
Astrocytes Exhibit a Protective Role in Neuronal Firing Patterns under Chemically Induced Seizures in Neuron–Astrocyte Co-Cultures

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
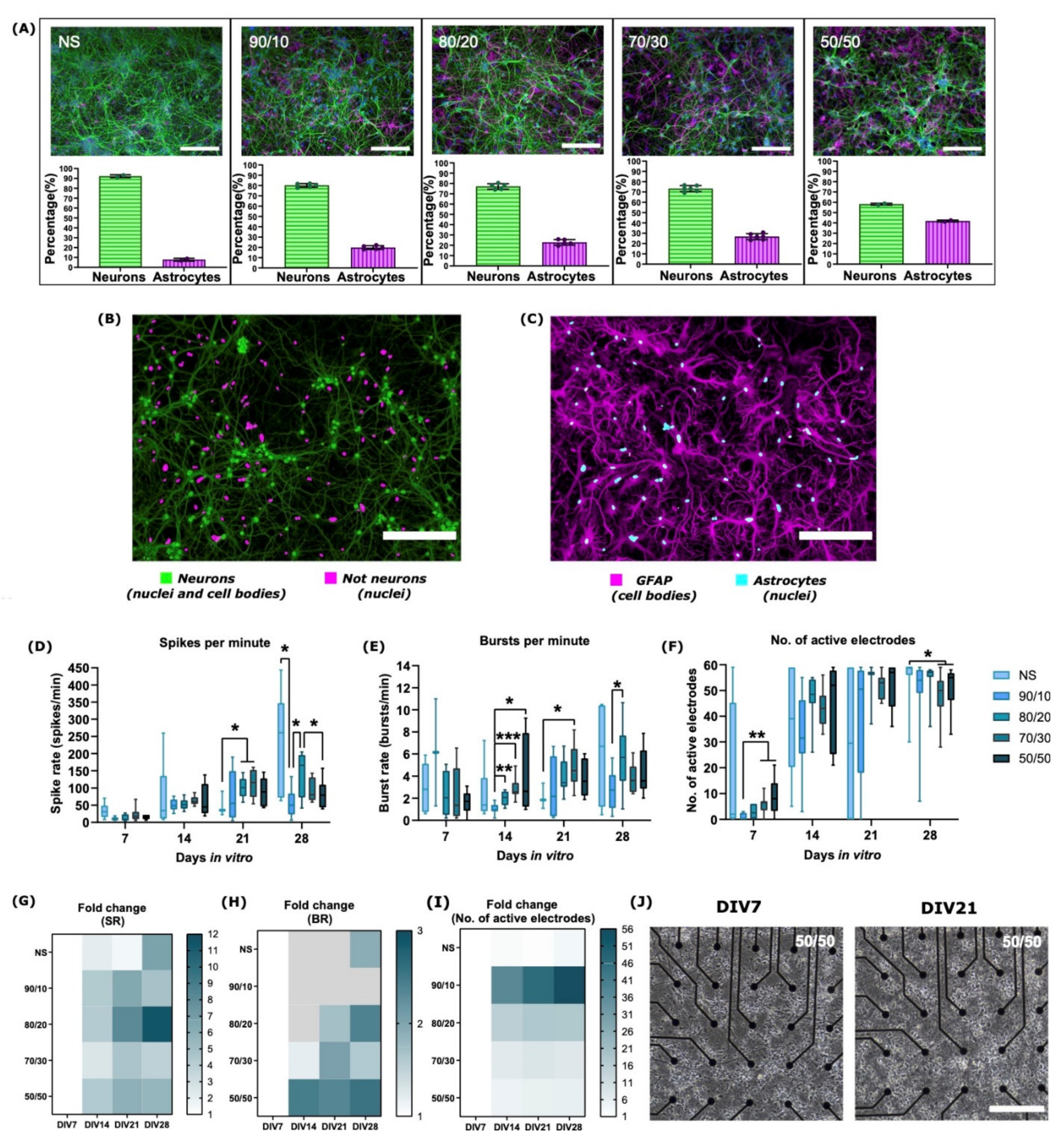
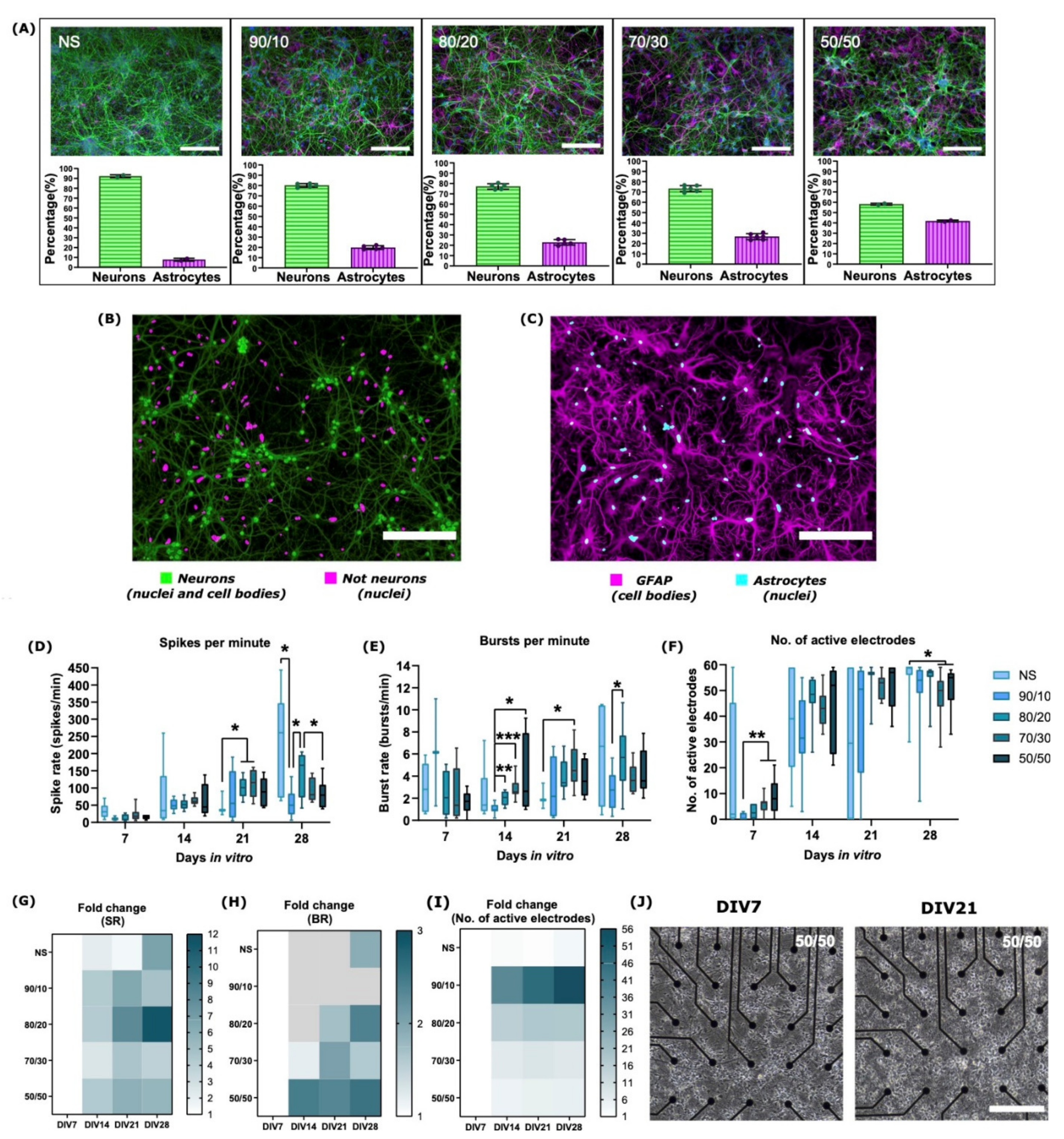
2.1. Neuron–Astrocyte Culture Ratios Corresponded Well to the Cell Plating Ratios
2.2. The Development of Neuronal Baseline Activity in NS vs. Neuron–Astrocyte Co-Cultures
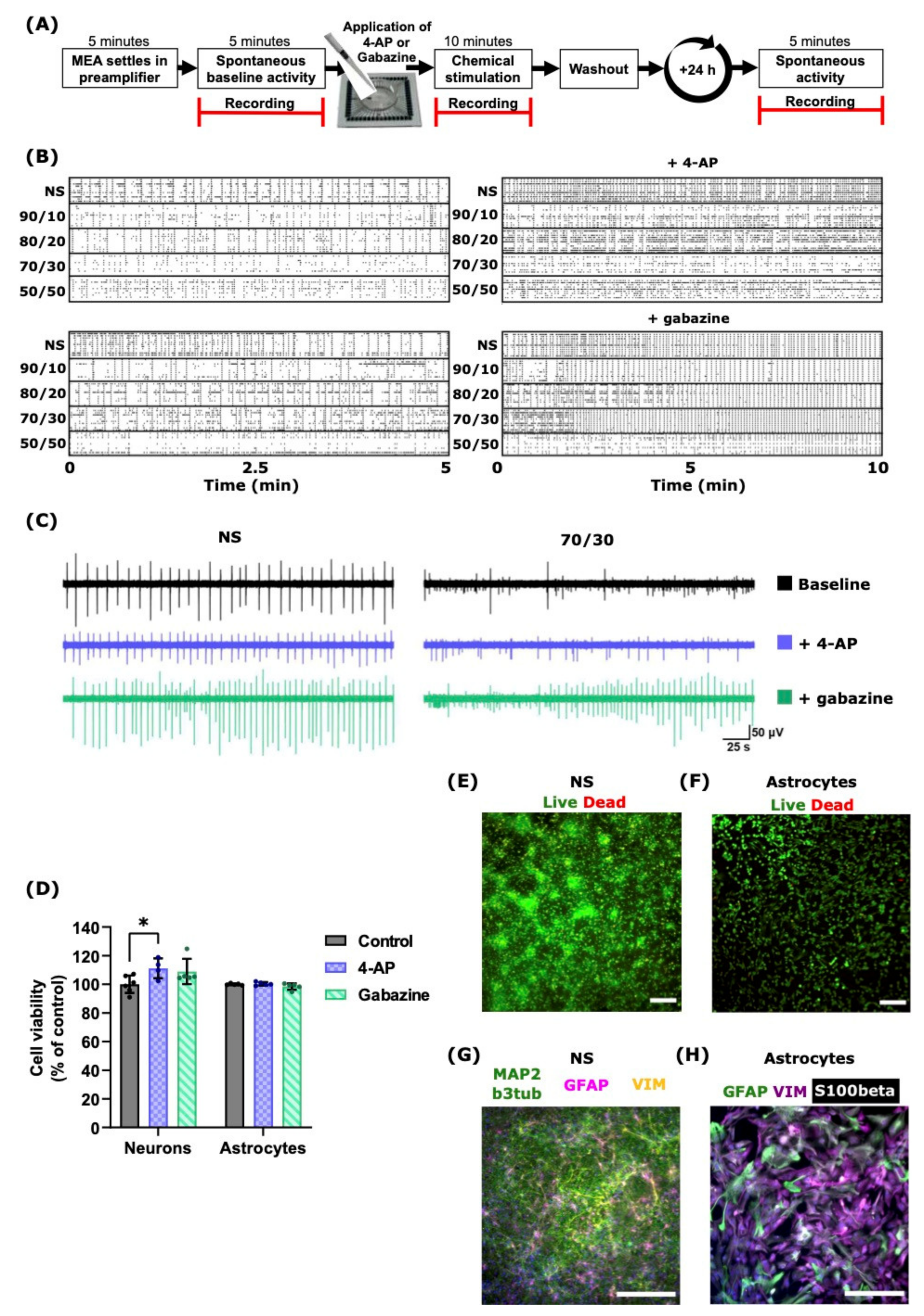
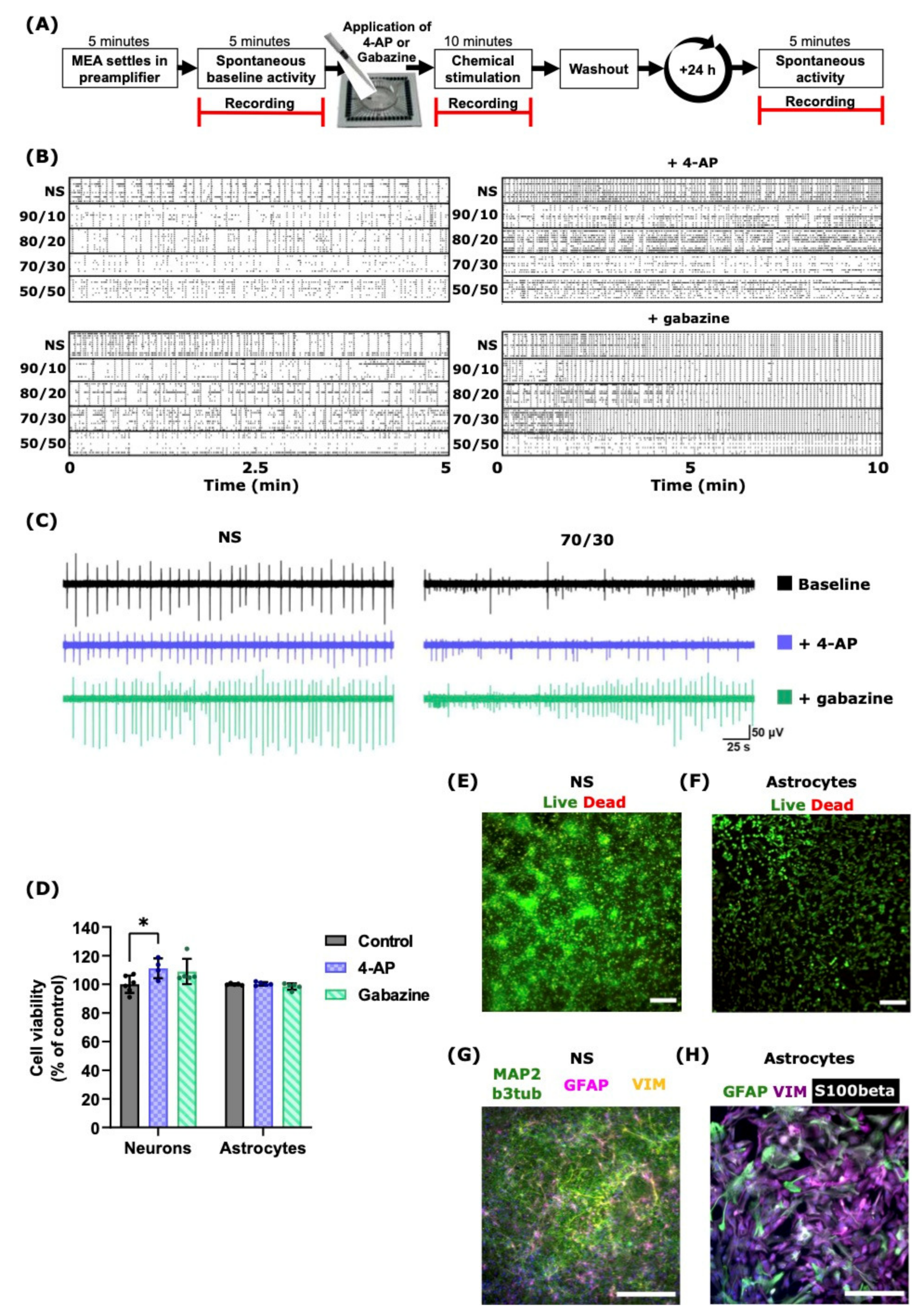
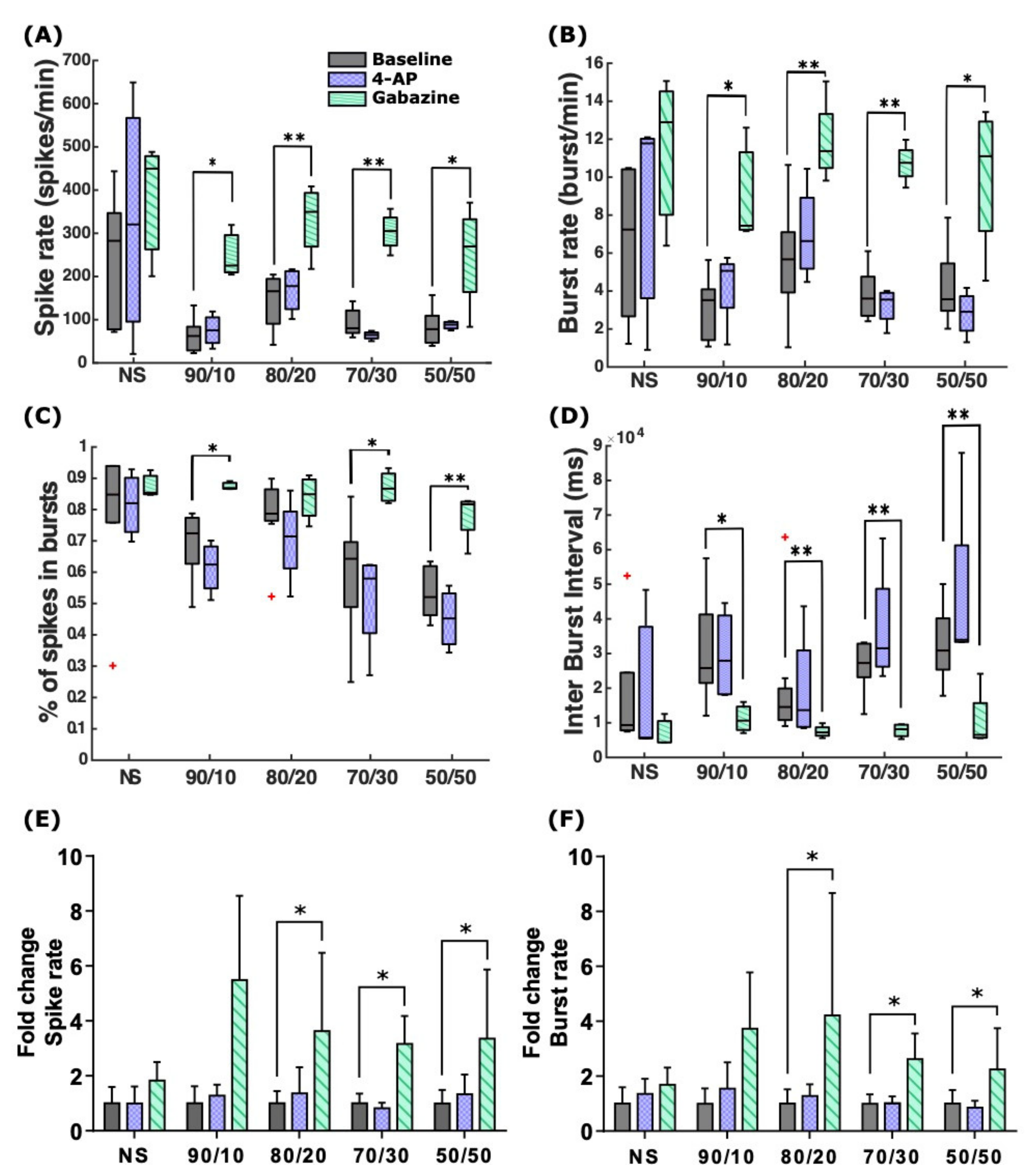
2.3. 4-AP and Gabazine Affected Neuronal Activity but Did Not Decrease Cell Viability
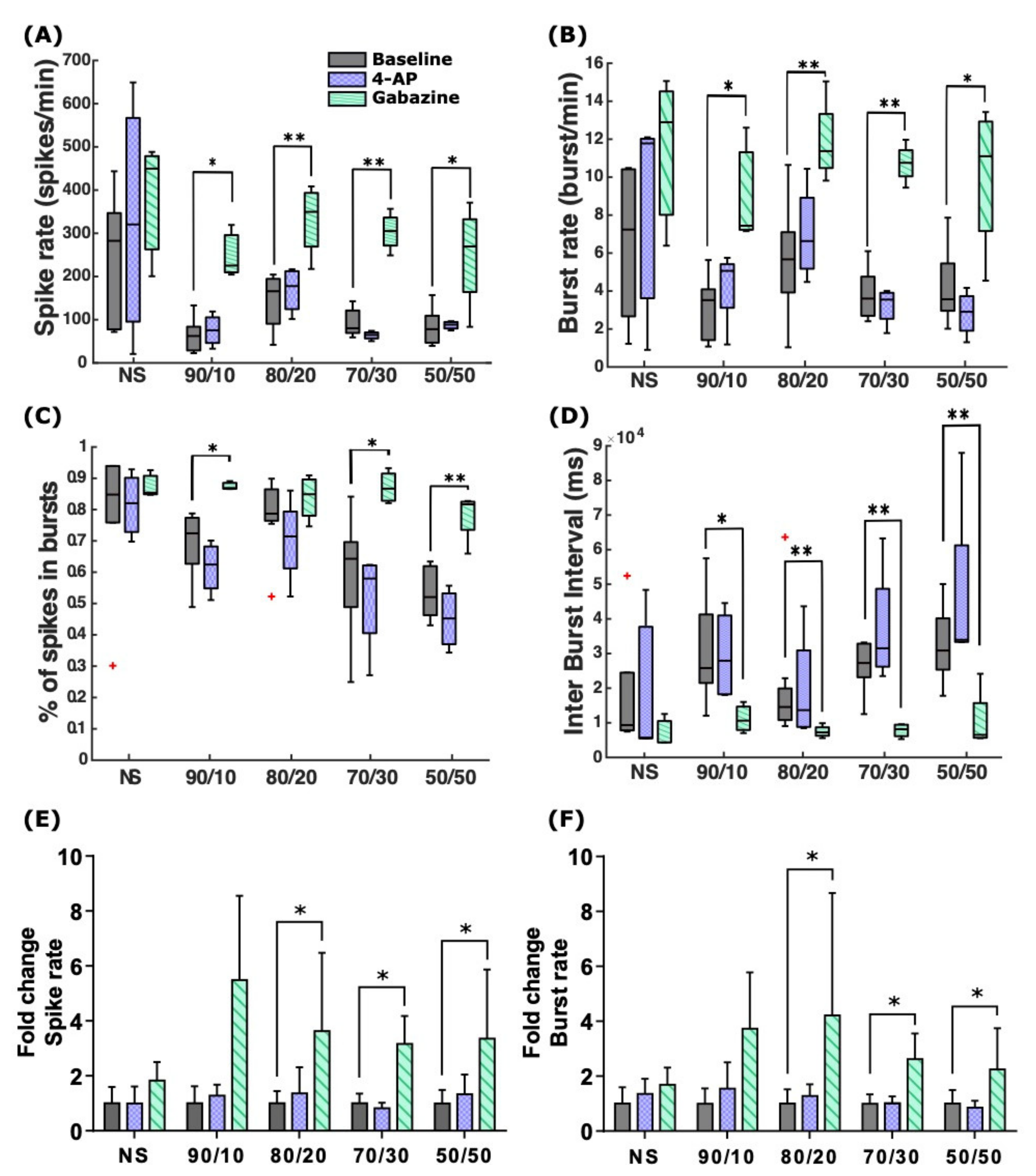
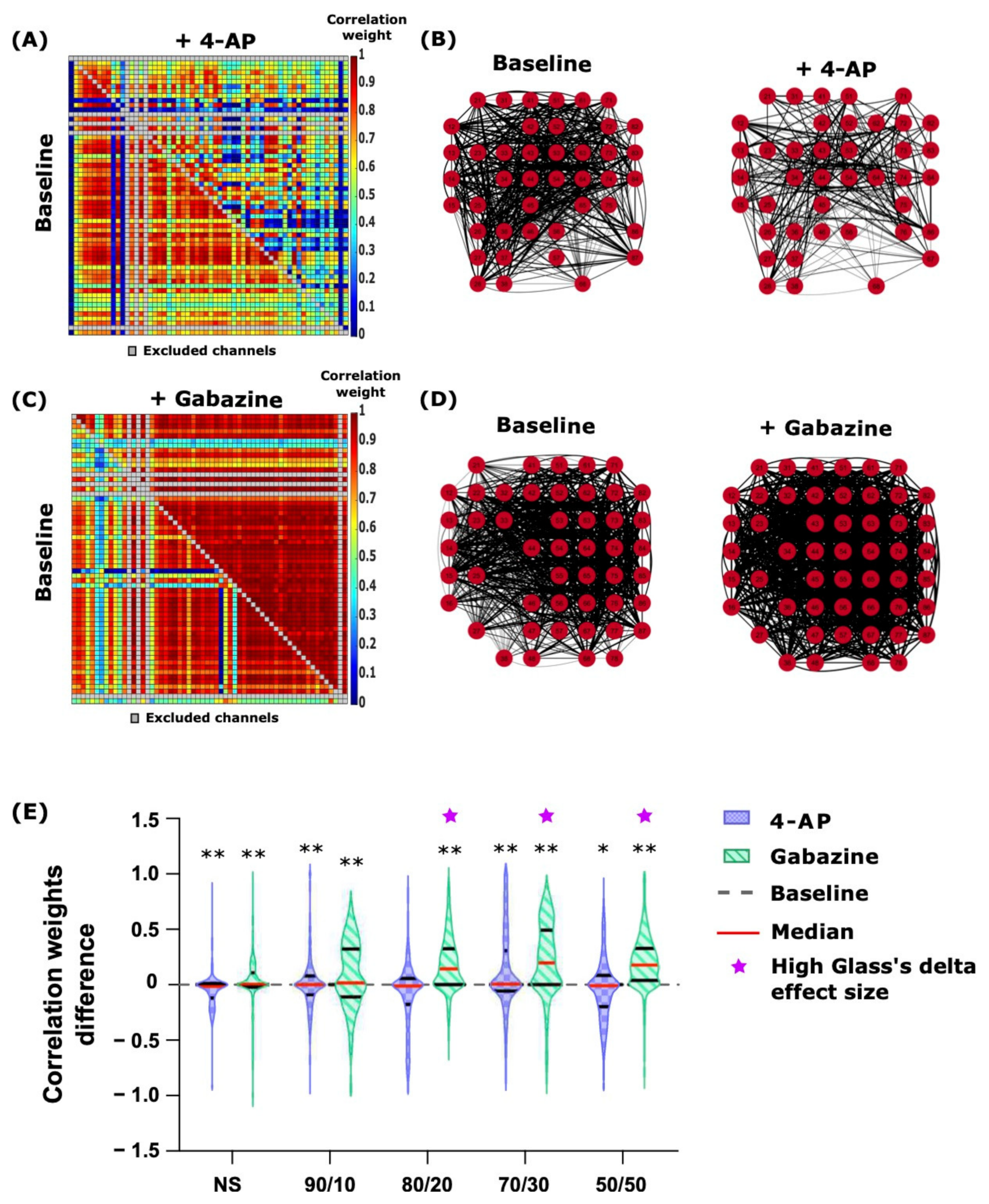
2.4. 4-AP and Gabazine Had Different Effects on Neuronal Network Synchronization
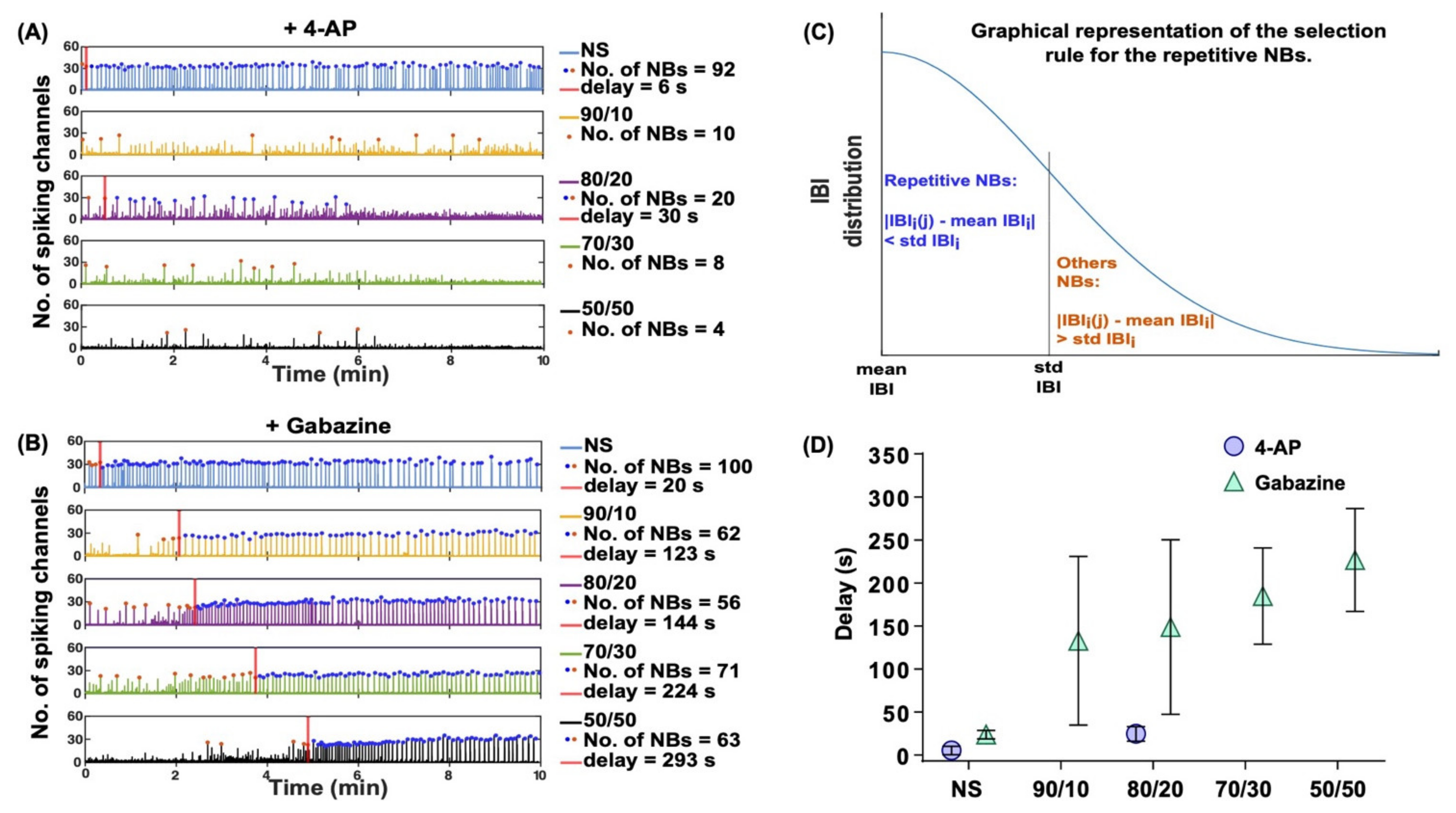
2.5. Gabazine Induced Synchronization and Neuron–Astrocyte Ratio Effects on Gabazine’s Delay in Response Time
3. Discussion
4. Materials and Methods
4.1. MEA and Coverslip Preparation
4.2. Cell Culture
4.3. Immunofluorescence and Imaging
4.4. Neuron and Astrocyte Viability
4.5. Chemical Stimulation
4.6. MEA Recordings
4.7. Data Analysis
4.7.1. Cell Count
4.7.2. Spiking and Bursting Analysis
4.7.3. Network Synchronicity by Channel Inter-Correlation Analysis
4.7.4. Network Synchronicity by Binned Activity Analysis
4.7.5. Delay in the Response Times of the Convulsants
4.7.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takeuchi, H.; Suzumura, A. Gap junctions and hemichannels composed of connexins: Potential therapeutic targets for neurodegenerative diseases. Front. Cell. Neurosci. 2014, 8, 189. [Google Scholar] [CrossRef] [Green Version]
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Tripartite synapses: Glia, the unacknowledged partner. Trends Neurosci. 1999, 22, 208–215. [Google Scholar] [CrossRef]
- Mayorquin, L.C.; Rodriguez, A.V.; Sutachan, J.-J.; Albarracín, S.L. Connexin-Mediated Functional and Metabolic Coupling Between Astrocytes and Neurons. Front. Mol. Neurosci. 2018, 11, 118. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Nedergaard, M. Physiology of Astroglia. Physiol. Rev. 2018, 98, 239–389. [Google Scholar] [CrossRef]
- Bazargani, N.; Attwell, D. Astrocyte calcium signaling: The third wave. Nat. Neurosci. 2016, 19, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Fraser, D.D.; Mudrick-Donnon, L.A.; MacVicar, B. Astrocytic GABA receptors. Glia 1994, 11, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Boddum, K.; Jensen, T.; Magloire, V.; Kristiansen, U.; Rusakov, D.A.; Pavlov, I.; Walker, M.C. Astrocytic GABA transporter activity modulates excitatory neurotransmission. Nat. Commun. 2016, 7, 13572. [Google Scholar] [CrossRef]
- Meier, S.D.; Kafitz, K.W.; Rose, C.R. Developmental profile and mechanisms of GABA-induced calcium signaling in hippocampal astrocytes. Glia 2008, 56, 1127–1137. [Google Scholar] [CrossRef]
