
DNA Methylation in Huntington’s Disease


Abstract
:1. Introduction
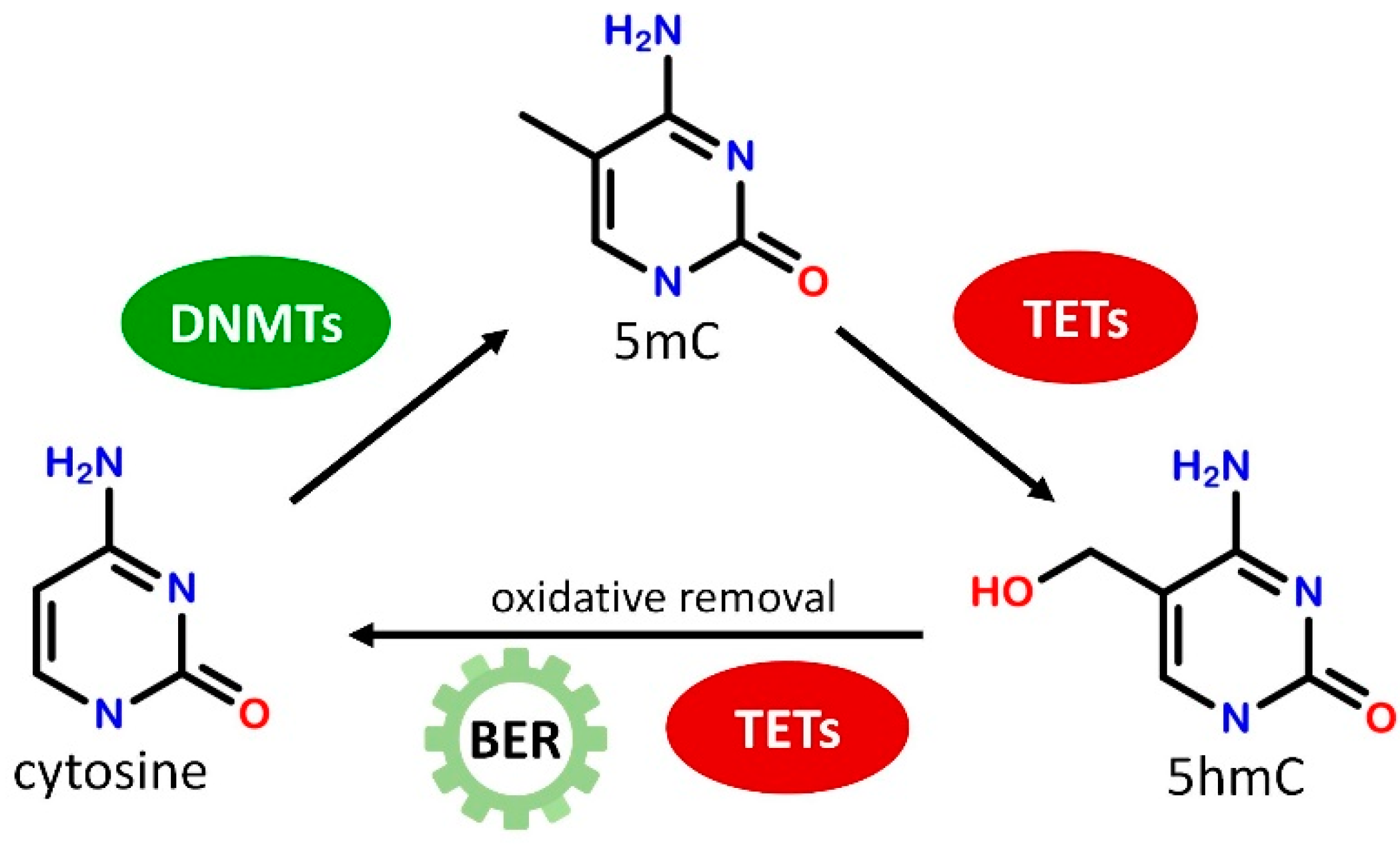
2. DNA Methylation and Its Role in Transcriptional Regulation
3. Lessons from Genome-Wide DNA Methylation Studies
4. Targeted Methylation Studies
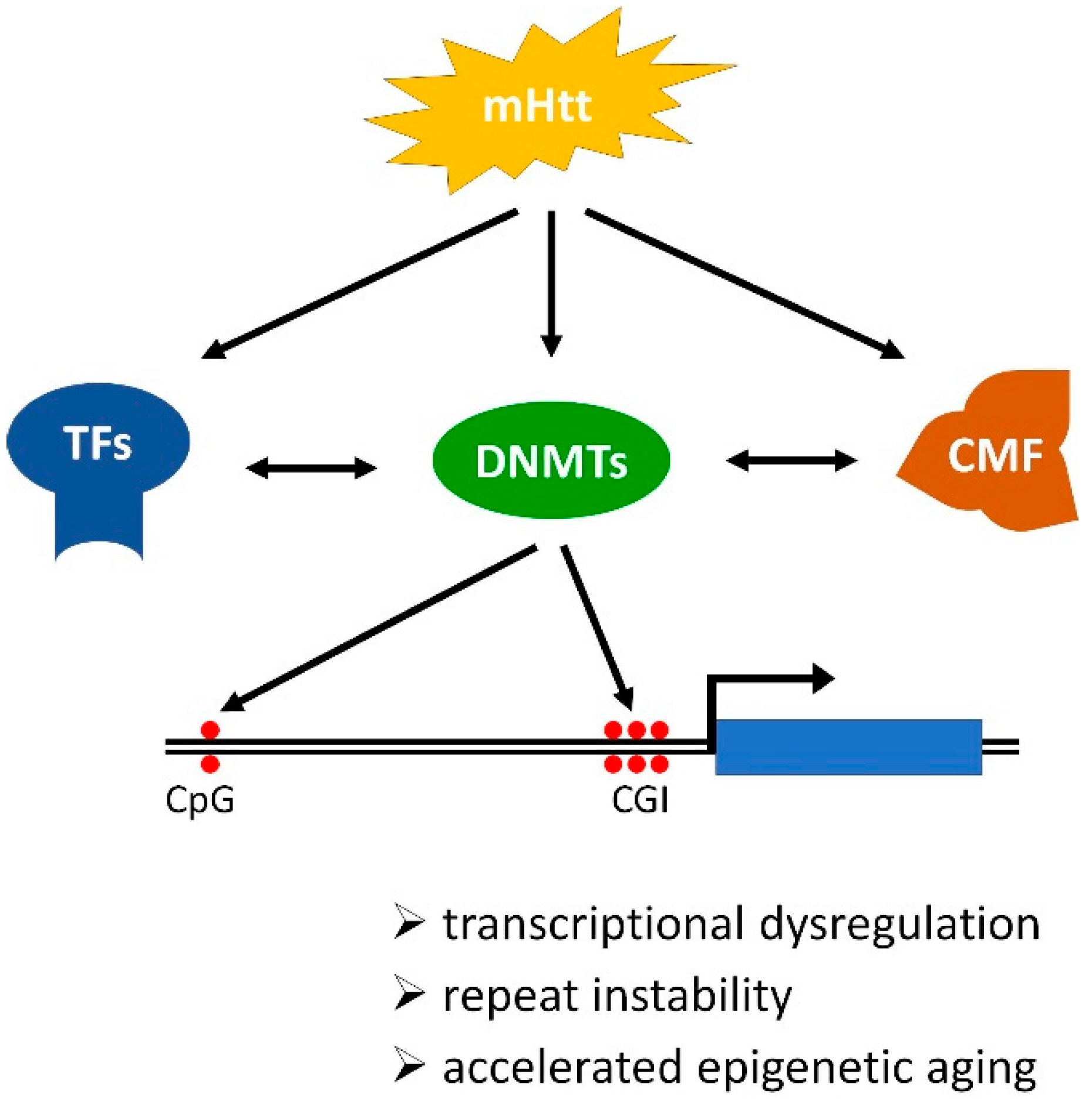
5. HD Impacts DNA Methylation Writers, Readers, and Erasers
6. Potential Role of DNA Methylation in Transgenerational Effects
7. DNA Methylation and Genome Maintenance in HD
8. Alternative Base Modifications in Huntington’s Disease
9. Possible Application of DNA Methylation as Biomarker in HD
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Model | Description | Reference |
|---|---|---|
| CAG140 KI | knock-in murine model expressing full-length Hdh gene with exon1 from human HTT with 140 CAG repeats | [79] |
| HD-hiPSCs | human-induced pluripotent stem cells derived from HD patients differentiated towards medium spiny neurons | [32] |
| Htt-72Q | murine primary cortical neuron model expressing HTT exon1 with 72 CAG repeats | [8] |
| OVT73 | transgenic sheep model expressing full-length human HTT cDNA with 73 CAG/CAA repeats under the control of the human promoter | [80] |
| R6/1 | murine model transgenic for the 5′ end of the HTT gene carrying 116 CAG repeats | [81] |
| R6/2 | murine model transgenic for the 5′ end of the HTT gene carrying 144 CAG repeats | [81] |
| N171-82Q | Transgenic murine model expressing N-terminal truncated human HTT cDNA with 82 CAG repeats under the influence of mouse prion protein promoter | [82] |
| STHdhQ111 | cell culture model derived from knock-in mouse embryos expressing full-length Hdh 1 with 111 CAG repeats | [83] |
| YAC128 | yeast artificial chromosome transgenic murine model expressing full-length HTT transgene with 128 CAG repeats | [84] |
| zQ175 | knock-in murine model expressing full-length Hdh gene with exon1 from human HTT with 188 CAG repeats, derived from CAG140 KI | [85] |
References
- Ross, C.A.; Tabrizi, S.J. Huntington’s Disease: From Molecular Pathogenesis to Clinical Treatment. Lancet Neurol. 2011, 10, 83–98. [Google Scholar] [CrossRef]
- The Huntington’s Disease Collaborative Research Group. A Novel Gene Containing a Trinucleotide Repeat That Is Expanded and Unstable on Huntington’s Disease Chromosomes. Cell 1993, 72, 971–983. [Google Scholar] [CrossRef]
- Bates, G.P.; Dorsey, R.; Gusella, J.F.; Hayden, M.R.; Kay, C.; Leavitt, B.R.; Nance, M.; Ross, C.A.; Scahill, R.I.; Wetzel, R.; et al. Huntington Disease. Nat. Rev. Dis. Primers 2015, 1, 15005. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, F.; Ng, C.W.; Kappes, V.; Ehrenberger, T.; Rigby, S.K.; Stivanello, V.; Gipson, T.A.; Soltis, A.R.; Vanhoutte, P.; Caboche, J.; et al. Early Epigenomic and Transcriptional Changes Reveal Elk-1 Transcription Factor as a Therapeutic Target in Huntington’s Disease. Proc. Natl. Acad. Sci. USA 2019, 116, 24840–24851. [Google Scholar] [CrossRef]
