast cells in endometriosis lesions positive for corticotropin-releasing hormone and urocortin. Am. J. Reprod. Immunol. 2004, 52, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Vallvé-Juanico, J.; Houshdaran, S.; Giudice, L.C. The endometrial immune environment of women with endometriosis. Hum. Reprod. Update 2019, 25, 565–592. [Google Scholar] [CrossRef] [PubMed]
- Rocha, M.G.; e Silva, J.C.; Ribeiro da Silva, A.; Candido Dos Reis, F.J.; Nogueira, A.A.; Poli-Neto, O.B. TRPV1 expression on peritoneal Endometriosis Foci is associated with chronic pelvic pain. Reprod. Sci. 2011, 18, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, X.; Duan, K.; Zhang, Y.; Guo, S.-W. The expression and functionality of transient receptor potential vanilloid 1 in ovarian endometriomas. Reprod. Sci. 2012, 19, 1110–1124. [Google Scholar] [CrossRef]
- Nie, J.; Liu, X.; Guo, S.-W. Immunoreactivity of oxytocin receptor and transient receptor potential vanilloid type 1 and its correlation with dysmenorrhea in adenomyosis. Am. J. Obstet. Gynecol. 2010, 202, 346.e1–346.e8. [Google Scholar] [CrossRef] [PubMed]
- Arnold, J.; Barcena de Arellano, M.L.; Rüster, C.; Vercellino, G.F.; Chiantera, V.; Schneider, A.; Mechsner, S. Imbalance between sympathetic and sensory innervation in peritoneal endometriosis. Brain. Behav. Immun. 2012, 26, 132–141. [Google Scholar] [CrossRef]
- Ferrero, S.; Haas, S.; Remorgida, V.; Camerini, G.; Fulcheri, E.; Ragni, N.; Straub, R.H.; Capellino, S. Loss of sympathetic nerve fibers in intestinal endometriosis. Fertil. Steril. 2010, 94, 2817–2819. [Google Scholar] [CrossRef]
- Nance, D.M.; Sanders, V.M. Autonomic innervation and regulation of the immune system (1987–2007). Brain. Behav. Immun. 2007, 21, 736–745. [Google Scholar] [CrossRef]
- Sanders, V.M.; Kasprowicz, D.J.; Kohm, A.P.; Swanson, M. Neurotransmitter receptors on lymphocytes and other lymphoid cells. In Psychoneuroimmunology, 1st. ed.; Ader, R., Felten, D., Cohen, N., Eds.; Academic Press: New York, NY, USA, 2001; pp. 161–196. [Google Scholar]
- Baerwald, C.; Graefe, C.; von Wichert, P.; Krause, A. Decreased density of beta-adrenergic receptors on peripheral blood mononuclear cells in patients with rheumatoid arthritis. J. Rheumatol. 1992, 19, 204–210. [Google Scholar]
- Heijnen, C.J.; Rouppe van der Voort, C.; Wulffraat, N.; van der Net, J.; Kuis, W.; Kavelaars, A. Functional α1-adrenergic receptors on leukocytes of patients with polyarticular juvenile rheumatoid arthritis. J. Neuroimmunol. 1996, 71, 223–226. [Google Scholar] [CrossRef]
- Kohm, A.P.; Sanders, V.M. Norepinephrine: A messenger from the brain to the immune system. Immunol. Today 2000, 21, 539–542. [Google Scholar] [CrossRef]
- Straub, R.H. Autoimmune disease and innervation. Brain. Behav. Immun. 2007, 21, 528–534. [Google Scholar] [CrossRef]
- Yan, D.; Liu, X.; Guo, S.W. Neuropeptides substance p and calcitonin gene related peptide accelerate the development and fibrogenesis of endometriosis. Sci. Rep. 2019, 9, 1–22. [Google Scholar] [CrossRef] [PubMed]
- de Arellano, M.L.B.; Mechsner, S. The peritoneum—An important factor for pathogenesis and pain generation in endometriosis. J. Mol. Med. 2014, 92, 595–602. [Google Scholar] [CrossRef]
