
Smad7 Deficiency in Myeloid Cells Does Not Affect Liver Injury, Inflammation or Fibrosis after Chronic CCl4 Exposure in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
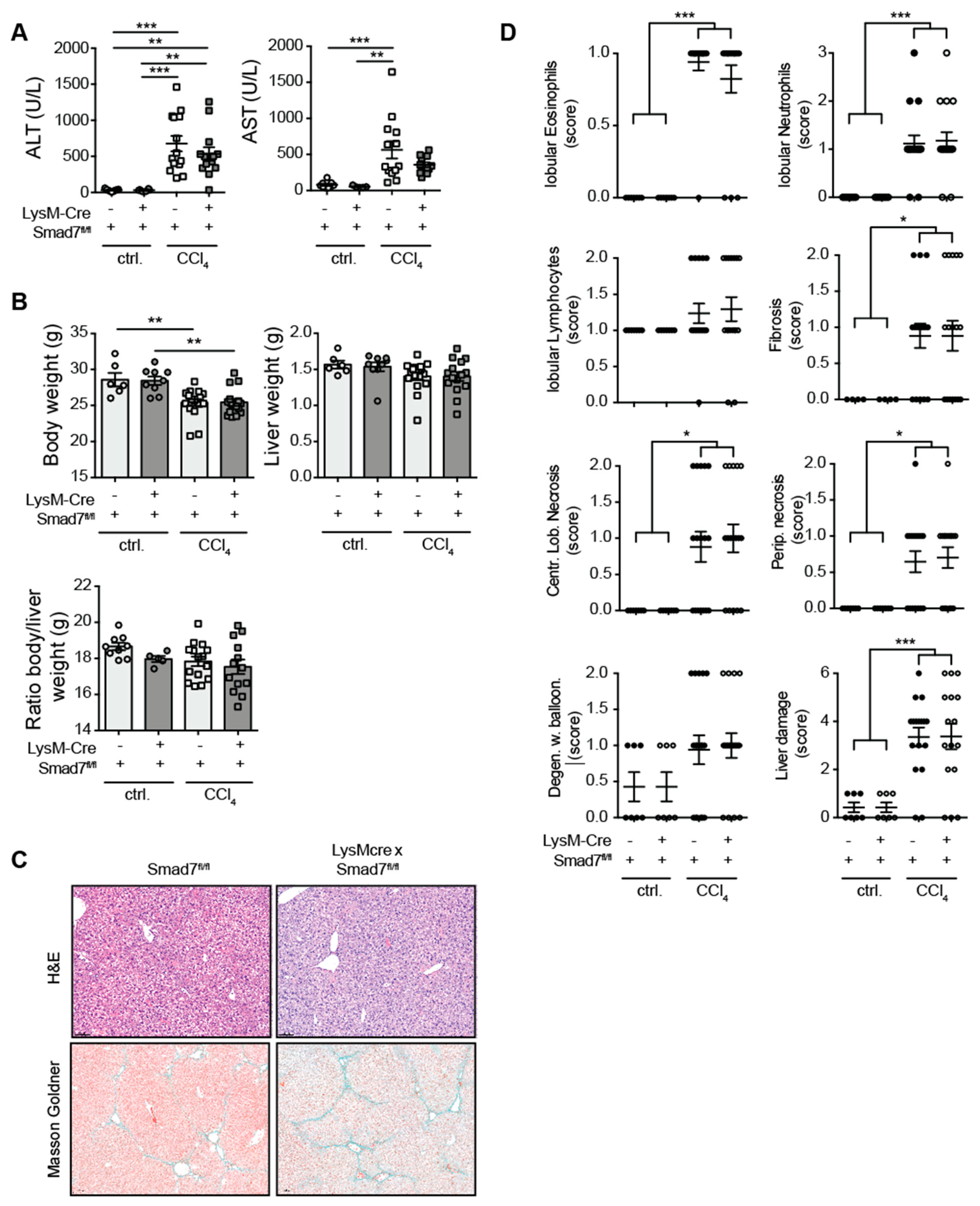
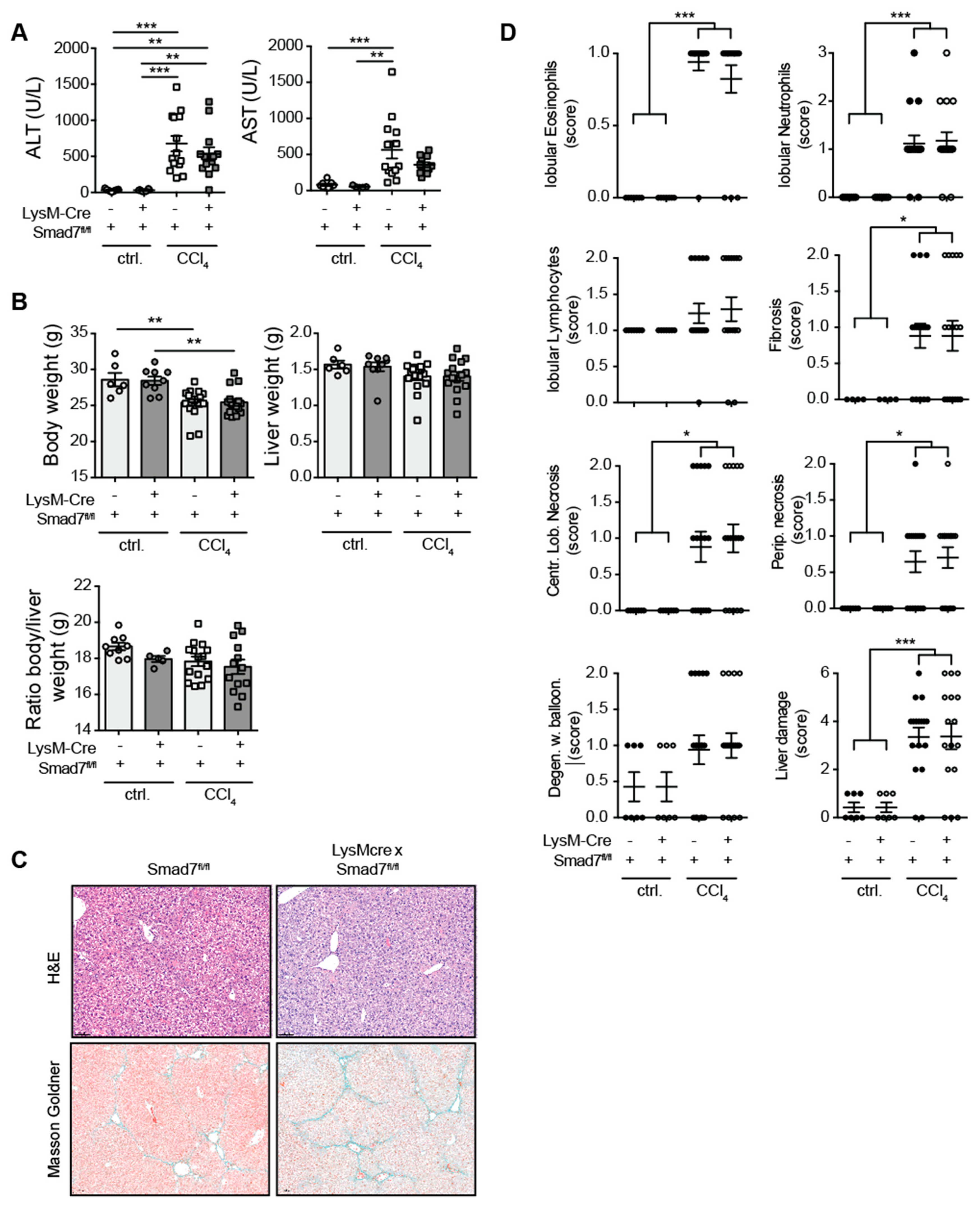
2.1. Myeloid-Specific Knock-Down of Smad7 Does Not Affect Liver Injury after Chronic CCl4 Exposure
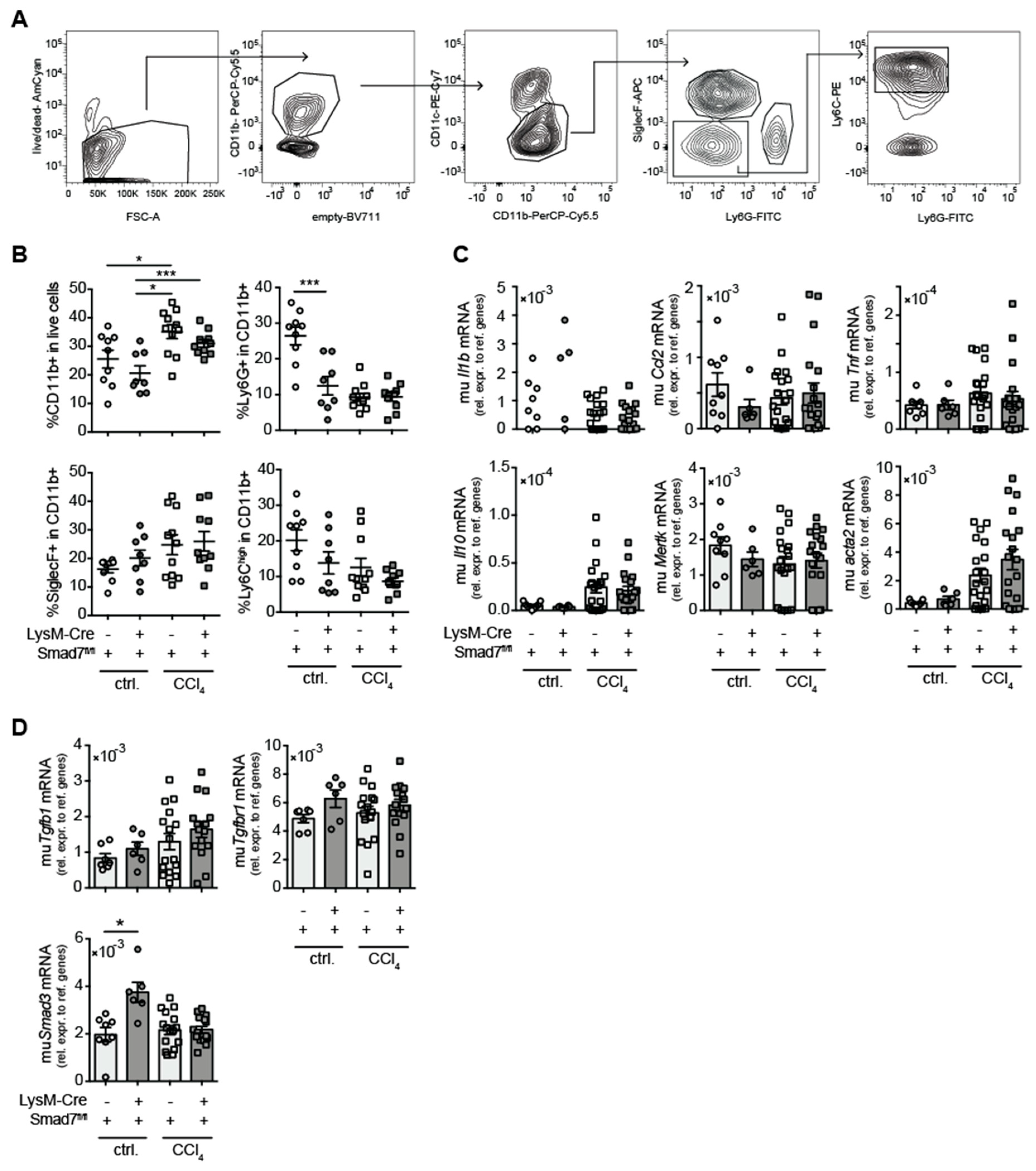
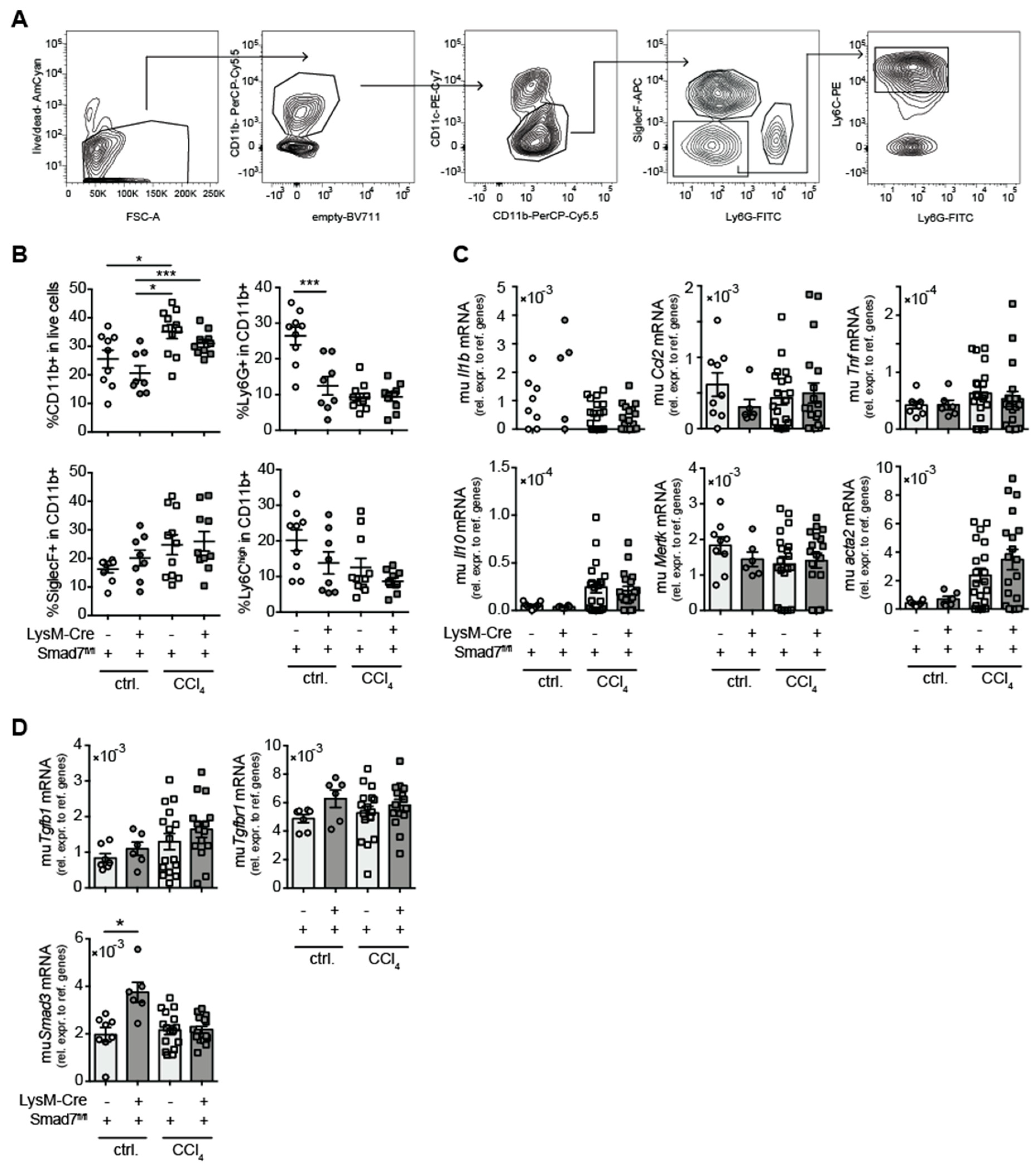
2.2. Comparable Myeloid-Cell Infiltration in Livers of Myeloid-Cell-Specific, Smad7-Deficient Mice and Wild-Type Littermates after Chronic CCl4 Treatment
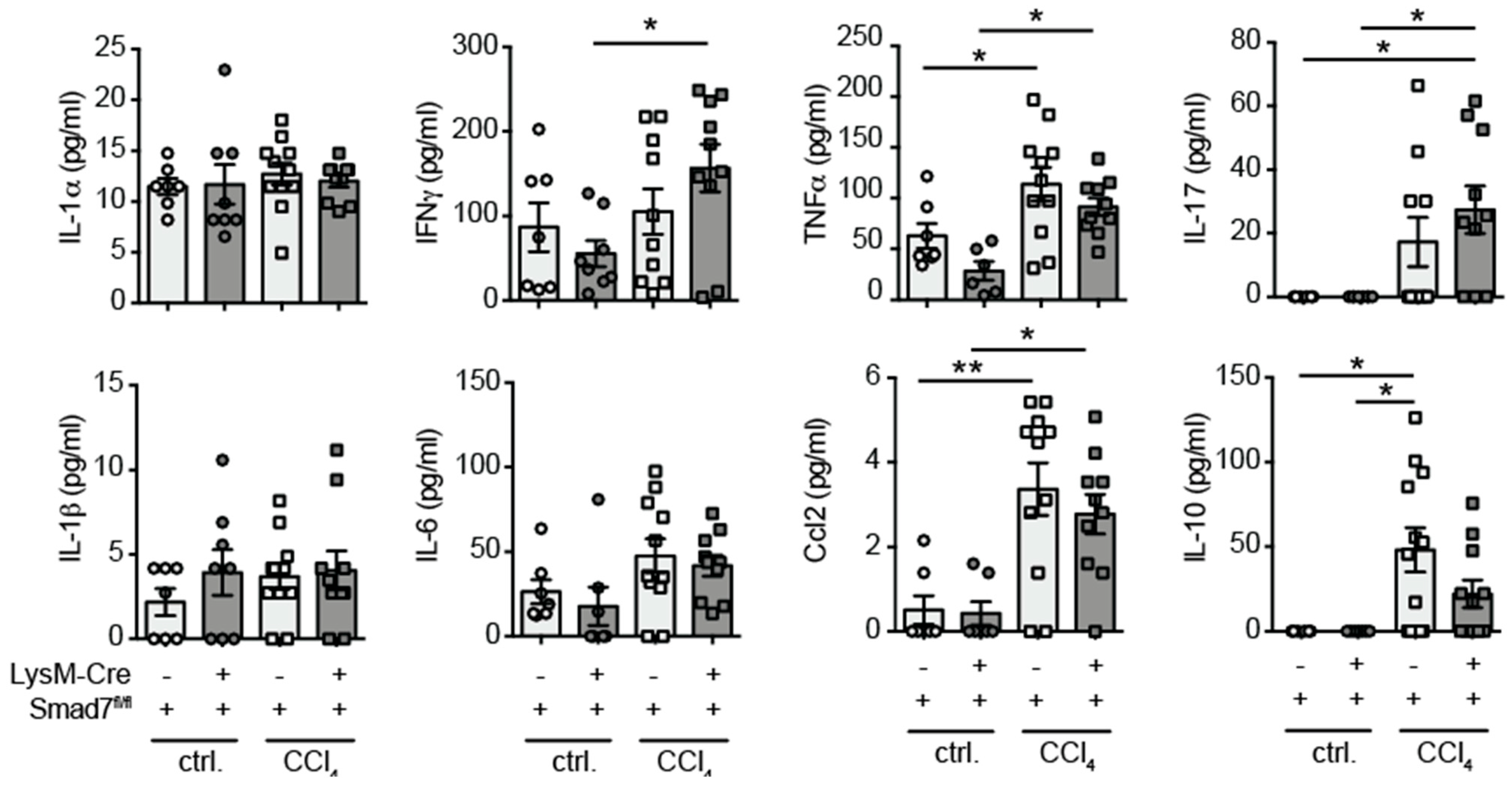
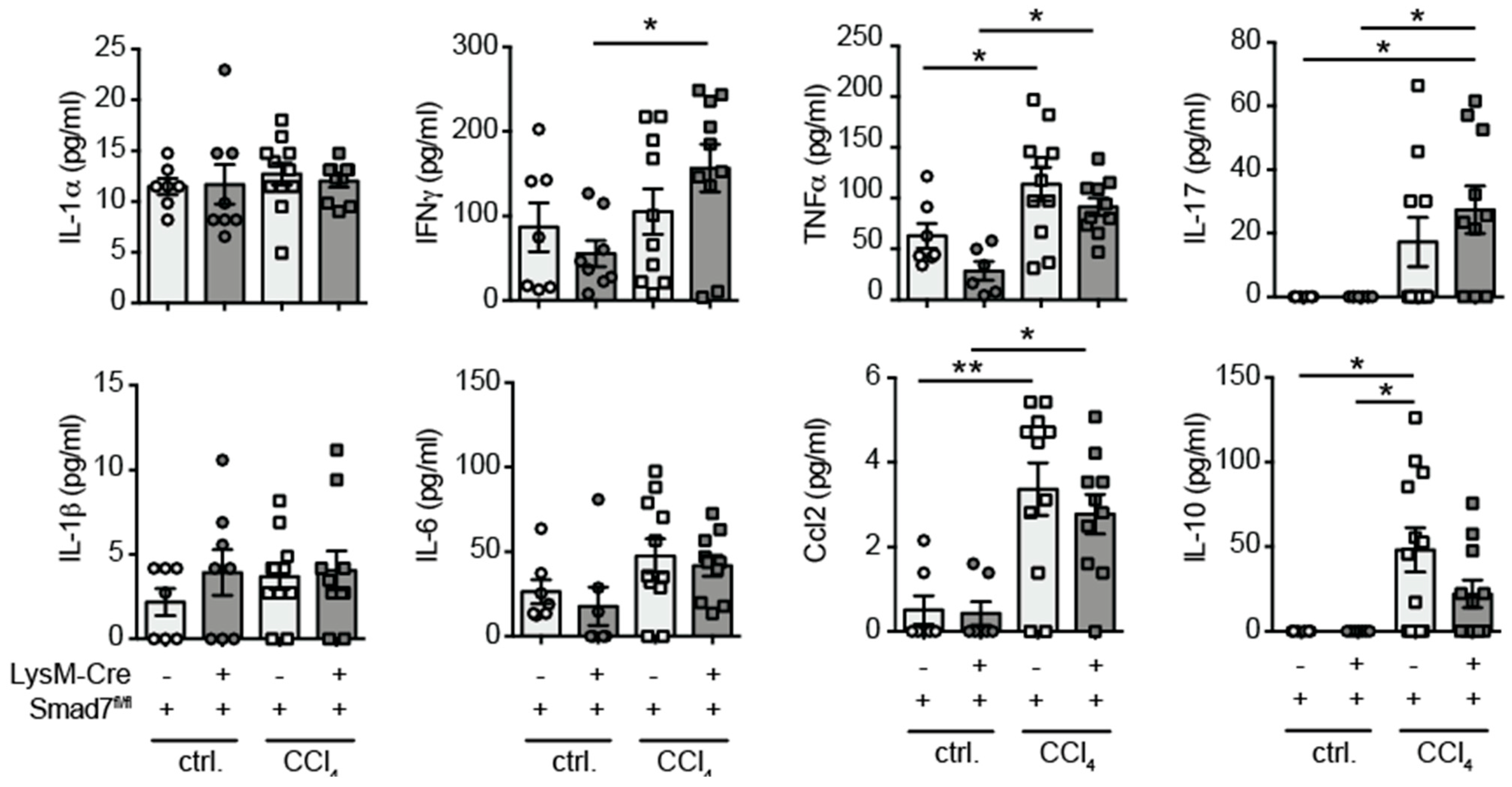
2.3. Similar Cytokine Production by Intrahepatic Non-Parenchymal Cells (NPCs) in the Presence and Absence of Myeloid-Expressed Smad7 after Chronic CCl4-Mediated Liver Injury
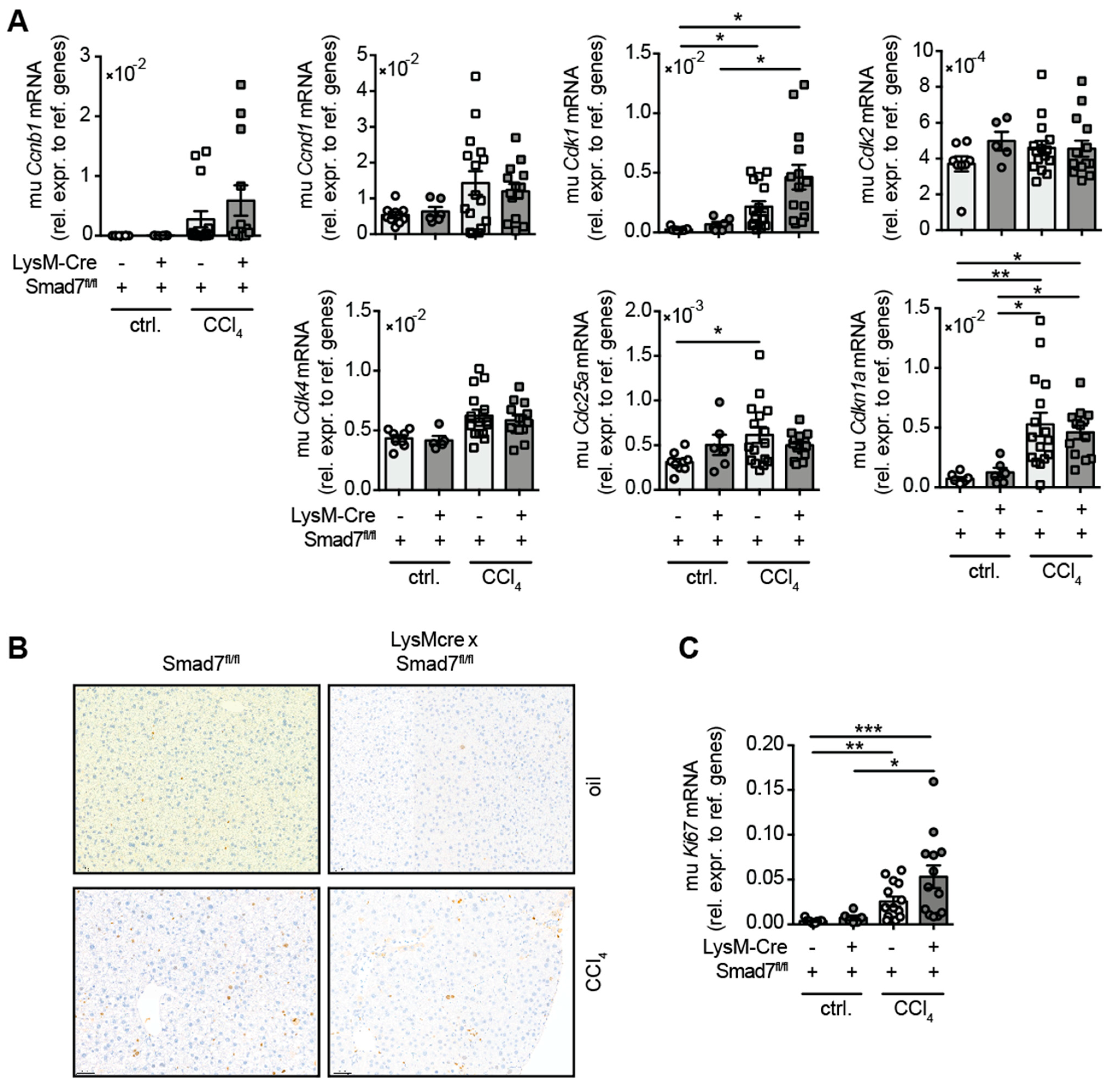
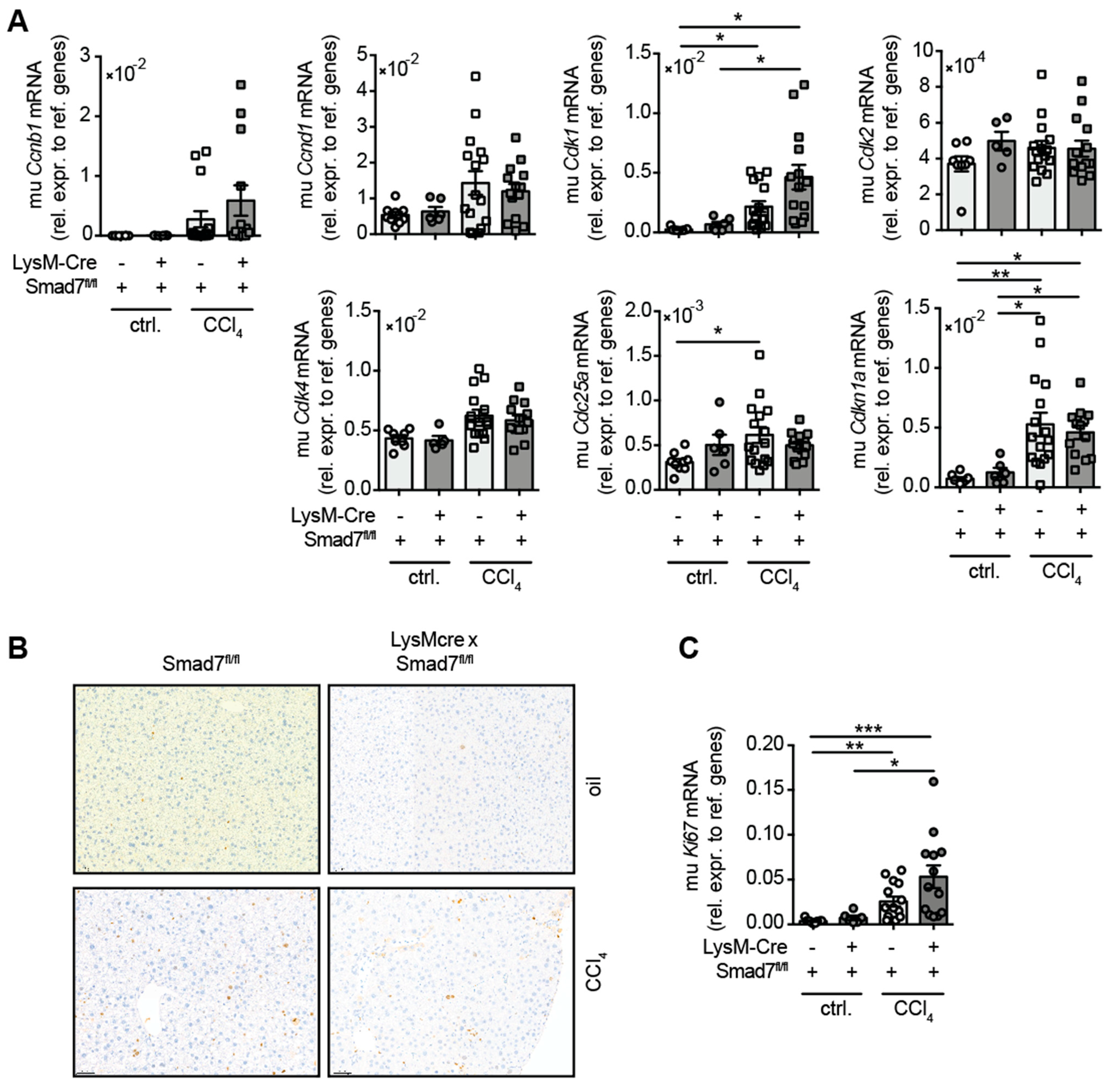
2.4. The Lack of Smad7 in Myeloid Cells Does Not Affect Liver Regeneration
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Induction and Evaluation of Chronic Liver Inflammation and Fibrosis Using Carbon Tetra Chloride
4.3. Isolation and Stimulation of Non-Parenchymal Liver Cells
