
The Extracellular Vesicles from the Commensal Staphylococcus Epidermidis ATCC12228 Strain Regulate Skin Inflammation in the Imiquimod-Induced Psoriasis Murine Model

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Results
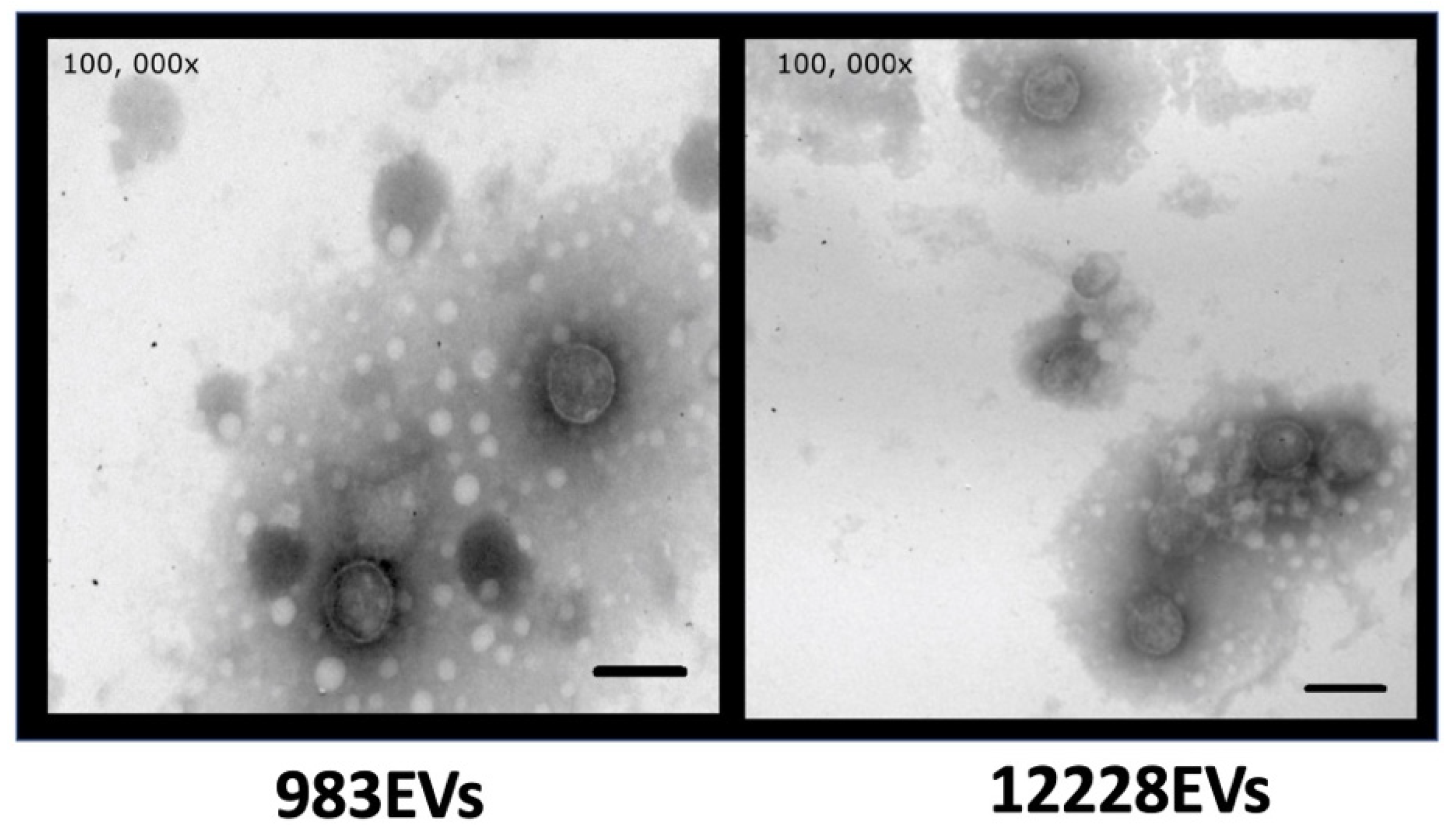
2.1. Characterization of S. epidermidis EVs
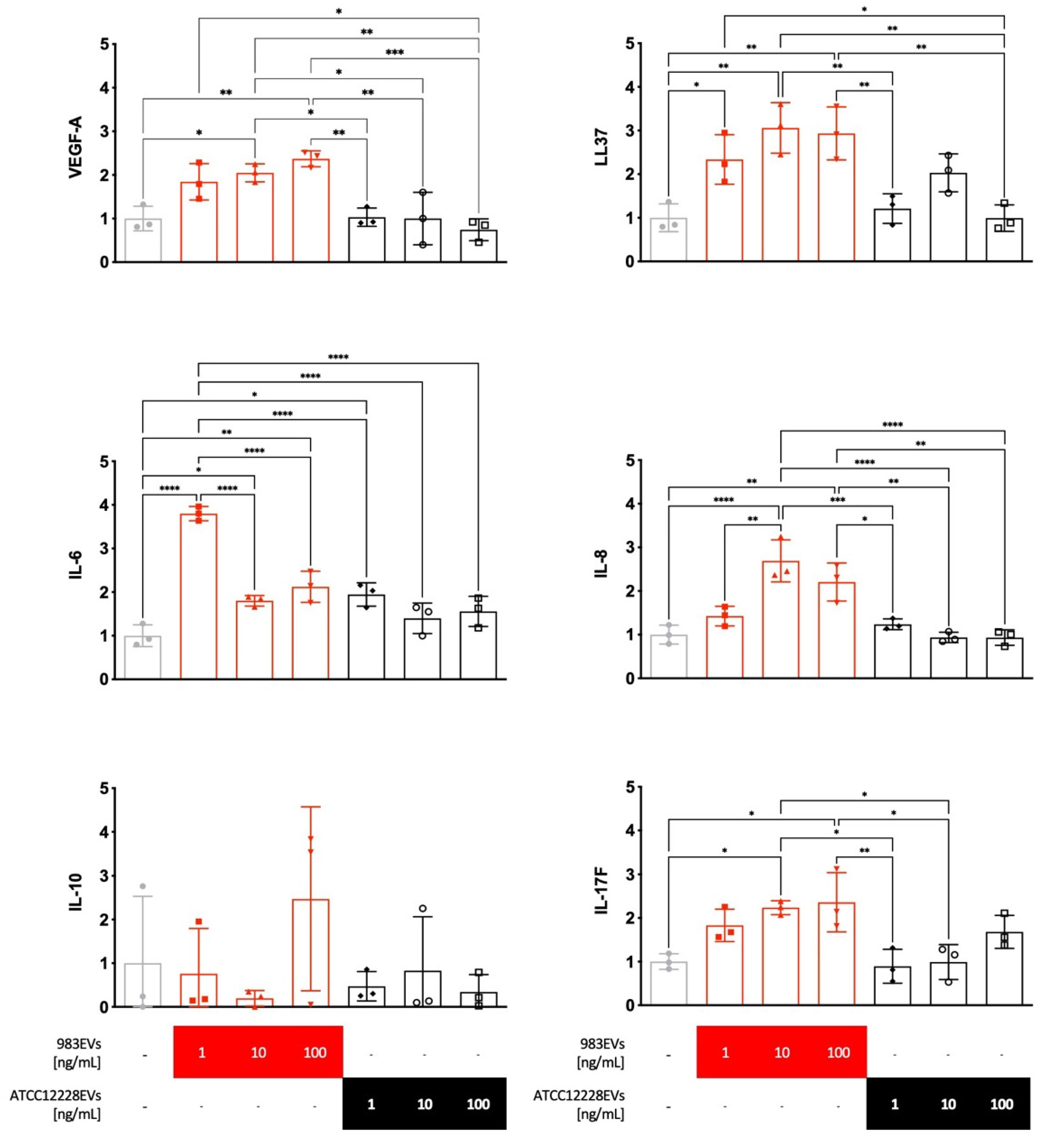
2.2. Effect of EVs on the Cytokine’s Expression in Human HaCaT Keratinocytes
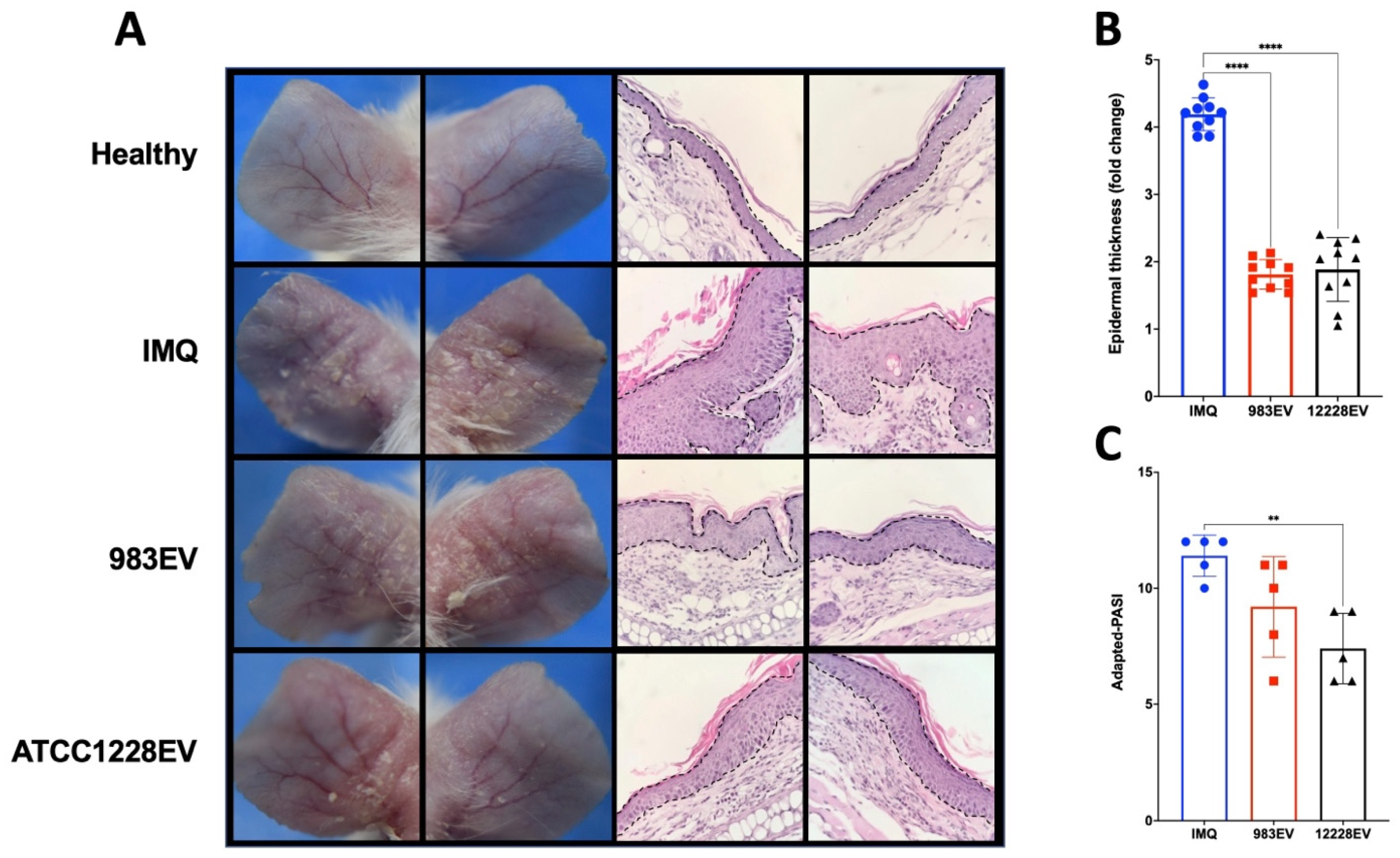
2.3. Effect of EVs on the Murine Imiquimod (IMQ)-Induced Psoriasis Model
2.4. ATCC12228EVs Decreased Cell Infiltration and Inflammatory Cytokine Expression in the IMQ-Induced Psoriasis Model
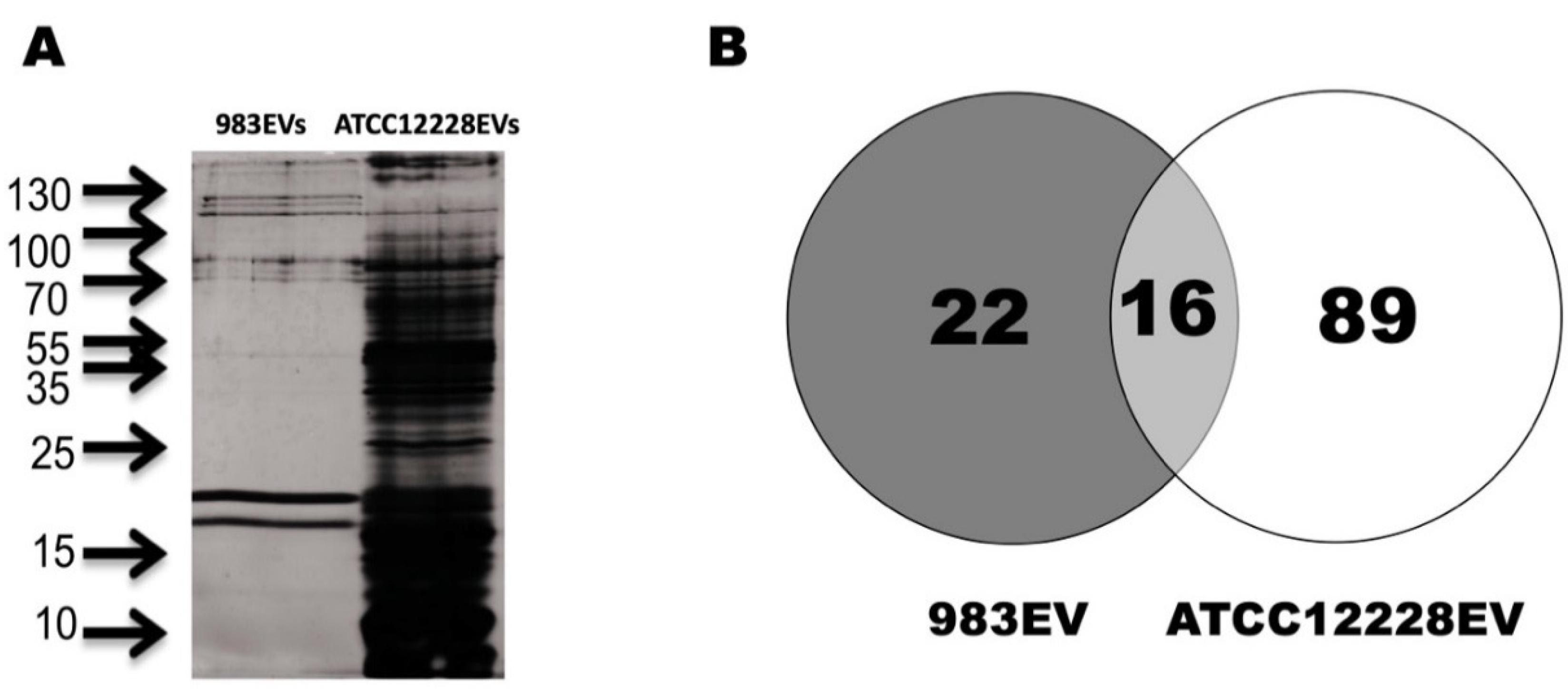
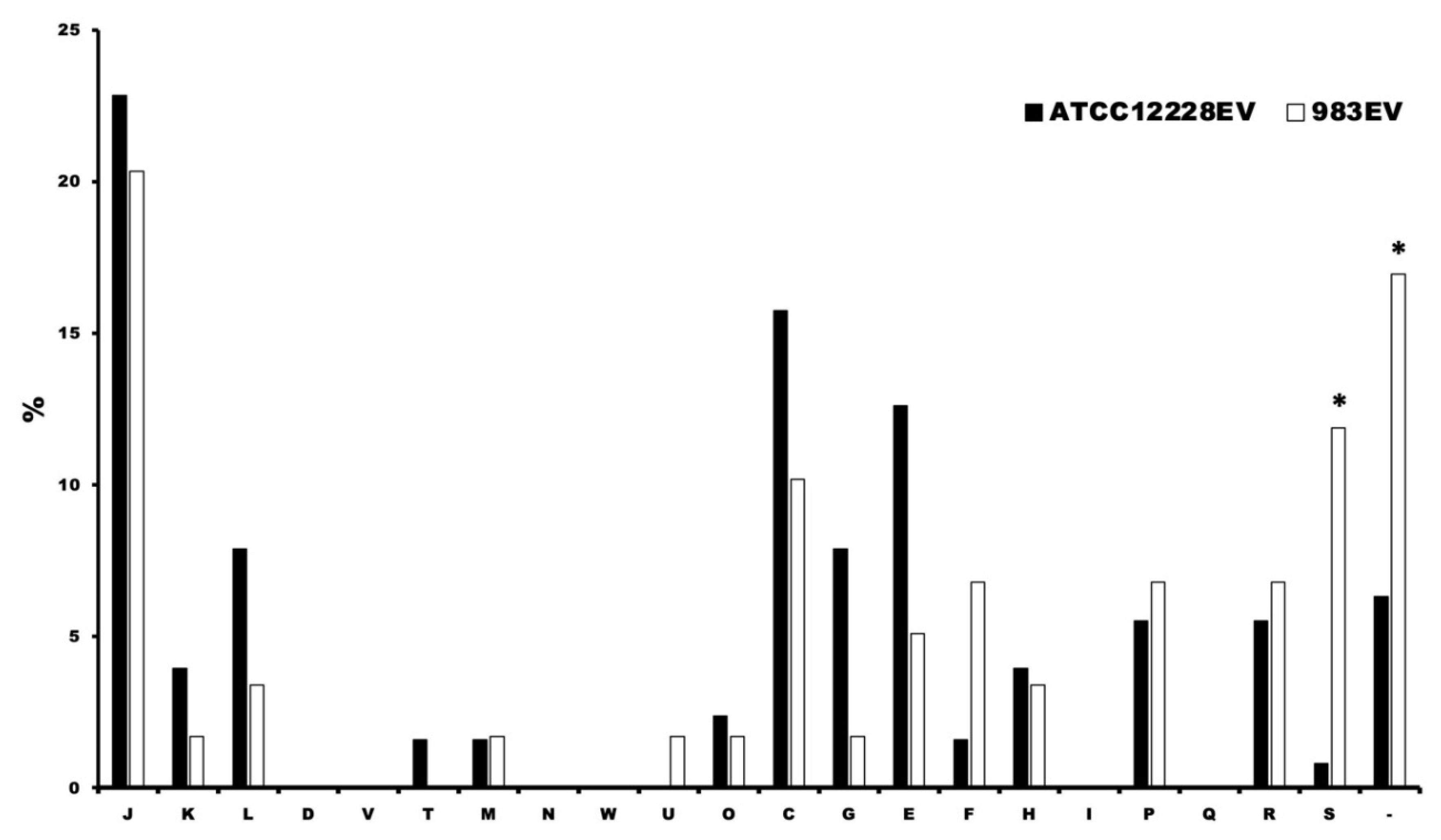
2.5. Protein Identification in EVs
2.6. Detection of Functional Protein Enrichment Using Gene Ontology Software
3. Discussion
4. Materials and Methods
4.1. EV Isolation
4.2. Observation of EVs by Electron Microscopy
4.3. Stimulation of HaCaT Keratinocytes with S. epidermidis EVs
4.4. HaCaT Keratinocyte RNA Extraction, cDNA Synthesis, and PCR
4.5. Imiquimod (IMQ)-Induced Murine Psoriasis Model Treated with Staphylococcus Epidermidis EVs
4.6. Histochemistry
4.7. Skin RT-PCR
4.8. GR1+ Cell Detection in the Skin of Ears from Treated Mice
4.9. Protein Identification by Liquid Chromatography Coupled with Tandem Mass Spectrometry (LC–MS/MS)
4.10. In Silico Analysis
4.11. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ulhuq, F.R.; Gomes, M.C.; Duggan, G.M.; Guo, M.; Mendonca, C.; Buchanan, G.; Chalmers, J.D.; Cao, Z.; Kneuper, H.; Murdoch, S.; et al. A membrane-depolarizing toxin substrate of the Staphylococcus aureus type VII secretion system mediates intraspecies competition. Proc. Natl. Acad. Sci. USA 2020, 117, 20836–20847. [Google Scholar] [CrossRef] [PubMed]
- Briaud, P.; Carroll, R.K. Extracellular vesicle biogenesis and functions in gram-positive bacteria. Infect. Immun. 2020, 88, 12. [Google Scholar] [CrossRef] [PubMed]
- Mashburn-Warren, L.M.; Whiteley, M. Special delivery: Vesicle trafficking in prokaryotes. Mol. Microbiol. 2006, 61, 839–846. [Google Scholar] [CrossRef]
- Lee, E.Y.; Choi, D.Y.; Kim, D.K.; Kim, J.W.; Park, J.O.; Kim, S.; Kim, S.H.; Desiderio, D.M.; Kim, Y.K.; Kim, K.P.; et al. Gram-positive bacteria produce membrane vesicles: Proteomics-based characterization of Staphylococcus aureus-derived membrane vesicles. Proteomics 2009, 9, 5425–5436. [Google Scholar] [CrossRef]
- Hong, S.W.; Kim, M.R.; Lee, E.Y.; Kim, J.H.; Kim, Y.S.; Jeon, S.G.; Yang, J.M.; Lee, B.J.; Pyun, B.Y.; Gho, Y.S.; et al. Extracellular vesicles derived from Staphylococcus aureus induce atopic dermatitis-like skin inflammation. Allergy 2011, 66, 351–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Defourny, K.A.Y.; Smid, E.J.; Abee, T. Gram-positive bacterial extracellular vesicles and their impact on health and disease. Front. Microbiol. 2018, 9, 1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schooling, S.R.; Hubley, A.; Beveridge, T.J. Interactions of DNA with biofilm-derived membrane vesicles. J. Bacteriol. 2009, 191, 4097–4102. [Google Scholar] [CrossRef] [Green Version]
- Maredia, R.; Devineni, N.; Lentz, P.; Dallo, S.F.; Yu, J.; Guentzel, N.; Chambers, J.; Arulanandam, B.; Haskins, W.E.; Weitao, T. Vesiculation from Pseudomonas aeruginosa under SOS. Sci. World J. 2012, 2012, 402919. [Google Scholar] [CrossRef] [Green Version]
- Gamalier, J.P.; Silva, T.P.; Zarantonello, V.; Dias, F.F.; Melo, R.C. Increased production of outer membrane vesicles by cultured freshwater bacteria in response to ultraviolet radiation. Microbiol. Res. 2017, 194, 38–46. [Google Scholar] [CrossRef]
- Manning, A.J.; Kuehn, M.J. Contribution of bacterial outer membrane vesicles to innate bacterial defense. BMC Microbiol. 2011, 11, 258. [Google Scholar] [CrossRef] [Green Version]
- Mashburn, L.M.; Whiteley, M. Membrane vesicles traffic signals and facilitate group activities in a prokaryote. Nat. Cell Biol. 2005, 437, 422–425. [Google Scholar] [CrossRef]
- Perez-Cruz, C.; Carrion, O.; Delgado, L.; Martinez, G.; Lopez-Iglesias, C.; Mercade, E. New type of outer membrane vesicle produced by the Gram-negative bacterium Shewanella vesiculosa M7T: Implications for DNA content. Appl. Environ. Microbiol. 2013, 79, 1874–1881. [Google Scholar] [CrossRef] [Green Version]
