
Wnt-Signaling Regulated by Glucocorticoid-Induced miRNAs


Abstract
1. Introduction
2. Results
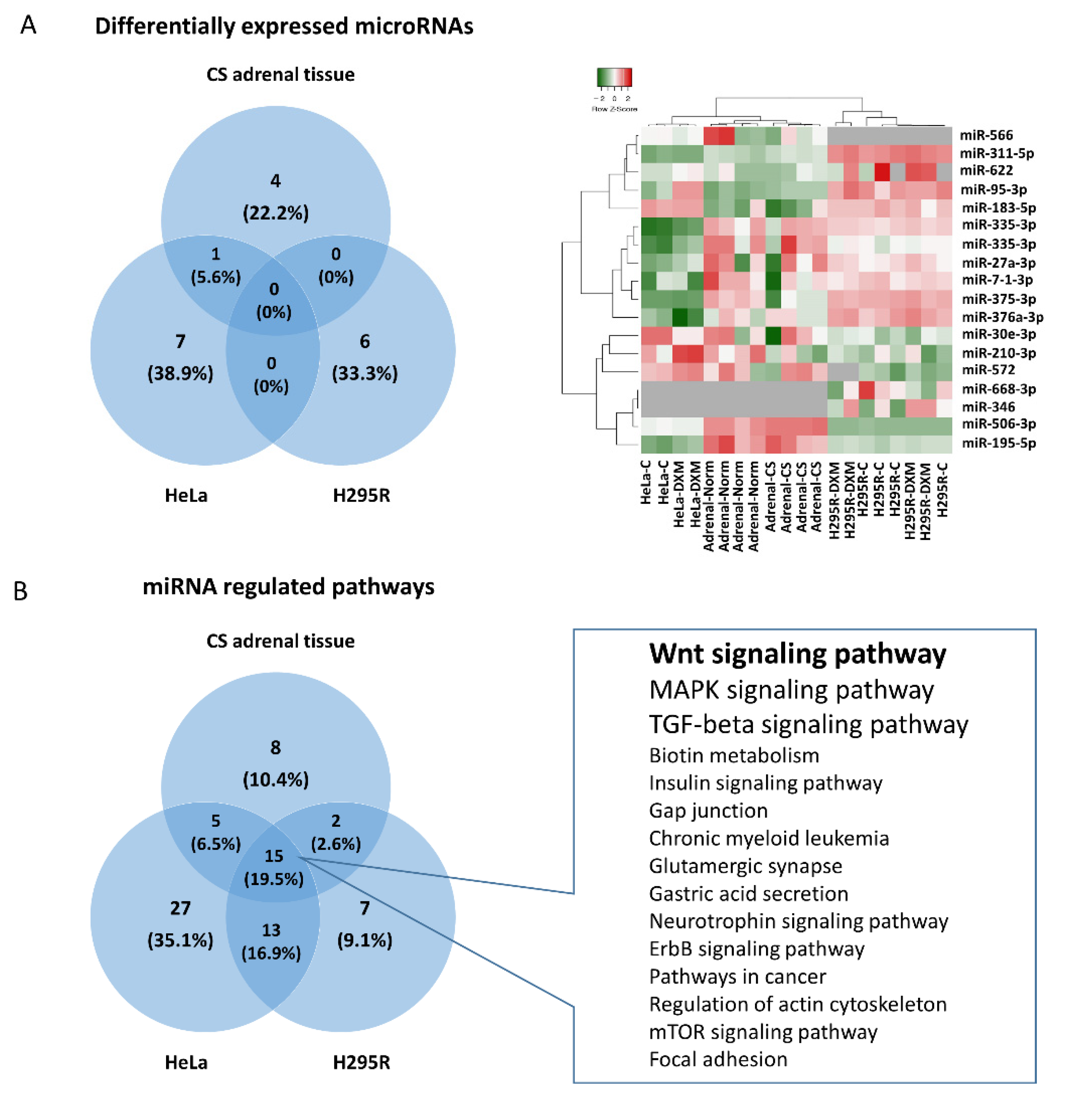
2.1. Differentially Expressed miRNAs in Glucocorticoid Secreting Adrenal Adenoma Tissues
2.2. Expression Change of miRNAs in H295R and HeLa Cell Lines after Dexamethasone Treatment
2.3. Common Pathways Regulated by Hypercortisolism through Different miRNAs in Different Cell Types
2.4. Gene Expression Profiling of GC Producing and GC Target Tissue Types
2.5. Investigation of Global Gene Expression Alteration upon Glucocorticoid Effect on Wnt Pathway Alteration in GC Target Tissues
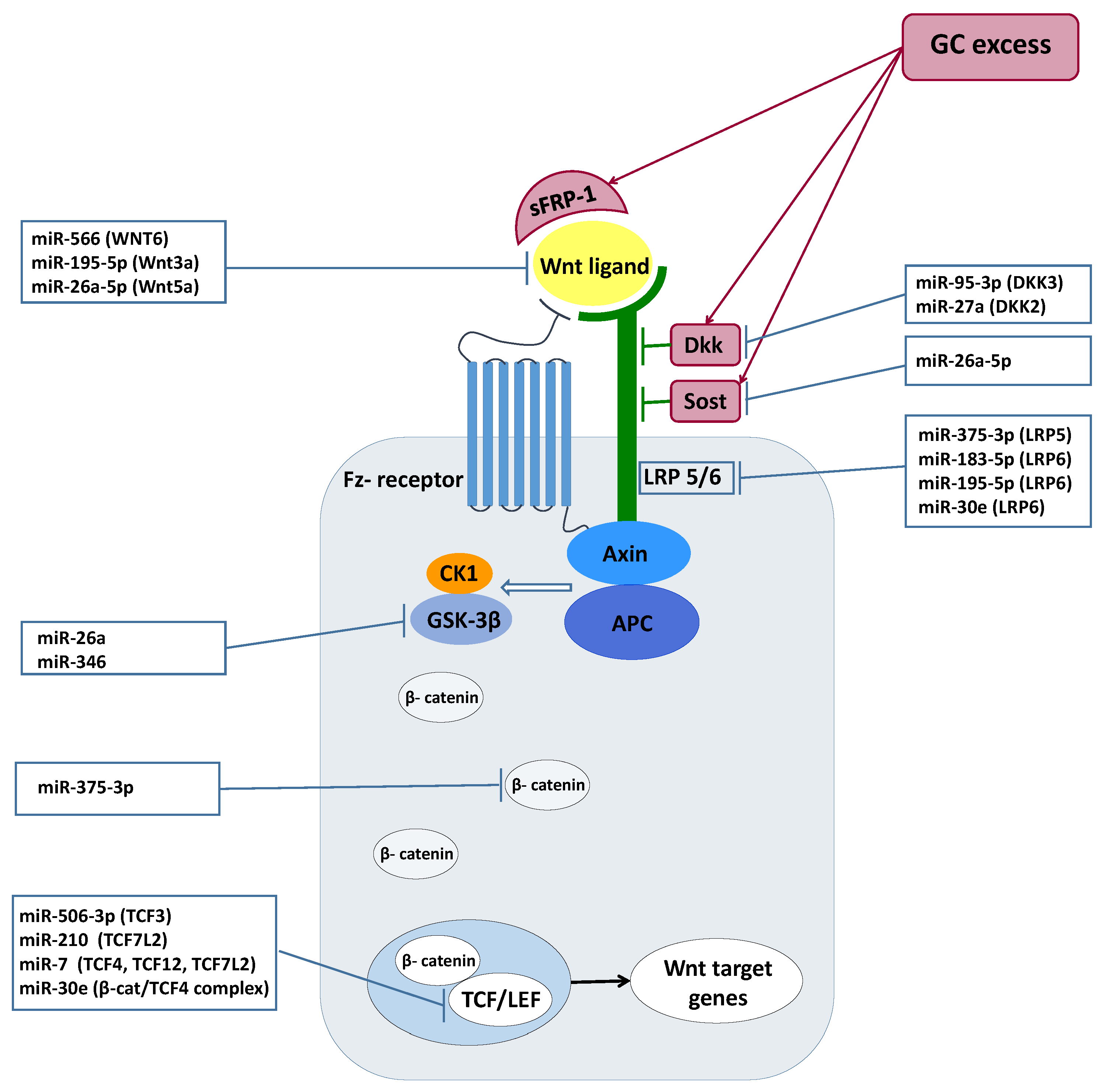
2.6. Tissue-Specific Target Prediction of Wnt Pathway in GC-Producing Adrenal Adenoma
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. In Vitro Cell Culture Experiments
4.3. Gene Expression Profiling and Gene Expression Re-Analysis
4.4. RNA Extraction
4.5. miRNA Profiling in Adrenocortical Tissues Using TLDA Cards
4.6. Gene Set Enrichment and Pathway Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guaraldi, F.; Salvatori, R. Cushing Syndrome: Maybe Not So Uncommon of an Endocrine Disease. J. Am. Board Fam. Med. 2012, 25, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Wengander, S.; Trimpou, P.; Papakokkinou, E.; Ragnarsson, O. The Incidence of Endogenous Cushing’s Syndrome in the Modern Era. Clin. Endocrinol. 2019, 91, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, S.; Cidlowski, J.A. Corticosteroids. Rheum. Dis. Clin. N. Am. 2016, 42, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Whirledge, S.; DeFranco, D.B. Glucocorticoid Signaling in Health and Disease: Insights from Tissue-Specific GR Knockout Mice. Endocrinology 2018, 159, 46–64. [Google Scholar] [CrossRef] [PubMed]
- Hadjidakis, D.J.; Androulakis, I.I. Bone Remodeling. Ann. N. Y. Acad. Sci. 2006, 1092, 385–396. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Otsuka, F.; Takano, M.; Mukai, T.; Yamanaka, R.; Takeda, M.; Miyoshi, T.; Inagaki, K.; Sada, K.; Makino, H. Estrogen and Glucocorticoid Regulate Osteoblast Differentiation through the Interaction of Bone Morphogenetic Protein-2 and Tumor Necrosis Factor-α in C2C12 Cells. Mol. Cell. Endocrinol. 2010, 325, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Grugan, K.D.; Ma, C.; Singhal, S.; Krett, N.L.; Rosen, S.T. Dual Regulation of Glucocorticoid-Induced Leucine Zipper (GILZ) by the Glucocorticoid Receptor and the PI3-Kinase/AKT Pathways in Multiple Myeloma. J. Steroid Biochem. Mol. Biol. 2008, 110, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Ramshanker, N.; Aagaard, M.; Hjortebjerg, R.; Voss, T.S.; Møller, N.; Jørgensen, J.O.L.; Jessen, N.; Bjerring, P.; Magnusson, N.E.; Bjerre, M.; et al. Effects of Prednisolone on Serum and Tissue Fluid IGF-I Receptor Activation and Post-Receptor Signaling in Humans. J. Clin. Endocrinol. Metab. 2017, 102, 4031–4040. [Google Scholar] [CrossRef] [PubMed]
- Ronchetti, S.; Migliorati, G.; Riccardi, C. GILZ as a Mediator of the Anti-Inflammatory Effects of Glucocorticoids. Front. Endocrinol. 2015, 6, 170. [Google Scholar] [CrossRef]
- Mak, W.; Shao, X.; Dunstan, C.R.; Seibel, M.J.; Zhou, H. Biphasic Glucocorticoid-Dependent Regulation of Wnt Expression and Its Inhibitors in Mature Osteoblastic Cells. Calcif. Tissue Int. 2009, 85, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.; Kumar, R. Crosstalk between NFkB and Glucocorticoid Signaling: A Potential Target of Breast Cancer Therapy. Cancer Lett. 2012, 322, 119–126. [Google Scholar] [CrossRef]