4.4. Flow Cytometry
4.5. mRNA Isolation and Quantitative RT-PCR
4.6. Histology
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frangogiannis, N. Transforming growth factor-β in tissue fibrosis. J. Exp. Med. 2020, 217, e20190103. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Liu, C.; Zhou, D.; Zhang, L. TGF-β/SMAD Pathway and Its Regulation in Hepatic Fibrosis. J. Histochem. Cytochem. 2016, 64, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A.; Vannella, K.M. Macrophages in Tissue Repair, Regeneration, and Fibrosis. Immunity 2016, 44, 450–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, A.C.K.; Huang, X.R.; Zhou, L.; Heuchel, R.; Lai, K.N.; Lan, H.Y. Disruption of the Smad7 gene promotes renal fibrosis and inflammation in unilateral ureteral obstruction (UUO) in mice. Nephrol. Dial. Transplant. 2009, 24, 1443–1454. [Google Scholar] [CrossRef] [Green Version]
- Latella, G.; Vetuschi, A.; Sferra, R.; Catitti, V.; D’Angelo, A.; Zanninelli, G.; Flanders, K.C.; Gaudio, E. Targeted disruption of Smad3 confers resistance to the development of dimethylnitrosamine-induced hepatic fibrosis in mice. Liver Int. 2009, 29, 997–1009. [Google Scholar] [CrossRef]
