
Quantitative Proteomics and Differential Protein Abundance Analysis after the Depletion of PEX3 from Human Cells Identifies Additional Aspects of Protein Targeting to the ER

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Results
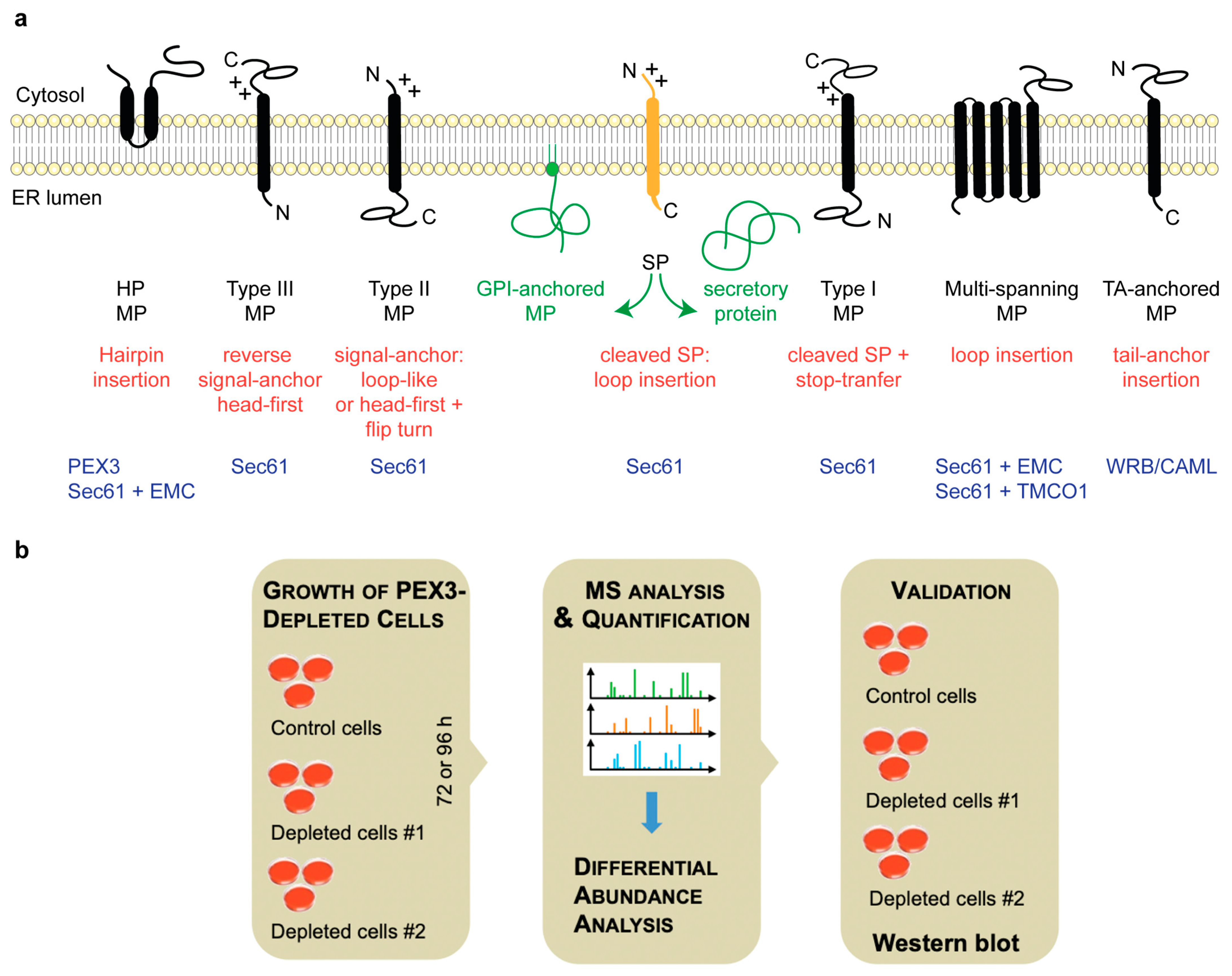
2.1. Quantitative Proteomic Analysis of HeLa Cells after the Transient and Partial Depletion of PEX3 by siRNA
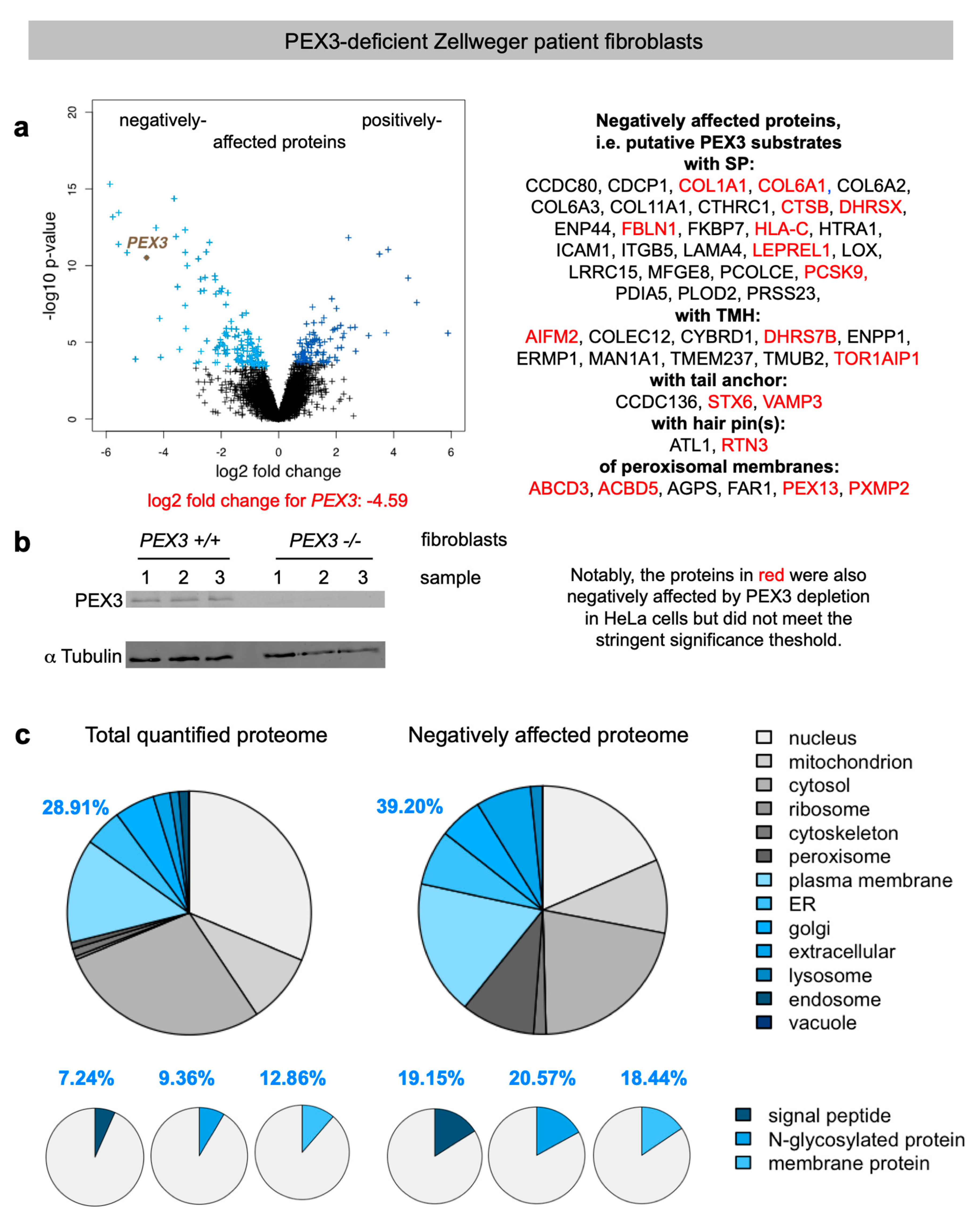
2.2. Quantitative Proteomic Analysis of PEX3-Deficient Zellweger Patient Fibroblasts
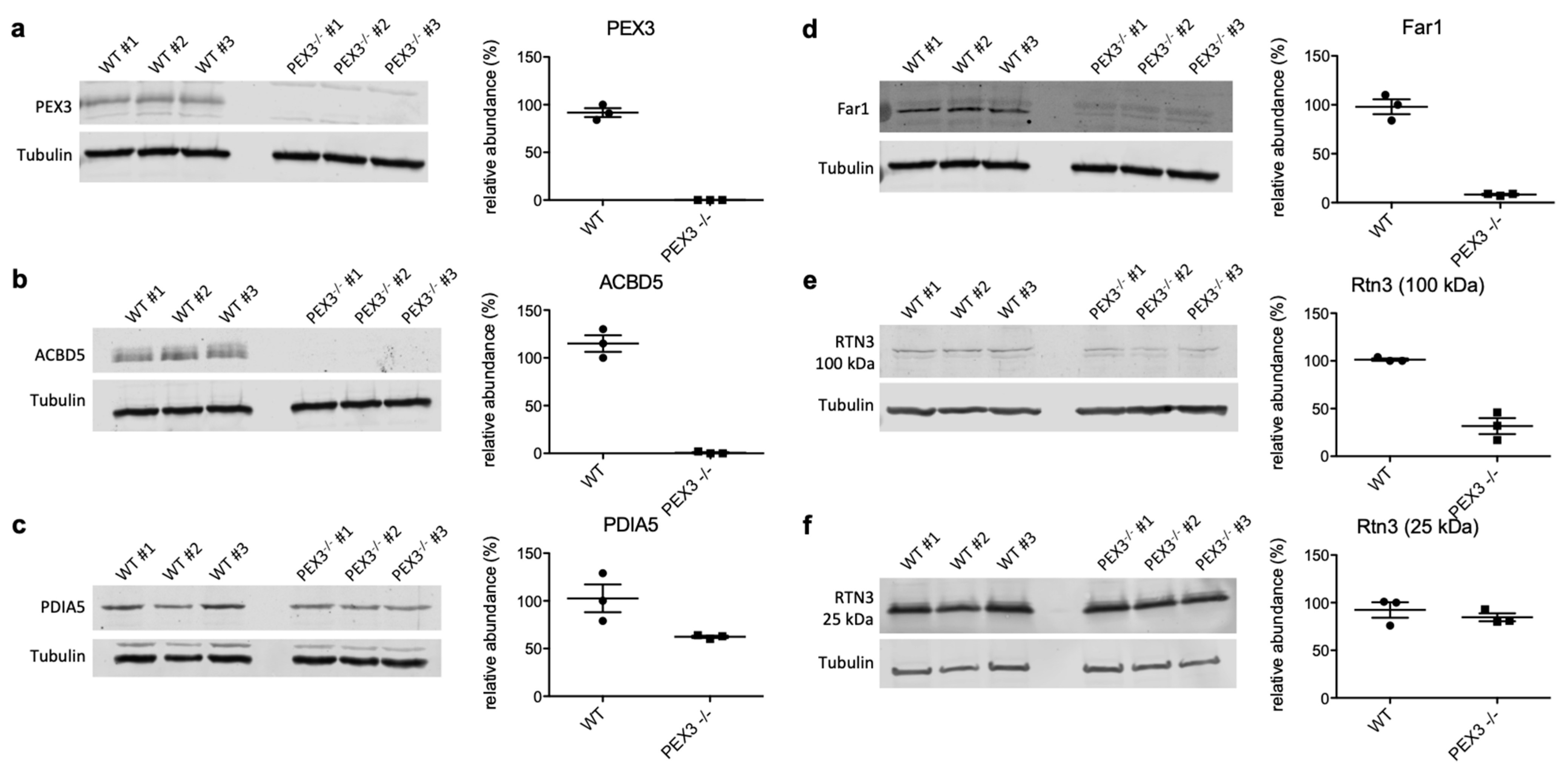
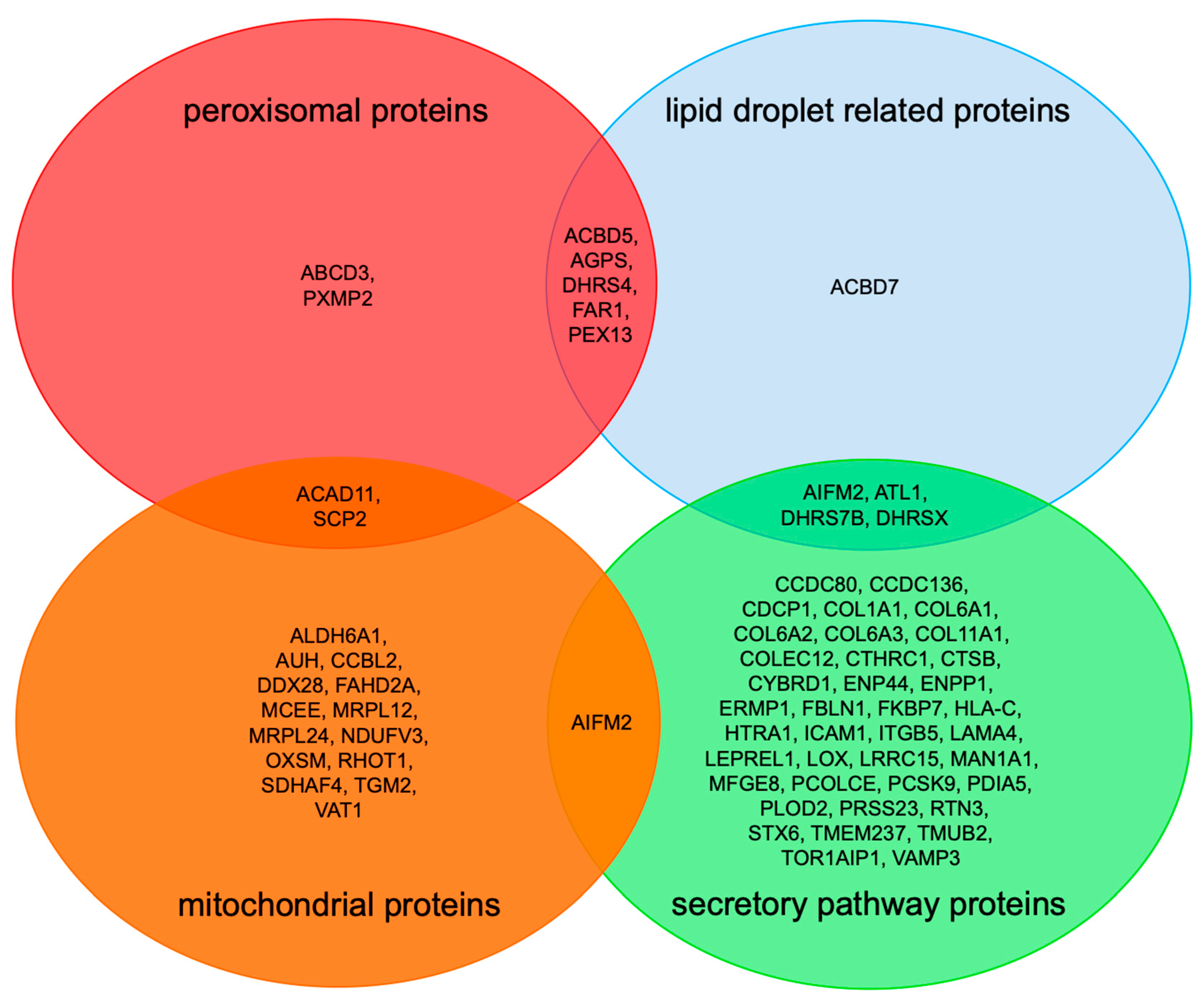
2.3. Negatively Affected Precursor Proteins in Zellweger Patient Fibroblasts Are Specific for PEX3- Deficiency, and Are Partially Affected in PEX3-Depleted HeLa Cells
3. Discussion
4. Materials and Methods
4.1. Cell Growth and Analysis
4.2. Label-Free Quantitative Proteomic Analysis
4.3. Data Analysis
4.4. Validation of Putative PEX3 Substrates by Quantitative Western Blotting
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ATL | Atlastin |
| CDG | Congenital disorder of glycosylation |
| EMC | ER membrane complex |
| ER | Endoplasmic reticulum |
| GET | Guided entry of tail-anchored proteins |
| GO | Gene ontology |
| GPI | Glycosylphosphatidylinositol |
| LD | Lipid droplet |
| PEX | Peroxin |
| RAMP | Ribosome-associated membrane protein |
| RNC | Ribosome-nascent chain complex |
| RTN | Reticulon |
| SEC | (Protein involved in) secretion |
| SND | SRP-independent |
| SP | Signal peptide |
| SR | SRP receptor |
| SRP | Signal recognition particle |
| SSR | Signal sequence receptor |
| TMEM | Transmembrane (protein) |
| TMH | Transmembrane helix |
| TRAM | translocating chain-associating membrane (protein) |
| TRAP | Translocon-associated protein |
| TRC | Transmembrane recognition complex |
| Z | Zellweger (patient fibroblasts) |
References
- Dudek, J.; Pfeffer, S.; Lee, P.-H.; Jung, M.; Cavalié, A.; Helms, V.; Förster, F.; Zimmermann, R. Protein transport into the human endoplasmic reticulum. J. Mol. Biol. 2015, 427, 1159–1175. [Google Scholar] [CrossRef]
- Aviram, N.; Schuldiner, M. Targeting and translocation of proteins to the endoplasmic reticulum at a glance. J. Cell Sci. 2017, 130, 4079–4085. [Google Scholar] [CrossRef] [PubMed]
- Gemmer, M.; Förster, F. A clearer picture of the ER translocon complex. J. Cell Sci. 2020, 133, jcs231340. [Google Scholar] [CrossRef] [PubMed]
- O´Keefe, S.; Pool, M.R.; High, S. Membrane protein biogenesis at the ER: The highways and byways. FEBS J. 2021. [Google Scholar] [CrossRef]
- Budnik, A.; Stephens, D.J. ER exit sites—Localization and control of COPII vesicle formation. FEBS Lett. 2009, 583, 3796–3803. [Google Scholar] [CrossRef] [PubMed]
- Stephens, D.J. Collagen secretion explained. Nature 2012, 482, 474–475. [Google Scholar] [CrossRef]
- Schrul, B.; Schliebs, W. Intracellular communication between lipid droplets and peroxisomes: The Janus face of PEX19. Biol. Chem. 2018, 399, 741–749. [Google Scholar] [CrossRef]
- Jansen, R.L.M.; van der Klei, I.J. The peroxisome biogenesis factors Pex3 and Pex19: Multitasking proteins with disputed functions. FEBS Lett. 2019, 593, 457–474. [Google Scholar] [CrossRef]
- Dhimann, R.; Caesar, S.; Thiam, A.R.; Schrul, B. Mechanisms of protein targeting to lipid droplets: A unified cell biological and biophysical perspective. Sem. Cell Dev. Biol. 2020, 108, 4–13. [Google Scholar] [CrossRef]
- Goodman, J.M. Building the lipid droplet assembly complex. J. Cell Biol. 2020, 219, e202006025. [Google Scholar] [CrossRef] [PubMed]
