
Comparison of Selenium Nanoparticles and Sodium Selenite on the Alleviation of Early Atherosclerosis by Inhibiting Endothelial Dysfunction and Inflammation in Apolipoprotein E-Deficient Mice



Abstract
:1. Introduction
2. Results
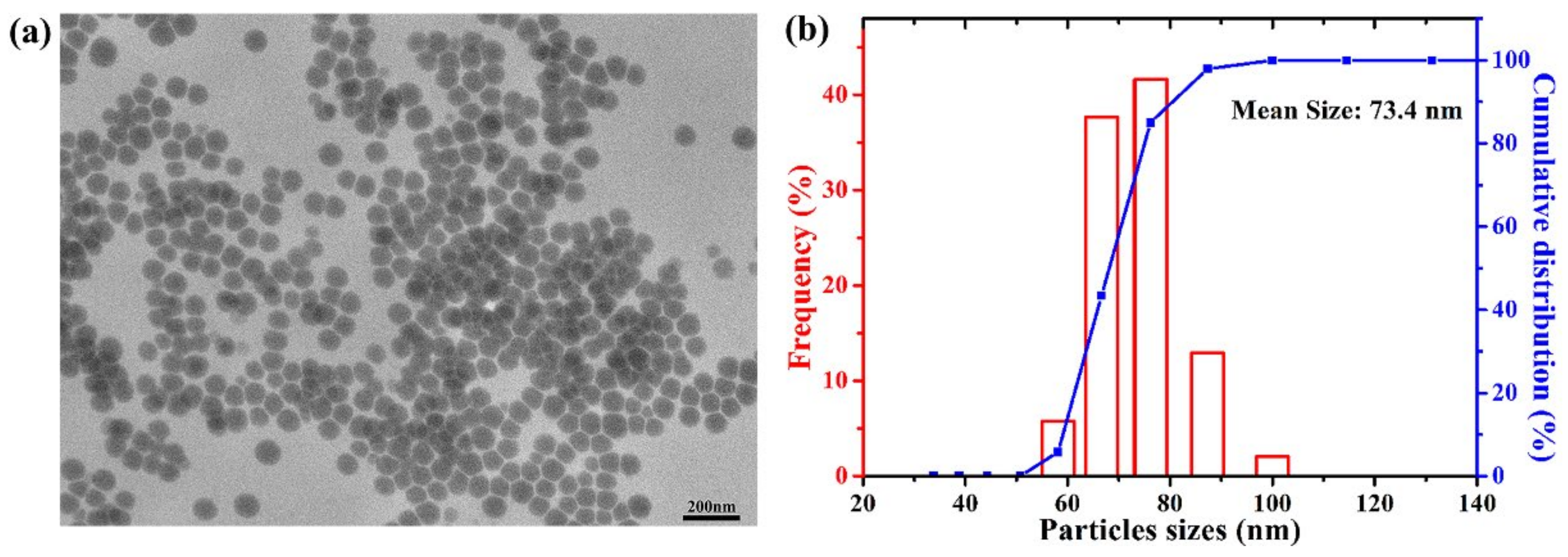
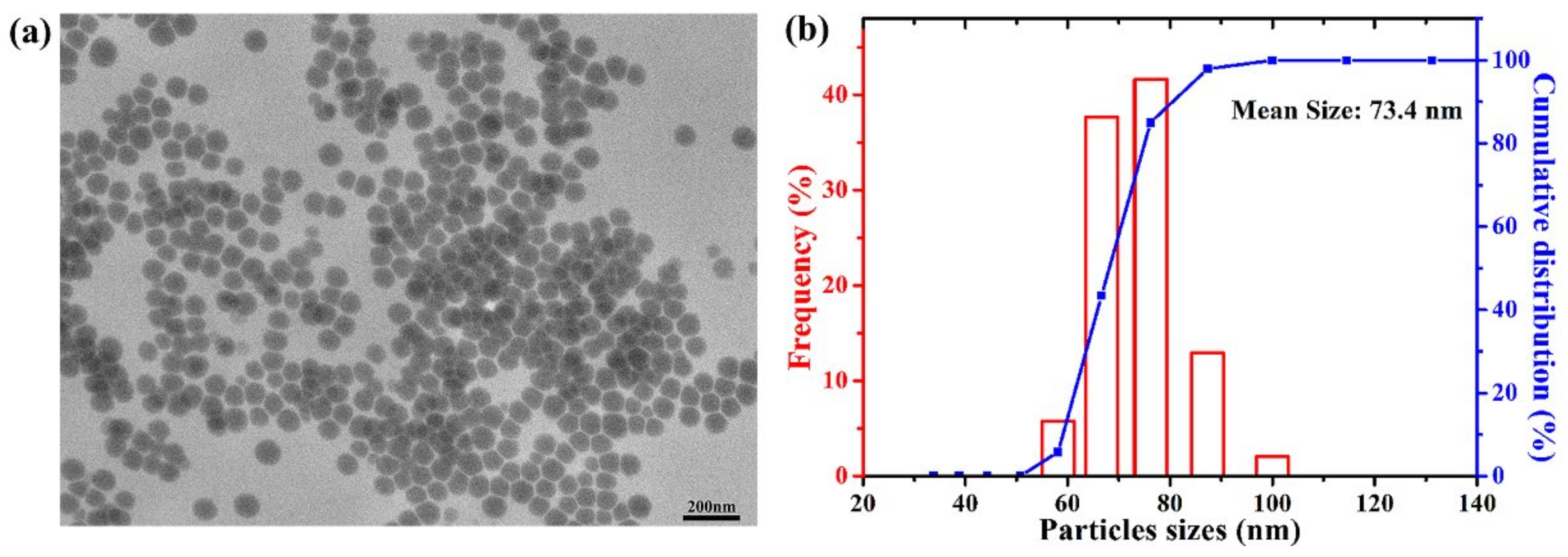
2.1. Characterization of CS-SeNPs
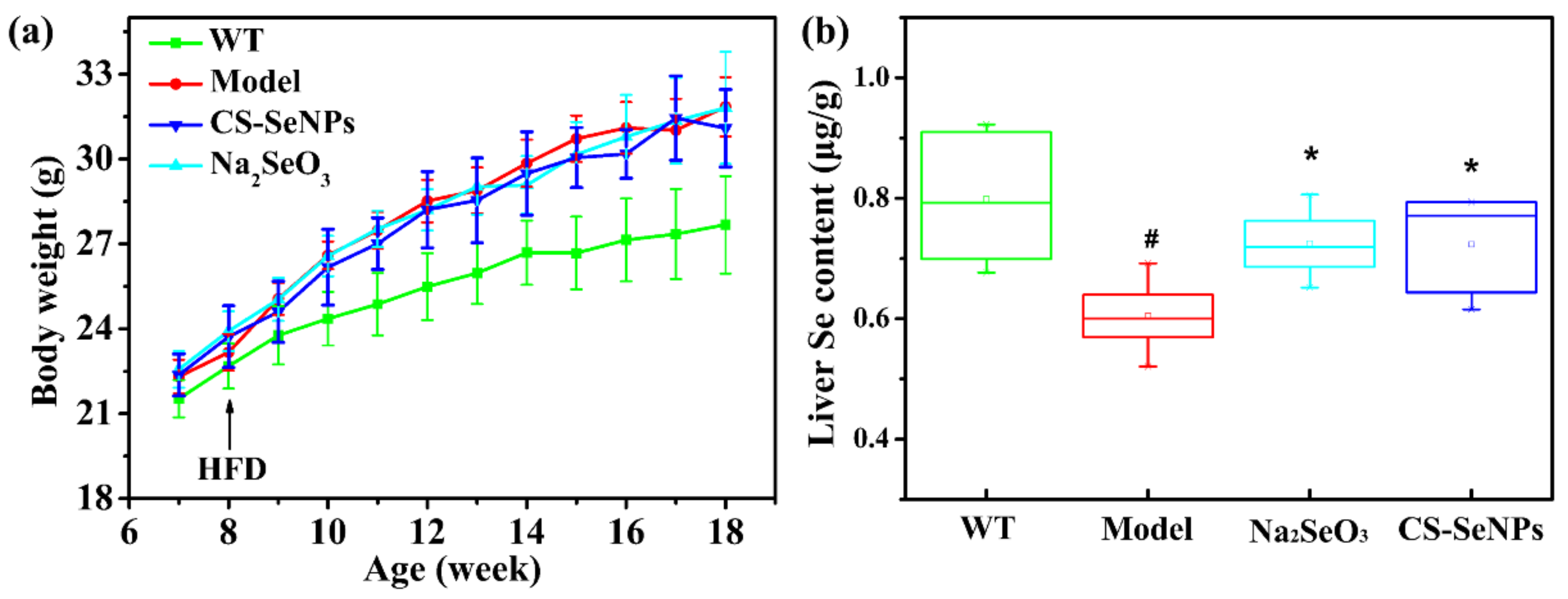
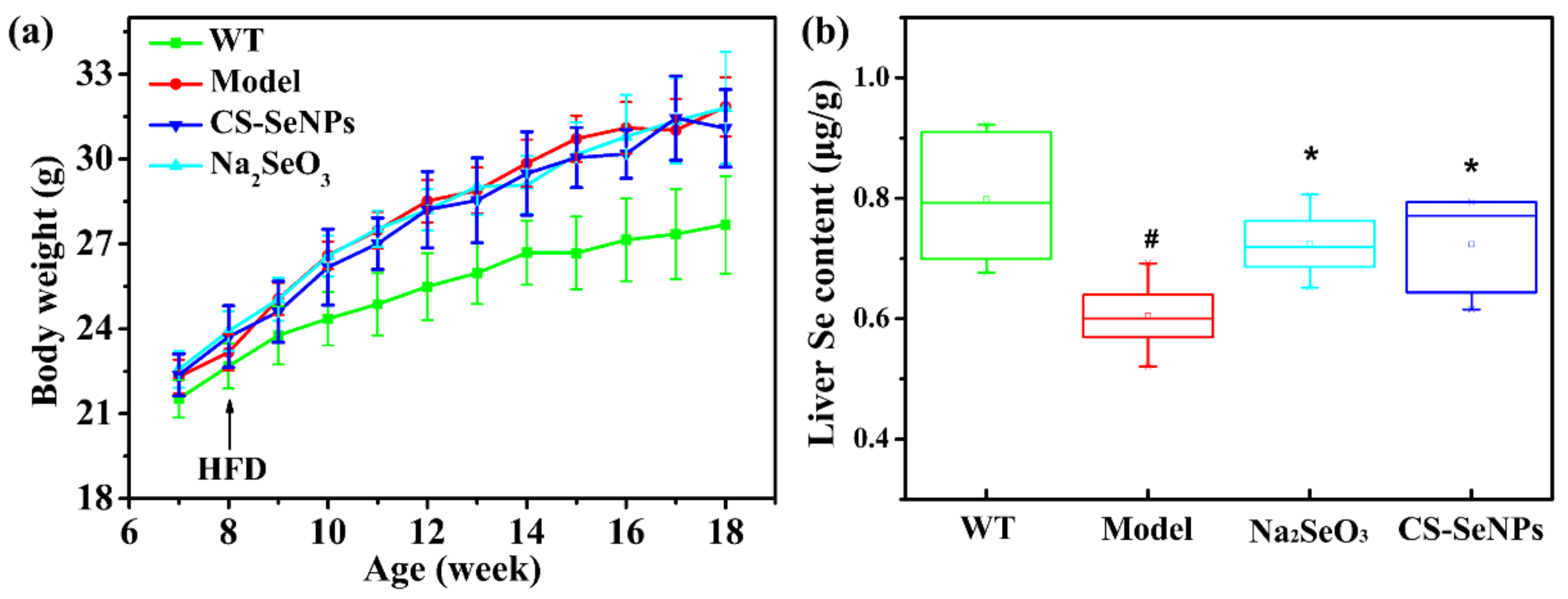
2.2. Both CS-SeNPs and Na2SeO3 Enhanced the Liver Se Contents in ApoE-/- Mice
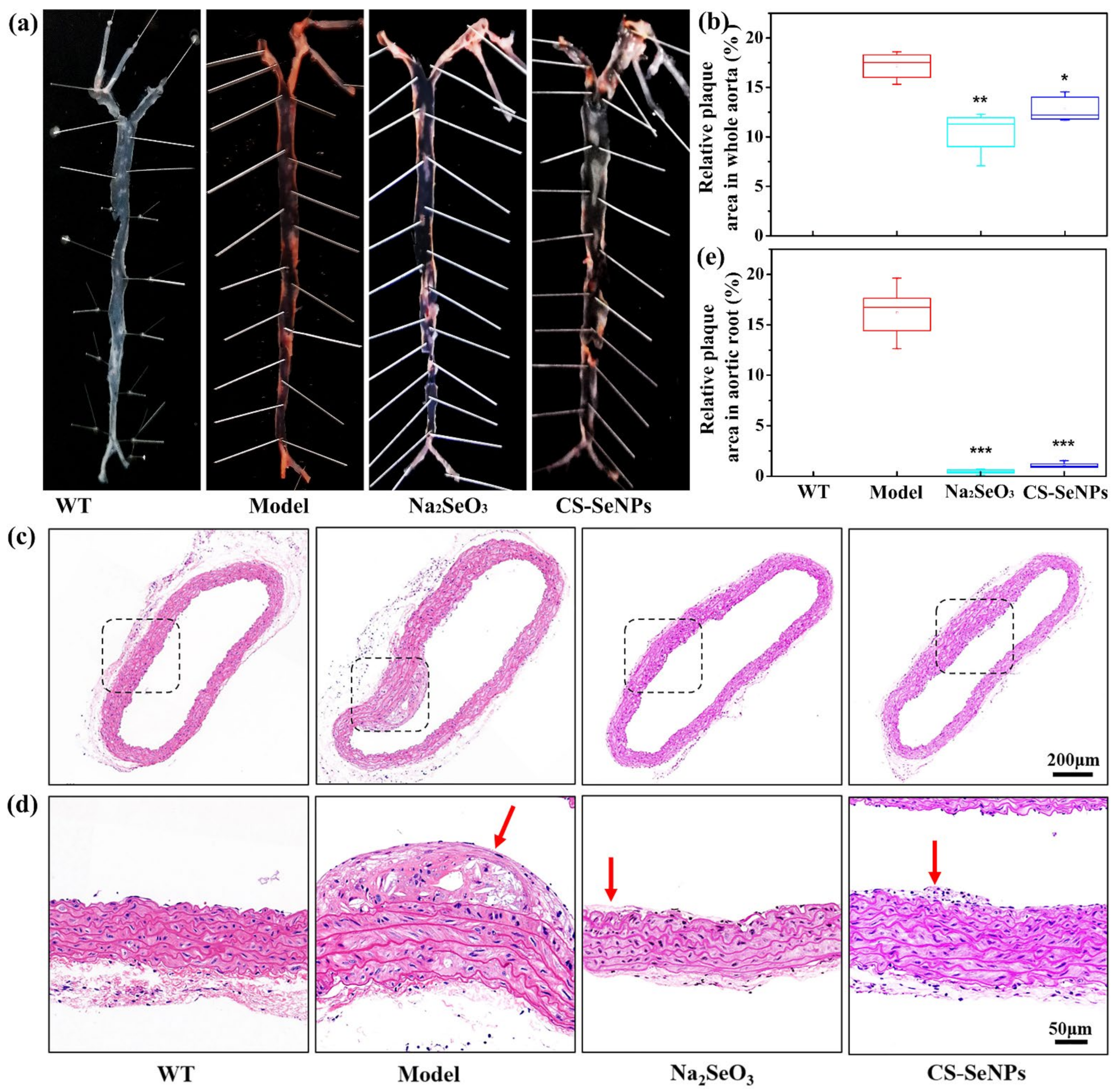
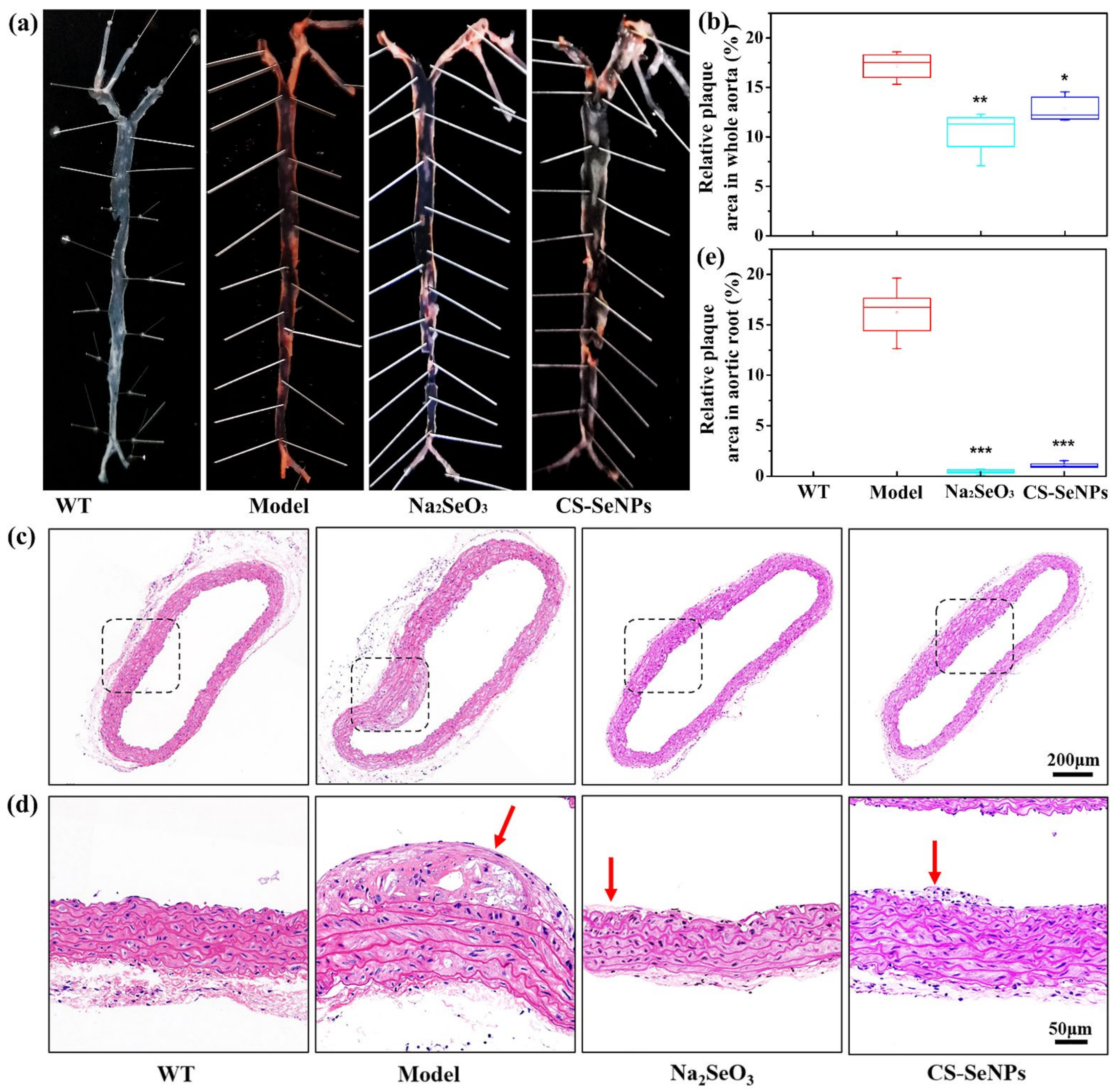
2.3. Both CS-SeNPs and Na2SeO3 Alleviated The Atherosclerotic Lesions in ApoE-/- Mice
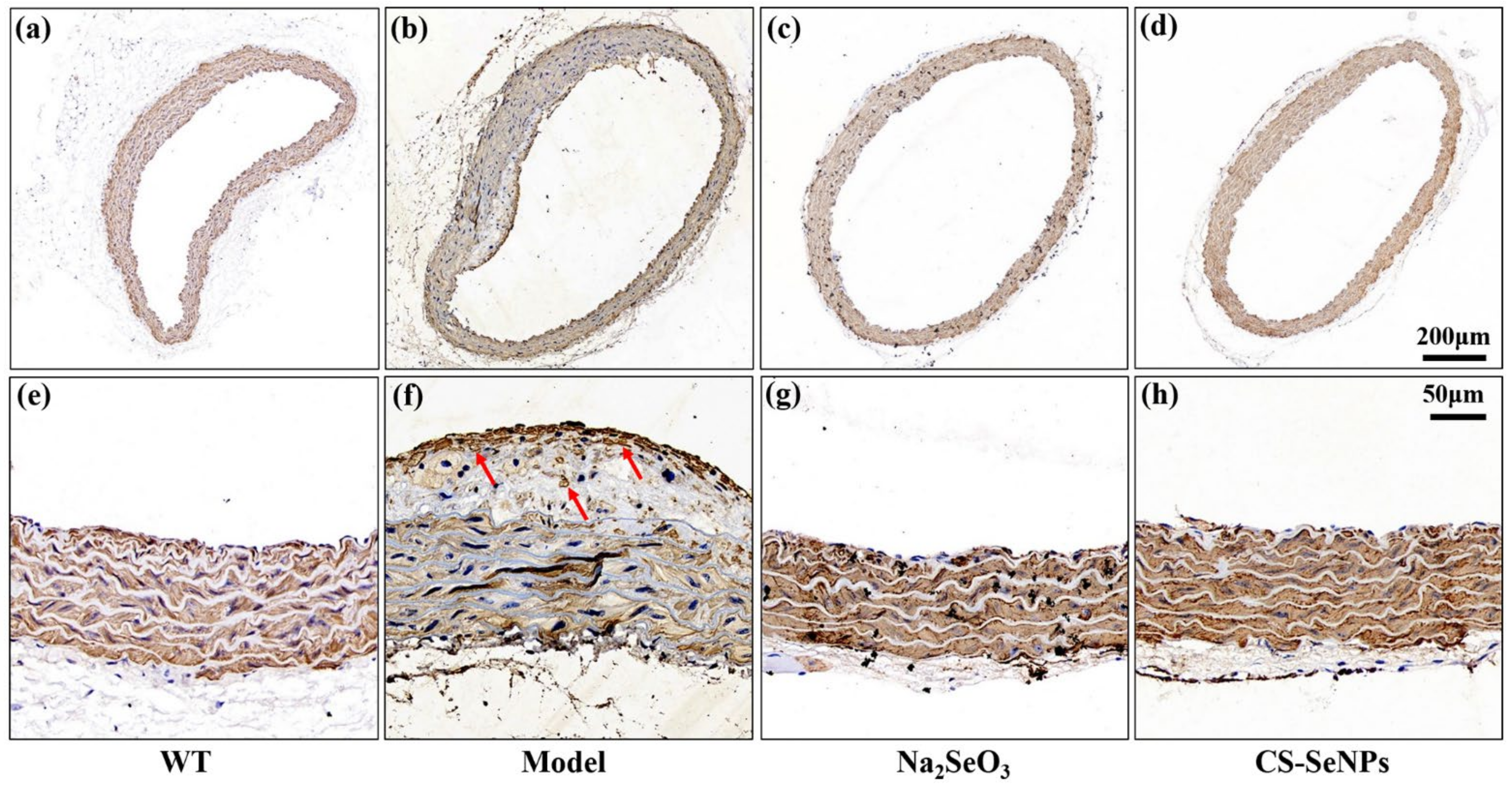
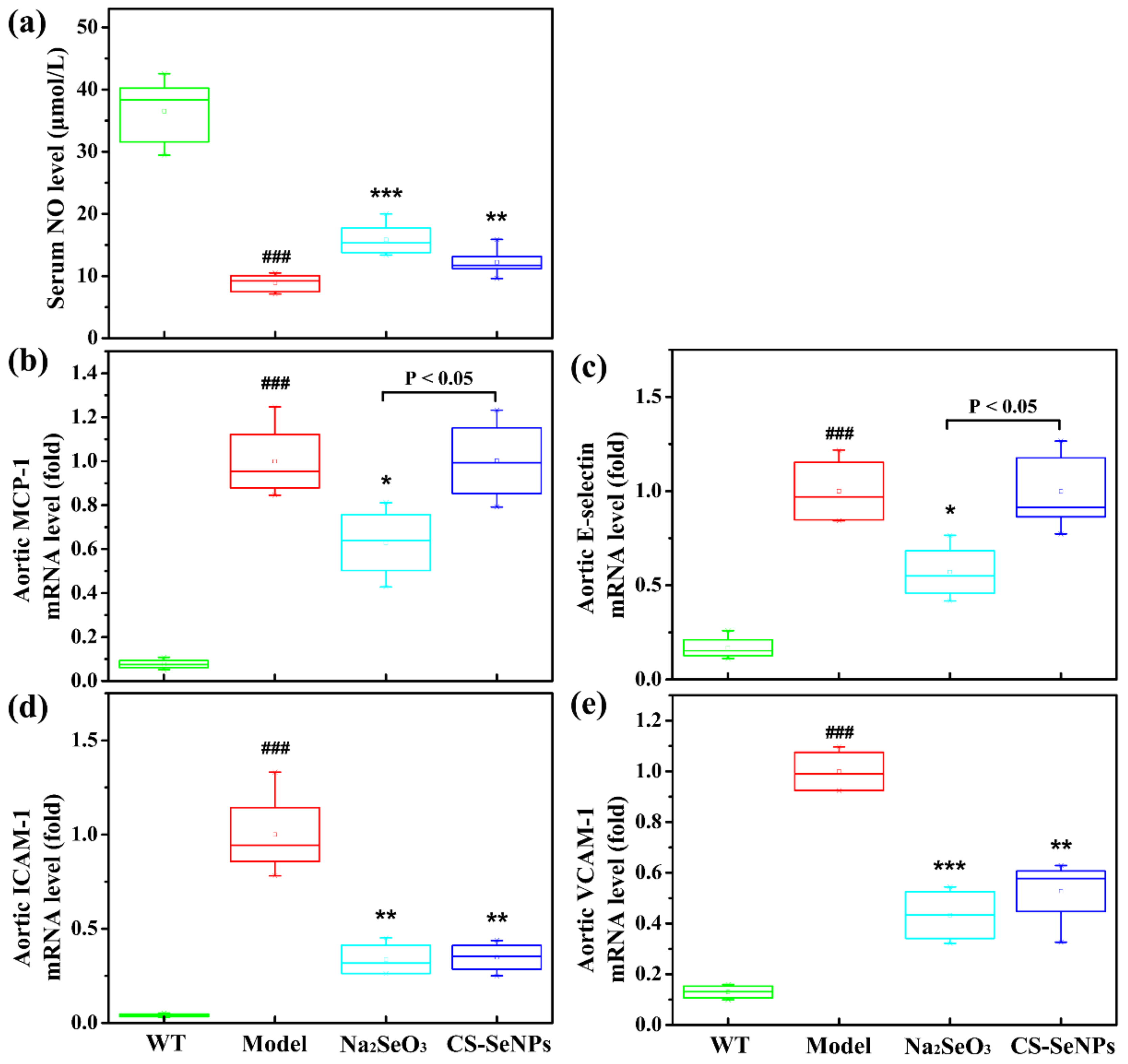
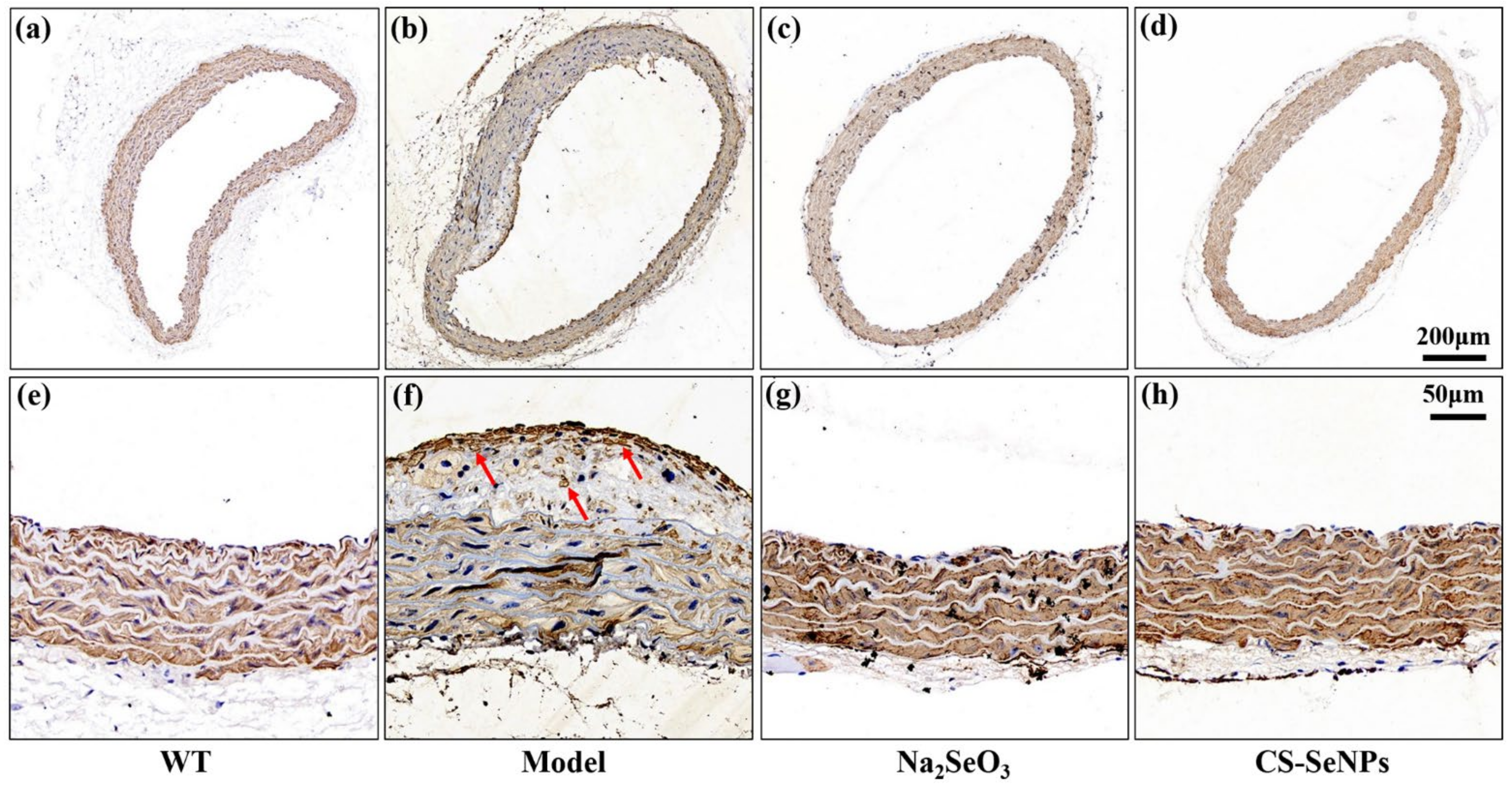
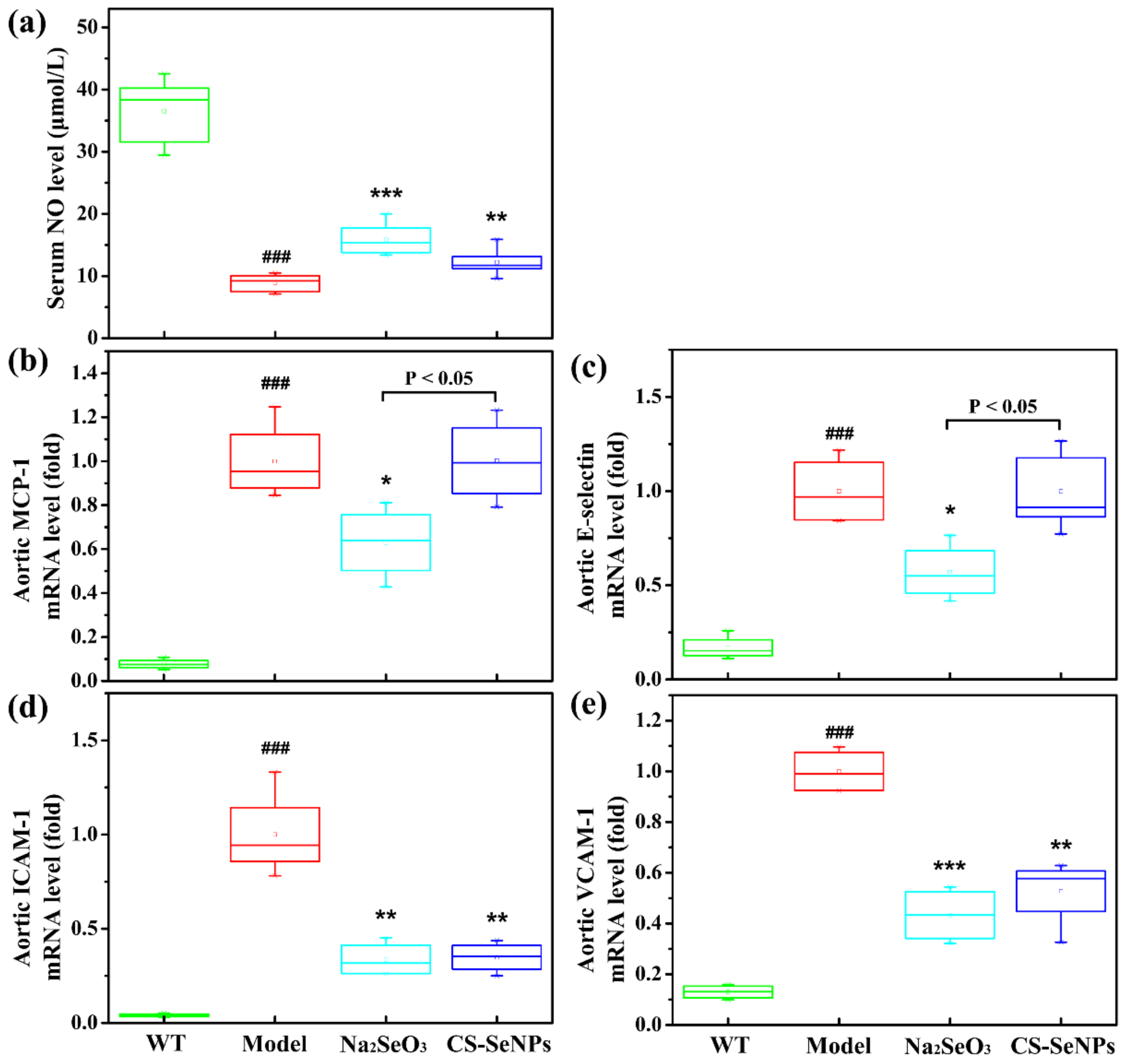
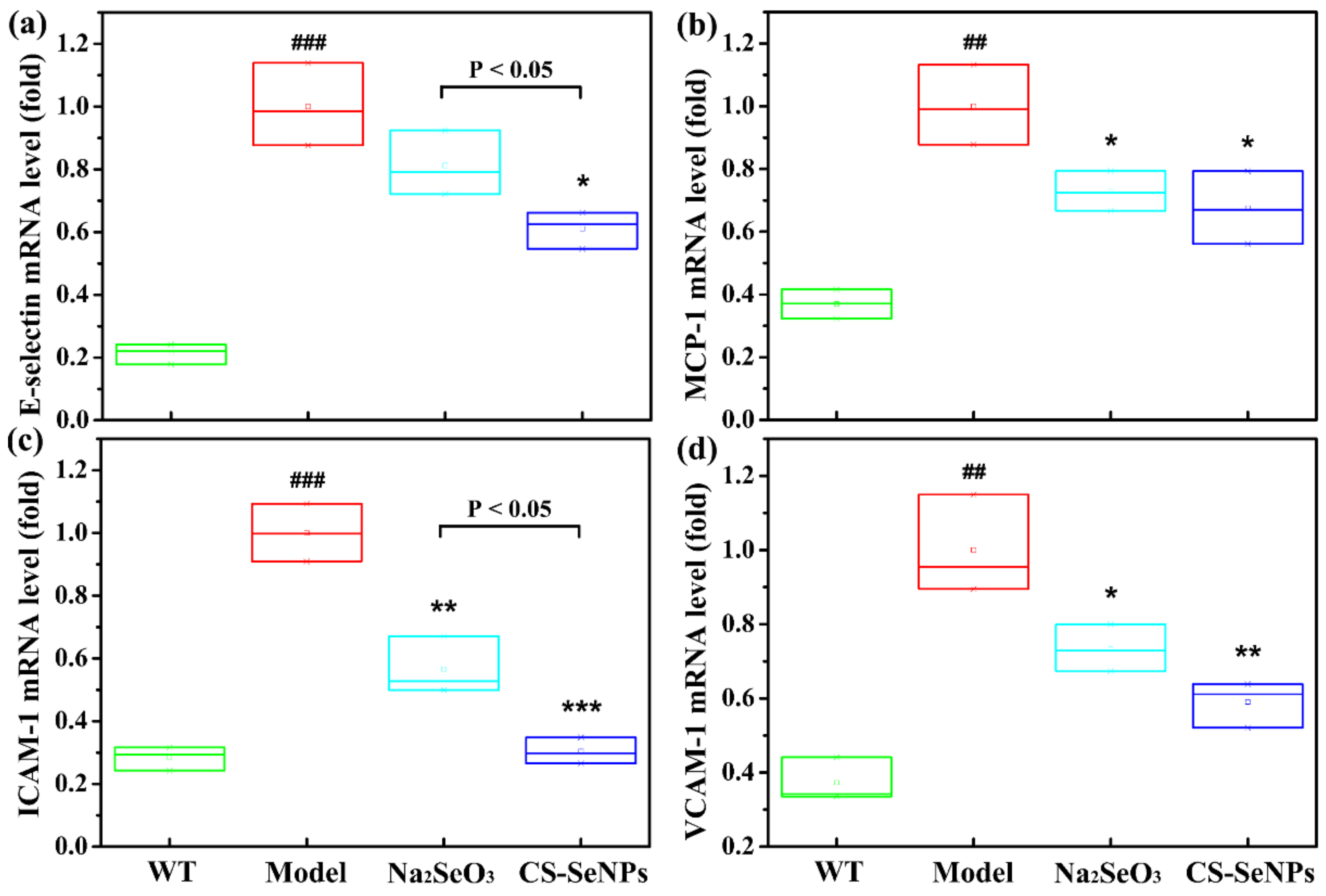
2.4. Both CS-SeNPs and Na2SeO3 Alleviated Vascular Endothelial Dysfunction in ApoE-/- Mice
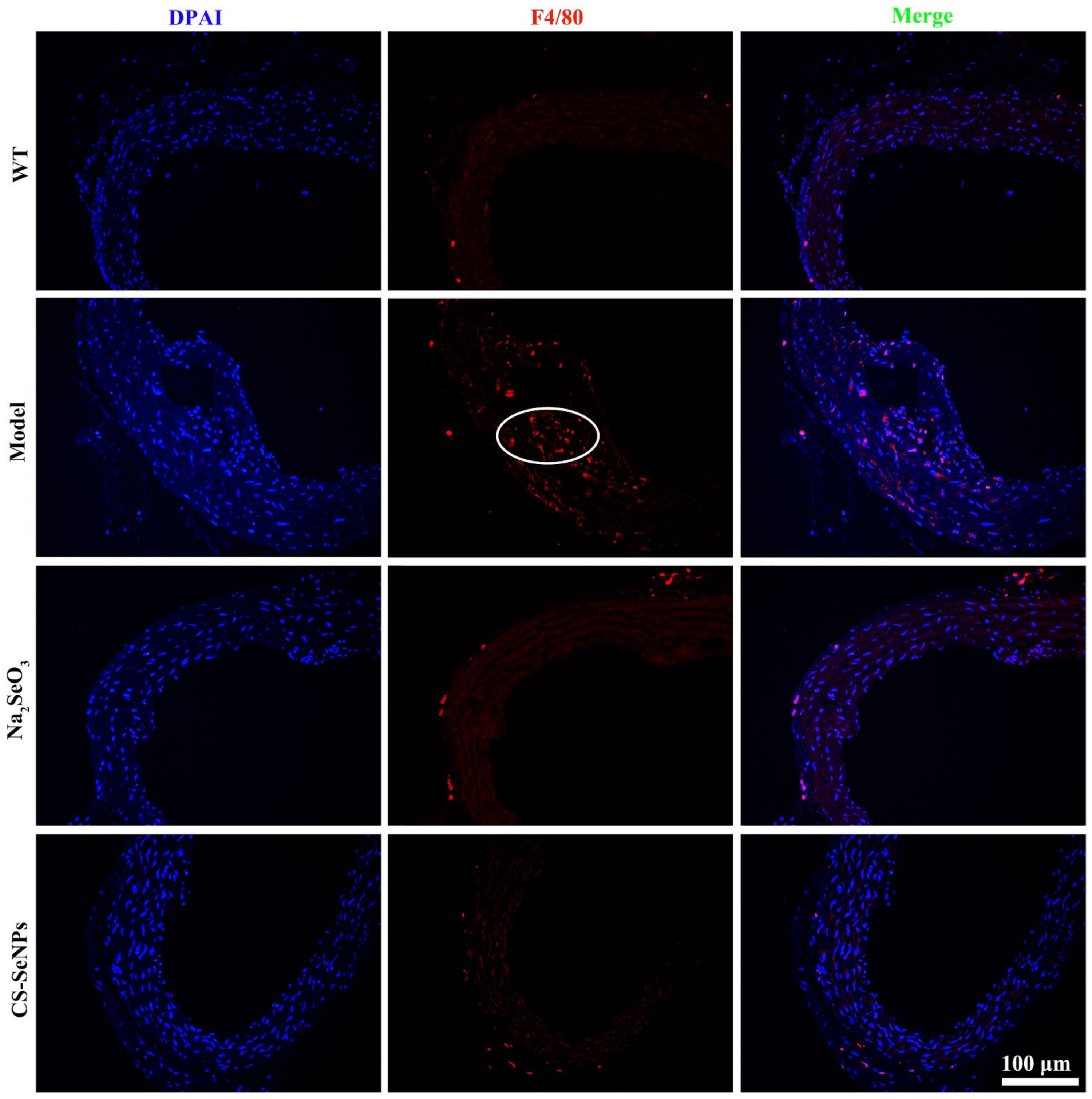
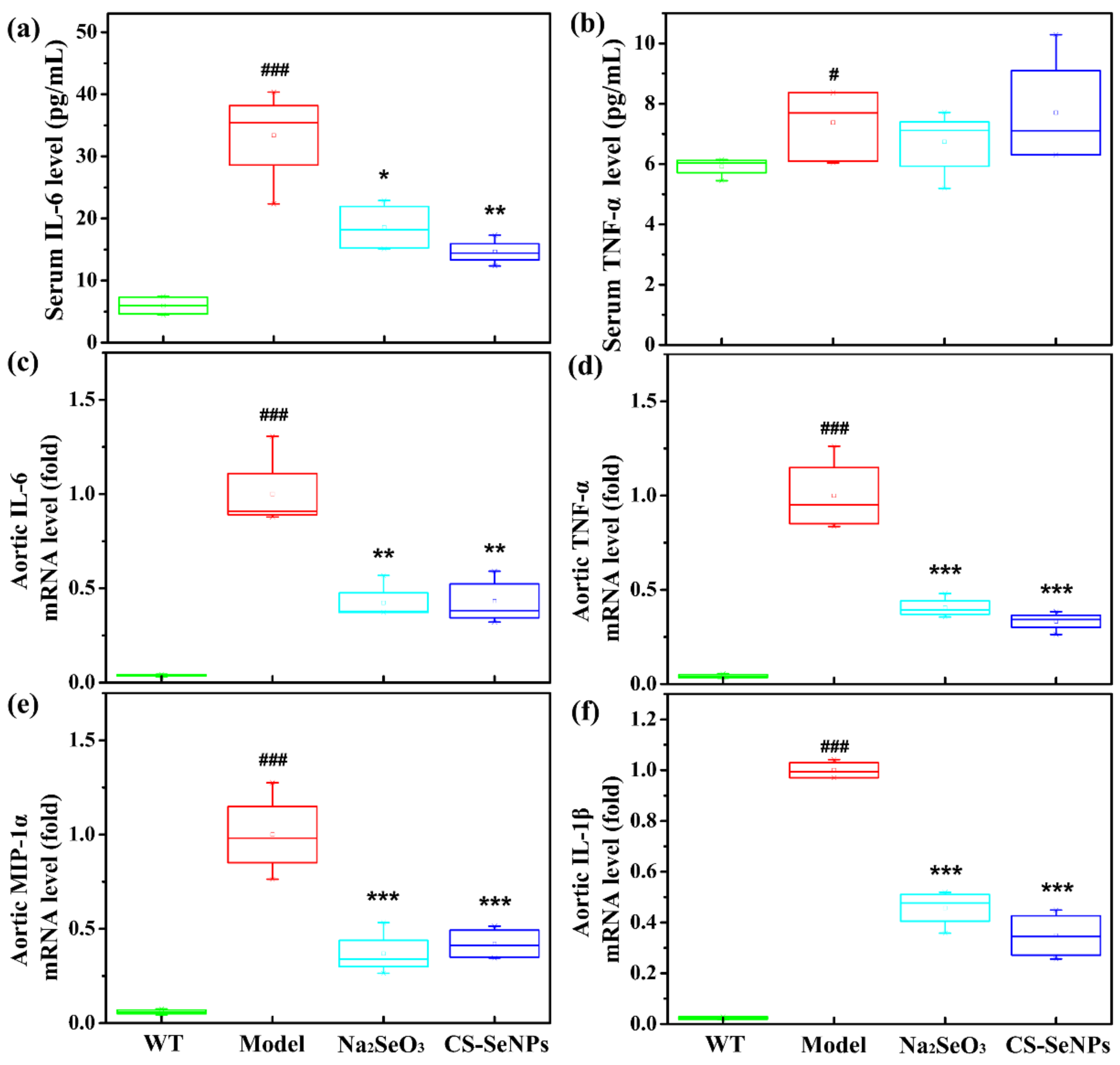
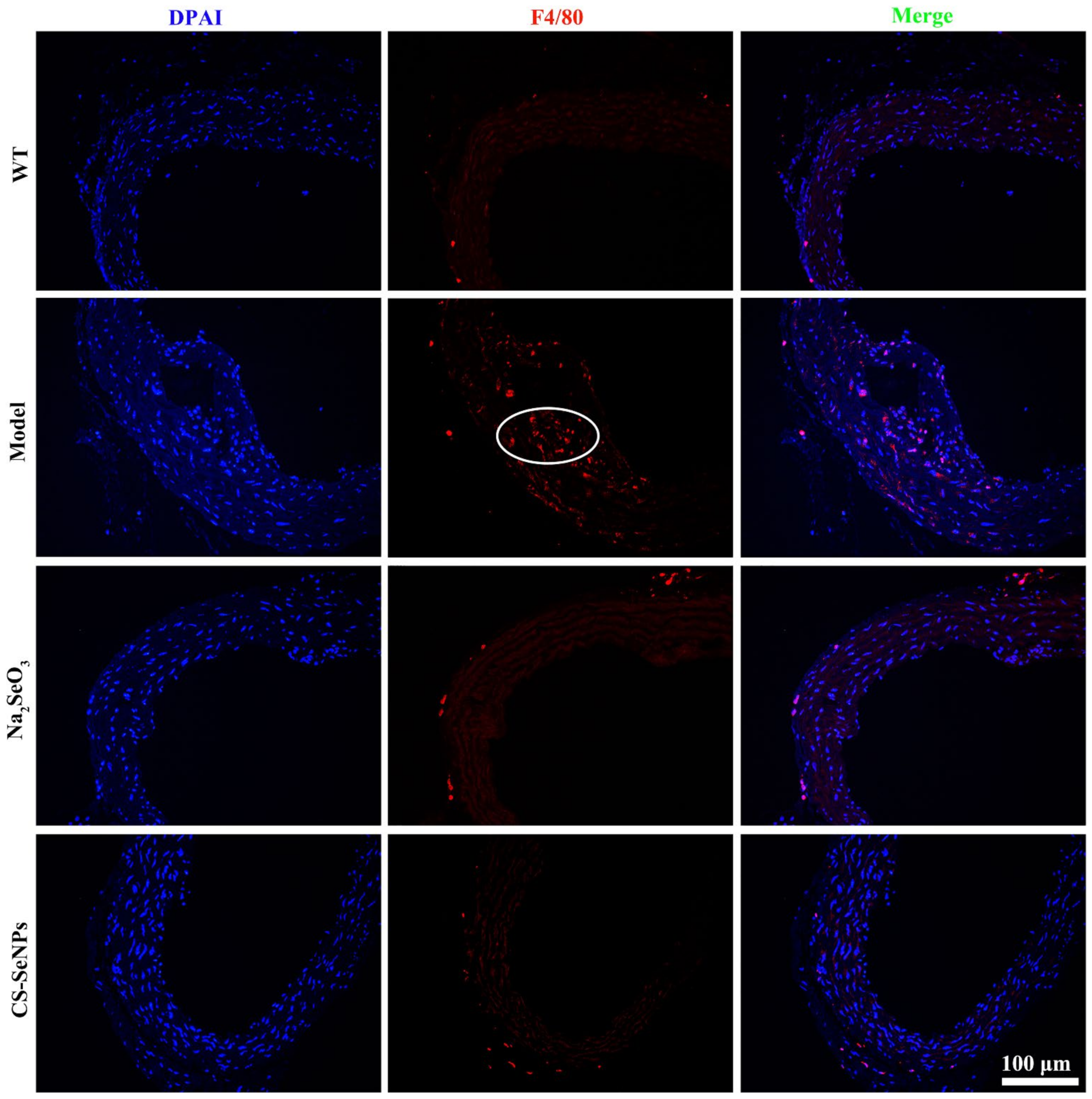
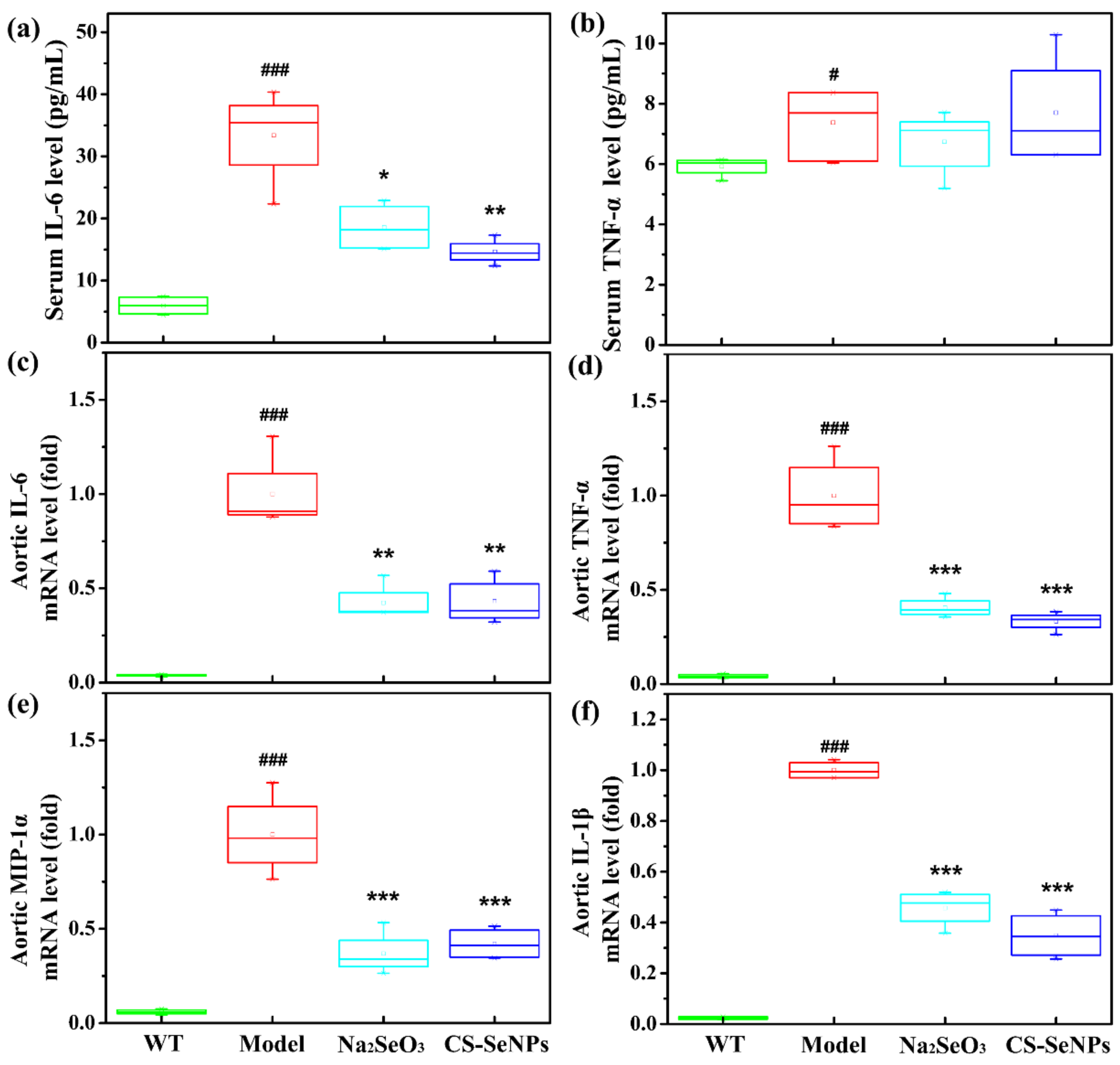
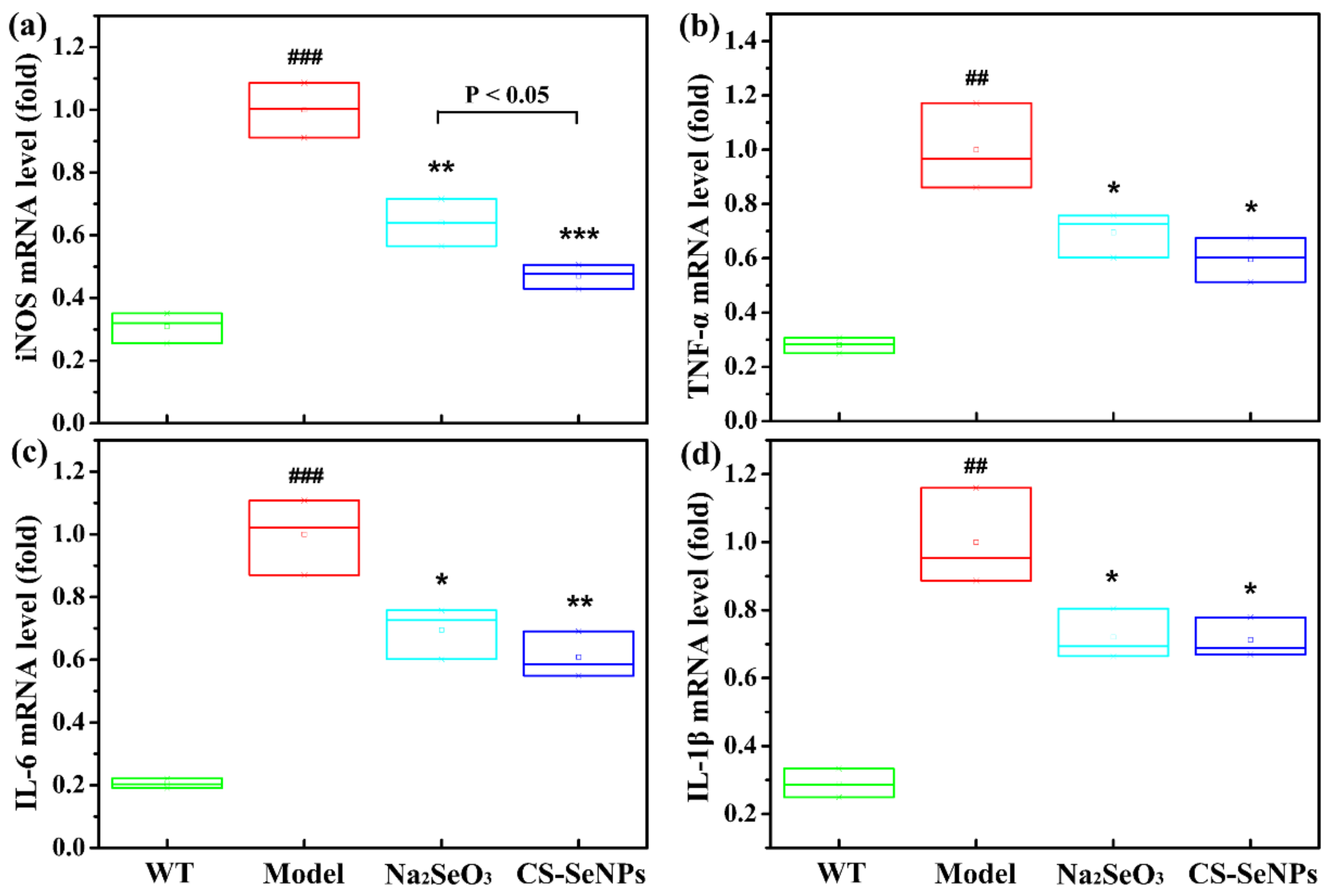
2.5. Both CS-SeNPs and Na2SeO3 Inhibited Macrophage Recruitment and the Inflammatory Response in ApoE-/- Mice
2.6. CS-SeNPs and Na2SeO3 Inhibited Oxidative Stress in ApoE-/- Mice
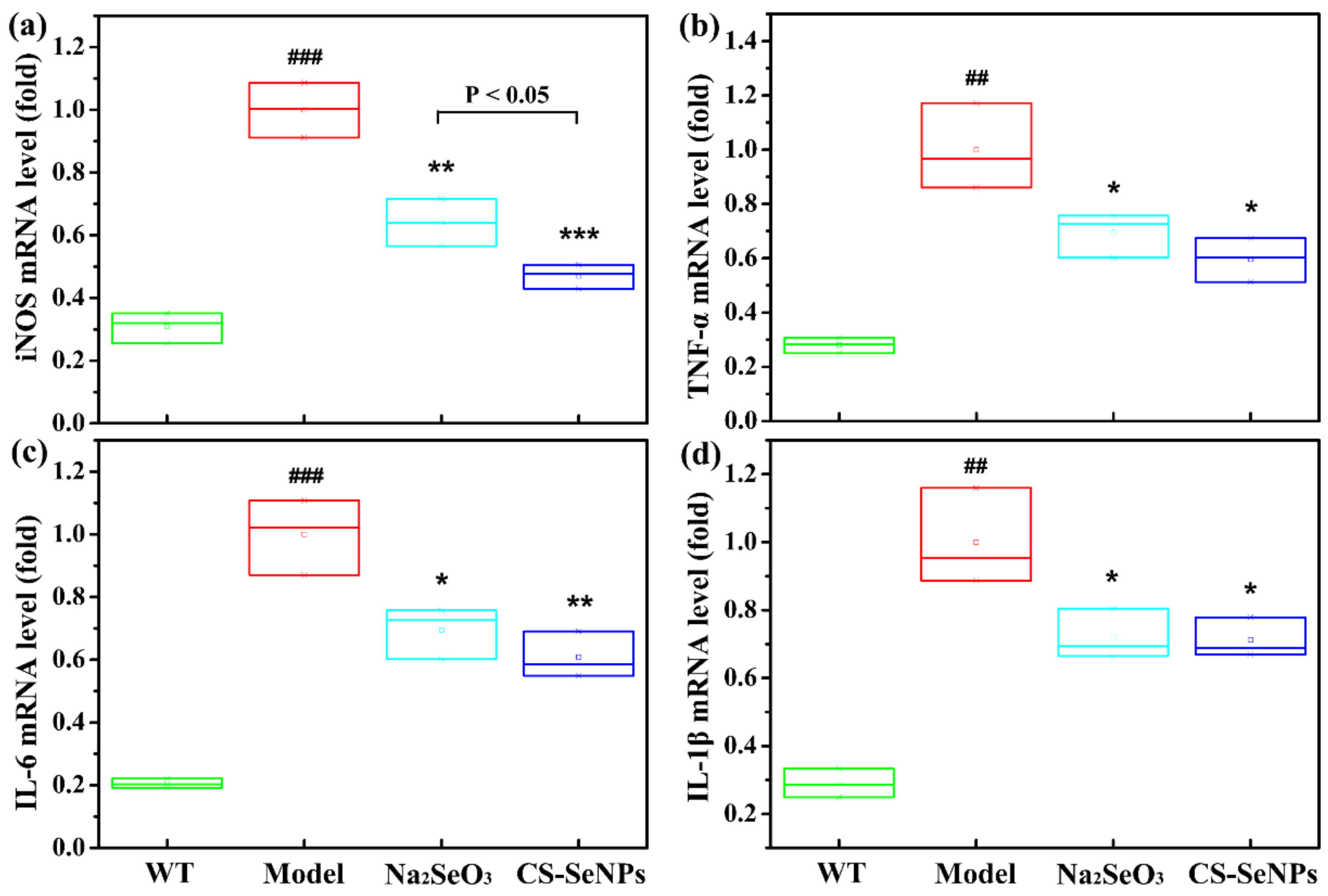
2.7. Both CS-SeNPs and Na2SeO3 Inhibited Lipopolysaccharide (LPS)-Induced Endothelial Dysfunction and Inflammatory Response in Human ECs
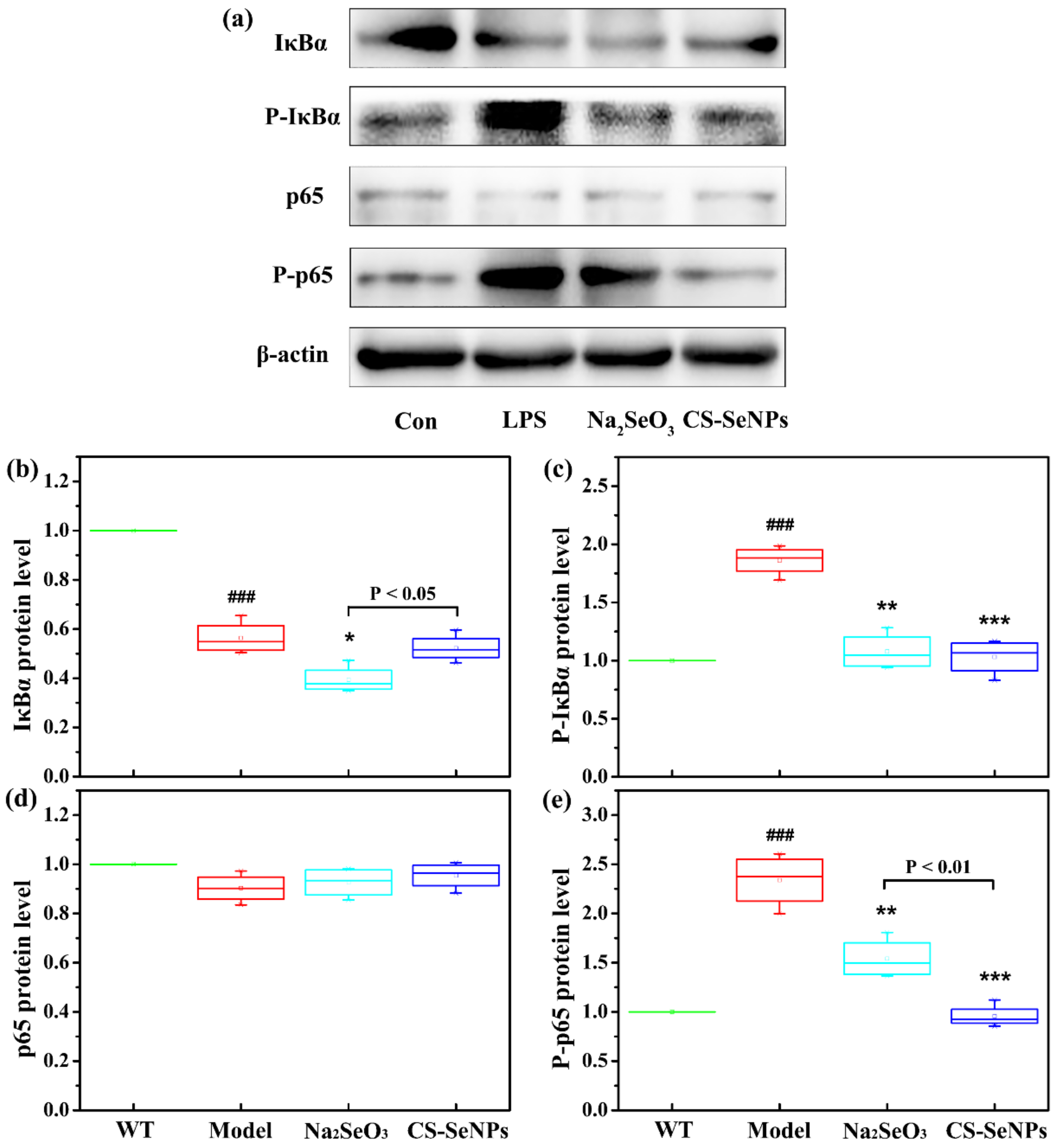
2.8. Both CS-SeNPs and Na2SeO3 Inhibited the Activation of Nuclear Factor-κB (NF-κB) Signaling Pathway Induced by LPS in Human ECs
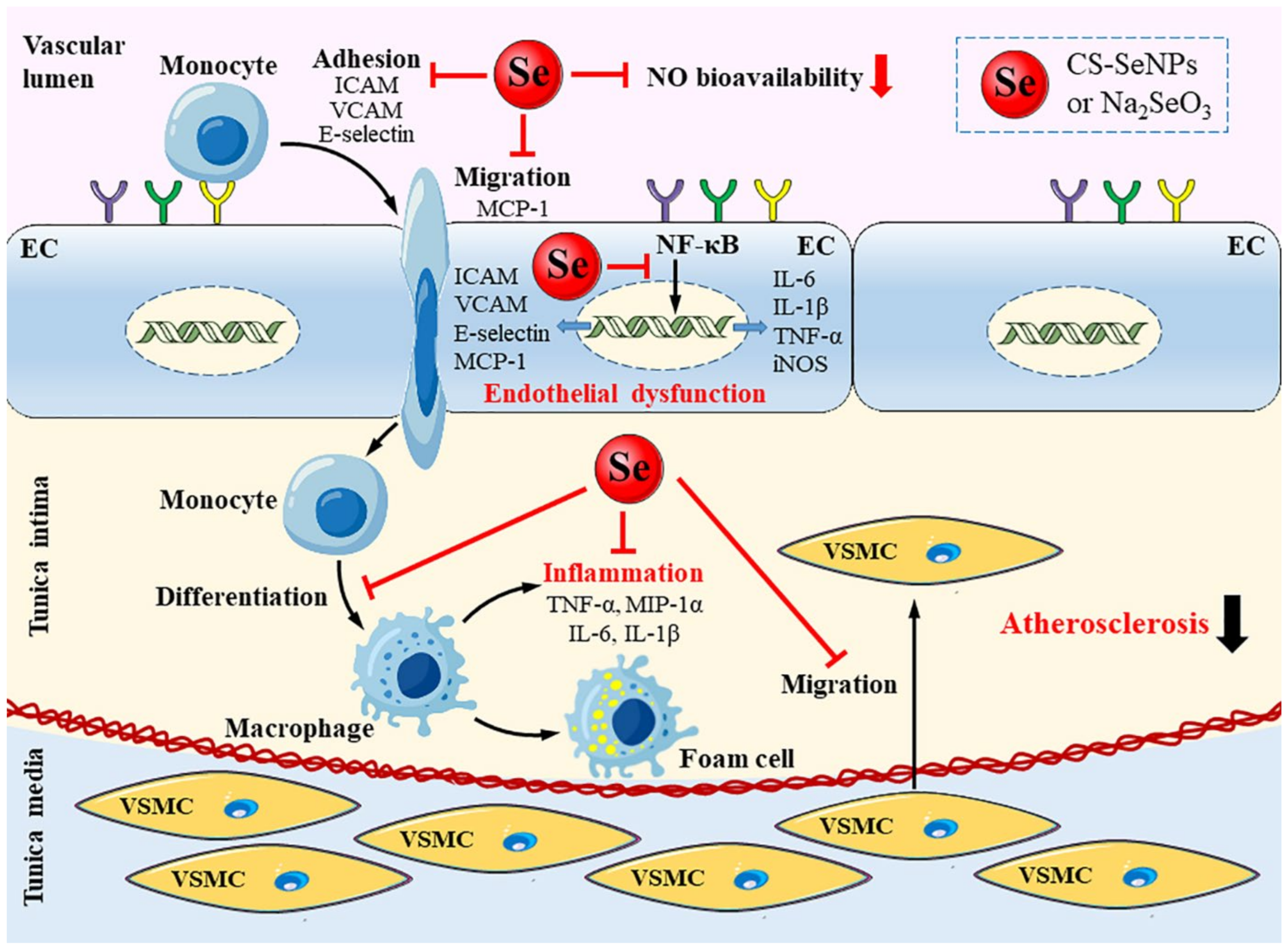
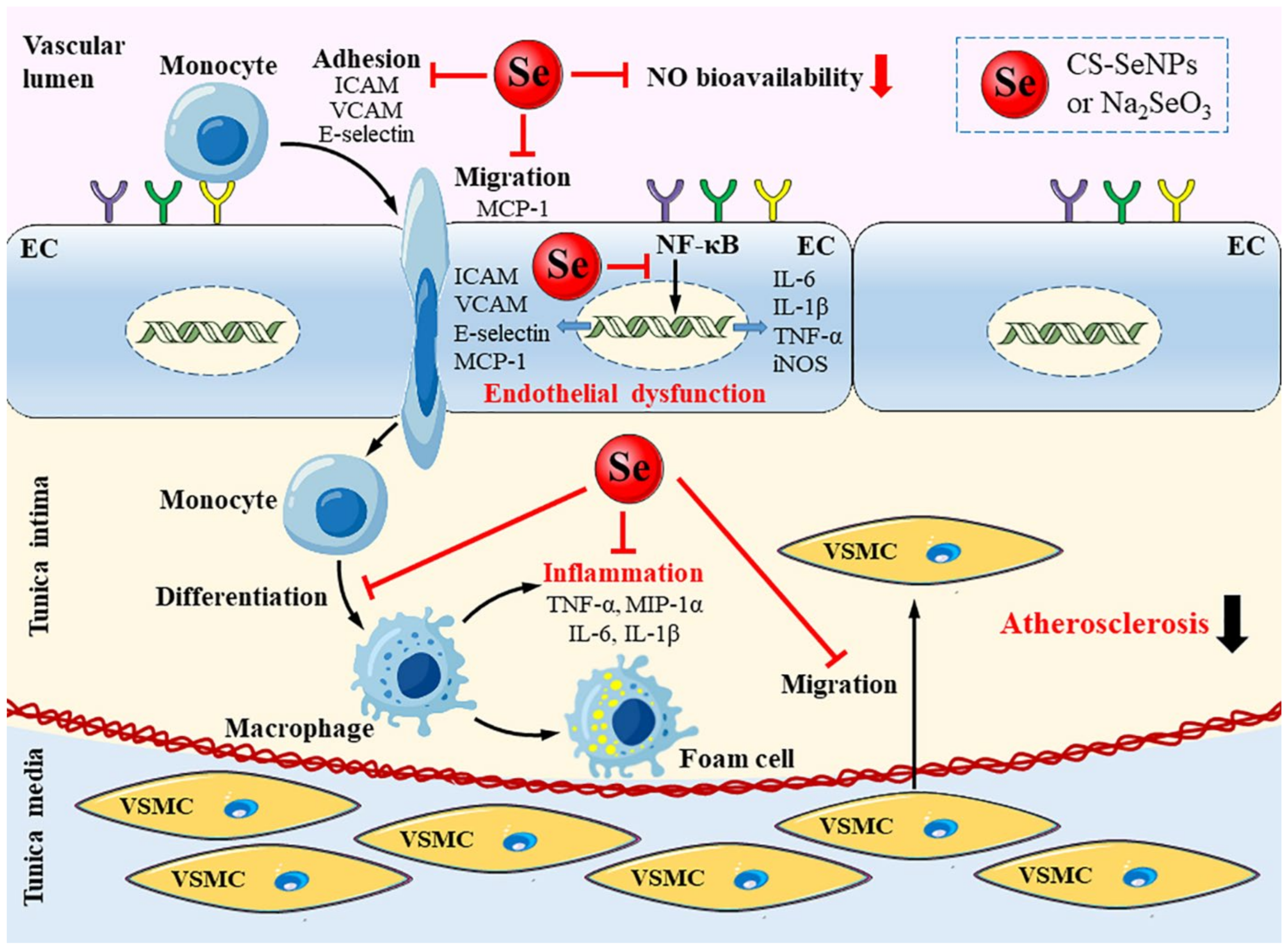
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of Selenium Nanoparticles Decorated with Chitosan
4.3. Characterization of CS-SeNPs
4.4. Animals
4.5. Sampling
4.6. Atherosclerotic Lesion Analysis
4.7. Biochemical Analysis
4.8. Immunofluorescence and Immunohistochemistry Analysis