- Komori, T. Glucocorticoid Signaling and Bone Biology. Horm. Metab. Res. 2016, 48, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.Y.; Cregor, M.; Delgado-Calle, J.; Condon, K.W.; Allen, M.R.; Peacock, M.; Plotkin, L.I.; Bellido, T. Protection from Glucocorticoid-Induced Osteoporosis by Anti-Catabolic Signaling in the Absence of Sost/Sclerostin. J. Bone Min. Res. 2016, 31, 1791–1802. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.-S.; Ko, J.-Y.; Yeh, D.-W.; Ke, H.-C.; Wu, H.-L. Modulation of Dickkopf-1 Attenuates Glucocorticoid Induction of Osteoblast Apoptosis, Adipocytic Differentiation, and Bone Mass Loss. Endocrinology 2008, 149, 1793–1801. [Google Scholar] [CrossRef]
- Wang, F.-S.; Lin, C.-L.; Chen, Y.-J.; Wang, C.-J.; Yang, K.D.; Huang, Y.-T.; Sun, Y.-C.; Huang, H.-C. Secreted Frizzled-Related Protein 1 Modulates Glucocorticoid Attenuation of Osteogenic Activities and Bone Mass. Endocrinology 2005, 146, 2415–2423. [Google Scholar] [CrossRef] [PubMed]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An Overview of MicroRNAs: Biology, Functions, Therapeutics, and Analysis Methods. J. Cell. Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef] [PubMed]
- Clayton, S.A.; Jones, S.W.; Kurowska-Stolarska, M.; Clark, A.R. The Role of MicroRNAs in Glucocorticoid Action. J. Biol. Chem. 2018, 293, 1865–1874. [Google Scholar] [CrossRef]
- Kawa, M.P.; Sobuś, A.; Litwińska, Z.; Osowicz-Korolonek, L.; Cymbaluk-Płoska, A.; Stecewicz, I.; Zagrodnik, E.; Romanowska, H.; Walczak, M.; Syrenicz, A.; et al. Expression of Selected Angiogenesis-Related Small MicroRNAs in Patients with Abnormally Increased Secretion of Glucocorticoids. Endokrynol. Pol. 2019, 70, 489–495. [Google Scholar] [CrossRef]
- Ren, J.; Gu, C.; Yang, Y.; Xue, J.; Sun, Y.; Jian, F.; Chen, D.; Bian, L.; Sun, Q. TSP-1 Is Downregulated and Inversely Correlates with MiR-449c Expression in Cushing’s Disease. J. Cell. Mol. Med. 2019, 23, 4097–4110. [Google Scholar] [CrossRef] [PubMed]
- Belaya, Z.; Khandaeva, P.; Nonn, L.; Nikitin, A.; Solodovnikov, A.; Sitkin, I.; Grigoriev, A.; Pikunov, M.; Lapshina, A.; Rozhinskaya, L.; et al. Circulating Plasma MicroRNA to Differentiate Cushing’s Disease from Ectopic ACTH Syndrome. Front. Endocrinol. 2020, 11, 331. [Google Scholar] [CrossRef]
- Chantzichristos, D.; Svensson, P.-A.; Garner, T.; Glad, C.A.; Walker, B.R.; Bergthorsdottir, R.; Ragnarsson, O.; Trimpou, P.; Stimson, R.H.; Borresen, S.W.; et al. Identification of Human Glucocorticoid Response Markers Using Integrated Multi-Omic Analysis from a Randomized Crossover Trial. eLife 2021, 10, e62236. [Google Scholar] [CrossRef]
- Landry, J.J.M.; Pyl, P.T.; Rausch, T.; Zichner, T.; Tekkedil, M.M.; Stütz, A.M.; Jauch, A.; Aiyar, R.S.; Pau, G.; Delhomme, N.; et al. The Genomic and Transcriptomic Landscape of a HeLa Cell Line. G3 Genes Genomes Genet. 2013, 3, 1213–1224. [Google Scholar] [CrossRef]
- Masters, J.R. HeLa Cells 50 Years on: The Good, the Bad and the Ugly. Nat. Rev. Cancer 2002, 2, 315–319. [Google Scholar] [CrossRef]
- Vrzal, R.; Ulrichová, J.; Dvorák, Z.; Pávek, P. Glucocorticoid Receptor Functions in HeLa Cells Are Perturbed by 2,3,8,9-Tetrachlorodibenzo-p-Dioxin (TCDD). Drug Metab. Lett. 2007, 1, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Shimojo, M.; Hiroi, N.; Yakushiji, F.; Ueshiba, H.; Yamaguchi, N.; Miyachi, Y. Differences in Down-Regulation of Glucocorticoid Receptor MRNA by Cortisol, Prednisolone and Dexamethasone in HeLa Cells. Endocr. J. 1995, 42, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Wagner, B.J. Expression of the Functional Glucocorticoid Receptor in Mouse and Human Lens Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2041–2046. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dvořák, Z.; Vrzal, R.; Maurel, P.; Ulrichová, J. Differential Effects of Selected Natural Compounds with Anti-Inflammatory Activity on the Glucocorticoid Receptor and NF-ΚB in HeLa Cells. Chem. Biol. Interact. 2006, 159, 117–128. [Google Scholar] [CrossRef]