- Hansen, K.G.; Aviram, N.; Laborenz, J.; Bibi, C.; Meyer, M.; Spang, A.; Schuldiner, M.; Herrmann, J.M. An ER surface retrieval pathway safeguaerds the import of mitochondrial membrane proteins in yeast. Science 2018, 361, 1118–1122. [Google Scholar] [CrossRef]
- Hegde, R.S.; Bernstein, H. The surprising complexity of signal peptides. Trends Biochem. Sci. 2006, 31, 563–571. [Google Scholar] [CrossRef]
- Goder, V.; Spiess, M. Molecular mechanism of signal sequence orientation in the endoplasmic reticulum. EMBO J. 2003, 22, 3645–3653. [Google Scholar] [CrossRef] [PubMed]
- Goder, V.; Junne, T.; Spiess, M. Sec61p contributes to signal sequence orientation according to the positive-inside rule. Mol. Biol. Cell 2004, 15, 1470–1478. [Google Scholar] [CrossRef]
- Borgese, N.; Fasana, E. Targeting pathways of C-tail-anchored proteins. Biochim. Biophys. Acta 2011, 1808, 937–946. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Sakisaka, T. Molecular machinery for insertion of tail-anchored membrane proteins into the endoplasmic reticulum membrane in mammalian cells. Mol. Cell 2012, 48, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Borgese, N.; Coy-Vergara, J.; Colombo, S.F.; Schwappach, B. The ways of tails: The GET pathway and more. Proteins 2019, 38, 289–305. [Google Scholar] [CrossRef]
- Abell, B.M.; High, S.; Moloney, M.M. Membrane protein topology of oleosin is constrained by its long hydrophobic domain. J. Biol. Chem. 2002, 277, 8602–8610. [Google Scholar] [CrossRef]
- Leznicki, P.; Schneider, H.O.; Harvey, J.V.; Shi, W.Q.; High, S. Co-translational biogenesis of lipid droplet integral membrane proteins. J. Cell Sci. 2021, 132, jcs.259220. [Google Scholar] [CrossRef]
- Gamerdinger, M.; Hanebuth, M.A.; Frickey, T.; Deuerling, E. The principle of antagonism ensures protein targeting specificity at the endoplasmic reticulum. Science 2015, 348, 201–207. [Google Scholar] [CrossRef]
- Hsieh, H.-H.; Lee, J.H.; Chandrasekar, S.; Shan, S.-o. A ribosome-associated chaperone enables sustrate triage in a cotranslational protein targeting complex. Nat. Commun. 2020, 11, 5840. [Google Scholar] [CrossRef]
- Aviram, N.; Ast, T.; Costa, E.A.; Arakel, E.; Chuartzman, S.G.; Jan, C.H.; Haßdenteufel, S.; Dudek, J.; Jung, M.; Schorr, S.; et al. The SND proteins constitute an alternative targeting route to the endoplasmic reticulum. Nature 2016, 540, 134–138. [Google Scholar] [CrossRef]
- Casson, J.; McKenna, M.; Haßdenteufel, S.; Aviram, N.; Zimmermann, R.; High, S. Multiple pathways facilitate the biogenesis of mammalian tail-anchored proteins. J. Cell Sci. 2017, 130, 3851–3861. [Google Scholar] [CrossRef]
- Haßdenteufel, S.; Sicking, M.; Schorr, S.; Aviram, N.; Fecher-Trost, C.; Schuldiner, M.; Jung, M.; Zimmermann, R.; Lang, S. hSnd2 protein represents an alternative targeting factor to the endoplasmic reticulum in human cells. FEBS Lett. 2017, 591, 3211–3224. [Google Scholar] [CrossRef]
- Haßdenteufel, S.; Johnson, N.; Paton, A.W.; Paton, J.C.; High, S.; Zimmermann, R. Chaperone-mediated Sec61 channel gating during ER import of small precursor proteins overcomes Sec61 inhibitor-reinforced energy barrier. Cell Rep. 2018, 23, 1373–1386. [Google Scholar] [CrossRef]
- Haßdenteufel, S.; Nguyen, D.; Helms, V.; Lang, S.; Zimmermann, R. Components and mechanisms for ER import of small human presecretory proteins. FEBS Lett. 2019, 593, 2506–2524. [Google Scholar] [CrossRef]
- Cui, X.A.; Zhang, H.; Palazzo, A.F. p180 promotes the ribosome-independent localization of a subset of mRNA to the endoplasmic reticulum. PLoS Biol. 2012, 10, e1001336. [Google Scholar] [CrossRef]
- Cui, X.A.; Zhang, Y.; Hong, S.J.; Palazzo, A.F. Identification of a region within the placental alkaline phosphatase mRNA that mediates p180-dependent targeting to the endoplasmic reticulum. J. Biol. Chem. 2013, 288, 29633–29641. [Google Scholar] [CrossRef] [PubMed]