4.9. Cell Culture and Treatment
4.10. Determination of Gene Expression by qPCR
4.11. Western Blot Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- World Health Organization. Cardiovascular Diseases (CVDs) Fact Sheet. 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 11 July 2021).
- Benjamin, E.J.; Blaha, M.J.; Chiuve, S.E.; Cushman, M.; Muntner, P. Heart disease and stroke statistics-2017 update: A report from the American Heart Association. Circulation 2017, 136, e146–e603. [Google Scholar] [CrossRef]
- Glass, C.K.; Witztum, J.L. Atherosclerosis: The road ahead. Cell 2001, 104, 503–516. [Google Scholar] [CrossRef] [Green Version]
- Libby, P.; Buring, J.E.; Badimon, L.; Hansson, G.K.; Deanfield, J.; Bittencourt, M.S.; Tokgzolu, L.; Lewis, E.F. Atherosclerosis. Nat. Rev. Dis. Primers 2019, 5, 56–73. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Ridker, P.M.; Hansson, G.K. Progress and challenges in translating the biology of atherosclerosis. Nature 2011, 473, 317–325. [Google Scholar] [CrossRef]
- Bäck, M.; Yurdagul, A., Jr.; Tabas, I.; Öörni, K.; Kovanen, P.T. Inflammation and its resolution in atherosclerosis: Mediators and therapeutic opportunities. Nat. Rev. Cardiol. 2019, 16, 384–406. [Google Scholar] [CrossRef] [PubMed]
- Mezentsev, A.; Bezsonov, E.; Kashirskikh, D.; Baig, M.S.; Eid, A.H.; Orekhov, A. Proatherogenic sialidases and desialylated lipoproteins: 35 years of research and current state from bench to bedside. Biomedicines 2021, 9, 600. [Google Scholar] [CrossRef] [PubMed]
- Bezsonov, E.E.; Sobenin, I.A.; Orekhov, A.N. Immunopathology of atherosclerosis and related diseases: Focus on molecular biology. Int. J. Mol. Sci. 2021, 22, 4080. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Liu, H.; Xu, H.; Huang, K. Selenium in the prevention of atherosclerosis and its underlying mechanisms. Metallomics 2017, 9, 21–37. [Google Scholar] [CrossRef]
- Wojcicki, J.; Rozewicka, L.; Barcew-Wiszniewska, B.; Samochowiec, L.; Juzwiak, S.; Kadlubowska, D.; Tustanowski, S.; Juzyszyn, Z. Effect of selenium and vitamin E on the development of experimental atherosclerosis in rabbits. Atherosclerosis 1991, 87, 9–16. [Google Scholar] [CrossRef]