- Hervás-Corpión, I.; Guiretti, D.; Alcaraz-Iborra, M.; Olivares, R.; Campos-Caro, A.; Barco, Á.; Valor, L.M. Early Alteration of Epigenetic-Related Transcription in Huntington’s Disease Mouse Models. Sci. Rep. 2018, 8, 9925. [Google Scholar] [CrossRef] [PubMed]
- Ament, S.A.; Pearl, J.R.; Cantle, J.P.; Bragg, R.M.; Skene, P.J.; Coffey, S.R.; Bergey, D.E.; Wheeler, V.C.; MacDonald, M.E.; Baliga, N.S.; et al. Transcriptional Regulatory Networks Underlying Gene Expression Changes in Huntington’s Disease. Mol. Syst. Biol. 2018, 14, e7435. [Google Scholar] [CrossRef]
- Dunah, A.W.; Jeong, H.; Griffin, A.; Kim, Y.-M.; Standaert, D.G.; Hersch, S.M.; Mouradian, M.M.; Young, A.B.; Tanese, N.; Krainc, D. Sp1 and TAFII130 Transcriptional Activity Disrupted in Early Huntington’s Disease. Science 2002, 296, 2238–2243. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Zhu, Y.; Yang, W.; Tycksen, E.; Liu, S.; Palucki, J.; Zhu, L.; Sasaki, Y.; Sharma, M.K.; Kim, A.H.; et al. The Role of Twist1 in Mutant Huntingtin-Induced Transcriptional Alterations and Neurotoxicity. J. Biol. Chem. 2018, 293, 11850–11866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valor, L.M. Understanding Histone Deacetylation in Huntington’s Disease. Oncotarget 2016, 8, 5660–5661. [Google Scholar] [CrossRef] [PubMed]
- Vashishtha, M.; Ng, C.W.; Yildirim, F.; Gipson, T.A.; Kratter, I.H.; Bodai, L.; Song, W.; Lau, A.; Labadorf, A.; Vogel-Ciernia, A.; et al. Targeting H3K4 Trimethylation in Huntington Disease. Proc. Natl. Acad. Sci. USA 2013, 110, E3027–E3036. [Google Scholar] [CrossRef] [Green Version]
- Thomas, E.A. DNA Methylation in Huntington’s Disease: Implications for Transgenerational Effects. Neurosci. Lett. 2016, 625, 34–39. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Pallos, J.; Jacques, V.; Lau, A.; Tang, B.; Cooper, A.; Syed, A.; Purcell, J.; Chen, Y.; Sharma, S.; et al. Histone Deacetylase (HDAC) Inhibitors Targeting HDAC3 and HDAC1 Ameliorate Polyglutamine-Elicited Phenotypes in Model Systems of Huntington’s Disease. Neurobiol. Dis. 2012, 46, 351–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.; Zsindely, N.; Faragó, A.; Marsh, J.L.; Bodai, L. Systematic Genetic Interaction Studies Identify Histone Demethylase Utx as Potential Target for Ameliorating Huntington’s Disease. Hum. Mol. Genet. 2018, 27, 649–666. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Daito, T.; Sasaki, Y.; Chung, Y.H.; Xing, X.; Pondugula, S.; Swamidass, S.J.; Wang, T.; Kim, A.H.; Yano, H. Inhibition of DNA Methyltransferases Blocks Mutant Huntingtin-Induced Neurotoxicity. Sci. Rep. 2016, 6, 31022. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.A. Functions of DNA Methylation: Islands, Start Sites, Gene Bodies and Beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA Methylation and Its Basic Function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Leonhardt, H.; Page, A.W.; Weier, H.U.; Bestor, T.H. A Targeting Sequence Directs DNA Methyltransferase to Sites of DNA Replication in Mammalian Nuclei. Cell 1992, 71, 865–873. [Google Scholar] [CrossRef] [Green Version]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA Methyltransferases Dnmt3a and Dnmt3b Are Essential for de Novo Methylation and Mammalian Development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Zhou, Y.; Campbell, S.L.; Le, T.; Li, E.; Sweatt, J.D.; Silva, A.J.; Fan, G. Dnmt1 and Dnmt3a Maintain DNA Methylation and Regulate Synaptic Function in Adult Forebrain Neurons. Nat. Neurosci. 