- Molina, M.L.; Guerrero, J.; Cidlowski, J.A.; Gatica, H.; Goecke, A. LPS Regulates the Expression of Glucocorticoid Receptor α and β Isoforms and Induces a Selective Glucocorticoid Resistance in Vitro. J. Inflamm. 2017, 14, 22. [Google Scholar] [CrossRef]
- Kino, T.; Manoli, I.; Kelkar, S.; Wang, Y.; Su, Y.A.; Chrousos, G.P. Glucocorticoid Receptor (GR) Beta Has Intrinsic, GRalpha-Independent Transcriptional Activity. Biochem. Biophys. Res. Commun. 2009, 381, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Li, C.-T.; Xiong, L.; Ning, Z.; Leung, F.; Peng, S.; Lu, W.W. MiR-375-3p Negatively Regulates Osteogenesis by Targeting and Decreasing the Expression Levels of LRP5 and β-Catenin. PLoS ONE 2017, 12, e0171281. [Google Scholar] [CrossRef]
- Zhao, M.-C.; Zhang, M.-M.; Li, T.; Tao, Z.-H.; Du, Y.-Q.; Wang, L.-P.; Zhang, J.; Wang, B.-Y.; Hu, X.-C. MiR-566 Protects the Malignant Progression of Breast Cancer by Negatively Regulating WNT6. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 6185–6194. [Google Scholar] [CrossRef] [PubMed]
- Xi, M.; Cheng, L.; Hua, W.; Zhou, Y.-L.; Gao, Q.-L.; Yang, J.-X.; Qi, S.-Y. MicroRNA-95-3p Promoted the Development of Prostatic Cancer via Regulating DKK3 and Activating Wnt/β-Catenin Pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jiaqi, C.; Zhaoying, C.; Huimin, C. MicroRNA-506-3p Regulates Neural Stem Cell Proliferation and Differentiation through Targeting TCF3. Gene 2016, 593, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Peng, B.; Zhu, X.; Wang, P.; Sun, K.; Lei, X.; He, H.; Tian, Y.; Mo, S.; Zhang, R.; et al. MiR-210-3p Inhibits Osteogenic Differentiation and Promotes Adipogenic Differentiation Correlated with Wnt Signaling in ERα-deficient RBMSCs. J. Cell. Physiol. 2019, 234, 23475–23484. [Google Scholar] [CrossRef]
- Hu, B.; Li, Y.; Wang, M.; Zhu, Y.; Zhou, Y.; Sui, B.; Tan, Y.; Ning, Y.; Wang, J.; He, J.; et al. Functional Reconstruction of Critical-Sized Load-Bearing Bone Defects Using a Sclerostin-Targeting MiR-210-3p-Based Construct to Enhance Osteogenic Activity. Acta Biomater. 2018, 76, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Xiang, H.; Peng, Y.; Peng, J.; Jiang, S. Mature MiR-183, Negatively Regulated by Transcription Factor GATA3, Promotes 3T3-L1 Adipogenesis through Inhibition of the Canonical Wnt/β-Catenin Signaling Pathway by Targeting LRP6. Cell. Signal. 2014, 26, 1155–1165. [Google Scholar] [CrossRef]
- Ueno, K.; Hirata, H.; Shahryari, V.; Deng, G.; Tanaka, Y.; Tabatabai, Z.L.; Hinoda, Y.; Dahiya, R. MicroRNA-183 Is an Oncogene Targeting Dkk-3 and SMAD4 in Prostate Cancer. Br. J. Cancer 2013, 108, 1659–1667. [Google Scholar] [CrossRef]
- Zhu, N.; Huang, K.; Liu, Y.; Zhang, H.; Lin, E.; Zeng, Y.; Li, H.; Xu, Y.; Cai, B.; Yuan, Y.; et al. MiR-195-5p Regulates Hair Follicle Inductivity of Dermal Papilla Cells by Suppressing Wnt/β -Catenin Activation. BioMed Res. Int. 2018, 2018, 1–13. [Google Scholar] [CrossRef]