- Dong, H.L.; Yuan, N.; Sun, T.; Dun, A.S.; Hou, H.F. Effects of selenium supplement on atherogenesis of ApoE-knockout mice fed high fat diet. Zhonghua Xin Xue Guan Bing Za Zhi 2016, 44, 244–249. [Google Scholar]
- Mehta, U.; Kang, B.; Kukreja, R.S.; Bansal, M.P. Ultrastructural examination of rabbit aortic wall following high-fat diet feeding and selenium supplementation: A transmission electron microscopy study. J. Appl. Toxicol. 2002, 22, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Krohn, R.M.; Lemaire, M.; Silva, L.; Lemarié, C.; Smits, J.E. High-selenium lentil diet protects against arsenic-induced atherosclerosis in a mouse model. J. Nutr. Biochem. 2016, 27, 9–15. [Google Scholar] [CrossRef]
- Weekley, C.M.; Harris, H.H. Which form is that? The importance of selenium speciation and metabolism in the prevention and treatment of disease. Chem. Soc. Rev. 2013, 42, 8870–8894. [Google Scholar] [CrossRef] [PubMed]
- Maiyo, F.; Singh, M. Selenium nanoparticles: Potential in cancer gene and drug delivery. Nanomedicine 2017, 12, 1075–1089. [Google Scholar] [CrossRef] [PubMed]
- Constantinescu-Aruxandei, D.; Frincu, R.M.; Capra, L.; Oancea, F. Selenium analysis and speciation in dietary supplements based on next-generation selenium ingredients. Nutrients 2018, 10, 1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakr, T.M.; Korany, M.; Katti, K.V. Selenium nanomaterials in biomedicine-An overview of new opportunities in nanomedicine of selenium. J. Drug Deliv. Sci. Technol. 2018, 46, 223–233. [Google Scholar] [CrossRef]
- Kieliszek, M.; Lipinski, B.; Blazejak, S. Application of sodium selenite in the prevention and treatment of cancers. Cells 2017, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Soumya, M.; Devi, K.S.; Santhiya, R.; Rajeshkumar, S.; Kumar, S.V. Selenium nanoparticles: A potent chemotherapeutic agent and an elucidation of its mechanism. Colloid. Surface. B 2018, 170, 280–292. [Google Scholar]