- Rivera, J.; Cordero, R.J.; Nakouzi, A.S.; Frases, S.; Nicola, A.; Casadevall, A. Bacillus anthracis produces membrane-derived vesicles containing biologically active toxins. Proc. Natl. Acad. Sci. USA 2010, 107, 19002–19007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciofu, O.; Beveridge, T.J.; Kadurugamuwa, J.; Walther-Rasmussen, J.; Hoiby, N. Chromosomal beta-lactamase is packaged into membrane vesicles and secreted from Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2000, 45, 9–13. [Google Scholar] [CrossRef]
- Lee, J.; Lee, E.Y.; Kim, S.H.; Kim, D.K.; Park, K.S.; Kim, K.P.; Kim, Y.K.; Roh, T.Y.; Gho, Y.S. Staphylococcus aureus extracellular vesicles carry biologically active beta-lactamase. Antimicrob. Agents Chemother. 2013, 57, 2589–2595. [Google Scholar] [CrossRef] [Green Version]
- Askarian, F.; Lapek, J.D., Jr.; Dongre, M.; Tsai, C.-M.; Kumaraswamy, M.; Kousha, A.; Valderrama, J.A.; Ludviksen, J.A.; Cavanagh, P.; Uchiyama, S.; et al. Staphylococcus aureus membrane-derived vesicles promote bacterial virulence and confer protective immunity in murine infection models. Front. Microbiol. 2018, 9, 262. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.H.; Lee, J.H.; Kim, S.I.; Choi, C.W.; Park, T.I.; Jung, H.R.; Cho, J.W.; Kim, S.H.; Lee, J.C. Staphylococcus aureus-derived membrane vesicles exacerbate skin inflammation in atopic dermatitis. Clin. Exp. Allergy 2017, 47, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Gurung, M.; Moon, D.C.; Choi, C.W.; Lee, J.H.; Bae, Y.C.; Kim, J.; Lee, Y.C.; Seol, S.Y.; Cho, D.T.; Kim, S.I.; et al. Staphylococcus aureus produces membrane-derived vesicles that induce host cell death. PLoS ONE 2011, 6, e27958. [Google Scholar] [CrossRef] [Green Version]
- Thay, B.; Wai, S.N.; Oscarsson, J. Staphylococcus aureus alpha-toxin-dependent induction of host cell death by membrane-derived vesicles. PLoS ONE 2013, 8, e54661. [Google Scholar] [CrossRef]
- Chebotar, I.; Konchakova, E.D.; Maianskii, A.N. Vesicle formation as a result of interaction between polymorphonuclear neutrophils and Staphylococcus aureus biofilm. J. Med. Microbiol. 2013, 62 Pt 8, 1153–1159. [Google Scholar] [CrossRef]
- Deng, S.; Wang, Y.; Liu, S.; Chen, T.; Hu, Y.; Zhang, G.; Zhang, X.; Yu, B. Extracellular vesicles: A potential biomarker for quick identification of infectious osteomyelitis. Front. Cell. Infect. Microbiol. 2020, 10, 323. [Google Scholar] [CrossRef]
- Wang, Y.; Kuo, S.; Shu, M.; Yu, J.; Huang, S.; Dai, A.; Two, A.; Gallo, R.L.; Huang, C.-M. Staphylococcus epidermidis in the human skin microbiome mediates fermentation to inhibit the growth of Propionibacterium acnes: Implications of probiotics in acne vulgaris. Appl. Microbiol. Biotechnol. 2013, 98, 411–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leech, J.M.; Dhariwala, M.O.; Lowe, M.M.; Chu, K.; Merana, G.R.; Cornuot, C.; Weckel, A.; Ma, J.M.; Leitner, E.G.; Gonzalez, J.R.; et al. Toxin-triggered interleukin-1 receptor signaling enables early-life discrimination of pathogenic versus commensal skin bacteria. Cell Host Microbe 2019, 26, 795–809.e5. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Li, Z.; Liu, K.; Wu, Y.; Jiang, D.; Lai, Y. Staphylococcal LTA-induced miR-143 inhibits Propionibacterium acnes-Mediated inflammatory response in skin. J. Investig. Dermatol. 2016, 136, 621–630. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.-W.; Yan, D.; Singh, R.; Liu, J.; Lu, X.; Ucmak, D.; Lee, K.; Afifi, L.; Fadrosh, D.; Leech, J.M.; et al. Alteration of the cutaneous microbiome in psoriasis and potential role in Th17 polarization. Microbiome 2018, 6, 1–27. [Google Scholar] [CrossRef]
- Lowes, M.A.; Bowcock, A.M.; Krueger, J.G. Pathogenesis and therapy of psoriasis. Nat. Cell Biol. 2007, 445, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Nestle, F.O.; Kaplan, D.H.; Barker, J. Psoriasis. N. Engl. J. Med. 2009, 361, 496–509. [Google Scholar] [CrossRef]
- Al-Harbi, N.O.; Nadeem, A.; Ahmad, S.F.; Bakheet, S.A.; El-Sherbeeny, A.M.; Ibrahim, K.E.; Alzahrani, K.S.; Al-Harbi, M.M.; Mahmood, H.M.; Alqahtani, F.; et al. Therapeutic treatment with Ibrutinib attenuates imiquimod-induced psoriasis-like inflammation in mice through downregulation of oxidative and inflammatory mediators in neutrophils and dendritic cells. Eur. J. Pharmacol. 2020, 877, 173088. [Google Scholar] [CrossRef]