- Doengi, M.; Hirnet, D.; Coulon, P.; Pape, H.-C.; Deitmer, J.W.; Lohr, C. GABA uptake-dependent Ca2+ signaling in developing olfactory bulb astrocytes. Proc. Natl. Acad. Sci. USA 2009, 106, 17570–17575. [Google Scholar] [CrossRef] [Green Version]
- Mederos, S.; González-Arias, C.; Perea, G. Astrocyte–Neuron Networks: A Multilane Highway of Signaling for Homeostatic Brain Function. Front. Synaptic Neurosci. 2018, 10, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Meur, K.; Mendizabal-Zubiaga, J.; Grandes, P.; Audinat, E. GABA release by hippocampal astrocytes. Front. Comput. Neurosci. 2012, 6, 59. [Google Scholar] [CrossRef] [Green Version]
- Rose, C.R.; Felix, L.; Zeug, A.; Dietrich, D.; Reiner, A.; Henneberger, C. Astroglial Glutamate Signaling and Uptake in the Hippocampus. Front. Mol. Neurosci. 2018, 10, 451. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Yoon, B.-E.; Berglund, K.; Oh, S.-J.; Park, H.; Shin, H.-S.; Augustine, G.J.; Lee, C.J. Channel-Mediated Tonic GABA Release from Glia. Science 2010, 330, 790–796. [Google Scholar] [CrossRef]
- Dossi, E.; Vasile, F.; Rouach, N. Human astrocytes in the diseased brain. Brain Res. Bull. 2018, 136, 139–156. [Google Scholar] [CrossRef]
- Bedner, P.; Steinhäuser, C. Altered Kir and gap junction channels in temporal lobe epilepsy. Neurochem. Int. 2013, 63, 682–687. [Google Scholar] [CrossRef] [PubMed]
- Steinhäuser, C.; Grunnet, M.; Carmignoto, G. Crucial role of astrocytes in temporal lobe epilepsy. Neuroscience 2016, 323, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Walrave, L.; Vinken, M.; Leybaert, L.; Smolders, I. Astrocytic Connexin43 Channels as Candidate Targets in Epilepsy Treatment. Biomolecules 2020, 10, 1578. [Google Scholar] [CrossRef] [PubMed]
- Shneker, B.F.; Fountain, N.B. Epilepsy. Dis. Mon. 2003, 49, 426–478. [Google Scholar] [CrossRef]
- Bhalla, D.; Godet, B.; Druet-Cabanac, M.; Preux, P.-M. Etiologies of epilepsy: A comprehensive review. Expert Rev. Neurother. 2011, 11, 861–876. [Google Scholar] [CrossRef]
- Ono, T.; Galanopoulou, A.S. Epilepsy and Epileptic Syndrome. In Neurodegenerative Diseases; Ahmad, S.I., Ed.; Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2012; Volume 724, pp. 99–113. ISBN 978-1-4614-0652-5. [Google Scholar]
- Blank, L.J.; Acton, E.K.; Thibault, D.; Willis, A.W. Neurodegenerative disease is associated with increased incidence of epilepsy: A population based study of older adults. Age Ageing 2021, 50, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Timmermann, A.; Henning, L.; Müller, H.; Steinhäuser, C.; Bedner, P. Astrocytic GABA Accumulation in Experimental Temporal Lobe Epilepsy. Front. Neurol. 2020, 11, 614923. [Google Scholar] [CrossRef]
- Tanskanen, J.M.; Ahtiainen, A.; Hyttinen, J.A. Toward Closed-Loop Electrical Stimulation of Neuronal Systems: A Review. Bioelectricity 2020, 2, 328–347. [Google Scholar] [CrossRef]