- Chen, W.; Li, Y.; Yang, S.; Yue, L.; Jiang, Q.; Xia, W. Synthesis and antioxidant properties of chitosan and carboxymethyl chitosan-stabilized selenium nanoparticles. Carbohydr. Polym. 2015, 132, 574–581. [Google Scholar] [CrossRef]
- Zeng, S.; Yu, K.; Liu, Y.; Shen, Y.; Lan, Z.; Cheng, L.; Liu, A.; Li, S.; Hu, X.; Wu, H. Synthesis and antidiabetic properties of chitosan-stabilized selenium nanoparticles. Colloids Surf. B Biointerf. 2018, 170, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Asri-Rezaei, S.; Nourian, A.; Shalizar-Jalali, A.; Najafi, G.; Nazarizadeh, A.; Koohestani, M.; Karimi, A. Selenium supplementation in the form of selenium nanoparticles and selenite sodium improves mature male mice reproductive performances. Iran. J. Basic. Med. Sci. 2018, 21, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Guo, K.; Zhang, C.; Talukder, M.; Lv, M.W.; Li, J.Y.; Li, J.L. Comparison of nanoparticle-selenium, selenium-enriched yeast and sodium selenite on the alleviation of cadmium-induced inflammation via NF-kB/IkappaB pathway in heart. Sci. Total. Environ. 2021, 773, 145442–145452. [Google Scholar] [CrossRef] [PubMed]
- Skalickova, S.S.; Milosavljevic, V.; Cihalova, K.; Horky, P.H.; Richtera, L.; Adam, V. Selenium nanoparticles as a nutritional supplement. Nutrition 2017, 33, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Xiao, J.; Liu, H.; Liu, H. Selenium nanoparticles alleviate hyperlipidemia and vascular injury in ApoE-deficient mice by regulating cholesterol metabolism and reducing oxidative stress. Metallomics 2020, 12, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Mao, L.; Xiao, J.; Wu, Y.; Liu, H. Selenium nanoparticles inhibit the formation of atherosclerosis in apolipoprotein E deficient mice by alleviating hyperlipidemia and oxidative stress. Eur. J. Pharmacol. 2021, 902, 174120. [Google Scholar] [CrossRef] [PubMed]