- Nadeem, A.; Ahmad, S.F.; Al-Harbi, N.O.; El-Sherbeeny, A.M.; Alasmari, A.F.; Alanazi, W.A.; Alasmari, F.; Ibrahim, K.E.; Al-Harbi, M.M.; Bakheet, S.A.; et al. Bruton’s tyrosine kinase inhibitor suppresses imiquimod-induced psoriasis-like inflammation in mice through regulation of IL-23/IL-17A in innate immune cells. Int. Immunopharmacol. 2020, 80, 106215. [Google Scholar] [CrossRef] [PubMed]
- Stacy, A.; Belkaid, Y. Microbial guardians of skin health. Science 2019, 363, 227–228. [Google Scholar] [CrossRef]
- Zhou, W.; Spoto, M.; Hardy, R.; Guan, C.; Fleming, E.; Larson, P.; Brown, J.S.; Oh, J. Host-specific evolutionary and transmission dynamics shape the functional diversification of staphylococcus epidermidis in human skin. Cell 2020, 180, 454–470.e18. [Google Scholar] [CrossRef] [PubMed]
- Zaborowska, M.; Flores, C.T.; Vazirisani, F.; Shah, F.A.; Thomsen, P.; Trobos, M. Extracellular vesicles influence the growth and adhesion of staphylococcus epidermidis under antimicrobial selective pressure. Front. Microbiol. 2020, 11, 1132. [Google Scholar] [CrossRef] [PubMed]
- Tovar-Castillo, L.E.; Cancino-Díaz, J.C.; García-Vázquez, F.; Cancino-Gómez, F.G.; León-Dorantes, G.; Blancas-González, F.; Jiménez-Zamudio, L.; García-Latorre, E.; Cancino-Díaz, M.E. Under-expression of VHL and over-expression of HDAC-1, HIF-1alpha, LL-37 and IAP-2 in affected skin biopsies of patients with psoriasis. Int. J. Dermatol. 2007, 46, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Lande, R.; Gregorio, J.; Facchinetti, V.; Chatterjee, B.; Wang, Y.H.; Homey, B.; Cao, W.; Wang, Y.H.; Su, B.; Nestle, F.O.; et al. Plasmacytoid dendritic cells sense self-DNA coupled with antimicrobial peptide. Nature 2007, 449, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, D.; Chamilos, G.; Lande, R.; Gregorio, J.; Meller, S.; Facchinetti, V.; Homey, B.; Barrat, F.J.; Zal, T.; Gilliet, M. Self-RNA–antimicrobial peptide complexes activate human dendritic cells through TLR7 and TLR8. J. Exp. Med. 2009, 206, 1983–1994. [Google Scholar] [CrossRef]
- Li, N.; Yamasaki, K.; Saito, R.; Fukushi-Takahashi, S.; Shimada-Omori, R.; Asano, M.; Aiba, S. Alarmin function of cathelicidin antimicrobial peptide LL37 through IL-36gamma induction in human epidermal keratinocytes. J. Immunol. 2014, 193, 5140–5148. [Google Scholar] [CrossRef] [Green Version]
- Hansel, A.; Gunther, C.; Ingwersen, J.; Starke, J.; Schmitz, M.; Bachmann, M.; Meurer, M.; Rieber, E.P.; Schakel, K. Human slan (6-sulfo LacNAc) dendritic cells are inflammatory dermal dendritic cells in psoriasis and drive strong TH17/TH1 T-cell responses. J. Allergy Clin. Immunol. 2011, 127, 787–794.e1–9. [Google Scholar] [CrossRef]
- Goodman, W.A.; Levine, A.D.; Massari, J.V.; Sugiyama, H.; McCormick, T.S.; Cooper, K.D. IL-6 Signaling in psoriasis prevents immune suppression by regulatory T Cells. J. Immunol. 2009, 183, 3170–3176. [Google Scholar] [CrossRef] [Green Version]
- Bartoccioni, E.; Scuderi, F.; Marino, M.; Provenzano, C. IL-6, monocyte infiltration and parenchymal cells. Trends Immunol. 2003, 24, 298–299. [Google Scholar] [CrossRef]
- Lindroos, J.; Svensson, L.; Norsgaard, H.; Lovato, P.; Moller, K.; Hagedorn, P.H.; Olsen, G.M.; Labuda, T. IL-23-mediated epidermal hyperplasia is dependent on IL-6. J. Investig. Dermatol. 2011, 131, 1110–1118. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, H.; Kawaguchi, M.; Fujishima, S.; Ogura, M.; Matsukura, S.; Takeuchi, H.; Ohba, M.; Sueki, H.; Kokubu, F.; Hizawa, N.; et al. Functional characterization of IL-17F as a selective neutrophil attractant in psoriasis. J. Investig. Dermatol. 2009, 129, 650–656. [Google Scholar] [CrossRef] [Green Version]
- Tortola, L.; Rosenwald, E.; Abel, B.; Blumberg, H.; Schäfer, M.; Coyle, A.J.; Renauld, J.-C.; Werner, S.; Kisielow, J.; Kopf, M. Psoriasiform dermatitis is driven by IL-36–mediated DC-keratinocyte crosstalk. J. Clin. Investig. 2012, 122, 3965–3976. [Google Scholar] [CrossRef]