- Keller, J.M.; Frega, M. Past, Present, and Future of Neuronal Models In Vitro. In In Vitro Neuronal Networks; Chiappalone, M., Pasquale, V., Frega, M., Eds.; Advances in Neurobiology; Springer International Publishing: Cham, Switzerland, 2019; Volume 22, pp. 3–17. ISBN 978-3-030-11134-2. [Google Scholar]
- Thomas, C.A., Jr.; Springer, P.A.; Loeb, G.E.; Berwald-Netter, Y.; Okun, L.M. A miniature microelectrode array to monitor the bioelectric activity of cultured cells. Exp. Cell Res. 1972, 74, 61–66. [Google Scholar] [CrossRef]
- Ylä-Outinen, L.; Tanskanen, J.M.A.; Kapucu, F.E.; Hyysalo, A.; Hyttinen, J.A.K.; Narkilahti, S. Advances in Human Stem Cell-Derived Neuronal Cell Culturing and Analysis. In In Vitro Neuronal Networks; Chiappalone, M., Pasquale, V., Frega, M., Eds.; Advances in Neurobiology; Springer International Publishing: Cham, Switzerland, 2019; Volume 22, pp. 299–329. [Google Scholar] [CrossRef]
- Herculano-Houzel, S. The glia/neuron ratio: How it varies uniformly across brain structures and species and what that means for brain physiology and evolution. Glia 2014, 62, 1377–1391. [Google Scholar] [CrossRef] [PubMed]
- Von Bartheld, C.S.; Bahney, J.; Herculano-Houzel, S. The search for true numbers of neurons and glial cells in the human brain: A review of 150 years of cell counting: Quantifying Neurons and Glia in Human Brain. J. Comp. Neurol. 2016, 524, 3865–3895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enright, H.A.; Lam, D.; Sebastian, A.; Sales, A.P.; Cadena, J.; Hum, N.R.; Osburn, J.J.; Peters, S.K.G.; Petkus, B.; Soscia, D.A.; et al. Functional and transcriptional characterization of complex neuronal co-cultures. Sci. Rep. 2020, 10, 11007. [Google Scholar] [CrossRef] [PubMed]
- Odawara, A.; Matsuda, N.; Ishibashi, Y.; Yokoi, R.; Suzuki, I. Toxicological evaluation of convulsant and anticonvulsant drugs in human induced pluripotent stem cell-derived cortical neuronal networks using an MEA system. Sci. Rep. 2018, 8, 10416. [Google Scholar] [CrossRef]
- Aebersold, M.J.; Thompson-Steckel, G.; Joutang, A.; Schneider, M.; Burchert, C.; Forró, C.; Weydert, S.; Han, H.; Vörös, J. Simple and Inexpensive Paper-Based Astrocyte Co-culture to Improve Survival of Low-Density Neuronal Networks. Front. Neurosci. 2018, 12, 94. [Google Scholar] [CrossRef] [PubMed]
- Myers, T.L.; González, O.; Stein, J.B.; Bazhenov, M. Characterizing Concentration-Dependent Neural Dynamics of 4-Aminopyridine-Induced Epileptiform Activity. Epilepsy J. 2018, 4, 128. [Google Scholar] [CrossRef]
- Žiburkus, J.; Cressman, J.R.; Schiff, S.J. Seizures as imbalanced up states: Excitatory and inhibitory conductances during seizure-like events. J. Neurophysiol. 2013, 109, 1296–1306. [Google Scholar] [CrossRef] [Green Version]