- Albanese, A.; Tang, P.S.; Chan, W.C.W. The effect of nanoparticle size, shape, and surface chemistry on biological systems. Annu. Rev. Biomed. Eng. 2012, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Zhang, Y.; Zheng, W.; Fan, C.; Chen, T. Positive surface charge enhances selective cellular uptake and anticancer efficacy of selenium nanoparticles. Lnorg. Chem. 2012, 51, 8956–8963. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Xiao, J.; Liu, H. Enhanced oxidase-like activity of selenium nanoparticles stabilized by chitosan and application in a facile colorimetric assay for mercury (II). Biochem. Eng. J. 2019, 152, 107384. [Google Scholar] [CrossRef]
- Gimbrone, M.A., Jr.; Garcia-Cardena, G. Endothelial cell dysfunction and the pathobiology of atherosclerosis. Circ. Res. 2016, 118, 620–636. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S.; Hamann, J.; Lin, H.H.; Stacey, M. F4/80 and the related adhesion-GPCRs. Eur. J. Immunol. 2011, 41, 2472–2476. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant. Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Praticò, D. Lipid peroxidation in mouse models of atherosclerosis. Trends Cardiovasc. Med. 2001, 11, 112–116. [Google Scholar] [CrossRef]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 in health and disease: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Sign. 2010, 15, 1957–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colasanti, M.; Suzuki, H. The dual personality of NO. Trends Pharmacol. Sci. 2000, 21, 249–252. [Google Scholar] [CrossRef]
- Förstermann, U.; Xia, N.; Li, H. Roles of vascular oxidative stress and nitric oxide in the pathogenesis of atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef] [PubMed]
- Dauphinee, S.M.; Karsan, A. Lipopolysaccharide signaling in endothelial cells. Lab. Invest. 2006, 86, 9–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, H.W.; Juan, H.F.; Huang, H.C.; Lin, J.Y.; Sinchaikul, S.; Lai, T.C.; Chen, C.F.; Chen, S.T.; Wang, G.J. Lipopolysaccharide-stimulated responses in rat aortic endothelial cells by a systems biology approach. Proteomics 2006, 6, 5915–5928. [Google Scholar] [CrossRef] [PubMed]