- Chang, M.; Lin, H.; Fu, H.; Wang, B.; Han, G.; Fan, M. MicroRNA-195-5p Regulates Osteogenic Differentiation of Periodontal Ligament Cells Under Mechanical Loading. J. Cell. Physiol. 2017, 232, 3762–3774. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Li, Z.; Xue, H.; Ma, T.; Ren, C.; Li, M.; Lu, Y.; Sun, H.; Zhang, K. MiR-26a Promotes Fracture Healing of Nonunion Rats Possibly by Targeting SOSTDC1 and Further Activating Wnt/β-Catenin Signaling Pathway. Mol. Cell. Biochem. 2019, 460, 165–173. [Google Scholar] [CrossRef]
- Li, S.; Hu, C.; Li, J.; Liu, L.; Jing, W.; Tang, W.; Tian, W.; Long, J. Effect of MiR-26a-5p on the Wnt/Ca2+ Pathway and Osteogenic Differentiation of Mouse Adipose-Derived Mesenchymal Stem Cells. Calcif. Tissue Int. 2016, 99, 174–186. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Liao, L.; Shuai, Y.; Jing, H.; Liu, S.; Zhou, H.; Liu, Y.; Jin, Y. MiR-26a Functions Oppositely in Osteogenic Differentiation of BMSCs and ADSCs Depending on Distinct Activation and Roles of Wnt and BMP Signaling Pathway. Cell Death Dis. 2015, 6, e1851. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Guan, X.; Guo, F.; Zhou, J.; Chang, A.; Sun, B.; Cai, Y.; Ma, Z.; Dai, C.; Li, X.; et al. MiR-30e Reciprocally Regulates the Differentiation of Adipocytes and Osteoblasts by Directly Targeting Low-Density Lipoprotein Receptor-Related Protein 6. Cell Death Dis. 2013, 4, e845. [Google Scholar] [CrossRef] [PubMed]
- Tomé, M.; López-Romero, P.; Albo, C.; Sepúlveda, J.C.; Fernández-Gutiérrez, B.; Dopazo, A.; Bernad, A.; González, M.A. MiR-335 Orchestrates Cell Proliferation, Migration and Differentiation in Human Mesenchymal Stem Cells. Cell Death Differ. 2011, 18, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Jiménez, C.; Carrascoso, I.; Barrero, J.; Izquierdo, J.M. Identification of a Set of MiRNAs Differentially Expressed in Transiently TIA-Depleted HeLa Cells by Genome-Wide Profiling. BMC Mol. Biol. 2013, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Deng, L.; Su, D.; Xiao, J.; Ge, D.; Bao, Y.; Jing, H. Identification of Crucial MicroRNAs and Genes in Hypoxia-Induced Human Lung Adenocarcinoma Cells. Onco. Targets 2016, 9, 4605–4616. [Google Scholar] [CrossRef]
- Adusumilli, L.; Facchinello, N.; Teh, C.; Busolin, G.; Le, M.T.; Yang, H.; Beffagna, G.; Campanaro, S.; Tam, W.L.; Argenton, F.; et al. MiR-7 Controls the Dopaminergic/Oligodendroglial Fate through Wnt/β-Catenin Signaling Regulation. Cells 2020, 9, 711. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Wang, L.; Zhou, J.; Pang, P.; Cai, S.; Li, J.; Mei, S.; Li, F. The Transcription Factor Ccaat/Enhancer Binding Protein β (C/EBPβ) and MiR-27a Regulate the Expression of Porcine Dickkopf2 (DKK2). Sci. Rep. 2015, 5, 17972. [Google Scholar] [CrossRef][Green Version]
- Wang, Q.; Cai, J.; Cai, X.; Chen, L. MiR-346 Regulates Osteogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells by Targeting the Wnt/β-Catenin Pathway. PLoS ONE 2013, 8, e72266. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Willems, E.; Singh, A.; Ong, I.M.; Verma, A.K. Ultraviolet Radiation-Induced Differential MicroRNA Expression in the Skin of Hairless SKH1 Mice, a Widely Used Mouse Model for Dermatology Research. Oncotarget 2016, 7, 84924–84937. [Google Scholar] [CrossRef]
- Tömböl, Z.; Szabó, P.M.; Molnár, V.; Wiener, Z.; Tölgyesi, G.; Horányi, J.; Riesz, P.; Reismann, P.; Patócs, A.; Likó, I.; et al. Integrative Molecular Bioinformatics Study of Human Adrenocortical Tumors: MicroRNA, Tissue-Specific Target Prediction, and Pathway Analysis. Endocr. Relat. Cancer 2009, 16, 895–906. [Google Scholar] [CrossRef] [PubMed]
- GEO Accession Viewer. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE64826 (accessed on 12 October 2021).