- Tsunawaki, S.; Sporn, M.; Ding, A.; Nathan, C. Deactivation of macrophages by transforming growth factor-beta. Nature 1988, 334, 260–262. [Google Scholar] [CrossRef]
- Chen, B.; Huang, S.; Su, Y.; Wu, Y.-J.; Hanna, A.; Brickshawana, A.; Graff, J.; Frangogiannis, N.G. Macrophage Smad3 Protects the Infarcted Heart, Stimulating Phagocytosis and Regulating Inflammation. Circ. Res. 2019, 125, 55–70. [Google Scholar] [CrossRef]
- Chu, P.-S.; Nakamoto, N.; Ebinuma, H.; Usui, S.; Saeki, K.; Matsumoto, A.; Mikami, Y.; Sugiyama, K.; Tomita, K.; Kanai, T.; et al. C-C motif chemokine receptor 9 positive macrophages activate hepatic stellate cells and promote liver fibrosis in mice. Hepatology 2013, 58, 337–350. [Google Scholar] [CrossRef]
- Derynck, R.; Budi, E.H. Specificity, versatility, and control of TGF-β family signaling. Sci. Signal. 2019, 12, eaav5183. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Cui, C.; Xie, F.; Kiełbasa, S.; Mei, H.; van Dinther, M.; van Dam, H.; Bauer, A.; Zhang, L.; ten Dijke, P. VprBP mitigates TGF-β and Activin signaling by promoting Smurf1-mediated type I receptor degradation. J. Mol. Cell Biol. 2020, 12, 138–151. [Google Scholar] [CrossRef]