2010, 13, 423–430. [Google Scholar] [CrossRef]
- Bhutani, N.; Burns, D.M.; Blau, H.M. DNA Demethylation Dynamics. Cell 2011, 146, 866–872. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Song, C.-X.; He, C.; Zhang, Y. Mechanism and Function of Oxidative Reversal of DNA and RNA Methylation. Annu. Rev. Biochem. 2014, 83, 585–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jobe, E.M.; Zhao, X. DNA Methylation and Adult Neurogenesis. Brain Plast. 2017, 3, 5–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nan, X.; Ng, H.H.; Johnson, C.A.; Laherty, C.D.; Turner, B.M.; Eisenman, R.N.; Bird, A. Transcriptional Repression by the Methyl-CpG-Binding Protein MeCP2 Involves a Histone Deacetylase Complex. Nature 1998, 393, 386–389. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.L.; Veenstra, G.J.; Wade, P.A.; Vermaak, D.; Kass, S.U.; Landsberger, N.; Strouboulis, J.; Wolffe, A.P. Methylated DNA and MeCP2 Recruit Histone Deacetylase to Repress Transcription. Nat. Genet. 1998, 19, 187–191. [Google Scholar] [CrossRef]
- Kimura, H.; Shiota, K. Methyl-CpG-Binding Protein, MeCP2, Is a Target Molecule for Maintenance DNA Methyltransferase, Dnmt1. J. Biol. Chem. 2003, 278, 4806–4812. [Google Scholar] [CrossRef] [Green Version]
- Bostick, M.; Kim, J.K.; Estève, P.-O.; Clark, A.; Pradhan, S.; Jacobsen, S.E. UHRF1 Plays a Role in Maintaining DNA Methylation in Mammalian Cells. Science 2007, 317, 1760–1764. [Google Scholar] [CrossRef] [Green Version]
- Skene, P.J.; Illingworth, R.S.; Webb, S.; Kerr, A.R.W.; James, K.D.; Turner, D.J.; Andrews, R.; Bird, A.P. Neuronal MeCP2 Is Expressed at near Histone-Octamer Levels and Globally Alters the Chromatin State. Mol. Cell 2010, 37, 457–468. [Google Scholar] [CrossRef] [Green Version]
- Fan, G.; Hutnick, L. Methyl-CpG Binding Proteins in the Nervous System. Cell Res. 2005, 15, 255–261. [Google Scholar] [CrossRef]
- Ng, C.W.; Yildirim, F.; Yap, Y.S.; Dalin, S.; Matthews, B.J.; Velez, P.J.; Labadorf, A.; Housman, D.E.; Fraenkel, E. Extensive Changes in DNA Methylation Are Associated with Expression of Mutant Huntingtin. Proc. Natl. Acad. Sci. USA 2013, 110, 2354–2359. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Yang, Y.; Lin, X.; Wang, J.-Q.; Wu, Y.-S.; Xie, W.; Wang, D.; Zhu, S.; Liao, Y.-Q.; Sun, Q.; et al. Genome-Wide Loss of 5-HmC Is a Novel Epigenetic Feature of Huntington’s Disease. Hum. Mol. Genet. 2013, 22, 3641–3653. [Google Scholar] [CrossRef]
- Kumar, S.; Chinnusamy, V.; Mohapatra, T. Epigenetics of Modified DNA Bases: 5-Methylcytosine and Beyond. Front. Genet. 2018, 9, 640. [Google Scholar] [CrossRef] [Green Version]