- de Arellano, M.L.B.; Oldeweme, J.; Arnold, J.; Schneider, A.; Mechsner, S. Remodeling of estrogen-dependent sympathetic nerve fibers seems to be disturbed in adenomyosis. Fertil. Steril. 2013, 100, 801.e2–809.e2. [Google Scholar] [CrossRef]
- Kolodkin, A.L. Semaphorin-mediated neuronal growth cone guidance. Prog. Brain Res. 1998, 117, 115–132. [Google Scholar]
- Hui, D.H.F.; Tam, K.J.; Jiao, I.Z.F.; Ong, C.J. Semaphorin 3C as a therapeutic target in prostate and other cancers. Int. J. Mol. Sci. 2019, 20, 774. [Google Scholar] [CrossRef] [PubMed]
- Richeri, A.; Chalar, C.; Martínez, G.; Greif, G.; Bianchimano, P.; Brauer, M.M. Estrogen up-regulation of semaphorin 3F correlates with sympathetic denervation of the rat uterus. Auton. Neurosci. 2011, 164, 43–50. [Google Scholar] [CrossRef]
- Klatt, S.; Fassold, A.; Straub, R.H. Sympathetic nerve fiber repulsion: Testing norepinephrine, dopamine, and 17β-estradiol in a primary murine sympathetic neurite outgrowth assay. Ann. N. Y. Acad. Sci. 2012, 1261, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Scheerer, C.; Frangini, S.; Chiantera, V.; Mechsner, S. Reduced sympathetic innervation in endometriosis is associated to semaphorin 3C and 3F expression. Mol. Neurobiol. 2017, 54, 5131–5141. [Google Scholar] [CrossRef] [PubMed]
- Godin, S.K.; Wagner, J.; Huang, P.; Bree, D. The role of peripheral nerve signaling in endometriosis. FASEB BioAdv. 2021, 2021, 00063. [Google Scholar] [CrossRef]
- Aredo, J.; Heyrana, K.; Karp, B.; Shah, J.; Stratton, P. Relating chronic pelvic pain and endometriosis to signs of sensitization and myofascial pain and dysfunction. Semin. Reprod. Med. 2017, 35, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, D. Central and peripheral pain generators in women with chronic pelvic pain: Patient centered assessment and treatment. Curr. Rheumatol. Rev. 2015, 11, 146–166. [Google Scholar] [CrossRef] [PubMed]
- Vilotti, S.; Marchenkova, A.; Ntamati, N.; Nistri, A. B-type natriuretic peptide-induced delayed modulation of TRPV1 and P2X3 receptors of mouse trigeminal sensory neurons. PLoS ONE 2013, 8, e81138. [Google Scholar]
- Davenport, A.J.; Neagoe, I.; Bräuer, N.; Koch, M.; Rotgeri, A.; Nagel, J.; Laux-Biehlmann, A.; Machet, F.; Coelho, A.M.; Boyce, S.; et al. Eliapixant is a selective P2X3 receptor antagonist for the treatment of disorders associated with hypersensitive nerve fibers. Sci. Rep. 2021, 11, 1–13. [Google Scholar]
- Waldhoer, M.; Bartlett, S.E.; Whistler, J.L. Opioid receptors. Annu. Rev. Biochem. 2004, 73, 953–990. [Google Scholar] [CrossRef]
- Toll, L.; Bruchas, M.R.; Calo’, G.; Cox, B.M.; Zaveri, N.T. Nociceptin/orphanin FQ receptor structure, signaling, ligands, functions, and interactions with opioid systems. Pharmacol. Rev. 2016, 68, 419–457. [Google Scholar] [CrossRef]
- Totorikaguena, L.; Olabarrieta, E.; Matorras, R.; Alonso, E.; Agirregoitia, E.; Agirregoitia, N. Mu opioid receptor in the human endometrium: Dynamics of its expression and localization during the menstrual cycle. Fertil. Steril. 2017, 107, 1070.e1–1077.e1. [Google Scholar] [CrossRef]