- Amakhin, D.; Soboleva, E.; Ergina, J.L.; Malkin, S.; Chizhov, A.; Zaitsev, A.V. Seizure-Induced Potentiation of AMPA Receptor-Mediated Synaptic Transmission in the Entorhinal Cortex. Front. Cell. Neurosci. 2018, 12, 486. [Google Scholar] [CrossRef] [PubMed]
- Kasatkina, L.A. 4-Aminopyridine sequesters intracellular Ca2+ which triggers exocytosis in excitable and non-excitable cells. Sci. Rep. 2016, 6, 34749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hongo, Y.; Takasu, K.; Ikegaya, Y.; Hasegawa, M.; Sakaguchi, G.; Ogawa, K. Heterogeneous effects of antiepileptic drugs in anin vitroepilepsy model—A functional multineuron calcium imaging study. Eur. J. Neurosci. 2015, 42, 1818–1829. [Google Scholar] [CrossRef] [PubMed]
- Pacico, N.; Mingorance-Le Meur, A. New In Vitro Phenotypic Assay for Epilepsy: Fluorescent Measurement of Synchronized Neuronal Calcium Oscillations. PLoS ONE 2014, 9, e84755. [Google Scholar] [CrossRef]
- Wu, Z.-Z.; Li, D.-P.; Chen, S.-R.; Pan, H.-L. Aminopyridines Potentiate Synaptic and Neuromuscular Transmission by Targeting the Voltage-activated Calcium Channel β Subunit. J. Biol. Chem. 2009, 284, 36453–36461. [Google Scholar] [CrossRef] [Green Version]
- Hyvärinen, T.; Hyysalo, A.; Kapucu, F.E.; Aarnos, L.; Vinogradov, A.; Eglen, S.J.; Ylä-Outinen, L.; Narkilahti, S. Functional characterization of human pluripotent stem cell-derived cortical networks differentiated on laminin-521 substrate: Comparison to rat cortical cultures. Sci. Rep. 2019, 9, 17125. [Google Scholar] [CrossRef] [Green Version]
- Shmoel, N.; Rabieh, N.; Ojovan, S.M.; Erez, H.; Maydan, E.; Spira, M.E. Multisite electrophysiological recordings by self-assembled loose-patch-like junctions between cultured hippocampal neurons and mushroom-shaped microelectrodes. Sci. Rep. 2016, 6, 27110. [Google Scholar] [CrossRef] [Green Version]
- Behrens, M.M.; Ali, S.S.; Dao, D.N.; Lucero, J.; Shekhtman, G.; Quick, K.L.; Dugan, L.L. Ketamine-Induced Loss of Phenotype of Fast-Spiking Interneurons Is Mediated by NADPH-Oxidase. Science 2007, 318, 1645–1647. [Google Scholar] [CrossRef] [Green Version]
- Ueno, S.; Bracamontes, J.; Zorumski, C.; Weiss, D.S.; Steinbach, J.H. Bicuculline and Gabazine Are Allosteric Inhibitors of Channel Opening of the GABAAReceptor. J. Neurosci. 1997, 17, 625–634. [Google Scholar] [CrossRef] [Green Version]
- Lee, V.; Maguire, J. The impact of tonic GABAA receptor-mediated inhibition on neuronal excitability varies across brain region and cell type. Front. Neural Circuits 2014, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Farhy-Tselnicker, I.; Allen, N.J. Astrocytes, neurons, synapses: A tripartite view on cortical circuit development. Neural Dev. 2018, 13, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grainger, A.I.; King, M.C.; Nagel, D.A.; Parri, H.R.; Coleman, M.D.; Hill, E. In vitro Models for Seizure-Liability Testing Using Induced Pluripotent Stem Cells. Front. Neurosci. 2018, 12, 590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokal, D.M.; Mason, R.; Parker, T.L. Multi-neuronal recordings reveal a differential effect of thapsigargin on bicuculline- or gabazine-induced epileptiform excitability in rat hippocampal neuronal networks. Neuropharmacology 2000, 39, 2408–2417. [Google Scholar] [CrossRef]