- Baronchelli, S.; La Spada, A.; Ntai, A.; Barbieri, A.; Conforti, P.; Jotti, G.S.; Redaelli, S.; Bentivegna, A.; De Blasio, P.; Biunno, I. Epigenetic and Transcriptional Modulation of WDR5, a Chromatin Remodeling Protein, in Huntington’s Disease Human Induced Pluripotent Stem Cell (HiPSC) Model. Mol. Cell Neurosci. 2017, 82, 46–57. [Google Scholar] [CrossRef]
- Horvath, S.; Langfelder, P.; Kwak, S.; Aaronson, J.; Rosinski, J.; Vogt, T.F.; Eszes, M.; Faull, R.L.M.; Curtis, M.A.; Waldvogel, H.J.; et al. Huntington’s Disease Accelerates Epigenetic Aging of Human Brain and Disrupts DNA Methylation Levels. Aging 2016, 8, 1485–1512. [Google Scholar] [CrossRef] [Green Version]
- Landwehrmeyer, G.B.; Fitzer-Attas, C.J.; Giuliano, J.D.; Gonçalves, N.; Anderson, K.E.; Cardoso, F.; Ferreira, J.J.; Mestre, T.A.; Stout, J.C.; Sampaio, C. Data Analytics from Enroll-HD, a Global Clinical Research Platform for Huntington’s Disease. Mov. Disord. Clin. Pract. 2017, 4, 212–224. [Google Scholar] [CrossRef] [Green Version]
- Orth, M.; Handley, O.J.; Schwenke, C.; Dunnett, S.B.; Craufurd, D.; Ho, A.K.; Wild, E.; Tabrizi, S.J.; Landwehrmeyer, G.B.; Investigators of the European Huntington’s Disease Network. Observing Huntington’s Disease: The European Huntington’s Disease Network’s REGISTRY. PLoS Curr. 2010, 2, RRN1184. [Google Scholar] [CrossRef]
- Lu, A.T.; Narayan, P.; Grant, M.J.; Langfelder, P.; Wang, N.; Kwak, S.; Wilkinson, H.; Chen, R.Z.; Chen, J.; Simon Bawden, C.; et al. DNA Methylation Study of Huntington’s Disease and Motor Progression in Patients and in Animal Models. Nat. Commun. 2020, 11, 4529. [Google Scholar] [CrossRef] [PubMed]
- De Souza, R.A.G.; Islam, S.A.; McEwen, L.M.; Mathelier, A.; Hill, A.; Mah, S.M.; Wasserman, W.W.; Kobor, M.S.; Leavitt, B.R. DNA Methylation Profiling in Human Huntington’s Disease Brain. Hum. Mol. Genet. 2016, 25, 2013–2030. [Google Scholar] [CrossRef] [Green Version]
- Horvath, S. DNA Methylation Age of Human Tissues and Cell Types. Genome Biol. 2013, 14, 3156. [Google Scholar] [CrossRef] [Green Version]
- Vonsattel, J.P.; Myers, R.H.; Stevens, T.J.; Ferrante, R.J.; Bird, E.D.; Richardson, E.P. Neuropathological Classification of Huntington’s Disease. J. Neuropathol. Exp. Neurol. 1985, 44, 559–577. [Google Scholar] [CrossRef] [PubMed]
- Chi, S.; Song, J.-H.; Tan, M.-S.; Zhang, W.; Wang, Z.-X.; Jiang, T.; Tan, L.; Yu, J.-T. Association of Single-Nucleotide Polymorphism in ANK1 with Late-Onset Alzheimer’s Disease in Han Chinese. Mol. Neurobiol. 2016, 53, 6476–6481. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.R.; Smith, R.G.; Burrage, J.; Troakes, C.; Al-Sarraj, S.; Kalaria, R.N.; Sloan, C.; Robinson, A.C.; Mill, J.; Lunnon, K. A Cross-Brain Regions Study of ANK1 DNA Methylation in Different Neurodegenerative Diseases. Neurobiol. Aging 2019, 74, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Zhang, Y.; Wang, C.; Xu, Z.; Liang, Q.; An, L.; Li, J.; Liu, Z.; You, Y.; He, M.; et al. The Zinc Finger Transcription Factor Sp9 Is Required for the Development of Striatopallidal Projection Neurons. Cell Rep. 2016, 16, 1431–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarditi, A.; Camurri, A.; Varani, K.; Borea, P.A.; Woodman, B.; Bates, G.; Cattaneo, E.; Abbracchio, M.P. Early and Transient Alteration of Adenosine A2A Receptor Signaling in a Mouse Model of Huntington Disease. Neurobiol. Dis. 2006, 23, 44–53. [Google Scholar] [CrossRef]