- Uribe-Herranz, M.; Lian, L.-H.; Hooper, K.M.; Milora, K.A.; Jensen, L.E. IL-1R1 Signaling Facilitates Munro’s Microabscess Formation in Psoriasiform Imiquimod-Induced Skin Inflammation. J. Investig. Dermatol. 2013, 133, 1541–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, J.D.; Bassoy, E.Y.; Caruso, A.; Palomo, J.; Rodriguez, E.; Lemeille, S.; Gabay, C. IL-36 signaling in keratinocytes controls early IL-23 production in psoriasis-like dermatitis. Life Sci. Alliance 2020, 3, e202000688. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Santana, Y.E.; Leon, G.; Leger, D.S.; Fallon, P.; Walsh, P.T. Keratinocyte interleukin-36 receptor expression orchestrates psoriasiform inflammation in mice. Life Sci. Alliance 2020, 3, e201900586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furue, K.; Yamamura, K.; Tsuji, G.; Mitoma, C.; Uchi, H.; Nakahara, T.; Kido-Nakahara, M.; Kadono, T.; Furue, M. Highlighting Interleukin-36 signalling in plaque psoriasis and pustular psoriasis. Acta Derm. Venereol. 2018, 98, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Foster, A.M.; Baliwag, J.; Chen, C.S.; Guzman, A.M.; Stoll, S.W.; Gudjonsson, J.E.; Ward, N.L.; Johnston, A. IL-36 promotes myeloid cell infiltration, activation, and inflammatory activity in skin. J. Immunol. 2014, 192, 6053–6061. [Google Scholar] [CrossRef] [Green Version]
- Hashiguchi, Y.; Yabe, R.; Chung, S.-H.; Murayama, M.A.; Yoshida, K.; Matsuo, K.; Kubo, S.; Saijo, S.; Nakamura, Y.; Matsue, H.; et al. IL-36α from skin-resident cells plays an important role in the pathogenesis of imiquimod-induced psoriasiform dermatitis by forming a local autoamplification loop. J. Immunol. 2018, 201, 167–182. [Google Scholar] [CrossRef]
- Bassoy, E.Y.; Towne, J.E.; Gabay, C. Regulation and function of interleukin-36 cytokines. Immunol. Rev. 2018, 281, 169–178. [Google Scholar] [CrossRef]
- Mercurio, L.; Morelli, M.; Scarponi, C.; Eisenmesser, E.Z.; Doti, N.; Pagnanelli, G.; Gubinelli, E.; Mazzanti, C.; Cavani, A.; Ruvo, M.; et al. IL-38 has an anti-inflammatory action in psoriasis and its expression correlates with disease severity and therapeutic response to anti-IL-17A treatment. Cell Death Dis. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Canavese, M.; Altruda, F.; Silengo, L.; Castiglioni, V.; Scanziani, E.; Radaelli, E. Clinical, pathological and immunological features of psoriatic-like lesions affecting keratin 14-vascular endothelial growth factor transgenic mice. Histol. Histopathol. 2011, 26, 285–296. [Google Scholar]
- Hvid, H.; Teige, I.; Kvist, P.H.; Svensson, L.; Kemp, K. TPA induction leads to a Th17-like response in transgenic K14/VEGF mice: A novel in vivo screening model of psoriasis. Int. Immunol. 2008, 20, 1097–1106. [Google Scholar] [CrossRef] [Green Version]
- Schlatterer, K.; Beck, C.; Hanzelmann, D.; Lebtig, M.; Fehrenbacher, B.; Schaller, M.; Ebner, P.; Nega, M.; Otto, M.; Kretschmer, D.; et al. The mechanism behind bacterial lipoprotein release: Phenol-soluble modulins mediate toll-like receptor 2 activation via extracellular vesicle release from Staphylococcus aureus. mBio 2018, 9, e01851-18. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, K.; Vasconcellos, C.I.; Soares, R.P.; Mendes, M.T.; Ellis, C.C.; Aguilera-Flores, M.; Almeida, I.; Schenkman, S.; Iwai, L.K.; Torrecilhas, A.C. Proteomic analysis reveals different composition of extracellular vesicles released by two Trypanosoma cruzi strains associated with their distinct interaction with host cells. J. Extracell. Vesicles 2018, 7, 1463779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.-Q.; Ren, S.-X.; Li, H.-L.; Wang, Y.-X.; Fu, G.; Yang, J.; Qin, Z.-Q.; Miao, Y.-G.; Wang, W.-Y.; Chen, R.-S.; et al. Genome-based analysis of virulence genes in a non-biofilm-forming Staphylococcus epidermidis strain (ATCC 12228). Mol. Microbiol. 2003, 49, 1577–1593. [Google Scholar] [CrossRef] [Green Version]
- Hussain, H.; Ziegler, J.; Mrestani, Y.; Neubert, R.H.H. Studies of the corneocytary pathway across the stratum corneum. Part I: Diffusion of amino acids into the isolated corneocytes. Die Pharm. Int. J. Pharm. Sci. 2019, 74, 340–344. [Google Scholar]