- Bradley, J.A.; Luithardt, H.H.; Metea, M.R.; Strock, C.J. In Vitro Screening for Seizure Liability Using Microelectrode Array Technology. Toxicol. Sci. 2018, 163, 240–253. [Google Scholar] [CrossRef]
- Young, S.Z.; Platel, J.-C.; Nielsen, J.V.; Jensen, N.A.; Bordey, A. GABAA increases calcium in subventricular zone astrocyte-like cells through L- and T-type voltage-gated calcium channels. Front. Cell. Neurosci. 2010, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Halassa, M.M.; Fellin, T.; Haydon, P.G. The tripartite synapse: Roles for gliotransmission in health and disease. Trends Mol. Med. 2007, 13, 54–63. [Google Scholar] [CrossRef]
- Turovskaya, M.V.; Epifanova, E.A.; Tarabykin, V.S.; Babaev, A.A.; Turovsky, E.A. Interleukin-10 restores glutamate receptor-mediated Ca2+-signaling in brain circuits under loss of Sip1 transcription factor. Int. J. Neurosci. 2020, 1–12. [Google Scholar] [CrossRef]
- Turovsky, E.A.; Varlamova, E.G.; Plotnikov, E.Y. Mechanisms Underlying the Protective Effect of the Peroxiredoxin-6 Are Mediated via the Protection of Astrocytes during Ischemia/Reoxygenation. Int. J. Mol. Sci. 2021, 22, 8805. [Google Scholar] [CrossRef]
- Dietrich, M.; Hartung, H.-P.; Albrecht, P. Neuroprotective Properties of 4-Aminopyridine. Neurol. Neuroimmunol. Neuroinflamm. 2021, 8, e976. [Google Scholar] [CrossRef]
- Hu, D.; Liu, J.; Keblesh, J.; Xiong, H. Involvement of the 4-aminopyridine-sensitive transient A-type K+current in macrophage-induced neuronal injury. Eur. J. Neurosci. 2010, 31, 214–222. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.J.; Tauskela, J.S.; Stone, T.W.; Smith, R.A. Preconditioning with 4-aminopyridine protects cerebellar granule neurons against excitotoxicity. Brain Res. 2009, 1294, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Liu, Z.; Liu, Z.-Q.; Zeng, X.-M.; Chen, X.-H.; Zhang, Z.-H.; Mei, Y.-A. 4-Aminopyridine, a Kv channel antagonist, prevents apoptosis of rat cerebellar granule neurons. Neuropharmacology 2006, 51, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.-M.; Sun, L.-N.; Zhou, H.-Y.; Wang, X.-L. Voltage-dependent potassium channels are involved in glutamate-induced apoptosis of rat hippocampal neurons. Neurosci. Lett. 2006, 398, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Tukker, A.M.; Van Kleef, R.G.D.M.; Wijnolts, F.M.J.; De Groot, A.; Westerink, R.H.S. Towards animal-free neurotoxicity screening: Applicability of hiPSC-derived neuronal models for in vitro seizure liability assessment. ALTEX 2020, 37, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Genocchi, B.; Lenk, K.; Hyttinen, J. Influence of Astrocytic Gap Junction Coupling on in Silico Neuronal Network Activity. In XV Mediterranean Conference on Medical and Biological Engineering and Computing—MEDICON 2019; Henriques, J., Neves, N., de Carvalho, P., Eds.; IFMBE Proceedings; Springer International Publishing: Cham, Switzerland, 2020; Volume 76, pp. 480–487. ISBN 978-3-030-31634-1. [Google Scholar]