- Glass, M.; Dragunow, M.; Faull, R.L. The Pattern of Neurodegeneration in Huntington’s Disease: A Comparative Study of Cannabinoid, Dopamine, Adenosine and GABA(A) Receptor Alterations in the Human Basal Ganglia in Huntington’s Disease. Neuroscience 2000, 97, 505–519. [Google Scholar] [CrossRef]
- Villar-Menéndez, I.; Blanch, M.; Tyebji, S.; Pereira-Veiga, T.; Albasanz, J.L.; Martín, M.; Ferrer, I.; Pérez-Navarro, E.; Barrachina, M. Increased 5-Methylcytosine and Decreased 5-Hydroxymethylcytosine Levels Are Associated with Reduced Striatal A2AR Levels in Huntington’s Disease. Neuromol. Med. 2013, 15, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Zuccato, C.; Ciammola, A.; Rigamonti, D.; Leavitt, B.R.; Goffredo, D.; Conti, L.; MacDonald, M.E.; Friedlander, R.M.; Silani, V.; Hayden, M.R.; et al. Loss of Huntingtin-Mediated BDNF Gene Transcription in Huntington’s Disease. Science 2001, 293, 493–498. [Google Scholar] [CrossRef] [Green Version]
- Zuccato, C.; Cattaneo, E. Brain-Derived Neurotrophic Factor in Neurodegenerative Diseases. Nat. Rev. Neurol. 2009, 5, 311–322. [Google Scholar] [CrossRef]
- Qin, Q.; Xu, Y.; He, T.; Qin, C.; Xu, J. Normal and Disease-Related Biological Functions of Twist1 and Underlying Molecular Mechanisms. Cell Res. 2012, 22, 90–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labadorf, A.; Hoss, A.G.; Lagomarsino, V.; Latourelle, J.C.; Hadzi, T.C.; Bregu, J.; MacDonald, M.E.; Gusella, J.F.; Chen, J.-F.; Akbarian, S.; et al. RNA Sequence Analysis of Human Huntington Disease Brain Reveals an Extensive Increase in Inflammatory and Developmental Gene Expression. PLoS ONE 2015, 10, e0143563. [Google Scholar] [CrossRef] [Green Version]
- Mollica, P.A.; Zamponi, M.; Reid, J.A.; Sharma, D.K.; White, A.E.; Ogle, R.C.; Bruno, R.D.; Sachs, P.C. Epigenetic Alterations Mediate IPSC-Induced Normalization of DNA Repair Gene Expression and TNR Stability in Huntington’s Disease Cells. J. Cell Sci. 2018, 131, jcs215343. [Google Scholar] [CrossRef] [Green Version]
- Bayer, C.; Pitschelatow, G.; Hannemann, N.; Linde, J.; Reichard, J.; Pensold, D.; Zimmer-Bensch, G. DNA Methyltransferase 1 (DNMT1) Acts on Neurodegeneration by Modulating Proteostasis-Relevant Intracellular Processes. Int. J. Mol. Sci. 2020, 21, 5420. [Google Scholar] [CrossRef]
- McFarland, K.N.; Huizenga, M.N.; Darnell, S.B.; Sangrey, G.R.; Berezovska, O.; Cha, J.-H.J.; Outeiro, T.F.; Sadri-Vakili, G. MeCP2: A Novel Huntingtin Interactor. Hum. Mol. Genet. 2014, 23, 1036–1044. [Google Scholar] [CrossRef] [Green Version]