- Chatzaki, E.; Margioris, A.N.; Makrigiannakis, A.; Castanas, E.; Georgoulias, V.; Gravanis, A. Kappa opioids and TGFbeta1 interact in human endometrial cells. Mol. Hum. Reprod. 2000, 6, 602–609. [Google Scholar] [CrossRef][Green Version]
- Hatzoglou, A.; Gravanis, A.; Margioris, A.N.; Zoumakis, E.; Castanas, E. Identification and characterization of opioid-binding sites present in the Ishikawa human endometrial adenocarcinoma cell line. J. Clin. Endocrinol. Metab. 1995, 80, 418–423. [Google Scholar]
- Zhu, Y.; Pintar, J.E. Expression of opioid receptors and ligands in pregnant mouse uterus and placenta. Biol. Reprod. 1998, 59, 925–932. [Google Scholar] [CrossRef]
- Matsuzaki, S.; Canis, M.; Vaurs-Barrière, C.; Boespflug-Tanguy, O.; Dastugue, B.; Mage, G. DNA microarray analysis of gene expression in eutopic endometrium from patients with deep endometriosis using laser capture microdissection. Fertil. Steril. 2005, 84, 1180–1190. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, S.; Canis, M.; Pouly, J.L.; Botchorishvili, R.; Déchelotte, P.J.; Mage, G. Differential expression of genes in eutopic and ectopic endometrium from patients with ovarian endometriosis. Fertil. Steril. 2006, 86, 548–553. [Google Scholar] [CrossRef]
- Matsuzaki, S.; Canis, M.; Pouly, J.L.; Botchorishvili, R.; Déchelotte, P.J.; Mage, G. Both GnRH agonist and continuous oral progestin treatments reduce the expression of the tyrosine kinase receptor B and mu-opioid receptor in deep infiltrating endometriosis. Hum. Reprod. 2007, 22, 124–128. [Google Scholar] [CrossRef][Green Version]
- Marcu, I.; Gee, A.; Lynn, B. Cannabinoids and chronic pelvic pain in women: Focus on endometriosis. J. Endometr. Pelvic Pain Disord. 2021, 13, 155–165. [Google Scholar] [CrossRef]
- Tanaka, K.; Mayne, L.; Khalil, A.; Baartz, D.; Eriksson, L.; Mortlock, S.A.; Montgomery, G.; McKinnon, B.; Amoako, A.A. The role of the endocannabinoid system in aetiopathogenesis of endometriosis: A potential therapeutic target. Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 44, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Muller, C.; Morales, P.; Reggio, P.H. Cannabinoid ligands targeting TRP channels. Front. Mol. Neurosci. 2018, 11, 487. [Google Scholar] [CrossRef] [PubMed]
- Bouaziz, J.; Bar On, A.; Seidman, D.S.; Soriano, D. The clinical significance of endocannabinoids in endometriosis pain management. Cannabis Cannabinoid Res. 2017, 2, 72–80. [Google Scholar] [CrossRef]
- Small-Howard, A.L.; Shimoda, L.M.N.; Adra, C.N.; Turner, H. Anti-inflammatory potential of CB1-mediated cAMP elevation in mast cells. Biochem. J. 2005, 388, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Leconte, M.; Nicco, C.; Ngô, C.; Arkwright, S.; Chéreau, C.; Guibourdenche, J.; Weill, B.; Chapron, C.; Dousset, B.; Batteux, F. Antiproliferative effects of cannabinoid agonists on deep infiltrating endometriosis. Am. J. Pathol. 2010, 177, 2963–2970. [Google Scholar] [CrossRef]
- Bilgic, E.; Guzel, E.; Kose, S.; Aydin, M.C.; Karaismailoglu, E.; Akar, I.; Usubutun, A.; Korkusuz, P. Endocannabinoids modulate apoptosis in endometriosis and adenomyosis. Acta Histochem. 2017, 119, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, A.M.; Cioffi, R.; Viganò, P.; Candiani, M.; Verde, R.; Piscitelli, F.; Di Marzo, V.; Garavaglia, E.; Panina-Bordignon, P. Elevated systemic levels of endocannabinoids and related mediators across the menstrual cycle in women with endometriosis. Reprod. Sci. 2016, 23, 1071–1079. [Google Scholar] [CrossRef]