- Vincent, K.; Tauskela, J.S.; Mealing, G.A.; Thivierge, J.-P. Altered Network Communication Following a Neuroprotective Drug Treatment. PLoS ONE 2013, 8, e54478. [Google Scholar] [CrossRef] [Green Version]
- Murakami, S.; Kurachi, Y. Mechanisms of astrocytic K+ clearance and swelling under high extracellular K+ concentrations. J. Physiol. Sci. 2016, 66, 127–142. [Google Scholar] [CrossRef]
- Mahmoud, S.; Gharagozloo, M.; Simard, C.; Gris, D. Astrocytes Maintain Glutamate Homeostasis in the CNS by Controlling the Balance between Glutamate Uptake and Release. Cells 2019, 8, 184. [Google Scholar] [CrossRef] [Green Version]
- Bellot-Saez, A.; Stevenson, R.; Kékesi, O.; Samokhina, E.; Ben-Abu, Y.; Morley, J.; Buskila, Y. Neuromodulation of Astrocytic K+ Clearance. Int. J. Mol. Sci. 2021, 22, 2520. [Google Scholar] [CrossRef]
- Bartolomei, F.; Chauvel, P.; Wendling, F. Epileptogenicity of brain structures in human temporal lobe epilepsy: A quantified study from intracerebral EEG. Brain 2008, 131, 1818–1830. [Google Scholar] [CrossRef] [Green Version]
- Aroniadou-Anderjaska, V.; Fritsch, B.; Qashu, F.; Braga, M.F. Pathology and pathophysiology of the amygdala in epileptogenesis and epilepsy. Epilepsy Res. 2008, 78, 102–116. [Google Scholar] [CrossRef] [Green Version]
- Olude, M.A.; Mustapha, O.A.; Aderounmu, O.A.; Olopade, J.O.; Ihunwo, A.O. Astrocyte morphology, heterogeneity, and density in the developing African giant rat (Cricetomys gambianus). Front. Neuroanat. 2015, 9, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allevi Protocols. Live/Dead Quantification Using Fiji—Step-by-Step Guide. Available online: https://www.allevi3d.com/livedead-assay-quantification-fiji/ (accessed on 24 September 2021).
- Chaure, F.; Rey, H.; Quiroga, R.Q. A novel and fully automatic spike-sorting implementation with variable number of features. J. Neurophysiol. 2018, 120, 1859–1871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Välkki, I.A.; Lenk, K.; Mikkonen, J.E.; Kapucu, F.E.; Hyttinen, J.A.K. Network-Wide Adaptive Burst Detection Depicts Neuronal Activity with Improved Accuracy. Front. Comput. Neurosci. 2017, 11, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapucu, F.E.; Tanskanen, J.M.A.; Mikkonen, J.E.; Ylä-Outinen, L.; Narkilahti, S.; Hyttinen, J.A.K. Burst analysis tool for developing neuronal networks exhibiting highly varying action potential dynamics. Front. Comput. Neurosci. 