- Calvin, H.J.; Williams, C.C.; Weissman, J.S. Principles of ER coranslational translocation revealed by proximity-specific ribosome profiling. Science 2014, 346, 1257521. [Google Scholar] [CrossRef]
- Ueno, T.; Tanaka, K.; Kaneko, K.; Taga, Y.; Sata, T.; Irie, S.; Shunji Hattori, S.; Ogawa-Goto, K. Enhancement of procollagen biosynthesis by p180 through augmented ribosome association on the endoplasmic reticulum in response to stimulated secretion. J. Biol. Chem. 2010, 285, 29942–29950. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.C.-C.; Reid, D.W.; Hoffman, A.M.; Sarkar, D.; Nicchitta, C.V. Oncoprotein AEG-1 is an endoplasmic reticulum RNA-binding protein whose interactome is enriched in organelle resident protein-encoding mRNAs. RNA 2018, 24, 688–703. [Google Scholar] [CrossRef]
- Hannigan, M.M.; Hoffman, A.M.; Thompson, J.W.; Zheng, T.; Nicchitta, C.V. Quantitative proteomics links the LRRC59 interactome to mRNA translation on the ER membrane. Mol. Cell. Proteom. 2020, 19, 1826–1849. [Google Scholar] [CrossRef]
- Bhadra, P.; Schorr, S.; Lerner, M.; Nguyen, D.; Dudek, J.; Förster, F.; Helms, V.; Lang, S.; Zimmermann, R. Quantitative proteomics and differential protein abundance analysis after depletion of putative mRNA receptors in the ER membrane of human cells identifies novel aspects of mRNA targeting to the ER. Molecules 2021, 26, 3591. [Google Scholar] [CrossRef] [PubMed]
- Sicking, M.; Lang, S.; Bochen, F.; Drenth, J.P.H.; Zacharia, M.; Zimmermann, R.; Roos, A.; Linxweiler, M. Complexity and specificity of sec61-channelopathies: Human diseases affecting gating of the Sec61 complex. Cells 2021, 10, 1036. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Sakisaka, T. The peroxisome biogenesis factors posttranslationally target reticulon homology-domain containing proteins to the endoplasmic reticulum membrane. Sci. Rep. 2018, 8, 2322. [Google Scholar] [CrossRef] [PubMed]
- Schrul, B.; Kopito, R.R. Peroxin-dependent targeting of a lipid-droplet-destined membrane protein to ER subdomains. Nat. Cell Biol. 2016, 18, 740–751. [Google Scholar] [CrossRef] [PubMed]
- Dahan, N.; Francisco, T.; Falter, C.; Rodrigues, T.; Kalel, V.; Kunze, M.; Hansen, T.; Schliebs, W.; Erdmann, R. Current advances in the function and biogenesis of peroxisomes and their roles I health and disease. Hist. Cell Biol. 2021, 155, 513–524. [Google Scholar] [CrossRef]
- Erdmann, R.; Veenhuis, M.; Mertens, D.; Kunau, W.-H. Isolation of peroxisome-deficient mutants of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1989, 86, 5419–5423. [Google Scholar] [CrossRef]
- Götte, K.; Girzalsky, W.; Linkert, M.; Baumgart, E.; Kammerer, S.; Hunau, W.-H.; Erdmann, R. Pex19p, a farnesylated protein essential for peroxisome biogenesis. Mol. Cell. Biol. 1998, 18, 616–628. [Google Scholar] [CrossRef] [PubMed]
- Hettema, E.H.; Girzalsky, W.; van den Berg, M.; Erdmann, R.; Distel, B. Saccharomyces cerevisiae Pex3p and Pex19p are required for proper localization and stability of peroxisomal membrane proteins. EMBO J. 2000, 19, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, F.; Dietrich, D.; Eylenstein, R.; Groemping, Y.; Stehle, T.; Dodt, G. The role of conserved PEX3 regions in PEX19-binding and peroxisome biogenesis. Traffic 2012, 13, 1244–1260. [Google Scholar] [CrossRef]
- Muntau, A.C.; Maerhofer, P.U.; Paton, B.C.; Kammerer, S.; Roscher, A.A. Defective peroxisome membrane synthesis due to mutations in human PEX3 causes Zellweger syndrome, complementation group G. Am. J. Hum. Genet. 2000, 67, 967–975. [Google Scholar] [CrossRef]