- Armstrong, A.W.; Wu, J.; Johnson, M.A.; Grapov, D.; Azizi, B.; Dhillon, J.; Fiehn, O. Metabolomics in psoriatic disease: Pilot study reveals metabolite differences in psoriasis and psoriatic arthritis. F1000Research 2014, 3, 248. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.K.; Ryder, J.E. Photodynamic therapy and topical aminolevulinic acid: An overview. Am. J. Clin. Dermatol. 2003, 4, 699–708. [Google Scholar] [CrossRef]
- Luqman, A.; Muttaqin, M.Z.; Yulaipi, S.; Ebner, P.; Matsuo, M.; Zabel, S.; Tribelli, P.M.; Nieselt, K.; Hidayati, D.; Götz, F. Trace amines produced by skin bacteria accelerate wound healing in mice. Commun. Biol. 2020, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Zhao, W.; Zhang, B.; Tu, Y.; Wang, Q.; Li, J. Cimifugin ameliorates imiquimod-induced psoriasis by inhibiting oxida-tive stress and inflammation via NF-kappaB/MAPK pathway. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef]
- Zhang, Y.; Jia, H.; Jin, Y.; Liu, N.; Chen, J.; Yang, Y.; Dai, Z.; Wang, C.; Wu, G.; Wu, Z. Glycine attenuates LPS-induced apoptosis and inflammatory cell infiltration in mouse liver. J. Nutr. 2020, 150, 1116–1125. [Google Scholar] [CrossRef]
- Wang, W.-M.; Jin, H.-Z. Role of neutrophils in psoriasis. J. Immunol. Res. 2020, 2020, 3709749. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Di Nardo, A.; Nakatsuji, T.; Leichtle, A.; Yang, Y.; Cogen, A.L.; Wu, Z.R.; Hooper, L.V.; Schmidt, R.R.; von Au-lock, S.; et al. Commensal bacteria regulate Toll-like receptor 3-dependent inflammation after skin injury. Nat. Med. 2009, 15, 1377–1382. [Google Scholar] [CrossRef]
- Iovieno, A.; Lambiase, A.; Sacchetti, M.; Stampachiacchiere, B.; Micera, A.; Bonini, S. Preliminary evidence of the efficacy of probiotic eye-drop treatment in patients with vernal keratoconjunctivitis. Graefes Arch. Clin. Exp. Ophthalmol. 2008, 246, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Cosseau, C.; Devine, D.A.; Dullaghan, E.; Gardy, J.L.; Chikatamarla, A.; Gellatly, S.; Yu, L.L.; Pistolic, J.; Falsafi, R.; Tagg, J.; et al. The commensal Streptococcus salivarius K12 downregulates the innate immune responses of human epithelial cells and promotes host-microbe homeostasis. Infect. Immun. 2008, 76, 4163–4175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volz, T.; Skabytska, Y.; Guenova, E.; Chen, K.M.; Frick, J.S.; Kirschning, C.J.; Kaesler, S.; Rocken, M.; Biedermann, T. Nonpathogenic bacteria alleviating atopic dermatitis inflammation induce IL-10-producing dendritic cells and regulatory Tr1 cells. J. Investig. Dermatol. 2014, 134, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Flores-Páez, L.A.; Zenteno, J.C.; Alcántar-Curiel, M.D.; Vargas-Mendoza, C.F.; Rodriguez, S.; Cancino-Diaz, M.E.; Jan-Roblero, J.; Cancino-Diaz, J.C. Molecular and phenotypic characterization of staphylococcus epidermidis isolates from healthy conjunctiva and a comparative analysis with isolates from ocular infection. PLoS ONE 2015, 10, e0135964. [Google Scholar] [CrossRef]
- Avila-Calderon, E.D.; Lopez-Merino, A.; Jain, N.; Peralta, H.; Lopez-Villegas, E.O.; Sriranganathan, N.; Boyle, S.M.; Witonsky, S.; Contreras-Rodriguez, A. Characterization of outer membrane vesicles from Brucella melitensis and protection induced in mice. Clin. Dev. Immunol. 2012, 2012, 352493. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Palma, M.D.S.; Avila-Calderón, E.D.; Aguilera-Arreola, M.G.; López-Merino, A.; Ruiz, E.A.; Morales-García, M.D.R.; López-Villegas, E.O.; Gomez-Lunar, Z.; Arellano-Reynoso, B.; Contreras-Rodríguez, A. Comparative proteomic analysis of outer membrane vesicles from Brucella suis, Brucella ovis, Brucella canis and Brucella neotomae. Arch. Microbiol. 2021, 203, 1611–1626. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Garcia, S.; Ortega-Pena, S.; De Haro-Cruz, M.J.; Aguilera-Arreola, M.G.; Alcantar-Curiel, M.D.; Betan-zos-Cabrera, G.; Jan-Roblero, J.; Perez-Tapia, S.M.; Rodriguez-Martinez, S.; Cancino-Diaz, M.E.; et al. Non-biofilm-forming commensal Staphylococcus epidermidis isolates produce biofilm in the presence of trypsin. Microbiologyopen 2019, 8, e906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Fits, L.; Mourits, S.; Voerman, J.S.; Kant, M.; Boon, L.; Laman, J.D.; Cornelissen, F.; Mus, A.M.; Florencia, E.; Prens, E.P.; et al. Imiquimod-induced psoriasis-like skin inflammation in mice is mediated via the IL-23/IL-17 axis. J. Immunol. 2009, 182, 5836–5845. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to imagej: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Accession | COG Number | Gene Name | Function |