- MohanKumar, K.; Namachivayam, K.; Chapalamadugu, K.C.; Garzon, S.A.; Premkumar, M.H.; Tipparaju, S.M.; Maheshwari, A. Smad7 interrupts TGF-β signaling in intestinal macrophages and promotes inflammatory activation of these cells during necrotizing enterocolitis. Pediatr. Res. 2016, 79, 951–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedda, S.; De Simone, V.; Marafini, I.; Bevivino, G.; Izzo, R.; Paoluzi, O.A.; Colantoni, A.; Ortenzi, A.; Giuffrida, P.; Corazza, G.R.; et al. High Smad7 sustains inflammatory cytokine response in refractory coeliac disease. Immunology 2017, 150, 356–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, K.; Matsuzaki, K. Differential Regulation of TGF-β/Smad Signaling in Hepatic Stellate Cells between Acute and Chronic Liver Injuries. Front. Physiol. 2012, 3, 53. [Google Scholar] [CrossRef] [Green Version]
- Hamzavi, J.; Ehnert, S.; Godoy, P.; Ciuclan, L.; Weng, H.; Mertens, P.R.; Heuchel, R.; Dooley, S. Disruption of the Smad7 gene enhances CCI4-dependent liver damage and fibrogenesis in mice. J. Cell. Mol. Med. 2008, 12, 2130–2144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, T.; Dzieran, J.; Yuan, X.; Dropmann, A.; Maass, T.; Teufel, A.; Marhenke, S.; Gaiser, T.; Rückert, F.; Kleiter, I.; et al. Hepatocyte-specific Smad7 deletion accelerates DEN-induced HCC via activation of STAT3 signaling in mice. Oncogenesis 2017, 6, e294. [Google Scholar] [CrossRef]