- Shimozawa, N.; Suzuki, Y.; Zhang, Z.; Imamura, A.; Ghaedi, K.; Fujiki, Y.; Kondo, N. Identification of PEX3 as the gene mutated in a Zellweger syndrome patient lacking peroxisomal remnant structures. Hum. Mol. Genet. 2000, 9, 1995–1999. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ghaedi, K.; Honsho, M.; Shimozawa, N.; Suzuki, Y.; Kondo, N.; Fujiki, Y. PEX3 is the causal gene responsible for peroxisome membrane assembly-defective Zellweger syndrome of complementation group G. Am. J. Hum. Genet. 2000, 67, 976–981. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hein, M.Y.; Hubner, N.C.; Poser, I.; Cox, J.; Nagaraj, N.; Toyoda, Y.; Gak, I.A.; Weisswange, I.; Mansfeld, J.; Buchholz, F.; et al. A human interactome in three quantitative dimensions organized by stoichiometries and abundances. Cell 2015, 163, 712–723. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.; Stutz, R.; Schorr, S.; Lang, S.; Pfeffer, S.; Freeze, H.F.; Förster, F.; Helms, V.; Dudek, J.; Zimmermann, R. Proteomics reveals signal peptide features determining the client specificity in human TRAP-dependent ER protein import. Nat. Commun. 2018, 9, 37639. [Google Scholar] [CrossRef]
- Schorr, S.; Nguyen, D.; Haßdenteufel, S.; Nagaraj, N.; Cavalié, A.; Greiner, M.; Weissgerber, P.; Loi, M.; Paton, A.W.; Paton, J.C.; et al. Proteomics identifies signal peptide features determining the substrate specificity in human Sec62/Sec63-dependent ER protein import. FEBS J. 2020, 287, 4612–4640. [Google Scholar] [CrossRef]
- Klein, M.-C.; Lerner, M.; Nguyen, D.; Pfeffer, S.; Dudek, J.; Förster, F.; Helms, V.; Lang, S.; Zimmermann, R. TRAM1 protein may support ER protein import by modulating the phospholipid bilayer near the lateral gate of the Sec61 channel. Channels 2020, 14, 28–44. [Google Scholar] [CrossRef] [PubMed]
- Vizcaíno, J.A.; Csordas, A.; del-Toro, N.; Dianes, J.A.; Griss, J.; Lavidas, I.; Mayer, G.; Perez-Riverol, Y.; Reisinger, F.; Ternent, T.; et al. 2016 update of the PRIDE database and related tools. Nucleic Acids Res. 2016, 44, D447–D456. [Google Scholar] [CrossRef]
- Cichocki, B.A.; Krumpe, K.; Vitali, D.G.; Rapaport, D. Pex19 is involved in importing dually targeted tail-anchored proteins to both mitochondria and peroxisomes. Traffic 2018, 19, 770–785. [Google Scholar] [CrossRef]
- Exner, T.; Romero-Brey, I.; Yifrach, E.; Rivera-Monroy, J.; Schrul, B.; Zouboulis, C.C.; Stremmel, W.; Honsho, M.; Bartenschlager, R.; Zalckvar, E.; et al. An alternative membrane topology permits lipid droplet localization of peroxisomal fatty acyl-CoA reductase 1. J. Cell Sci. 2021, 132, jcs223016. [Google Scholar] [CrossRef]
- Agarwal, A.K.; Sukumaran, S.; Cortés, V.A.; Tunison, K.; Mizrachi, D.; Sankella, S.; Gerard, R.D.; Horton, J.D.; Garg, A. Human 1-acylglycerol-3-phosphate O-acyltransferase isoforms 1 and 2: Biochemical characterization and inability to rescue hepatic steatosis in Agpat2(-/-) gene lipodystrophic mice. J. Biol. Chem. 2011, 286, 37676–37691. [Google Scholar] [CrossRef]
- He, W.; Shi, Q.; Hu, X.; Yan, R. The membrane topology of RTN3 and its effect on binding of RTN3 to BACE1. J. Biol. Chem. 2007, 282, 29144–29151. [Google Scholar] [CrossRef]
- Mayerhofer, P.U.; Bano-Polo, M.; Mingarro, I.; Johnson, A.E. Human peroxin PEX3 is co-translationally integrated into the ER and exits the ER in budding vesicles. Traffic 2016, 17, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Brar, G.A.; Weissman, J.S. Ribosome profiling reveals the what, when, where, and how of protein synthesis. Nat. Rev. Mol. Cell Biol. 2015, 16, 651–664. [Google Scholar] [CrossRef]
