
Long Term Immune Response Produced by the SputnikV Vaccine


 ,
,  , ,
, ,  , and
, and 
Abstract
:1. Introduction
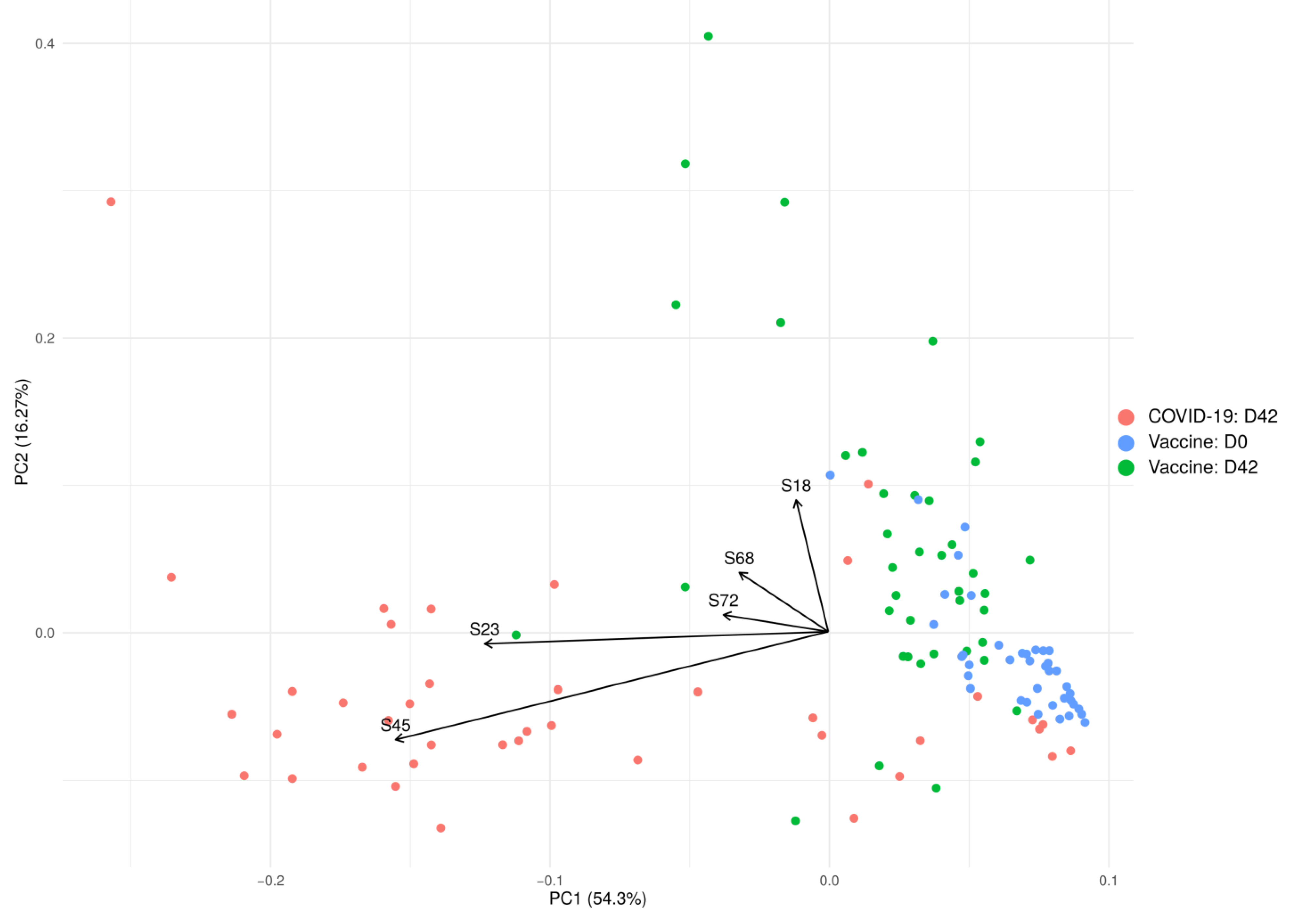
2. Results
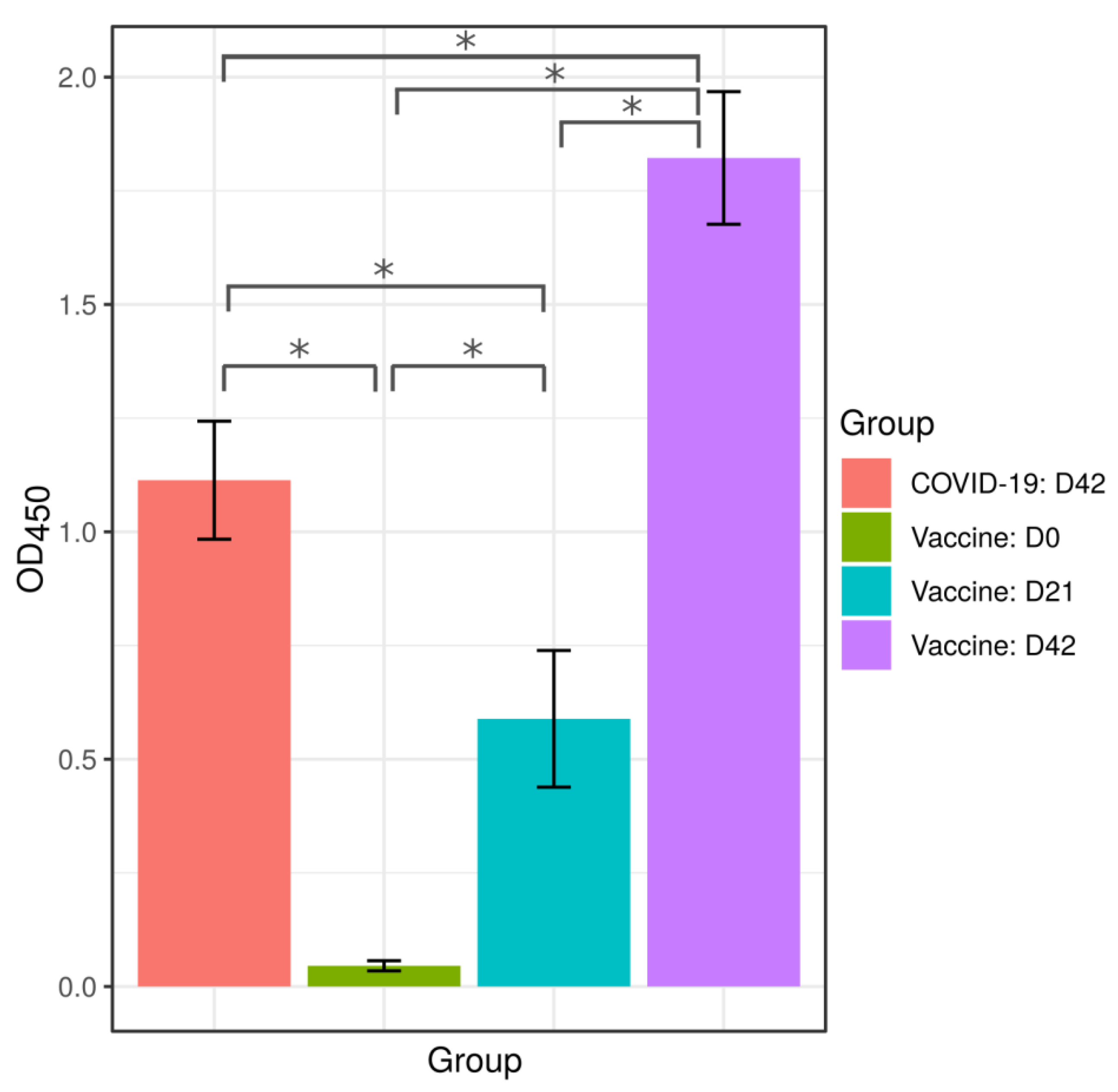