- Endig, J.; Unrau, L.; Sprezyna, P.; Rading, S.; Karsak, M.; Goltz, D.; Heukamp, L.; Tiegs, G.; Diehl, L. Acute liver injury after CCl4 administration is independent of Smad7 expression in myeloid cells. Int. J. Mol. Sci. 2019, 20, 5528. [Google Scholar] [CrossRef] [Green Version]
- Pellicoro, A.; Ramachandran, P.; Iredale, J.P.; Fallowfield, J.A. Liver fibrosis and repair: Immune regulation of wound healing in a solid organ. Nat. Rev. Immunol. 2014, 14, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Fabregat, I.; Caballero-Díaz, D. Transforming Growth Factor-β-Induced Cell Plasticity in Liver Fibrosis and Hepatocarcinogenesis. Front. Oncol. 2018, 8, 357. [Google Scholar] [CrossRef] [Green Version]
- Dennis, E.A.; Smythies, L.E.; Grabski, R.; Li, M.; Ballestas, M.E.; Shimamura, M.; Sun, J.J.; Grams, J.; Stahl, R.; Niederweis, M.E.; et al. Cytomegalovirus promotes intestinal macrophage-mediated mucosal inflammation through induction of Smad7. Mucosal Immunol. 2018, 11, 1694–1704. [Google Scholar] [CrossRef] [Green Version]