- Pfeiffer, N.V.; Dirndorfer, D.; Lang, S.; Resenberger, U.K.; Restelli, L.M.; Hemion, C.; Miesbauer, M.; Frank, S.; Neutzner, A.; Zimmermann, R.; et al. Structural features within the nascent chain regulate alternative targeting of secretory proteins to mitochondria. EMBO J. 2013, 32, 1036–1051. [Google Scholar] [CrossRef] [PubMed]
- Nuebel, E.; Morgan, J.T.; Fogarty, S.; Winter, J.M.; Lettlova, S.; Berg, J.A.; Chen, Y.-C.; Kidwell, C.U.; Maschek, J.A.; Clowers, K.J.; et al. The biochemical basis of mitochondrial dysfunction in Zellweger spectrum disorder. EMBO Rep. 2021, 22, e51991. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, J.; Qin, X.; Wang, W. Peroxisomal membrane contact sites in mammalian cells. Front. Cell Dev. Biol. 2020, 8, 512. [Google Scholar] [CrossRef]
- Saito, K.M.; Chen, F.; Bard, S.; Chen, H.; Zhou, D.; Woodley, R.; Polischuk, R.; Schekman, R.; Malhotra, V. TANGO1 facilitates cargo loading at endoplasmic reticulum exit sites. Cell 2009, 136, 891–902. [Google Scholar] [CrossRef] [PubMed]
- Raote, I.; Ortega-Bellido, M.; Santos, A.J.M.; Foresti, O.; Zhang, C.; Garcia-Parajo, M.F.; Campelo, F.; Malhotra, V. TANGO1 builds a machine for collagen export by recruiting and spatially organizing COPII, thethers and membranes. eLife 2018, 7, e32723. [Google Scholar] [CrossRef]
- Raote, I.; Ernst, A.M.; Campelo, F.; Rothman, J.; Pincet, F.; Malhotra, V. TANGO1 membrane helices create a lipid diffusion barrier at curved membranes. eLife 2020, 9, e57822. [Google Scholar] [CrossRef]
- Pataki, C.I.; Rodrigues, J.; Zhang, L.; Qian, J.; Efron, B.; Hastie, T.; Elias, J.E.; Levitt, M.; Kopito, R.R. Proteomic analysis of monolayer-integrated proteins of lipid droplets identifies amphipathic interfacial α-helical membrane anchors. Proc. Natl. Acad. Sci. USA 2018, 115, E8172–E8180. [Google Scholar] [CrossRef]
- Pinto, M.P.; Grou, C.P.; Fransen, M.; Fransen, M.; Sa-Miranda, C. The cytosolic domain of PEX3, a protein inolved in the biogenesis of peroxisomes, binds membrane lipids. Biochem. Biophys. Acta 2009, 1793, 1669–1675. [Google Scholar] [CrossRef] [PubMed]
- Poulos, A.; Christodoulou, J.; Chow, C.W.; Goldblatt, J.; Paton, B.C.; Orii, T.; Suzuki, Y.; Shimozawa, N. Peroxisomal assembly defects: Clinical, pathologic, and biochemical findings in two patients in a newly identified complementation group. J. Pediatr. 1995, 127, 596–599. [Google Scholar] [CrossRef]
- Rappsilber, J.; Mann, M.; Ishihama, Y. Protocol for micro-purification, enrichment, pre-fractionation and storage of peptides for proteomics using StageTips. Nat. Protoc. 2007, 2, 1896–1906. [Google Scholar] [CrossRef]
- Nagaraj, N.; Kulak, N.A.; Cox, J.; Neuhauser, N.; Mayr, K.; Hoerning, O.; Vorm, O.; Mann, M. System-wide perturbation analysis with nearly complete coverage of the yeast proteome by single-shot ultra HPLC runs on a bench top Orbitrap. Mol. Cell. Proteom. 2012, 11, M111.013722. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell. Proteom. 2014, 13, 2513–2526. [Google Scholar] [CrossRef]
- Nagaraj, N.; Wisniewski, J.R.; Geiger, T.; Cox, J.; Kircher, M.; Kelso, J.; Pääbo, S.; Mann, M. Deep proteome and transcriptome mapping of a human cancer cell line. Mol. Sys. Biol. 2011, 7, 548. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Coc, J. The Perseus computational platform for comprehensive analysis of (proteo)omics data. Nature Meth. 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Hyunsoo, K.; Golub, G.H.; Park, H. Missing value estimation for DNA microarray gene expression data: Local least squares imputation. Bioinformatics 2005, 21, 187–198. [Google Scholar] [CrossRef]
- Tusher, V.G.; Tibshirani, R.; Chu, G. Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. USA 2001, 98, 5116–5121. [Google Scholar] [CrossRef] [PubMed]