2.1. Anti-SARS-CoV-2 Antibodies in Serum of Convalescent COVID-19 and Vaccinated with Sputnikv
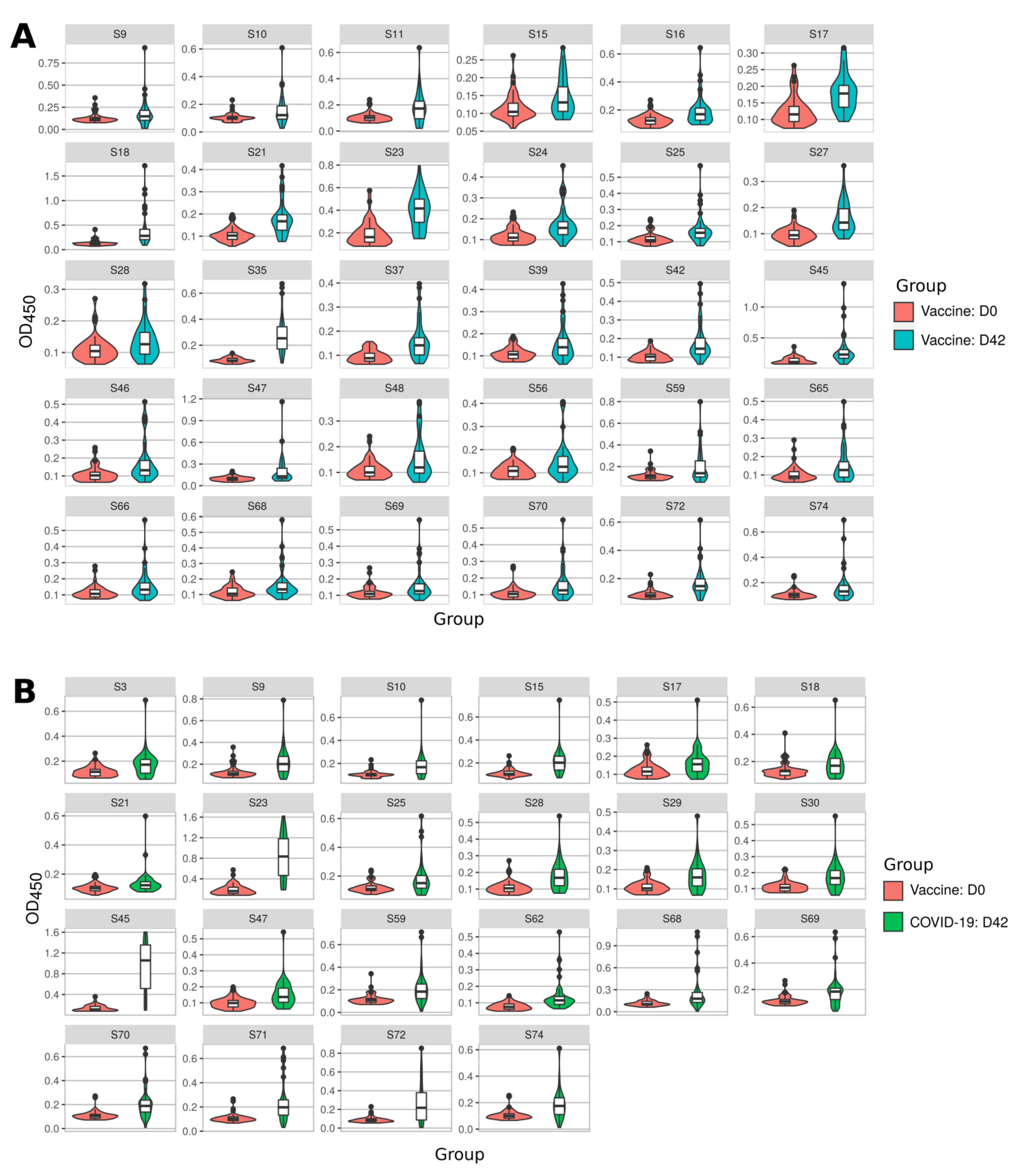
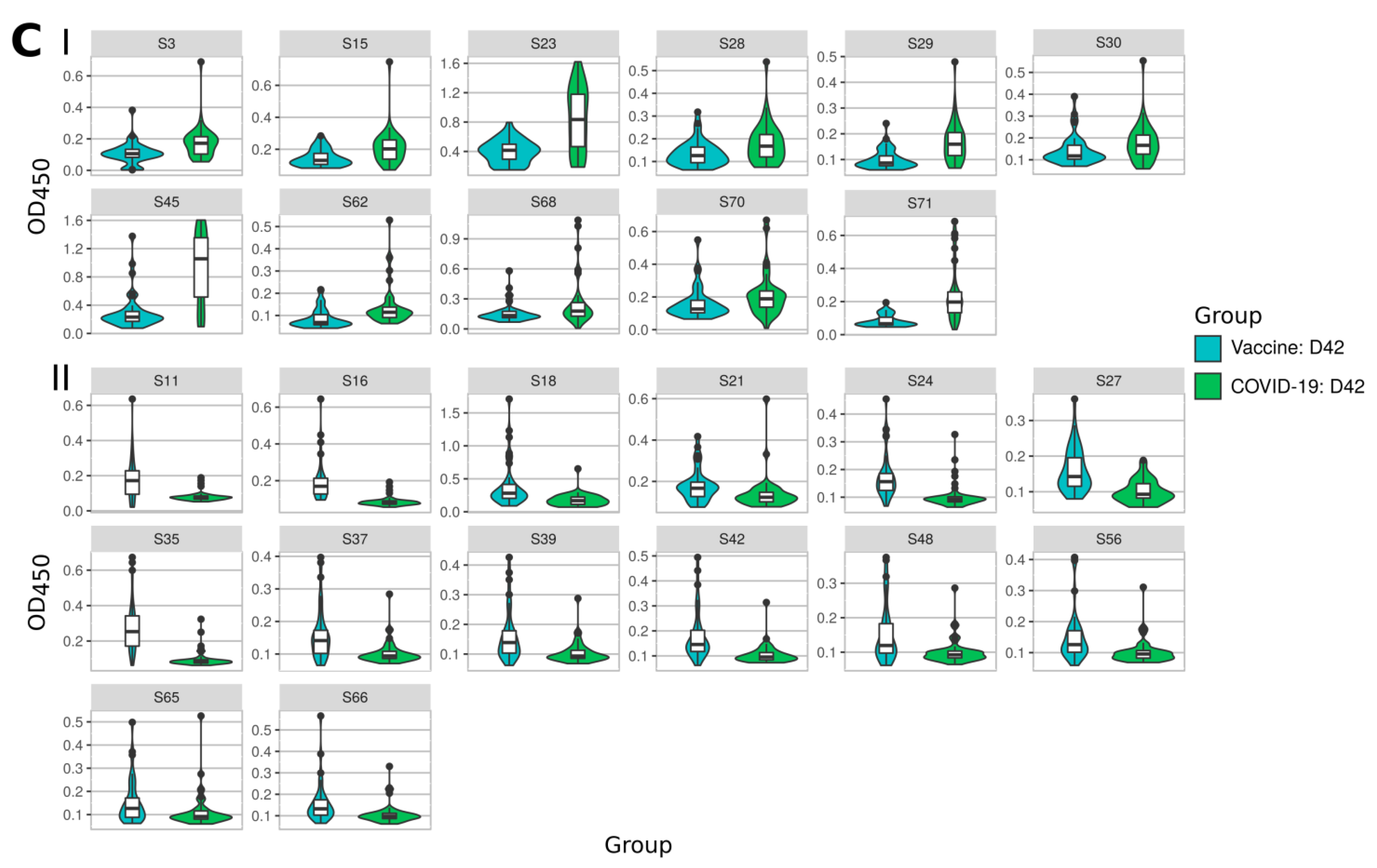
2.2. Immune Response to