- Martin, E.M.; Fry, R.C. Environmental Influences on the Epigenome: Exposure-Associated DNA Methylation in Human Populations. Annu. Rev. Public Health 2018, 39, 309–333. [Google Scholar] [CrossRef] [Green Version]
- Crider, K.S.; Yang, T.P.; Berry, R.J.; Bailey, L.B. Folate and DNA Methylation: A Review of Molecular Mechanisms and the Evidence for Folate’s Role. Adv. Nutr. 2012, 3, 21–38. [Google Scholar] [CrossRef] [Green Version]
- Ly, A.; Hoyt, L.; Crowell, J.; Kim, Y.-I. Folate and DNA Methylation. Antioxid. Redox Signal. 2012, 17, 302–326. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, P.; Singh, N. Homocysteine Excess: Delineating the Possible Mechanism of Neurotoxicity and Depression. Fundam. Clin. Pharmacol. 2015, 29, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Craenen, K.; Verslegers, M.; Baatout, S.; Abderrafi Benotmane, M. An Appraisal of Folates as Key Factors in Cognition and Ageing-Related Diseases. Crit. Rev. Food Sci. Nutr. 2020, 60, 722–739. [Google Scholar] [CrossRef]
- de Wilde, M.C.; Vellas, B.; Girault, E.; Yavuz, A.C.; Sijben, J.W. Lower Brain and Blood Nutrient Status in Alzheimer’s Disease: Results from Meta-Analyses. Alzheimers Dement. 2017, 3, 416–431. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, S.; Ji, L.; Wu, T.; Ji, Y.; Zhou, Y.; Zheng, M.; Zhang, M.; Xu, W.; Huang, G. Folic Acid Supplementation Mitigates Alzheimer’s Disease by Reducing Inflammation: A Randomized Controlled Trial. Mediat. Inflamm. 2016, 2016, 5912146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, V. Methylenetetrahydrofolate Reductase (MTHFR) C677T Polymorphism and Alzheimer Disease Risk: A Meta-Analysis. Mol. Neurobiol. 2017, 54, 1173–1186. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, E.; Estévez, J.; Suárez, H.; Morales, L.M.; Chacin de Bonilla, L.; Villalobos, R.; Dávila, J.O. Serum Ferritin Deficiency in Huntington’s Disease Patients. Neurosci. Lett. 1991, 129, 22–24. [Google Scholar] [CrossRef]
- von Albert, H.H. Futile treatment of Huntington’s chorea using folic acid. Nervenarzt 1976, 47, 628–629. [Google Scholar]
- Brune, N.; Andrich, J.; Gencik, M.; Saft, C.; Müller, T.; Valentin, S.; Przuntek, H.; Epplen, J.T. Methyltetrahydrofolate Reductase Polymorphism Influences Onset of Huntington’s Disease. J. Neural. Transm. Suppl. 2004, 105–110. [Google Scholar] [CrossRef]
- Hansen, W.; Saft, C.; Andrich, J.; Müller, T.; Wieczorek, S.; Epplen, J.T.; Arning, L. Failure to Confirm Influence of Methyltetrahydrofolate Reductase (MTHFR) Polymorphisms on Age at Onset of Huntington Disease. J. Negat. Results Biomed. 2005, 4, 12. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Morris, C.D.; Williams, R.M.; Loring, J.F.; Thomas, E.A. HDAC Inhibition Imparts Beneficial Transgenerational Effects in Huntington’s Disease Mice via Altered DNA and Histone Methylation. Proc. Natl. Acad. Sci. USA 2015, 112, E56–E64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, H.; Kast, R.J.; Steffan, J.S.; Thomas, E.A. Selective Histone Deacetylase (HDAC) Inhibition Imparts Beneficial Effects in Huntingtons Disease Mice: Implications for the Ubiquitinproteasomal and Autophagy Systems. Hum. Mol. Genet. 2012, 21, 5280–5293. [Google Scholar] [CrossRef] [PubMed]