- Lekva, T.; Ueland, T.; Bøyum, H.; Evang, J.A.; Godang, K.; Bollerslev, J. TXNIP Is Highly Regulated in Bone Biopsies from Patients with Endogenous Cushing’s Syndrome and Related to Bone Turnover. Eur. J. Endocrinol. 2012, 166, 1039–1048. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Grundberg, E.; Adoue, V.; Kwan, T.; Ge, B.; Duan, Q.L.; Lam, K.C.L.; Koka, V.; Kindmark, A.; Weiss, S.T.; Tantisira, K.; et al. Global Analysis of the Impact of Environmental Perturbation on Cis-Regulation of Gene Expression. PLoS Genet. 2011, 7, e1001279. [Google Scholar] [CrossRef]
- Grundberg, E.; Brändström, H.; Lam, K.C.L.; Gurd, S.; Ge, B.; Harmsen, E.; Kindmark, A.; Ljunggren, O.; Mallmin, H.; Nilsson, O.; et al. Systematic Assessment of the Human Osteoblast Transcriptome in Resting and Induced Primary Cells. Physiol. Genom. 2008, 33, 301–311. [Google Scholar] [CrossRef][Green Version]
- Hardy, R.; Juarez, M.; Naylor, A.; Tu, J.; Rabbitt, E.H.; Filer, A.; Stewart, P.M.; Buckley, C.D.; Raza, K.; Cooper, M.S. Synovial DKK1 Expression Is Regulated by Local Glucocorticoid Metabolism in Inflammatory Arthritis. Arthritis Res. 2012, 14, R226. [Google Scholar] [CrossRef]
- Pickering, R.T.; Lee, M.-J.; Karastergiou, K.; Gower, A.; Fried, S.K. Depot Dependent Effects of Dexamethasone on Gene Expression in Human Omental and Abdominal Subcutaneous Adipose Tissues from Obese Women. PLoS ONE 2016, 11, e0167337. [Google Scholar] [CrossRef] [PubMed]
- Stojadinovic, O.; Lee, B.; Vouthounis, C.; Vukelic, S.; Pastar, I.; Blumenberg, M.; Brem, H.; Tomic-Canic, M. Novel Genomic Effects of Glucocorticoids in Epidermal Keratinocytes. J. Biol. Chem. 2007, 282, 4021–4034. [Google Scholar] [CrossRef]
- Murani, E.; Trakooljul, N.; Hadlich, F.; Ponsuksili, S.; Wimmers, K. Transcriptome Responses to Dexamethasone Depending on Dose and Glucocorticoid Receptor Sensitivity in the Liver. Front. Genet. 2019, 10, 559. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Lönnerdal, B. Beta-Catenin/TCF4 Transactivates MiR-30e during Intestinal Cell Differentiation. Cell. Mol. Life Sci. 2010, 67, 2969–2978. [Google Scholar] [CrossRef]
- Smith, S.S.; Dole, N.S.; Franceschetti, T.; Hrdlicka, H.C.; Delany, A.M. MicroRNA-433 Dampens Glucocorticoid Receptor Signaling, Impacting Circadian Rhythm and Osteoblastic Gene Expression. J. Biol. Chem. 2016, 291, 21717–21728. [Google Scholar] [CrossRef] [PubMed]
- Son, G.H.; Chung, S.; Choe, H.K.; Kim, H.-D.; Baik, S.-M.; Lee, H.; Lee, H.-W.; Choi, S.; Sun, W.; Kim, H.; et al. Adrenal Peripheral Clock Controls the Autonomous Circadian Rhythm of Glucocorticoid by Causing Rhythmic Steroid Production. Proc. Natl. Acad. Sci. USA 2008, 105, 20970–20975. [Google Scholar] [CrossRef]
- Ceccato, F.; Boscaro, M. Cushing’s Syndrome: Screening and Diagnosis. High Blood Press. Cardiovasc. Prev. 2016, 23, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Sonino, N.; Fallo, F.; Fava, G.A. Psychosomatic Aspects of Cushing’s Syndrome. Rev. Endocr. Metab. Disord. 2010, 11, 95–104. [Google Scholar] [CrossRef]
- Donn, R.; Berry, A.; Stevens, A.; Farrow, S.; Betts, J.; Stevens, R.; Clayton, C.; Wang, J.; Warnock, L.; Worthington, J.; et al. Use of Gene Expression Profiling to Identify a Novel Glucocorticoid Sensitivity Determining Gene, BMPRII. FASEB J. 2007, 21, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Carter, B.S.; Meng, F.; Thompson, R.C. Glucocorticoid Treatment of Astrocytes Results in Temporally Dynamic Transcriptome Regulation and Astrocyte-Enriched MRNA Changes in Vitro. Physiol. Genom. 2012, 44, 1188–1200. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhou, J.; Lu, F.; Qiu, L.; Xu, L.; Yang, X.; Miao, Y. Downregulation of MicroRNA-183-5p Inhibits the Proliferation and Invasion of Colorectal Cancer Cells by Inactivating the Reticulocalbin-2/Wnt/Β-catenin Signaling Pathway. Mol. Med. Rep. 2019, 10, 59. [Google Scholar] [CrossRef]
- Falzone, L.; Scola, L.; Zanghì, A.; Biondi, A.; Di Cataldo, A.; Libra, M.; Candido, S. Integrated Analysis of Colorectal Cancer MicroRNA Datasets: Identification of MicroRNAs Associated with Tumor Development. Aging 2018, 10, 1000–1014. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Wu, L.; Chen, H.; Huang, Z.; Xu, J.; Zhou, K.; Zhang, Y.; Chen, J.; Xia, J.; Yin, X. Identification of Differentially Expressed MicroRNAs in the Bone Marrow of Osteoporosis Patients. Am. J. Transl. Res. 2019, 11, 2940–2954. [Google Scholar] [PubMed]
- Belaya, Z.E.; Grebennikova, T.A.; Melnichenko, G.A.; Nikitin, A.G.; Solodovnikov, A.G.; Brovkina, O.I.; Grigoriev, A.U.; Rozhinskaya, L.Y.; Dedov, I.I. Effects of Endogenous Hypercortisolism on Bone MRNA and MicroRNA Expression in Humans. Osteoporos. Int. 2018, 29, 211–221. [Google Scholar] [CrossRef]
- Ludwig, N.; Leidinger, P.; Becker, K.; Backes, C.; Fehlmann, T.; Pallasch, C.; Rheinheimer, S.; Meder, B.; Stähler, C.; Meese, E.; et al. Distribution of MiRNA Expression across Human Tissues. Nucleic Acids Res. 2016, 44, 3865–3877. [Google Scholar] [CrossRef]
- Sevignani, C.; Calin, G.A.; Siracusa, L.D.; Croce, C.M. Mammalian MicroRNAs: A Small World for Fine-Tuning Gene Expression. Mamm. Genome 2006, 17, 189–202. [Google Scholar] [CrossRef]
- Peláez, N.; Carthew, R.W. Biological Robustness and the Role of MicroRNAs. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2012; Volume 99, pp. 237–255. ISBN 9780123870384. [Google Scholar]
- Murani, E.; Trakooljul, N.; Hadlich, F.; Ponsuksili, S.; Wimmers, K. Brain Transcriptome Responses to Dexamethasone Depending on Dose and Sex Reveal Factors Contributing to Sex-Specific Vulnerability to Stress-Induced Disorders. Neuroendocrinology 2021, 14, 6500. [Google Scholar] [CrossRef]
- About GEO2R—GEO—NCBI. Available online: https://www.ncbi.nlm.nih.gov/geo/info/geo2r.html (accessed on 18 October 2021).