- Eden, E.; Navon, R.; Steinfeld, I.; Lipson, D.; Yakhini, Z. Gorilla: A tool for discovery and visualization of enriched GO terms in ranked gene lists. BMC Bioinform. 2009, 10, 48. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component/Subunit for ER Targeting | Abundance 1 | Localization 2 | Linked Diseases |
|---|---|---|---|
| #p34 (LRC59, LRRC59) 3 | 2480 | ERM | |
| #p180 (RRBP1) | 135 | ERM | Hepatocellular Carcinoma, Colorectal Cancer |
| Kinectin 1 (KTN1) | 263 | ERM | |
| AEG-1 (LYRIC, MTDH) | 575 | ERM | |
| #SRP4 | C | ||
| − SRP72 | 355 | Aplasia, Myelodysplasia | |
| − SRP68 | 197 | ||
| − SRP54 | 228 | Neutropenia, Pancreas Insufficiency | |
| − SRP19 | 33 | ||
| − SRP14 | 4295 | ||
| − SRP9 | 3436 | ||
| − 7SL RNA | |||
| SRP receptor | ERM | ||
| − SRα (docking protein) | 249 | ||
| − SRβ | 173 | ||
| Calmodulin | 9428 | C | |
| for ER targeting and, possibly, for membrane integration | |||
| hSnd1 | unknown | ||
| Snd receptor | ERM | ||
| − hSnd2 (TMEM208) | 81 | ||
| − hSnd3 § | 49 | ||
| PEX19 | 80 | C | Zellweger Syndrome |
| PEX3 | 103 | ERM, PexM | Zellweger Syndrome |
| for ER targeting plus membrane integration | |||
| #Bag6 complex | C | ||
| − TRC35 (Get4) | 171 | ||
| − Ubl4A | 177 | ||
| − Bag6 (Bat3) | 133 | ||
| SGTA | 549 | C | |
| TRC40 (Asna1, Get3) | 381 | C | |
| TA receptor | ERM | ||
| − CAML (CAMLG, Get2) | 5 | ||
| − WRB (CHD5, Get1) | 4 | Congenital Heart Disease | |
| for ER membrane integration | |||
| ERM protein complex | ERM | ||
| − EMC1 | 124 | ||
| − EMC2 | 300 | ||
| − EMC3 | 270 | ||
| − EMC4 | 70 | ||
| − EMC5 (MMGT1) | 35 | ||
| − EMC6 (TMEM93) | 5 | ||
| − EMC7 | 247 | ||
| − EMC8 | 209 | ||
| − EMC9 | 1 | ||
| − EMC10 | 3 | ||
| #TMCO1 complex § | ERM | Glaucoma, Cerebrofaciothoracic Dysplasia | |
| − TMCO1 | 2013 | ||
| − Nicalin | 99 | ||
| − TMEM147 | 21 | ||
| − CCDC47 (Calumin) | 193 | ||
| − NOMO | 267 | ||
| PAT complex | ERM | ||
| − PAT10 (Asterix) | |||
| − CCDC47 (Calumin) | 193 | ||
| for ER membrane integration plus translocation | |||
| #Sec61 complex§ | ERM | ||
| − Sec61α1 | 139 | Diabetes 5, CVID 6, TKD, Neutropenia | |
| − Sec61β | 456 | PLD, Colorectal cancer | |
| − Sec61γ | 400 | GBM, Hepatocellular carcinoma | |
| #Sec62 (TLOC1) | 26 | ERM | Breast-, Prostate-, Cervix-, Lung-cancer |
| ER Chaperones | |||
| − Sec63 (ERj2) | 168 | ERM | PLD, Colorectal cancer |
| − #ERj1 (DNAJC1) | 8 | ERM | |
| − BiP (Grp78, HSPA5) | 8253 | ERL | HUS |
| − Grp170 (HYOU1) | 923 | ERL | |
| − Sil1 (BAP) | 149 | ERL | MSS |
| #Calnexinpalmitoylated | 7278 | ERM | |
| #TRAM1 | 26 | ERM | |
| TRAM2 | 40 | ERM | |
| #TRAP complex | ERM | ||
| − TRAPα ((SSR1) | 568 | ||
| − TRAPβ (SSR2) | |||
| − TRAPγ (SSR3) | 1701 | CDG, Hepatocellular Carcinoma | |
| − TRAPδ (SSR4) | 3212 | CDG | |
| #RAMP4 (SERP1) | ERM | ||
| for covalent modification | |||
| #Oligosaccharyltransferase (OST-A) | ERM | ||
| − RibophorinI (Rpn1) | 1956 | ||
| − RibophorinII (Rpn2) | 527 | ||
| − OST48 | 273 | CDG | |
| − Dad1 | 464 | ||
| − OST4 | |||
| − TMEM258 | |||
| − Stt3A * | 430 | CDG | |
| − DC2 | |||
| − Kcp2 | |||
| Oligosaccharyltransferase (OST-B) | |||
| − RibophorinI (Rpn1) | 1956 | ||
| − RibophorinII (Rpn2) | 527 | ||
| − OST48 | 273 | CDG | |
| − Dad1 | 464 | ||
| − OST4 | |||
| − TMEM258 | |||
| − Stt3B * | 150 | CDG | |
| − TUSC3 | CDG | ||
| − MagT1 | 33 | ||
| Signal peptidase (SPC-A) | ERM | ||
| − SPC12 | 2733 | ||
| − SPC18 * (SEC11A) | |||
| − SPC22/23 | 334 | ||
| − SPC25 | 94 | ||
| Signal peptidase (SPC-C) | ERM | ||
| − SPC12 | 2733 | ||
| − SPC21 * (SEC11C) | |||
| − SPC22/23 | 334 | ||
| − SPC25 | 94 | ||
| GPI transamidase (GPI-T) | ERM | ||