SARS-CoV-2 Peptides in Convalescent COVID-19 and Sputnikv Vaccinated Sera
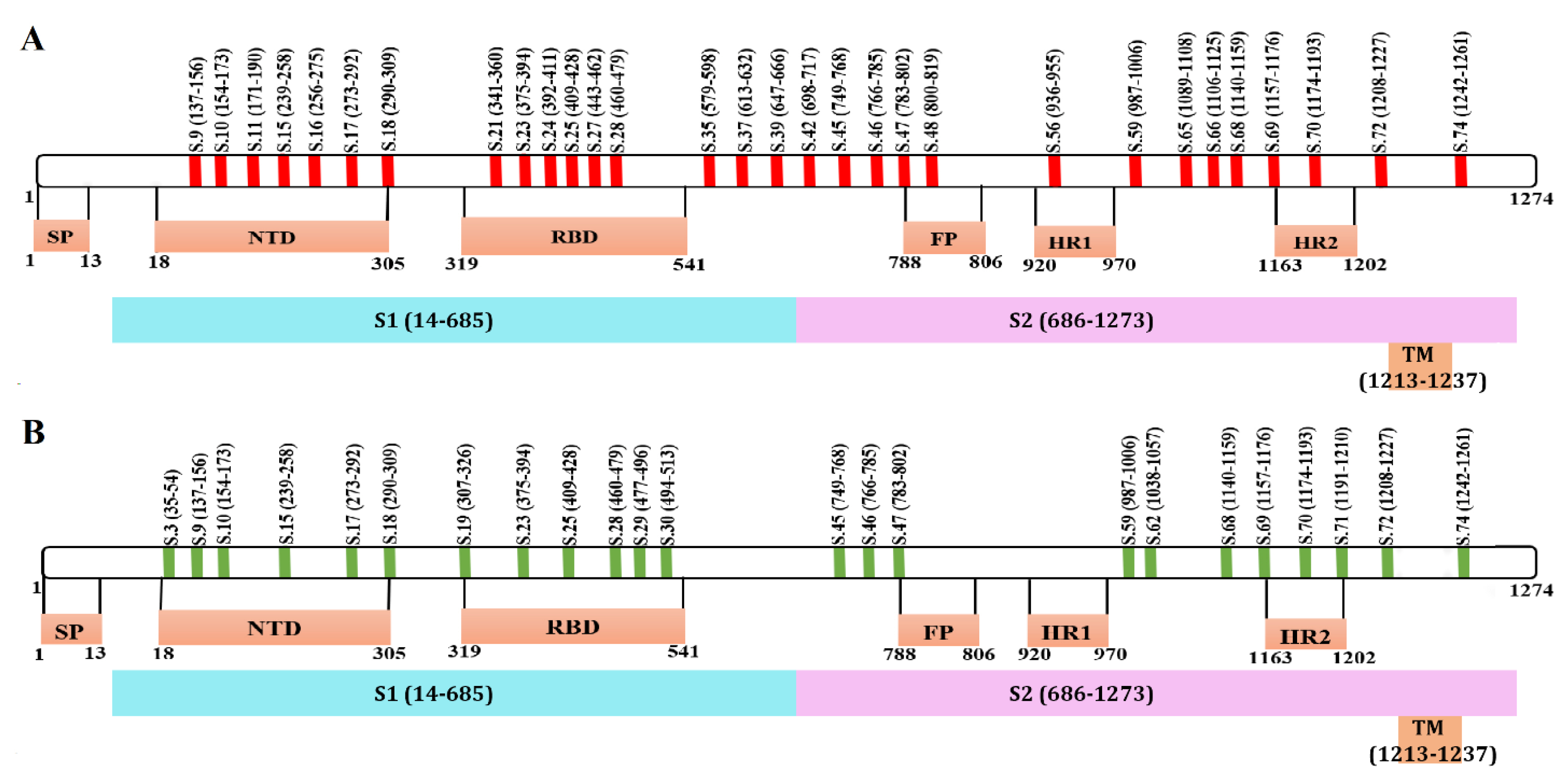