2012, 6, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapucu, F.E.; Välkki, I.; Mikkonen, J.E.; Leone, C.; Lenk, K.; Tanskanen, J.; Hyttinen, J. Spectral Entropy Based Neuronal Network Synchronization Analysis Based on Microelectrode Array Measurements. Front. Comput. Neurosci. 2016, 10, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagerland, M.W. T-tests, non-parametric tests, and large studies—a paradox of statistical practice? BMC Med. Res. Methodol. 2012, 12, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.; Lucas, H.C.; Shmueli, G. Research Commentary—Too Big to Fail: Large Samples and thep-Value Problem. Inf. Syst. Res. 2013, 24, 906–917. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.L.; Glass, G.V. Meta-analysis of psychotherapy outcome studies. Am. Psychol. 1977, 32, 752–760. [Google Scholar] [CrossRef]
- Lakens, D. Calculating and reporting effect sizes to facilitate cumulative science: A practical primer for t-tests and ANOVAs. Front. Psychol. 2013, 4, 863. [Google Scholar] [CrossRef] [Green Version]
- Hentschke, H.; Stüttgen, M.C. Computation of measures of effect size for neuroscience data sets. Eur. J. Neurosci. 2011, 34, 1887–1894. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Culture Platform | Cell Culture Type | n | Experiment/Analysis | Methods Section | Stimulation (4-AP 75 µM or Gabazine 30 µM) | Results Section | Results |
|---|---|---|---|---|---|---|---|
| MEAs | NS, 90/10, 80/20, 70/30, and 50/50 | 8 MEAs per cell culture type (total n = 40) | Spontaneous baseline activity recording | 4.6 | None (control) | 2.2 and 2.3 | NS cultures experienced more robust development at the later stages of culture (DIV21-28) compared to co-cultures. Development of electrical activity was more robust for co-cultures with higher ratios of astrocytes (70/30 and 50/50) at early stages of culture (DIV7-DIV14). Co-cultures with higher astrocyte numbers had lower spike and burst rates and a lower percentage of spikes in bursts. IBI increased with increasing neuron–astrocyte ratio. |
| Acute stimulation recording | 4.7 | 4-AP | 2.3 | Slight increase in SRs and BRs for NS, 90/10, and 80/20, whereas with 70/30 and 50/50 SRs remained mostly unchanged, and the BRs slightly decreased. The percentage of spikes in bursts was reduced in all the cultures, and the IBI and its variability were slightly increased. | |||
| Gabazine | 2.3 | SRs and BRs increased in all cultures. IBI values decreased for all cultures, and the percentage of spikes in bursts increased, synchronizing the network activity. | |||||
| Washout followed by recording after 24 h | 4.7 | 4-AP | 2.3 | SRs were generally lower than at baseline except for NS and 70/30. BRs were higher than at baseline for NS and all the co-cultures except 80/20. | |||
| Gabazine | 2.3 | SRs and BRs were generally lower for all cultures, except in the case of BRs in 90/10. | |||||