- Dooley, S.; Hamzavi, J.; Ciuclan, L.; Godoy, P.; Ilkavets, I.; Ehnert, S.; Ueberham, E.; Gebhardt, R.; Kanzler, S.; Geier, A.; et al. Hepatocyte-specific Smad7 expression attenuates TGF-beta-mediated fibrogenesis and protects against liver damage. Gastroenterology 2008, 135, 642–659. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Wang, L.; Wang, X.; Luo, X.; Yang, L.; Zhang, R.; Yin, H.; Xie, D.; Pan, Y.; Chen, Y. Hepatic deletion of Smad7 in mouse leads to spontaneous liver dysfunction and aggravates alcoholic liver injury. PLoS ONE 2011, 6, e17415. [Google Scholar] [CrossRef] [Green Version]
- Böhm, F.; Köhler, U.A.; Speicher, T.; Werner, S. Regulation of liver regeneration by growth factors and cytokines. EMBO Mol. Med. 2010, 2, 294–305. [Google Scholar] [CrossRef]
- Karkampouna, S.; Goumans, M.-J.; ten Dijke, P.; Dooley, S.; Kruithof-de Julio, M. Inhibition of TGFβ type I receptor activity facilitates liver regeneration upon acute CCl4 intoxication in mice. Arch. Toxicol. 2016, 90, 347–357. [Google Scholar] [CrossRef]
- Tacke, F.; Zimmermann, H.W. Macrophage heterogeneity in liver injury and fibrosis. J. Hepatol. 2014, 60, 1090–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horst, A.K.; Tiegs, G.; Diehl, L. Contribution of Macrophage Efferocytosis to Liver Homeostasis and Disease. Front. Immunol. 2019, 10, 2670. [Google Scholar] [CrossRef] [PubMed]