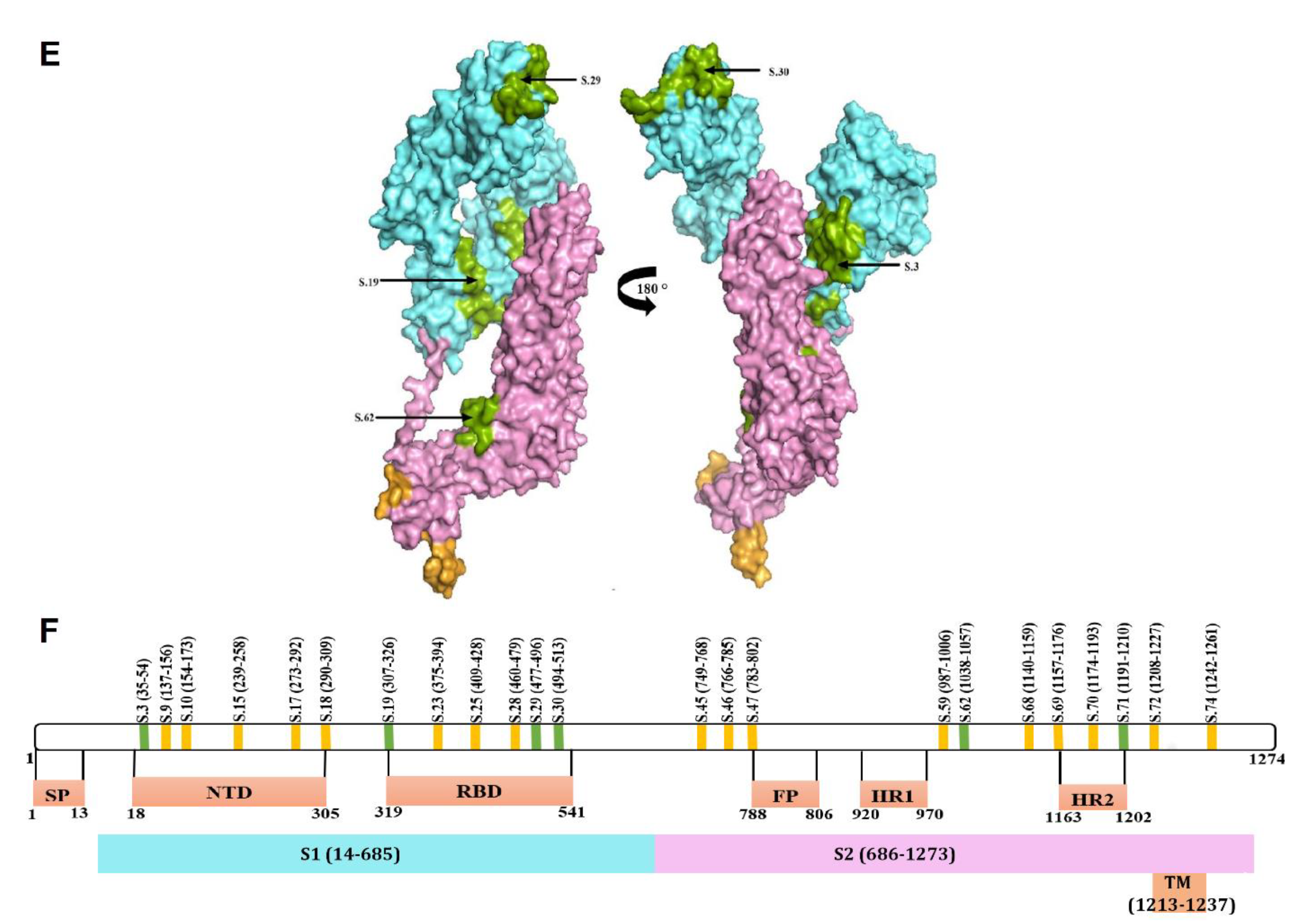
2.3. Mapping of Reactive Peptides on S Protein