|---|---|---|---|---|
| J | Q8CQV5 | COG1190 | lysS | Lysine–tRNA ligase |
| J | Q8CSD5 | COG0423 | glyQS | Anticodon-binding domain protein |
| J | Q8CSY9 | COG0016 | pheS | Phenylalanine–tRNA ligase alpha subunit |
| J | Q8CRH9 | COG0200 | rplO | 50S ribosomal protein L5 |
| J | Q8CS87 | COG0261 | rplU | 50S ribosomal protein L21 |
| J | Q8CRI3 | COG0099 | rpsM | 30S ribosomal protein S13 |
| J | P66618 | COG0049 | rpsG | 30S ribosomal protein S7 |
| J | Q8CRH2 | COG0094 | rplE | 50S ribosomal protein L5 |
| J | Q8CRH1 | COG0198 | rplX | 50S ribosomal protein L24 |
| J | Q8CTT4 | COG0081 | rplA | 50S ribosomal protein L1 |
| J | Q8CRH7 | COG0098 | rpsE | 30S ribosomal protein S5 |
| J | Q8CRG2 | COG0089 | rplW | 50S ribosomal protein L23 |
| J | Q8CS77 | COG0292 | rplT | 50S ribosomal protein L20 |
| J | Q8CRI5 | COG0203 | rplQ | 50S ribosomal protein L17 |
| J | Q8CTT5 | COG0080 | rplK | 50S ribosomal protein L11 |
| J | Q8CRJ0 | COG0103 | rpsI | 30S ribosomal protein S9 |
| J | Q8CRG4 | COG0185 | rpsS | 30S ribosomal protein S19 |
| J | P66336 | COG0051 | rpsJ | 30S ribosomal protein S10 |
| J | Q8CRH6 | COG0256 | rplR | 50S ribosomal protein L18 |
| J | Q8CRH4 | COG0096 | rpsH | 30S ribosomal protein S8 |
| K | Q8CQ84 | COG0085 | rpoB | DNA-directed RNA polymerase subunit beta |
| K | Q8CNU6 | COG1846 | Rot | HTH-type transcriptional regulator rot |
| K | Q8CRX8 | COG1476 | Cro | Cro/Cl family transcriptional regulator |
| K | Q8CRJ1 | COG1349 | lacR | Lactose phosphotransferase system repressor |
| L | Q8CNN0 | COG0582 | tnpA | Transposase A |
| L | Q8CP25 | COG0648 | Nfo | Probable endonuclease 4 |
| L | Q8CPC6 | COG0420 | sbcD | Exonuclease SbcCD, C subunit |
| L | Q8CP04 | COG0507 | - | Helicase, RecD/TraA family |
| L | Q8CRP6 | COG0513 | cshA | DEAD/DEAH box helicase domain protein |
| L | Q8CNX7 | COG0258 | polA | DNA polymerase I |
| L | Q8CPT9 | COG1074 | addA | Helicase-exonuclease AddAB, AddB subunit |
| L | Q8CNX7 | COG0258 | polA | DNA polymerase |
| L | Q8CPZ0 | COG0556 | uvrB | UvrABC system protein B |
| T | Q8CS61 | COG0589 | uspA | Universal stress protein UspA |
| T | Q8CQK0 | COG0745 | walR | Transcriptional regulatory protein WalR |
| M | Q8CP74 | COG0744 | - | Penicillin-binding protein 2 |
| M | Q8CMV0 | COG0438 | gtf1 | Glycosyltransferase |
| O | P0C0N7 | COG0459 | groEL | 60 kDa chaperonin |
| O | Q8CTA6 | COG0396 | - | ABC transporter (ATP-binding protein) |
| C | Q8CN04 | COG1012 | rocA | 1-Pyrroline-5-carboxylate dehydrogenase |
| C | Q8CSL9 | COG0508 | odhB | Dihydrolipoyllysine-residue succinyltransferase |
| C | Q8CPH5 | COG0045 | sucC | Succinyl-CoA ligase (ADP-forming) subunit beta |
| C | Q8CNX4 | COG0538 | Icd | Isocitrate dehydrogenase (NADP) |
| C | Q8CP83 | COG0567 | odhA | 2-Oxoglutarate dehydrogenase E1 component |
| C | Q8CPL2 | COG1053 | sdhA | Succinate dehydrogenase flavoprotein subunit |
| C | Q8CQA3 | COG1249 | lpdA | Dihydrolipoyl dehydrogenase |
| C | Q8CT13 | COG0508 | odp2 | Dihydrolipoyllysine-residue acetyltransferase component of pyruvatedehydrogenase complex |
| C | Q8CNJ7 | COG0055 | atpD | ATP synthase subunit beta |
| C | Q8CS25 | COG1866 | pckA | Phosphoenolpyruvate carboxykinase (ATP) |
| C | Q8PCP2 | COG1048 | acnA | Aconitate hydratase1 |
| C | Q8CNI5 | COG1012 | - | Aldehyde dehydrogenase (NAD) family protein |
| C | Q8CNJ1 | COG0356 | atpA | ATP synthase subunit alpha |
| C | Q8CN24 | COG1012 | aldA | Aldehyde dehydrogenase (NAD) family protein |
| C | Q8CQB2 | COG0371 | gldA | Glycerol dehydrogenase |
| C | Q8CQA1 | COG0022 | - | Branched-chain alpha-keto acid dehydrogenase E1 |
| C | Q8CMZ0 | COG0039 | idh2 | L-Lactate dehydrogenase |
| G | Q8CPY5 | COG0057 | Gap | Glyceraldehyde 3-phosphate dehydrogenase, C-terminal domain protein |
| G | Q8CPY3 | COG0148 | Eno | Enolase |
| G | Q8CTD6 | COG0126 | Pgk | Phosphoglycerate kinase |
| G | Q8CS69 | COG0469 | Pyk | Pyruvate kinase |
| G | Q8CN17 | COG1869 | rbsD | D-ribose pyranase |
| G | Q8CRJ4 | COG1105 | lacC | Tagatose-6-phosphate kinase |