- Guillot, A.; Tacke, F. The unexpected role of neutrophils for resolving liver inflammation by transmitting microRNA-223 to macrophages. Hepatology 2019, 71, 749–751. [Google Scholar] [CrossRef] [Green Version]
- Goh, Y.P.S.; Henderson, N.C.; Heredia, J.E.; Red Eagle, A.; Odegaard, J.I.; Lehwald, N.; Nguyen, K.D.; Sheppard, D.; Mukundan, L.; Locksley, R.M.; et al. Eosinophils secrete IL-4 to facilitate liver regeneration. Proc. Natl. Acad. Sci. USA 2013, 110, 9914–9919. [Google Scholar] [CrossRef] [Green Version]
- Proctor, W.R.; Chakraborty, M.; Chea, L.S.; Morrison, J.C.; Berkson, J.D.; Semple, K.; Bourdi, M.; Pohl, L.R. Eosinophils mediate the pathogenesis of halothane-induced liver injury in mice. Hepatology 2013, 57, 2026–2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Unrau, L.; Endig, J.; Goltz, D.; Sprezyna, P.; Ulrich, H.; Hagenstein, J.; Geers, B.; Kaftan, K.; Heukamp, L.C.; Tiegs, G.; et al. Smad7 Deficiency in Myeloid Cells Does Not Affect Liver Injury, Inflammation or Fibrosis after Chronic CCl4 Exposure in Mice. Int. J. Mol. Sci. 2021, 22, 11575. https://doi.org/10.3390/ijms222111575