- Butz, H.; Likó, I.; Czirják, S.; Igaz, P.; Korbonits, M.; Rácz, K.; Patócs, A. MicroRNA Profile Indicates Downregulation of the TGFβ Pathway in Sporadic Non-Functioning Pituitary Adenomas. Pituitary 2011, 14, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, I.S.; Kostoulas, N.; Vergoulis, T.; Georgakilas, G.; Reczko, M.; Maragkakis, M.; Paraskevopoulou, M.D.; Prionidis, K.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA MiRPath v.2.0: Investigating the Combinatorial Effect of MicroRNAs in Pathways. Nucleic Acids Res. 2012, 40, W498–W504. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| miRNA Name | log2FC | p-Value | Connection with Wnt-Signaling | |
|---|---|---|---|---|
| Cortisol Producing Adenoma vs. Normal Adrenal Tissue | Targeted Gene(s) | References | ||
| hsa-miR-375-3p | −3.18283 | 0.002756 | LRP5 and β-catenin | [30] |
| hsa-miR-566 | −2.0934 | 0.012993 | WNT6 | [31] |
| hsa-miR-95-3p | 0.435871 | 0.014638 | DKK3 | [32] |
| hsa-miR-506-3p | 1.96571 | 0.021431 | TCF3 | [33] |
| hsa-miR-210-3p | −1.18389 | 0.038611 | WNT7b, FZD5, Sclerostin | [34,35] |
| Dexamethasone Treated HeLa Cells vs. Control | ||||
| hsa-miR-183-5p | 0.203726 | 0.037677 | LRP6, Dkk-3 | [36,37] |
| hsa-miR-195-5p | 1.355332 | 0.043846 | LRP6, WNT3A | [38,39] |
| hsa-miR-26a-5p | 1.321971 | 0.020827 | SOSTDC1 *, Wnt5a, GSK3β | [40,41,42] |
| hsa-miR-30e-3p | −0.86986 | 0.02123 | LRP6 | [43] |
| hsa-miR-335-3p | 1.586908 | 0.045771 | RUNX2 | [44] |
| hsa-miR-572 | 1.58437 | 0.042381 | pathway analysis ** | [45] |
| hsa-miR-622 | 3.287344 | 0.03071 | pathway analysis ** | [46] |
| hsa-miR-95-3p | 2.307738 | 0.040524 | DKK3 | [32] |
| Dexamethasone Treated H295R Cells vs. Control | ||||
| miR-331-5p | 2.14926 | 0.018581 | no literature data available | |
| miR-7-1-3p | −0.25662 | 0.004913 | TCF4, TCF12, TCF7L2 | [47] |
| miR-27a-3p | 0.457272 | 0.010971 | DKK2 | [48] |
| miR-346 | 0.921683 | 0.017707 | GSK3β | [49] |
| miR-376a-3p | 0.581655 | 0.034872 | pathway analysis ** | [50] |
| miR-668-3p | −0.56929 | 0.041363 | no literature data available | |
| GC-Secreting Adrenal Adenoma | Hela Cells | H295R Cells | |||
|---|---|---|---|---|---|
| KEGG Pathway | p-Value | KEGG Pathway | p-Value | KEGG Pathway | p-Value |
| Biotin metabolism | 5.90E-06 | PI3K-Akt signaling pathway | 1.87E-26 | Focal adhesion | 5.71E-09 |
| Axon guidance | 5.90E-06 | Prostate cancer | 5.59E-20 | Wnt signaling pathway | 4.44E-08 |
| Long-term depression | 3.95E-04 | Wnt signaling pathway | 8.18E-14 | Neurotrophin signaling pathway | 4.44E-08 |
| Insulin signaling pathway | 3.23E-03 | mTOR signaling pathway | 5.65E-13 | ErbB signaling pathway | 1.08E-07 |
| Gap junction | 7.57E-03 | Insulin signaling pathway | 1.74E-12 | Gap junction | 1.08E-07 |
| Chronic myeloid leukemia | 8.75E-02 | Focal adhesion | 2.14E-11 | PI3K-Akt signaling pathway | 1.08E-07 |
| Glutamatergic synapse | 1.84E-01 | Ubiquitin mediated proteolysis | 8.16E-11 | GnRH signaling pathway | 1.85E-06 |
| Adipocytokine signaling pathway | 1.84E-01 | TGF-beta signaling pathway | 1.03E-10 | Circadian rhythm | 2.06E-05 |
| Wnt signaling pathway | 2.65E-01 | Pathways in cancer | 5.91E-10 | MAPK signaling pathway | 2.06E-05 |
| Gastric acids secretion | 2.65E-01 | Melanoma | 1.32E-09 | Ubiquitin mediated proteolysis | 3.30E-05 |
| Salmonella infection | 2.91E-01 | p53 signaling pathway | 1.69E-09 | Glioma | 5.05E-05 |
| Neurotrophin signaling pathway | 3.95E-01 | Regulation of actin cytoskeleton | 1.02E-08 | Long-term potentiation | 1.06E-04 |
| MAPK signaling pathway | 5.45E-01 | Glioma | 2.31E-08 | Pathways in cancer | 2.11E-04 |
| Retrograde endocannabinoid signaling | 6.21E-01 | Long-term potentiation | 2.58E-08 | Insulin signaling pathway | 2.58E-04 |
| ErbB signaling pathway | 6.21E-01 | Long-term depression | 2.76E-08 | mTOR signaling pathway | 2.08E-03 |
| TGF-beta signaling pathway | 6.77E-01 | Gap junction | 4.83E-08 | Axon guidance | 2.45E-03 |
| Pathways in cancer | 7.03E-01 | Endometrial cancer | 4.83E-08 | Dopaminergic synapse | 4.01E-03 |
| Acute myeloid leukemia | 9.43E-01 | Dopaminergic synapse | 5.80E-07 | Small cell lung cancer | 5.79E-03 |
| HTLV-I infection | 9.76E-01 | Fatty acid biosynthesis | 5.87E-07 | Lysine degradation | 6.25E-03 |
| GnRH signaling pathway | 1.34E+00 | Neurotrophin signaling pathway | 6.26E-07 | Protein processing in endoplasmic reticulum | 9.83E-03 |