- Dayang, E.Z.; Plantinga, J.; Ter Ellen, B.; van Meurs, M.; Molema, G.; Moser, J. Identification of LPS-activated endothelial subpopulations with distinct inflammatory phenotypes and regulatory signaling Mechanisms. Front. Immunol. 2019, 10, 1169–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pennathur, S.; Heinecke, J.W. Mechanisms for oxidative stress in diabetic cardiovascular disease. Antioxid. Redox Sign. 2007, 9, 955–969. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Verma, I.M. NF-kappaB regulation in the immune system. Nat. Rev. Immunolo. 2002, 2, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Gareus, R.; Kotsaki, E.; Xanthoulea, S.; Made, I.; Gijbels, M.; Kardakaris, R.; Polykratis, A.; Kollias, G.; Winther, M.; Pasparakis, M. Endothelial cell-specific NF-kappaB inhibition protects mice from atherosclerosis. Cell Metab. 2008, 8, 372–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Yu, W.; Hargrove, J.L.; Greenspan, P.; Dean, R.G.; Taylor, E.W.; Hartle, D.K. Inhibition of TNF-alpha induced ICAM-1, VCAM-1 and E-selectin expression by selenium. Atherosclerosis 2002, 161, 381–386. [Google Scholar] [CrossRef]
- Vunta, H.; Davis, F.; Palempalli, U.D.; Bhat, D.; Arner, R.J.; Thompson, J.T.; Peterson, D.G.; Reddy, C.C.; Prabhu, K.S. The anti-inflammatory effects of selenium are mediated through 15-deoxy-Δ12,14-prostaglandin J2 in macrophages. J. Biol. Chem. 2007, 282, 17964–17973. [Google Scholar] [CrossRef] [Green Version]
- Zamamiri-Davis, F.; Lu, Y.; Thompson, J.T.; Prabhu, K.S.; Reddy, C.C. Nuclear factor-κB mediates over-expression of cyclooxygenase-2 during activation of RAW 264.7 macrophages in selenium deficiency. Free Radical Biol. Med. 2002, 32, 890–897. [Google Scholar] [CrossRef]
- Pei, Z.; Hui, L.; Ying, G.; Jin, Y.; Lin, D. Sodium selenite inhibits the expression of VEGF, TGFbeta(1) and IL-6 induced by LPS in human PC3 cells via TLR4-NF-(K)B signaling blockage. Int. Immunopharmacol. 2010, 10, 50–56. [Google Scholar] [CrossRef]