- Mahalingam, S.; Levy, L.M. Genetics of Huntington Disease. Am. J. Neuroradiol. 2014, 35, 1070–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollica, P.A.; Reid, J.A.; Ogle, R.C.; Sachs, P.C.; Bruno, R.D. DNA Methylation Leads to DNA Repair Gene Down-Regulation and Trinucleotide Repeat Expansion in Patient-Derived Huntington Disease Cells. Am. J. Pathol. 2016, 186, 1967–1976. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.; Wu, C.; Jin, L. A Possible Role for Long Interspersed Nuclear Elements-1 (LINE-1) in Huntington’s Disease Progression. Med. Sci. Monit. 2018, 24, 3644–3652. [Google Scholar] [CrossRef]
- Boysen, G.; Pachkowski, B.F.; Nakamura, J.; Swenberg, J.A. The Formation and Biological Significance of N7-Guanine Adducts. Mutat. Res./Genet. Toxicol. Environ. Mutagenes. 2009, 678, 76–94. [Google Scholar] [CrossRef] [Green Version]
- Thomas, B.; Matson, S.; Chopra, V.; Sun, L.; Sharma, S.; Hersch, S.; Rosas, H.D.; Scherzer, C.; Ferrante, R.; Matson, W. A Novel Method for Detecting 7-Methyl Guanine Reveals Aberrant Methylation Levels in Huntington Disease. Anal. Biochem. 2013, 436, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, Y.P.; Andrew, S.E.; Clarke, L.A.; Hayden, M.R. A PCR Method for Accurate Assessment of Trinucleotide Repeat Expansion in Huntington Disease. Hum. Mol. Genet. 1993, 2, 635–636. [Google Scholar] [CrossRef] [PubMed]
- Przybyl, L.; Wozna-Wysocka, M.; Kozlowska, E.; Fiszer, A. What, When and How to Measure—Peripheral Biomarkers in Therapy of Huntington’s Disease. Int. J. Mol. Sci. 2021, 22, 1561. [Google Scholar] [CrossRef]
- Lin, P.-I.; Shu, H.; Mersha, T.B. Comparing DNA Methylation Profiles across Different Tissues Associated with the Diagnosis of Pediatric Asthma. Sci. Rep. 2020, 10, 151. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, A.; Corey-Bloom, J.; Thomas, E.A.; Desplats, P. Evaluation of Biochemical and Epigenetic Measures of Peripheral Brain-Derived Neurotrophic Factor (BDNF) as a Biomarker in Huntington’s Disease Patients. Front. Mol. Neurosci. 2019, 12, 335. [Google Scholar] [CrossRef] [Green Version]
- Zuccato, C.; Marullo, M.; Vitali, B.; Tarditi, A.; Mariotti, C.; Valenza, M.; Lahiri, N.; Wild, E.J.; Sassone, J.; Ciammola, A.; et al. Brain-Derived Neurotrophic Factor in Patients with Huntington’s Disease. PLoS ONE 2011, 6, e22966. [Google Scholar] [CrossRef] [Green Version]
- Ou, Z.-Y.A.; Byrne, L.M.; Rodrigues, F.B.; Tortelli, R.; Johnson, E.B.; Foiani, M.S.; Arridge, M.; De Vita, E.; Scahill, R.I.; Heslegrave, A.; et al. Brain-Derived Neurotrophic Factor in Cerebrospinal Fluid and Plasma Is Not a Biomarker for Huntington’s Disease. Sci. Rep. 2021, 11, 3481. [Google Scholar] [CrossRef] [PubMed]