| − GPAA1 | 9 | ||
| − PIG-K | 38 | ||
| − PIG-S | 86 | ||
| − PIG-T | 20 | ||
| − PIG-U | 42 |
| Proteins | PEX3 | Z 1 | RRBP1 2 | KTN1 2 | SEC61 2 | TRAP 2 | CDG 1,2 |
|---|---|---|---|---|---|---|---|
| Quantified proteins | 8178 | 6328 | 4813 | 4947 | 7212 | 7670 | 5920 |
| Statistically analyzed proteins | 6488 | 6328 | 4813 | 4947 | 5129 | 5911 | 5920 |
| representing the secretory pathway (%) | 29 | 29 | 26 | 27 | 26 | 27 | 36 |
| Proteins with SP (%) | 7 | 7 | 6 | 6 | 6 | 7 | nd 3 |
| N-Glycoproteins (%) | 9 | 9 | 8 | 8 | 8 | 8 | nd |
| Membrane proteins (%) | 13 | 13 | 12 | 13 | 12 | 13 | nd |
| Positively affected proteins | 0 | 97 | 157 | 25 | 342 | 77 | 39 |
| Negatively affected proteins | 13 | 141 | 141 | 45 | 482 | 180 | 279 |
| representing the secretory pathway (%) | 54 | 39 | 37 | 41 | 61 | 40 | 36 |
| Negatively affected proteins with SP (%) | 8 | 19 | 18 | 7 | 41 | 22 | 12 |
| Negatively affected N-glycoproteins (%) | 8 | 21 | 17 | 18 | 45 | 23 | 17 |
| Negatively affected membrane proteins (%) | 31 | 18 | 18 | 22 | 36 | 26 | 23 |
| Negatively affected proteins with SP | 1 | 27 | 21 | 3 | 197 | 38 | 34 |
| including N-glycoproteins | 0 | 23 | 16 | 3 | 158 | 28 | 30 |
| corresponding to % | 0 | 85 | 76 | 100 | 80 | 74 | 88 |
| including membrane proteins | 1 | 6 | 6 | 1 | 77 | 19 | 16 |
| corresponding to % | 100 | 22 | 29 | 33 | 39 | 50 | 53 |
| Negatively affected proteins with TMH | 3 | 16 | 18 | 8 | 98 | 22 | 41 |
| including N-glycoproteins | 1 | 6 | 7 | 4 | 56 | 11 | 17 |
| corresponding to % | 33 | 38 | 39 | 50 | 57 | 50 | 41 |
| Negatively affected peroxisomal proteins | 1 | 12 | 0 | 1 | 1 | 0 | 1 |
| corresponding to % | 8 | 9 | nd | 2 | 0 | nd | 0 |
| including membrane proteins | 1 | 6 | nd | 0 | 1 | 0 | 1 |
| corresponding to % | 100 | 50 | nd | nd | 100 | nd | 100 |
| Negatively affected mitochondrial proteins | 0 | 14 | 6 | 1 | 29 | 14 | 21 |
| corresponding to % | nd | 10 | 4 | 2 | 1 | 1 | 1 |
| including membrane proteins | nd | 4 | 3 | 0 | 11 | 3 | 8 |
| corresponding to % | nd | 29 | 50 | nd | 38 | 21 | 38 |
| Gene | Subcellular Location | Membrane Protein Type | SS or TMH |
|---|---|---|---|
| ACBD5 | Peroxisome membrane | Single-spanning membrane protein | |
| COLEC12 | Membrane | Single-spanning type II membrane protein | TMH |
| LRRC15 | Membrane | Single-spanning type I membrane protein | SP |
| PEX3 | Peroxisome membrane | Single-spanning membrane protein | |
| TOR1AIP1 | Nuclear envelope inner membrane | Single-spanning membrane protein | TMH |
| COL1A1 | Secreted, Extracellular space, Extracellular matrix | SP | |
| AGPS | Peroxisome membrane | ||
| ACAD11 | Peroxisome, Mitochondrion | ||
| STX6 | Golgi apparatus membrane | Tail-anchored membrane protein | tail anchor |
| CCDC136 | Acrosome membrane, Secretory vesicle, Cytoplasmic vesicle | Tail-anchored membrane protein | tail anchor |
| FAR1 | Peroxisome membrane | Tail-anchored membrane protein | tail anchor |
| PXMP2 | Peroxisome membrane | Multi-spanning membrane protein | |
| ATL1 | Cell projection, Golgi apparatus membrane, ER membrane, | Hairpin membrane protein with one HP | hairpin |
| COL6A2 | Extracellular matrix, Membrane, Secreted, Extracellular space | SP | |
| LOX | Extracellular space, Secreted | SP | |
| ERMP1 | ER membrane | Multi-spanning membrane protein | TMH |
| CYBRD1 | Membrane | Multi-spanning membrane protein | TMH |
| TMUB2 | Membrane | Multi-spanning membrane protein | TMH |
| ABCD3 | Peroxisome membrane | Multi-spanning membrane protein | |
| SCP2 | Peroxisome, Mitochondrion, Cytoplasm | ||
| CDCP1 | Secreted, Cell membrane | SP | |
| COL6A3 | Extracellular space, Secreted, Extracellular matrix | SP | |