2.4. Peptide Conservation Analysis
2.5. Mutational Analysis of Reactive Peptides
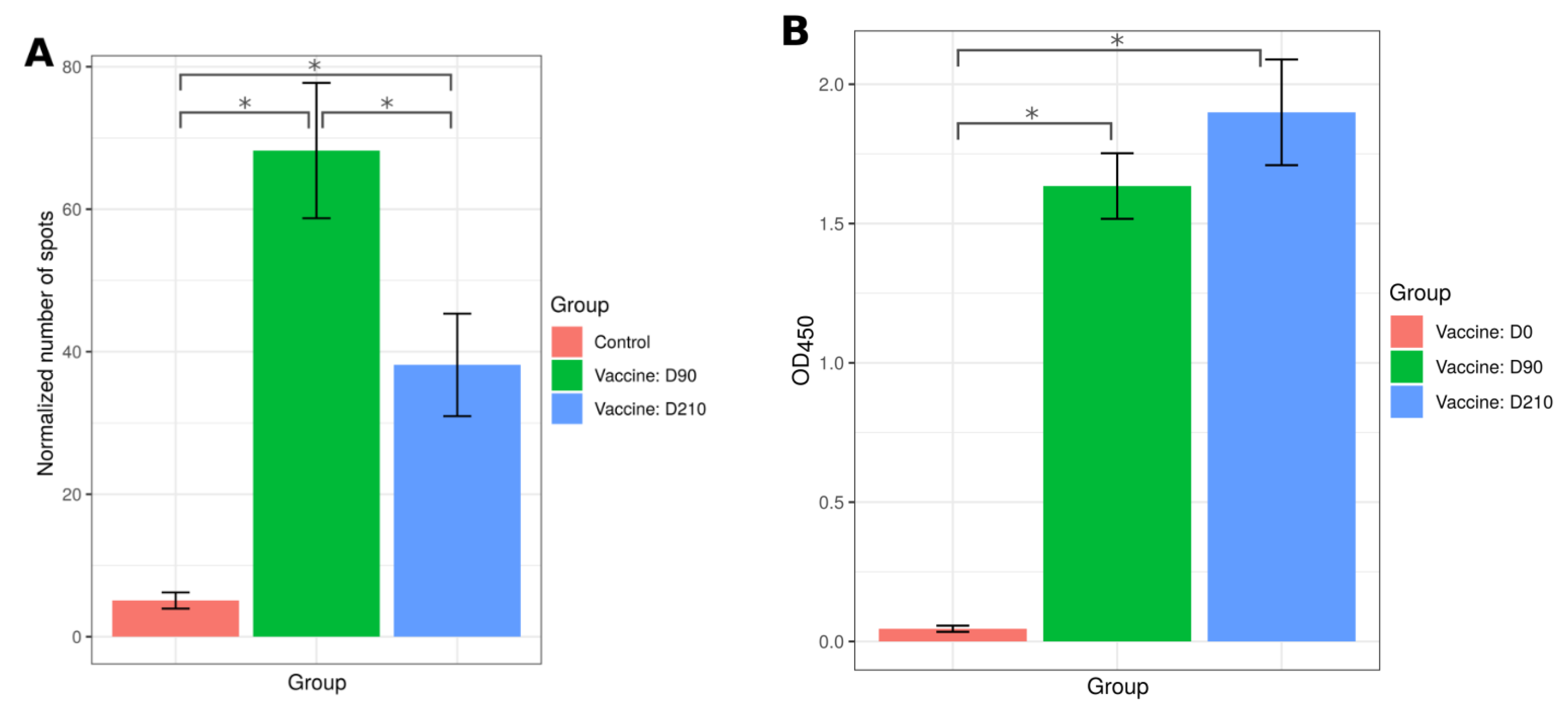