| G | Q8CN27 | COG3855 | Fbp | Fructose-1,6-bisphosphatase class 3 O |
| G | Q8CPC7 | COG0021 | Tkt | Transketolase |
| E | P0C0N1 | COG0078 | argF | Ornithine carbamoyltransferase |
| E | Q8CU41 | COG0078 | arcB | Ornithine carbamoyltransferase |
| E | Q8CSR8 | COG0174 | glnA | Glutamine synthetase |
| E | Q8CQG5 | COG2235 | arcA | Arginine deiminase |
| E | Q8CPU5 | COG0334 | gdhA | Glutamate dehydrogenase |
| E | Q8CSR8 | COG0174 | glnA | Glutamine synthetase |
| E | Q8CU41 | COG0078 | otcC1 | Ornithine carbamoyltransferase |
| E | Q8CMM1 | COG1003 | gcvPB | Glycine dehydrogenase subunit 2 |
| E | Q8CMM0 | COG0403 | gcvPA | Probable glycine dehydrogenase (decarboxylating) subunit 1 |
| E | Q8CP09 | COG0169 | aroE | AroE |
| E | Q8CPN0 | COG3842 | potA2 | ABC transporter, ATP-binding protein |
| E | Q8CU42 | COG0549 | arcC2 | Carbamate kinase |
| E | Q8CTA4 | COG0520 | Csd | Cysteine desulfurase |
| E | Q8CNQ9 | COG0834 | - | Glutamine ABC transporter, permease protein |
| F | Q8CPJ6 | COG0044 | pyrC | Dihydroorotase |
| F | Q8CMQ7 | COG0517 | guaB | Inosine-5’-monophosphate dehydrogenase |
| H | Q8CNU2 | COG0108 | ribBA | Riboflavin biosynthesis protein |
| H | Q8CQV7 | COG0214 | pdxS | Pyridoxal biosynthesis lyase |
| H | Q8CNB8 | COG1052 | pdxB | Putative 2-hydroxyacid dehydrogenase |
| H | Q8CNZ1 | COG0001 | hemL | Glutamate-1-semialdehyde 2,1-aminomutase |
| H | Q8CPQ6 | COG1169 | - | Isochorismate synthase |
| P | Q8CR71 | COG0855 | Ppk | Polyphosphate kinase |
| P | Q8CPN8 | COG1122 | - | ABC transporter, ATP-binding protein |
| P | Q8CPD0 | COG0753 | catA | Catalase |
| P | P0C0Q6 | COG0605 | soda | Superoxide dismutase |
| P | Q8CPD0 | COG0753 | katA | Catalase |
| P | Q8CN76 | COG0223 | narT | Nitrate ABC transporter substrate-binding protein |
| R | Q8CNT0 | COG1106 | - | Abortive phage resistance protein |
| R | Q8CQ56 | COG1064 | adhP | Acetaldehyde reductase |
| R | Q8CMY4 | COG0579 | mqo4 | Probable malate:quinone oxidoreductase 4 |
| R | Q8CTE4 | COG0457 | - | Uncharacterized protein |
| R | Q8CPB7 | COG1942 | - | 4-Oxalocrotonate tautomerase |
| S | Q8CT08 | COG4493 | - | UPF0637 protein HMPREF9956_0818 |
| - | Q8CMZ9 | nog70990 | isaA | Putative transglycosylase IsaA |
| - | Q8CMZ9 | NOG70990 | gseF | Transglycosylase-like domain protein |
| - | Q7CCK7 | - | - | Phenol soluble modulin beta 1/beta 2 |
| - | Q8CTA5 | - | sufD | FeS assembly protein SufD |
| - | Q8CQK8 | NOG27742 | - | Poly (glycerol-phosphate) alpha-glucosyltransferase |
| - | Q8CS14 | NOG28792 | - | Uncharacterized protein |
| - | Q8CTN6 | - | - | Uncharacterized protein |
| - | Q8CU14 | - | - | Putative uncharacterized protein |
| Class | Accession | COG | Gene Name | Function |
|---|---|---|---|---|
| J | Q8CS54 | COG0522 | rpsD | 30S ribosomal protein S4 |
| J | Q8CRG6 | COG0092 | rpsC | 30S ribosomal protein S3 |
| J | Q5HQE9 | COG1514 | - | 2’,5’ RNA ligase family protein |
| L | Q8CSH8 | COG0776 | - | Transcriptional regulator |
| M | Q8CP74 | COG0744 | - | Transglycosylase |
| U | Q8CPZ2 | COG0653 | secA1 | Protein translocase subunit |
| C | Q8CNJ3 | COG0711 | atpF | ATP synthase subunit b |
| C | Q8CT13 | COG0508 | pdhC | Dihydrolipoyllysine-residue acetyltransferase component of pyruvate dehydrogenase complex |
| C | Q8CPN3 | COG1071 | pdhA | Pyruvate dehydrogenase E1 component, alpha subunit |
| C | Q8CSL9 | COG0508 | odhB | Dihydrolipoyllysine-residue succinyltransferase, E2 component |
| G | Q8CPY5 | COG0057 | gapA1 | Glyceraldehyde-3-phosphate dehydrogenase, type I |
| E | Q8CSR8 | COG0174 | glnA | Putative uncharacterized protein |
| F | Q7CCJ0 | COG0503 | purR | Purine operon repressor |
| F | Q8CRN4 | COG0035 | Upp | Uracil phosphoribosyltransferase |
| F | Q8CPC9 | COG0516 | guaC | GMP reductase |
| F | Q5HL04 | COG1328 | nrdD | Anaerobic ribonucleoside-triphosphate reductase |
| H | Q8CRN9 | COG2145 | thiM | Hydroxyethylthiazole kinase |
| H | Q8CRM3 | COG5146 | coaW | Type II pantothenate kinase |
| P | Q5HM52 | COG4594 | fecB | Periplasmic-binding protein |
| P | Q8CTM7 | COG0025 | - | Putative Na+/H+ antiporter |
| R | Q8CN54 | COG1380 | lrgA | Antiholin-like protein LrgA |
| R | Q8CMN2 | COG3942 | sle1 | N-acetylmuramoyl-L-alanine amidase |