- Luschnig, P.; Schicho, R. Cannabinoids in gynecological diseases. Med. Cannabis Cannabinoids 2019, 2, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, A.M.; Vigano, P.; Mugione, A.; Panina-Bordignon, P.; Candiani, M. The molecular connections between the cannabinoid system and endometriosis. Mol. Hum. Reprod. 2012, 18, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Dmitrieva, N.; Nagabukuro, H.; Resuehr, D.; Zhang, G.; McAllister, S.L.; McGinty, K.A.; Mackie, K.; Berkley, K.J. Endocannabinoid involvement in endometriosis. Pain 2010, 151, 703. [Google Scholar] [CrossRef] [PubMed]
- Armour, M.; Sinclair, J.; Chalmers, K.J.; Smith, C.A. Self-management strategies amongst Australian women with endometriosis: A national online survey. BMC Complement. Altern. Med. 2019, 19, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Burney, R.O.; Lathi, R.B. Menstrual bleeding from an endometriotic lesion. Fertil. Steril. 2009, 91, 1926–1927. [Google Scholar] [CrossRef] [PubMed]
- Králíčková, M.; Vetvicka, V. Immunological aspects of endometriosis: A review. Ann. Transl. Med. 2015, 3, 153. [Google Scholar]
- Tran, L.V.P.; Tokushige, N.; Berbic, M.; Markham, R.; Fraser, I.S. Macrophages and nerve fibres in peritoneal endometriosis. Hum. Reprod. 2009, 24, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Schulke, L.; Berbic, M.; Manconi, F.; Tokushige, N.; Markham, R.; Fraser, I.S. Dendritic cell populations in the eutopic and ectopic endometrium of women with endometriosis. Hum. Reprod. 2009, 24, 1695–1703. [Google Scholar] [CrossRef] [PubMed]
- Bulun, S.E. Endometriosis. N. Engl. J. Med. 2009, 360, 268–279. [Google Scholar] [CrossRef]
- Scheerer, C.; Bauer, P.; Chiantera, V.; Sehouli, J.; Kaufmann, A.; Mechsner, S. Characterization of endometriosis-associated immune cell infiltrates (EMaICI). Arch. Gynecol. Obstet. 2016, 294, 657–664. [Google Scholar] [CrossRef]
- Galli, S.J.; Grimbaldeston, M.; Tsai, M. Immunomodulatory mast cells: Negative, as well as positive, regulators of immunity. Nat. Rev. Immunol. 2008, 8, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Binda, M.M.; Donnez, J.; Dolmans, M.M. Targeting mast cells: A new way to treat endometriosis. Expert Opin. Ther. Targets 2017, 21, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Berbic, M.; Fraser, I.S. Regulatory T cells and other leukocytes in the pathogenesis of endometriosis. J. Reprod. Immunol. 2011, 88, 149–155. [Google Scholar] [CrossRef]
- Crispim, P.C.A.; Jammal, M.P.; Murta, E.F.C.; Nomelini, R.S. Endometriosis: What is the influence of immune cells? Immunol. Investig. 2020, 50, 372–388. [Google Scholar] [CrossRef] [PubMed]
- Capobianco, A.; Rovere-Querini, P. Endometriosis, a disease of the macrophage. Front. Immunol. 2013, 4, 1–14. [Google Scholar] [CrossRef]