- Zhu, C.; Zhang, S.; Song, C.; Zhang, Y.; Ling, Q.; Hoffmann, P.R.; Li, J.; Chen, T.; Zheng, W.; Zhi, H. Selenium nanoparticles decorated with Ulva lactuca polysaccharide potentially attenuate colitis by inhibiting NF-κB mediated hyper inflammation. J. Nanobiotechnol. 2017, 15, 20–34. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, H.; Yan, X.; Zhang, L. Comparison of short-term toxicity between Nano-Se and selenite in mice. Life Sci. 2005, 76, 1099–1109. [Google Scholar] [CrossRef]
- Zhang, J.S.; Gao, X.Y.; Zhang, L.D.; Bao, Y.P. Biological effects of a nano red elemental selenium. Biofactors 2001, 15, 27–38. [Google Scholar] [CrossRef]
- Ormrod, D.J.; Holmes, C.C.; Miller, T.E. Dietary chitosan inhibits hypercholesterolaemia and atherogenesis in the apolipoprotein E-deficient mouse model of atherosclerosis. Atherosclerosis 1998, 138, 329–334. [Google Scholar] [CrossRef]
- Yu, Y.; Luo, T.; Liu, S.; Song, G.; Han, J.; Wang, Y.; Yao, S.; Feng, L.; Qin, S. Chitosan Oligosaccharides Attenuate Atherosclerosis and Decrease Non-HDL in ApoE-/- Mice. J. Atheroscler. Thromb. 2015, 22, 926–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, J.; Cao, H.; Guo, S.; Xiao, S.; Li, N.; Li, M.; Wu, Y.; Liu, H. Long-term administration of low-dose selenium nanoparticles with different sizes aggravated atherosclerotic lesions and exhibited toxicity in apolipoprotein E-deficient mice. Chem. Biol. Interact. 2021, 347, 109601. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Hu, J.; Wu, Y.; Luo, H.; Meng, W.; Xiao, B.; Xiao, X.; Zhou, Z.; Liu, F. Rheb (ras homolog enriched in brain 1) deficiency in mature macrophages prevents atherosclerosis by repressing macrophage proliferation, inflammation, and lipid uptake. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1787–1801. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, W.; Xuan, L.L.; Han, F.F.; Lv, Y.L.; Wan, Z.R.; Liu, H.; Ren, L.L.; Gong, L.L.; Liu, L.H. Akebia Saponin D inhibits the formation of atherosclerosis in ApoE(-/-) mice by attenuating oxidative stress-induced apoptosis in endothelial cells. Atherosclerosis 2019, 285, 23–30. [Google Scholar] [CrossRef]