| Shigellosis | 1.48E+00 | MAPK signaling pathway | 8.51E-07 | Prostate cancer | 2.00E-02 |
| Regulation of actin cytoskeleton | 1.86E+00 | Chronic myeloid leukemia | 4.10E-06 | TGF-beta signaling pathway | 2.13E-02 |
| Pancreatic cancer | 2.43E+00 | HIF-1 signaling pathway | 6.81E-06 | Amphetamine addiction | 6.20E-02 |
| Maturity onset diabetes of the young | 3.57E+00 | ErbB signaling pathway | 7.23E-06 | Hedgehog signaling pathway | 9.27E-02 |
| Cholinergic synapse | 4.00E+00 | Small cell lung cancer | 1.07E-05 | Chronic myeloid leukemia | 1.13E-01 |
| mTOR signaling pathway | 4.04E+00 | Non-small cell lung cancer | 4.37E-05 | RNA transport | 1.54E-01 |
| Focal adhesion | 4.79E+00 | Acute myeloid leukemia | 5.51E-05 | Melanogenesis | 2.37E-01 |
| Pancreatic secretion | 4.79E+00 | Prion diseases | 4.79E-04 | Regulation of actin cytoskeleton | 4.01E-01 |
| Non-small cell lung cancer | 5.54E+00 | mRNA surveillance pathway | 6.30E-04 | Renal cell carcinoma | 6.54E-01 |
| Salivary secretion | 7.96E+00 | Transcriptional misregulation in cancer | 1.48E-03 | Glutamatergic synapse | 6.54E-01 |
| Gastric acid secretion | 3.44E-03 | p53 signaling pathway | 9.56E-01 | ||
| Calcium signaling pathway | 6.18E-03 | Gastric acid secretion | 1.39E+00 | ||
| Aldosterone-regulated sodium reabsorption | 8.44E-03 | Biotin metabolism | 1.63E+00 | ||
| Hedgehog signaling pathway | 1.28E-02 | Transcriptional misregulation in cancer | 2.47E+00 | ||
| T cell receptor signaling pathway | 1.83E-02 | Bacterial invasion of epithelial cells | 4.44E+00 | ||
| RNA transport | 2.76E-02 | Fc gamma R-mediated phagocytosis | 7.41E+00 | ||
| Glutamatergic synapse | 3.14E-02 | Fc epsilon RI signaling pathway | 7.97E+00 | ||
| Renal cell carcinoma | 4.60E-02 | ||||
| Oocyte meiosis | 4.81E-02 | ||||
| Cell cycle | 4.89E-02 | ||||
| Biotin metabolism | 6.21E-02 | ||||
| Colorectal cancer | 1.11E-01 | ||||
| Basal cell carcinoma | 2.68E-01 | ||||
| Inositol phosphate metabolism | 2.68E-01 | ||||
| VEGF signaling pathway | 2.68E-01 | ||||
| B cell receptor signaling pathway | 2.68E-01 | ||||
| Pancreatic cancer | 2.68E-01 | ||||
| Cholinergic synapse | 3.50E-01 | ||||
| Melanogenesis | 3.72E-01 | ||||
| Arrhythmogenic right ventricular cardiomyopathy (ARVC) | 1.19E+00 | ||||
| Hypertrophic cardiomyopathy (HCM) | 1.21E+00 | ||||
| Progesterone-mediated oocyte maturation | 1.21E+00 | ||||
| Apoptosis | 1.52E+00 | ||||
| Tight junction | 1.52E+00 | ||||
| Thyroid cancer | 1.52E+00 | ||||
| Viral carcinogenesis | 2.40E+00 | ||||
| Phosphatidylinositol signaling system | 2.41E+00 | ||||
| Bladder cancer | 2.65E+00 | ||||
| Cocaine addiction | 3.14E+00 | ||||
| Fanconi anemia pathway | 3.92E+00 | ||||
| Gene Expression Study | Time-Dex Treatment | Dose-Dex Treatment | All Genes Measured | # of Not Regulated Genes | # of DEGs (p < 0.05) | Ratio of Regulated Genes | # of Upregulated Genes | # of Downregulated Genes |
|---|---|---|---|---|---|---|---|---|
| Adrenal adenoma vs. normal cortex [51] | na | na | 41,078 | 37,522 | 3556 | 0.08 | 1631 | 1925 |
| Human ACC cancer cell line (H295R) control vs. dex [52] | 6 h | 100 nM | 29,153 | 27,886 | 1267 | 0.04 | 585 | 682 |
| Human bone tissue biopsies from CS patients, before vs. 3 months after surgery [53] | na | before and mean 3 months after surgery | 54,675 | 53,417 | 1258 | 0.02 | 608 | 650 |
| Human primary osteoblast cell control vs. dex [54] | 24 h | 100 nM | 22,177 | 16,834 | 5343 | 0.24 | 2683 | 2660 |
| Human primary osteoblast-like cells (HObs) from bone explant control vs. dex [55] | 24 h | 100 nM | 54,675 | 47,866 | 6809 | 0.12 | 199 | 6610 |
| Human primary synovial fibroblasts control or dex * [56] | 24 h | 100 nM | 81 * | 74 * | 7 * | 0.08 * | 6 * | 1 * |
| human primary abdominal subcutaneous adipose cells control vs. dex [57] | 7 days | 1000 nM | 19,741 | 15,465 | 4276 | 0.21 | 2130 | 2146 |
| Human primary abdominal omental adipose cells control vs. dex [57] | 7 days | 1000 nM | 19,741 | 15,954 | 3787 | 0.19 | 2091 | 1696 |
| Human primary epidermal keratinocytes control vs. dex [58] | 24 h | 100 nM | 12,625 | 11,500 | 1125 | 0.08 | 656 | 469 |
| Piglet hippocampus tissue control vs. im. dex treated [59] | 3 h | 60 µg/kg | 16,764 | 14,361 | 2403 | 0.14 | 1156 | 1247 |
| Piglet hypothalamus tissue control vs. im. dex treated [59] | 3 h | 60 µg/kg | 16,764 | 14,103 | 2661 | 0.15 | 1311 | 1350 |