- Zadel, M.; Maver, A.; Kovanda, A.; Peterlin, B. DNA Methylation Profiles in Whole Blood of Huntington’s Disease Patients. Front. Neurol. 2018, 9, 655. [Google Scholar] [CrossRef] [PubMed]
- Menalled, L.B.; Sison, J.D.; Dragatsis, I.; Zeitlin, S.; Chesselet, M.-F. Time Course of Early Motor and Neuropathological Anomalies in a Knock-in Mouse Model of Huntington’s Disease with 140 CAG Repeats. J. Comp. Neurol. 2003, 465, 11–26. [Google Scholar] [CrossRef]
- Jacobsen, J.C.; Bawden, C.S.; Rudiger, S.R.; McLaughlan, C.J.; Reid, S.J.; Waldvogel, H.J.; MacDonald, M.E.; Gusella, J.F.; Walker, S.K.; Kelly, J.M.; et al. An Ovine Transgenic Huntington’s Disease Model. Hum. Mol. Genet. 2010, 19, 1873–1882. [Google Scholar] [CrossRef] [Green Version]
- Mangiarini, L.; Sathasivam, K.; Seller, M.; Cozens, B.; Harper, A.; Hetherington, C.; Lawton, M.; Trottier, Y.; Lehrach, H.; Davies, S.W.; et al. Exon 1 of the HD Gene with an Expanded CAG Repeat Is Sufficient to Cause a Progressive Neurological Phenotype in Transgenic Mice. Cell 1996, 87, 493–506. [Google Scholar] [CrossRef] [Green Version]
- Schilling, G.; Becher, M.W.; Sharp, A.H.; Jinnah, H.A.; Duan, K.; Kotzuk, J.A.; Slunt, H.H.; Ratovitski, T.; Cooper, J.K.; Jenkins, N.A.; et al. Intranuclear Inclusions and Neuritic Aggregates in Transgenic Mice Expressing a Mutant N-Terminal Fragment of Huntingtin. Hum. Mol. Genet. 1999, 8, 397–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trettel, F.; Rigamonti, D.; Hilditch-Maguire, P.; Wheeler, V.C.; Sharp, A.H.; Persichetti, F.; Cattaneo, E.; MacDonald, M.E. Dominant Phenotypes Produced by the HD Mutation in STHdhQ111 Striatal Cells. Hum. Mol. Genet. 2000, 9, 2799–2809. [Google Scholar] [CrossRef]
- Slow, E.J.; van Raamsdonk, J.; Rogers, D.; Coleman, S.H.; Graham, R.K.; Deng, Y.; Oh, R.; Bissada, N.; Hossain, S.M.; Yang, Y.-Z.; et al. Selective Striatal Neuronal Loss in a YAC128 Mouse Model of Huntington Disease. Hum. Mol. Genet. 2003, 12, 1555–1567. [Google Scholar] [CrossRef] [PubMed]
- Menalled, L.B.; Kudwa, A.E.; Miller, S.; Fitzpatrick, J.; Watson-Johnson, J.; Keating, N.; Ruiz, M.; Mushlin, R.; Alosio, W.; McConnell, K.; et al. Comprehensive Behavioral and Molecular Characterization of a New Knock-in Mouse Model of Huntington’s Disease: ZQ175. PLoS ONE 2012, 7, e49838. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zsindely, N.; Siági, F.; Bodai, L. DNA Methylation in Huntington’s Disease. Int. J. Mol. Sci. 2021, 22, 12736. https://doi.org/10.3390/ijms222312736
Zsindely N, Siági F, Bodai L. DNA Methylation in Huntington’s Disease. International Journal of Molecular Sciences. 2021; 22(23):12736. https://doi.org/10.3390/ijms222312736
Chicago/Turabian StyleZsindely, Nóra, Fruzsina Siági, and László Bodai. 2021. "DNA Methylation in Huntington’s Disease" International Journal of Molecular Sciences 22, no. 23: 12736. https://doi.org/10.3390/ijms222312736
APA StyleZsindely, N., Siági, F., & Bodai, L. (2021). DNA Methylation in Huntington’s Disease. International Journal of Molecular Sciences, 22(23), 12736. https://doi.org/10.3390/ijms222312736