Unrau L, Endig J, Goltz D, Sprezyna P, Ulrich H, Hagenstein J, Geers B, Kaftan K, Heukamp LC, Tiegs G, et al. Smad7 Deficiency in Myeloid Cells Does Not Affect Liver Injury, Inflammation or Fibrosis after Chronic CCl4 Exposure in Mice. International Journal of Molecular Sciences. 2021; 22(21):11575. https://doi.org/10.3390/ijms222111575
Chicago/Turabian StyleUnrau, Ludmilla, Jessica Endig, Diane Goltz, Paulina Sprezyna, Hanna Ulrich, Julia Hagenstein, Bernd Geers, Karina Kaftan, Lukas Carl Heukamp, Gisa Tiegs, and et al. 2021. "Smad7 Deficiency in Myeloid Cells Does Not Affect Liver Injury, Inflammation or Fibrosis after Chronic CCl4 Exposure in Mice" International Journal of Molecular Sciences 22, no. 21: 11575. https://doi.org/10.3390/ijms222111575
APA StyleUnrau, L., Endig, J., Goltz, D., Sprezyna, P., Ulrich, H., Hagenstein, J., Geers, B., Kaftan, K., Heukamp, L. C., Tiegs, G., & Diehl, L. (2021). Smad7 Deficiency in Myeloid Cells Does Not Affect Liver Injury, Inflammation or Fibrosis after Chronic CCl4 Exposure in Mice. International Journal of Molecular Sciences, 22(21), 11575. https://doi.org/10.3390/ijms222111575