| S | Q8CRV3 | NOG42366 | ytxH | Uncharacterized protein |
| S | Q8CS40 | NOG41643 | - | Uncharacterized protein |
| S | Q5HMW0 | NOG08342 | - | Uncharacterized protein |
| S | Q5HM45 | NOG40448 | - | Putative uncharacterized protein |
| S | Q5HMK5 | NOG80633 | - | Uncharacterized protein |
| S | Q8CN89 | NOG131938 | - | Putative uncharacterized protein |
| S | Q5HMK1 | NOG249085 | - | Uncharacterized protein |
| - | Q5HMJ2 | - | - | Uncharacterized protein |
| - | Q5HMJ9 | - | - | Uncharacterized protein |
| - | Q5HMK0 | - | - | Uncharacterized protein |
| - | Q7CCK7 | - | - | Antibacterial protein 2 |
| - | J0ZSL7 | - | - | Siphovirus Gp157 |
| - | Q5HK77 | - | - | Methylase S |
| - | Q8CT20 | NOG44554 | - | Putative uncharacterized protein |
| - | Q8CRB3 | - | - | Membrane protein |
| - | Q8CTN6 | - | - | Uncharacterized protein |
| GO ID | GO Term | ATCC12228EV | 983EV |
|---|---|---|---|
| Number of Molecules | Number of Molecules | ||
| GO:0006412 | Translation | 28 | 10 |
| GO:0044267 | cellular protein metabolic process | 31 | 11 |
| GO:0006091 | generation of precursor metabolites | 13 | 6 |
| GO:0019538 | protein metabolic process | 31 | 11 |
| GO:0046365 | monosaccharide catabolic process | 10 | 4 |
| GO:0006006 | glucose metabolic process | 11 | 4 |
| GO:0006096 | Glycolysis | 8 | 4 |
| GO:0005996 | monosaccharide metabolic process | 12 | 4 |
| GO:0016052 | carbohydrate catabolic process | 12 | 4 |
| GO:0006007 | glucose catabolic process | 9 | 4 |
| GO:0019318 | hexose metabolic process | 11 | 4 |
| GO:0019320 | hexose catabolic process | 9 | 4 |
| GO:0034645 | cellular macromolecule biosynthetic process | 35 | 12 |
| GO:0008152 | metabolic process | 102 | 0 |
| GO:0009056 | catabolic process | 28 | 0 |
| GO:0006099 | tricarboxylic acid cycle | 6 | 0 |
| GO:0046356 | acetyl-CoA catabolic process | 6 | 0 |
| GO:0009059 | macromolecule biosynthetic process | 35 | 0 |
| GO:0009109 | coenzyme catabolic process | 6 | 0 |
| GO:0009405 | Pathogenesis | 0 | 3 |
| GO:0009987 | cellular process | 0 | 33 |
| GO:0010467 | gene expression | 0 | 12 |
| GO:0051704 | multi-organism process | 0 | 3 |
| GO:0044238 | primary metabolic process | 0 | 28 |
| GO:0009112 | nucleobase metabolic process | 0 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Chávez, F.; Cedillo-Peláez, C.; Zapi-Colín, L.A.; Gutiérrez-González, G.; Martínez-Torres, I.; Peralta, H.; Chavez-Galan, L.; Avila-Calderón, E.D.; Contreras-Rodríguez, A.; Bartolo-Aguilar, Y.; et al. The Extracellular Vesicles from the Commensal Staphylococcus Epidermidis ATCC12228 Strain Regulate Skin Inflammation in the Imiquimod-Induced Psoriasis Murine Model. Int. J. Mol. Sci. 2021, 22, 13029. https://doi.org/10.3390/ijms222313029
Gómez-Chávez F, Cedillo-Peláez C, Zapi-Colín LA, Gutiérrez-González G, Martínez-Torres I, Peralta H, Chavez-Galan L, Avila-Calderón ED, Contreras-Rodríguez A, Bartolo-Aguilar Y, et al. The Extracellular Vesicles from the Commensal Staphylococcus Epidermidis ATCC12228 Strain Regulate Skin Inflammation in the Imiquimod-Induced Psoriasis Murine Model. International Journal of Molecular Sciences. 2021; 22(23):13029. https://doi.org/10.3390/ijms222313029
Chicago/Turabian StyleGómez-Chávez, Fernando, Carlos Cedillo-Peláez, Luis A. Zapi-Colín, Guadalupe Gutiérrez-González, Isaí Martínez-Torres, Humberto Peralta, Leslie Chavez-Galan, Erick D. Avila-Calderón, Araceli Contreras-Rodríguez, Yaneth Bartolo-Aguilar, and et al. 2021. "The Extracellular Vesicles from the Commensal Staphylococcus Epidermidis ATCC12228 Strain Regulate Skin Inflammation in the Imiquimod-Induced Psoriasis Murine Model" International Journal of Molecular Sciences 22, no. 23: 13029. https://doi.org/10.3390/ijms222313029
APA StyleGómez-Chávez, F., Cedillo-Peláez, C., Zapi-Colín, L. A., Gutiérrez-González, G., Martínez-Torres, I., Peralta, H., Chavez-Galan, L., Avila-Calderón, E. D., Contreras-Rodríguez, A., Bartolo-Aguilar, Y., Rodríguez-Martínez, S., Cancino-Diaz, M. E., & Cancino-Diaz, J. C. (2021). The Extracellular Vesicles from the Commensal Staphylococcus Epidermidis ATCC12228 Strain Regulate Skin Inflammation in the Imiquimod-Induced Psoriasis Murine Model. International Journal of Molecular Sciences, 22(23), 13029. https://doi.org/10.3390/ijms222313029