| TMEM237 | Cell projection, Membrane, Cilium | Multi-spanning membrane protein | TMH |
| ENPP4 | Cell membrane | Single-spanning type I membrane protein | SP |
| HTRA1 | Cell membrane, Secreted, Cytoplasm, Cytosol | SP | |
| VAMP3 | Synapse, Membrane, Cell junction, Synaptosome | Tail-anchored membrane protein | tail anchor |
| MFGE8 | Membrane, Secreted | SP | |
| PRSS23 | Secreted | SP | |
| DHRS4 | Peroxisome, Nucleus | ||
| ITGB5 | Membrane | Single-pass type I membrane protein | SP |
| FBLN1 | Extracellular space, Secreted, Extracellular matrix | SP | |
| COL6A1 | Extracellular space, Secreted, Extracellular matrix | SP | |
| PCSK9 | Endosome, Golgi apparatus, Cell surface, Secreted, ER, Lysosome | SP | |
| CTHRC1 | Extracellular space, Secreted, Extracellular matrix | SP | |
| DHRSX | Secreted | SP | |
| HLA-C | Membrane | Single-spanning type I membrane protein | SP |
| CCDC80 | Secreted, Extracellular space, Extracellular matrix | SP | |
| RTN3 | ER membrane, Golgi apparatus membrane | Hairpin membrane protein with two HP | hairpin |
| ENPP1 | Secreted, Basolateral cell membrane, Cell membrane | Single-spanning type II membrane protein | TMH |
| PLOD2 | Rough ER membrane | SP | |
| RHOT1 | Mitochondrion outer membrane | ||
| COL11A1 | Extracellular matrix, Extracellular space, Secreted | SP | |
| NDUFV3 | Mitochondrion inner membrane | ||
| PCOLCE | Secreted | SP | |
| AIFM2 | Membrane, Mitochondrion outer membrane, Lipid droplet | Single-spanning membrane protein | TMH |
| MAN1A1 | Golgi apparatus membrane | Single-spanning type II membrane protein | TMH |
| ACBD7 | Cytosol | ||
| ICAM1 | Membrane | Single-spanning type I membrane protein | SP |
| CTSB | Lysosome, Melanosome, Secreted, Extracellular space | SP | |
| DHRS7B | ER membrane | Single-spanning type II membrane protein | TMH |
| LAMA4 | Extracellular matrix, Extracellular space, Secreted | SP | |
| LEPREL1 | ER, Golgi apparatus | SP | |
| PEX13 | Peroxisome membrane | Single-spanning membrane protein | |
| PDIA5 | ER lumen | SP | |
| CTHRC1 | Extracellular space, Secreted, Extracellular matrix | ||
| FKBP7 | ER lumen | SP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zimmermann, R.; Lang, S.; Lerner, M.; Förster, F.; Nguyen, D.; Helms, V.; Schrul, B. Quantitative Proteomics and Differential Protein Abundance Analysis after the Depletion of PEX3 from Human Cells Identifies Additional Aspects of Protein Targeting to the ER. Int. J. Mol. Sci. 2021, 22, 13028. https://doi.org/10.3390/ijms222313028
Zimmermann R, Lang S, Lerner M, Förster F, Nguyen D, Helms V, Schrul B. Quantitative Proteomics and Differential Protein Abundance Analysis after the Depletion of PEX3 from Human Cells Identifies Additional Aspects of Protein Targeting to the ER. International Journal of Molecular Sciences. 2021; 22(23):13028. https://doi.org/10.3390/ijms222313028
Chicago/Turabian StyleZimmermann, Richard, Sven Lang, Monika Lerner, Friedrich Förster, Duy Nguyen, Volkhard Helms, and Bianca Schrul. 2021. "Quantitative Proteomics and Differential Protein Abundance Analysis after the Depletion of PEX3 from Human Cells Identifies Additional Aspects of Protein Targeting to the ER" International Journal of Molecular Sciences 22, no. 23: 13028. https://doi.org/10.3390/ijms222313028
APA StyleZimmermann, R., Lang, S., Lerner, M., Förster, F., Nguyen, D., Helms, V., & Schrul, B. (2021). Quantitative Proteomics and Differential Protein Abundance Analysis after the Depletion of PEX3 from Human Cells Identifies Additional Aspects of Protein Targeting to the ER. International Journal of Molecular Sciences, 22(23), 13028. https://doi.org/10.3390/ijms222313028