- Nie, M.F.; Xie, Q.; Wu, Y.H.; He, H.; Zou, L.J.; She, X.L.; Wu, X.Q. Serum and ectopic endometrium from women with endometriosis modulate macrophage M1/M2 polarization via the Smad2/Smad3 pathway. J. Immunol. Res. 2018, 2018, 6285813. [Google Scholar] [CrossRef] [PubMed]
- Laganà, A.S.; Salmeri, F.M.; Ban Frangež, H.; Ghezzi, F.; Vrtačnik-Bokal, E.; Granese, R. Evaluation of M1 and M2 macrophages in ovarian endometriomas from women affected by endometriosis at different stages of the disease. Gynecol. Endocrinol. 2020, 36, 441–444. [Google Scholar] [CrossRef]
- Sugamata, M.; Ihara, T.; Uchiide, I. Increase of activated mast cells in human endometriosis. Am. J. Reprod. Immunol. 2005, 53, 120–125. [Google Scholar] [CrossRef]
- Paula, R.; Oliani, A.H.; Vaz-Oliani, D.C.; D’Ávila, S.C.; Oliani, S.M.; Gil, C.D. The intricate role of mast cell proteases and the annexin A1-FPR1 system in abdominal wall endometriosis. J. Mol. Histol. 2015, 46, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Burns, K.A.; Thomas, S.Y.; Hamilton, K.J.; Young, S.L.; Cook, D.N.; Korach, K.S. Early endometriosis in females is directed by immune-Mediated estrogen receptor a and IL-6 cross-Talk. Endocrinology 2018, 159, 103–118. [Google Scholar] [CrossRef]
- Wang, X.-M.; Ma, Z.-Y.; Song, N. Inflammatory cytokines IL-6, IL-10, IL-13, TNF-α and peritoneal fluid flora were associated with infertility in patients with endometriosis. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 2513–2518. [Google Scholar]
- Riccio, L.D.G.C.; Santulli, P.; Marcellin, L.; Abrão, M.S.; Batteux, F.; Chapron, C. Immunology of endometriosis. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 50, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, H.; Jin, D.; Zhang, Y. Serum miR-17, IL-4, and IL-6 levels for diagnosis of endometriosis. Medicine 2018, 97, e10853. [Google Scholar] [CrossRef]
- McKinnon, B.D.; Bertschi, D.; Bersinger, N.A.; Mueller, M.D. Inflammation and nerve fiber interaction in endometriotic pain. Trends Endrocrinol. Metab. 2015, 26, 1–10. [Google Scholar] [CrossRef]
- Zhou, J.; Chern, B.S.M.; Barton-Smith, P.; Phoon, J.W.L.; Tan, T.Y.; Viardot-Foucault, V.; Ku, C.W.; Tan, H.H.; Chan, J.K.Y.; Lee, Y.H. Peritoneal fluid cytokines reveal new insights of endometriosis subphenotypes. Int. J. Mol. Sci. 2020, 21, 3515. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velho, R.V.; Taube, E.; Sehouli, J.; Mechsner, S. Neurogenic Inflammation in the Context of Endometriosis—What Do We Know? Int. J. Mol. Sci. 2021, 22, 13102. https://doi.org/10.3390/ijms222313102
Velho RV, Taube E, Sehouli J, Mechsner S. Neurogenic Inflammation in the Context of Endometriosis—What Do We Know? International Journal of Molecular Sciences. 2021; 22(23):13102. https://doi.org/10.3390/ijms222313102
Chicago/Turabian StyleVelho, Renata Voltolini, Eliane Taube, Jalid Sehouli, and Sylvia Mechsner. 2021. "Neurogenic Inflammation in the Context of Endometriosis—What Do We Know?" International Journal of Molecular Sciences 22, no. 23: 13102. https://doi.org/10.3390/ijms222313102
APA StyleVelho, R. V., Taube, E., Sehouli, J., & Mechsner, S. (2021). Neurogenic Inflammation in the Context of Endometriosis—What Do We Know? International Journal of Molecular Sciences, 22(23), 13102. https://doi.org/10.3390/ijms222313102