| GC Target Tissue Type’s Gene Expression | # of DEGs (p < 0.05) | Ratio of WNT Pathway Members * | # of Affected BP | # of Affected BP Related to WNT Signaling ** |
|---|---|---|---|---|
| Human bone tissue biopsies from CS patients, before vs. 3 months after surgery [53] | 1423 | 14/151 | 312 | 4 |
| Human primary osteoblast cell control vs. dex [54] | 5343 | 52/151 | 1573 | 11 |
| Human primary osteoblast-like cells (HObs) from bone explant control vs. dex [55] | 7279 | 53/151 | 1164 | 6 |
| Human primary synovial fibroblasts control or dex [56] | 10 | 8/151 | 880 | 41 |
| Human primary abdominal subcutaneous adipose cells control vs. dex [57] | 4276 | 37/151 | 2644 | 16 |
| Human primary abdominal omental adipose cells control vs. dex [57] | 3787 | 32/151 | 2422 | 6 |
| Human primary epidermal keratinocytes control vs. dex [58] | 1130 | 13/151 | 1753 | 7 |
| Piglet hippocampus tissue control vs. im. dex treated [59] | 2403 | 34/151 | 1913 | 10 |
| Piglet hypothalamus tissue control vs. im. dex treated [59] | 2661 | 33/151 | 1850 | 11 |
| Targeting miRNA | Gene Symbol | Gene Title | logFC | p-Value | GSE14922 Probe ID |
|---|---|---|---|---|---|
| miR-375 | SFRP4 | Secreted frizzled related protein 4 | 1.3979 | 0.0077694 | A_23_P215328 |
| miR-506 | APC | APC, WNT signaling pathway regulator | −2.1794 | 0.0278861 | A_23_P70213 |
| miR-506 | NFAT5 | Nuclear factor of activated T-cells 5 | −1.0592 | 0.0343637 | A_23_P359647 |
| miR-375 | CTBP2 | C-terminal binding protein 2 | 0.9465 | 0.037437 | A_23_P63897 |
| Sample Type | Experiment | # of Control | # of Dex Treatment/GC Affected | DataSet [Refs] | Platform |
|---|---|---|---|---|---|
| Human tissue | bone tissue biopsies from CS patients, before vs. 3 months after surgery | 9 | 9 | GSE30159 [53] | Affymetrix Human Genome U133 Plus 2.0 Array |
| Human tissue | adrenal cortex, normal tissue vs. GC secreting adenoma | 4 | 4 | GSE14922 [51] | Agilent-014850 Whole Human Genome Microarray 4 × 44 K G4112F |
| In vitro human cell line | adrenocortical cancer (H295R) cell line, control vs. dex treatment | 3 | 3 | GSE64826 [52] | Affymetrix Human Gene 1.0 ST Array |
| In vitro human primary cells | osteoblast cell, control vs. dex | 6 | 3 | GSE21727 [54] | Illumina humanRef-8 v2.0 expression bead chip |
| In vitro human primary cells | osteoblast-like cells (HObs) from bone explant, control vs. dex treatment | 6 | 3 | GSE10311 [55] | Affymetrix Human Genome U133 Plus 2.0 Array |
| In vitro human primary cells | synovial fibroblasts, control or dex treatment | 6 | 3 | GSE37520 [56] | Applied Biosystems/University of Birmingham Human RT-PCR array (custom made) |
| In vitro human primary cells | abdominal subcutaneous adipose cells, control vs. dex treatment | 3 | 3 | GSE88966 [57] | Affymetrix Human Gene 1.0 ST Array |
| In vitro human primary cells | abdominal omental adipose cells, control vs. dex treatment | 3 | 3 | GSE88966 [57] | Affymetrix Human Gene 1.0 ST Array |
| In vitro human primary cells | epidermal keratinocytes, control vs. dex treatment | 2 | 2 | GSE26487 [58] | Affymetrix Human Genome U95 Version 2 Array |
| German Landrace piglet tissue | 7-week old purebred piglet hippocampus tissue, control vs. im. dex treatment | 20 | 12 | Murani et al., 2021 [74] | TruSeqStranded mRNA sample preparation kit; HiSeq 2500 instrument |
| German Landrace piglet tissue | 7-week old purebred piglet hypothalamus tissue, control vs. im. dex treatment | 20 | 12 | Murani et al., 2021 [74] | TruSeqStranded mRNA sample preparation kit; HiSeq 2500 instrument |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Butz, H.; Mészáros, K.; Likó, I.; Patocs, A. Wnt-Signaling Regulated by Glucocorticoid-Induced miRNAs. Int. J. Mol. Sci. 2021, 22, 11778. https://doi.org/10.3390/ijms222111778
Butz H, Mészáros K, Likó I, Patocs A. Wnt-Signaling Regulated by Glucocorticoid-Induced miRNAs. International Journal of Molecular Sciences. 2021; 22(21):11778. https://doi.org/10.3390/ijms222111778
Chicago/Turabian StyleButz, Henriett, Katalin Mészáros, István Likó, and Attila Patocs. 2021. "Wnt-Signaling Regulated by Glucocorticoid-Induced miRNAs" International Journal of Molecular Sciences 22, no. 21: 11778. https://doi.org/10.3390/ijms222111778
APA StyleButz, H., Mészáros, K., Likó, I., & Patocs, A. (2021). Wnt-Signaling Regulated by Glucocorticoid-Induced miRNAs. International Journal of Molecular Sciences, 22(21), 11778. https://doi.org/10.3390/ijms222111778