| Cross-correlation | 4.7.3 | 4-AP | 2.4 | Either no change or a small decrease in the numbers of channels with synchronized activity and the correlation weights of the connections. | |||
| Gabazine | 2.4 | Increased connection weights in all co-cultures which shifted the weight difference distribution towards positive values. | |||||
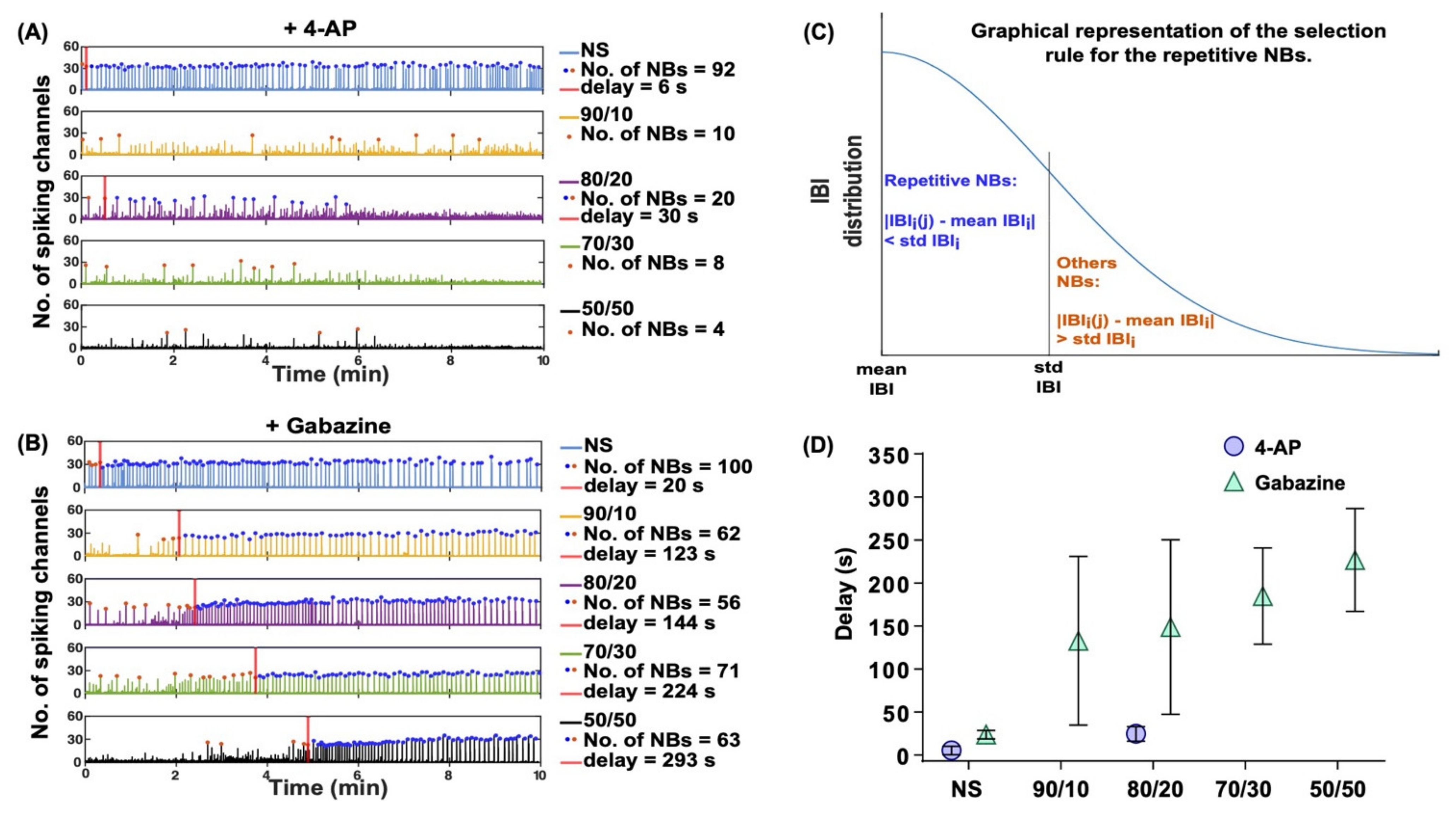
| NBs | 4.7.4 | 4-AP | 2.4 and 2.5 | The number of network-wide bursts decreased. | |||
| Gabazine | 2.4 and 2.5 | Repetitive synchronous NBs spanned over a constant number of channels (~40 channels). | |||||
| Convulsant response delay | 4.7.5 | 4-AP | 2.5 | No noticeable delays in the NB synchronization. | |||
| Gabazine | 2.5 | Increasing the relative numbers of astrocytes drastically delayed the effects of gabazine on neuronal firing (approximate delays were NS: 25 s; 90/10: 130 s; 80/20: 150 s; 70/30: 180 s; 50/50: 230 s). | |||||
| Coverslips | NS, 90/10, 80/20, 70/30, and 50/50 | 3–4 per staining | ICC | 4.3 | None (control) | 2.1 | Neuron–astrocyte ratios were coherent after DIV14 with plating densities, and neurons counts were homogeneous through all the coverslips counted. |
| Coverslips | NS | 3–4 | ICC | 4.3 | Control/4-AP/Gabazine | 2.3 | 4-AP slightly increased the neuronal viability. Neither 4-AP nor gabazine were toxic to neurons and did not decrease cell viability or affect their appearance. |
| 3–4 | Live/dead assay | 4.4 | |||||
| 24-Well Plates | Astrocytes | 2–3 | ICC | 4.3 | Control/4-AP/Gabazine | 2.3 | No differences in cell viability or appearance. |
| 2–3 | Live/dead assay | 4.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahtiainen, A.; Genocchi, B.; Tanskanen, J.M.A.; Barros, M.T.; Hyttinen, J.A.K.; Lenk, K. Astrocytes Exhibit a Protective Role in Neuronal Firing Patterns under Chemically Induced Seizures in Neuron–Astrocyte Co-Cultures. Int. J. Mol. Sci. 2021, 22, 12770. https://doi.org/10.3390/ijms222312770
Ahtiainen A, Genocchi B, Tanskanen JMA, Barros MT, Hyttinen JAK, Lenk K. Astrocytes Exhibit a Protective Role in Neuronal Firing Patterns under Chemically Induced Seizures in Neuron–Astrocyte Co-Cultures. International Journal of Molecular Sciences. 2021; 22(23):12770. https://doi.org/10.3390/ijms222312770
Chicago/Turabian StyleAhtiainen, Annika, Barbara Genocchi, Jarno M. A. Tanskanen, Michael T. Barros, Jari A. K. Hyttinen, and Kerstin Lenk. 2021. "Astrocytes Exhibit a Protective Role in Neuronal Firing Patterns under Chemically Induced Seizures in Neuron–Astrocyte Co-Cultures" International Journal of Molecular Sciences 22, no. 23: 12770. https://doi.org/10.3390/ijms222312770
APA StyleAhtiainen, A., Genocchi, B., Tanskanen, J. M. A., Barros, M. T., Hyttinen, J. A. K., & Lenk, K. (2021). Astrocytes Exhibit a Protective Role in Neuronal Firing Patterns under Chemically Induced Seizures in Neuron–Astrocyte Co-Cultures. International Journal of Molecular Sciences, 22(23), 12770. https://doi.org/10.3390/ijms222312770