- Dufailly, V.; Noël, L.; Guérin, T. Determination of chromium, iron and selenium in foodstuffs of animal origin by collision cell technology, inductively coupled plasma mass spectrometry (ICP-MS), after closed vessel microwave digestion. Anal. Chim. Acta 2006, 565, 214–221. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, H.J.; Farr, A.; Randall, R.J. Protein measurement with the pholin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | MDA (nmol/mL) | T-AOC (U/mL) | GPx (U/mL) |
|---|---|---|---|
| WT | 1.10 ± 0.19 | 5.09 ± 0.78 | 260.60 ± 59.53 |
| Model | 8.93 ± 0.57 ### | 1.76 ± 0.33 ### | 154.12 ± 33.55 ### |
| Na2SeO3 | 7.64 ± 0.46 ** | 2.29 ± 0.35 * | 280.48 ± 54.11 *** |
| CS-Se | 8.46 ± 1.32 | 2.78 ± 0.51 ** | 191.59 ± 33.45 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, J.; Li, N.; Xiao, S.; Wu, Y.; Liu, H. Comparison of Selenium Nanoparticles and Sodium Selenite on the Alleviation of Early Atherosclerosis by Inhibiting Endothelial Dysfunction and Inflammation in Apolipoprotein E-Deficient Mice. Int. J. Mol. Sci. 2021, 22, 11612. https://doi.org/10.3390/ijms222111612
Xiao J, Li N, Xiao S, Wu Y, Liu H. Comparison of Selenium Nanoparticles and Sodium Selenite on the Alleviation of Early Atherosclerosis by Inhibiting Endothelial Dysfunction and Inflammation in Apolipoprotein E-Deficient Mice. International Journal of Molecular Sciences. 2021; 22(21):11612. https://doi.org/10.3390/ijms222111612
Chicago/Turabian StyleXiao, Junying, Na Li, Shengze Xiao, Yuzhou Wu, and Hongmei Liu. 2021. "Comparison of Selenium Nanoparticles and Sodium Selenite on the Alleviation of Early Atherosclerosis by Inhibiting Endothelial Dysfunction and Inflammation in Apolipoprotein E-Deficient Mice" International Journal of Molecular Sciences 22, no. 21: 11612. https://doi.org/10.3390/ijms222111612
APA StyleXiao, J., Li, N., Xiao, S., Wu, Y., & Liu, H. (2021). Comparison of Selenium Nanoparticles and Sodium Selenite on the Alleviation of Early Atherosclerosis by Inhibiting Endothelial Dysfunction and Inflammation in Apolipoprotein E-Deficient Mice. International Journal of Molecular Sciences, 22(21), 11612. https://doi.org/10.3390/ijms222111612