2.6. T Cell Immune Response to SputnikV Vaccine
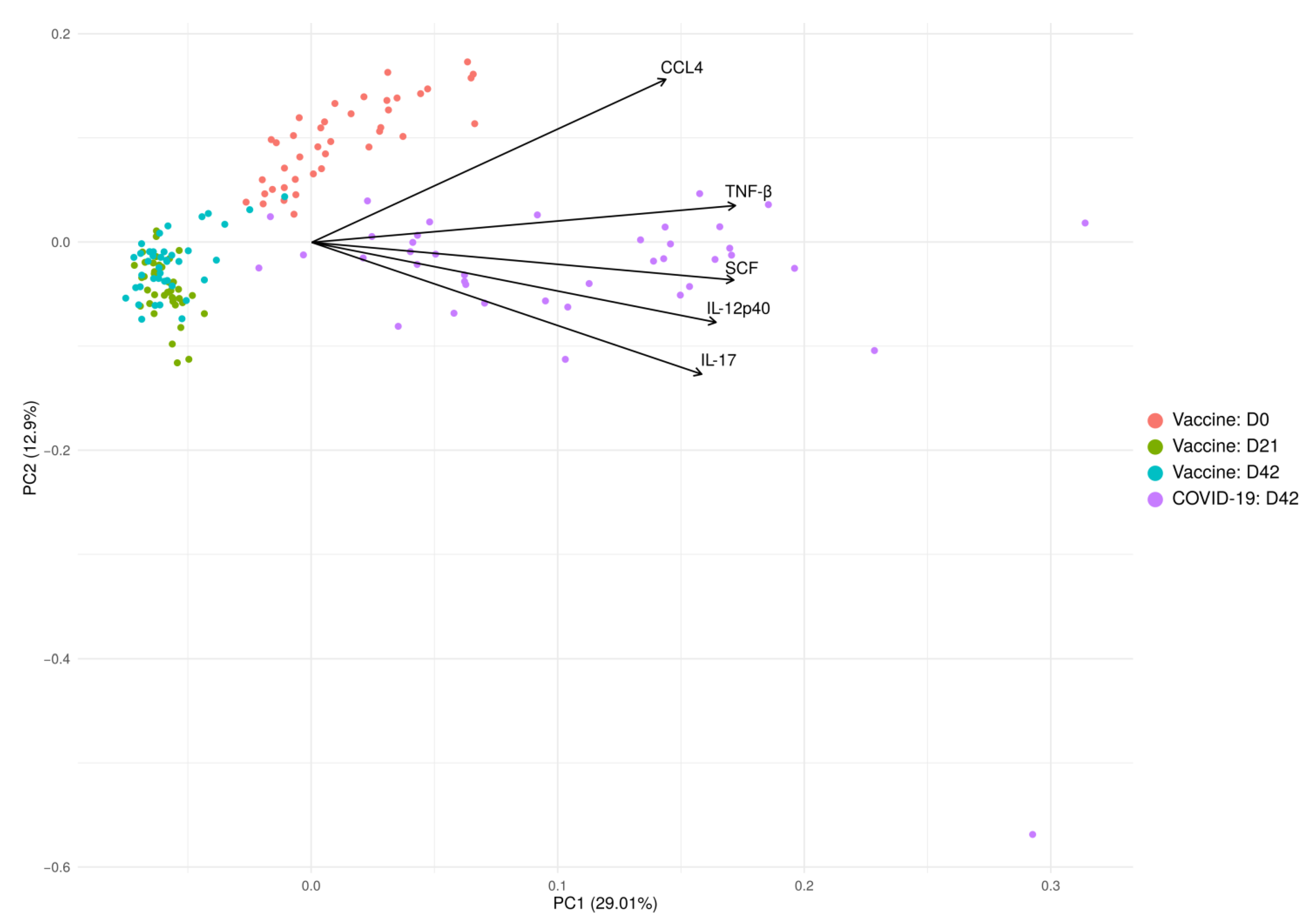
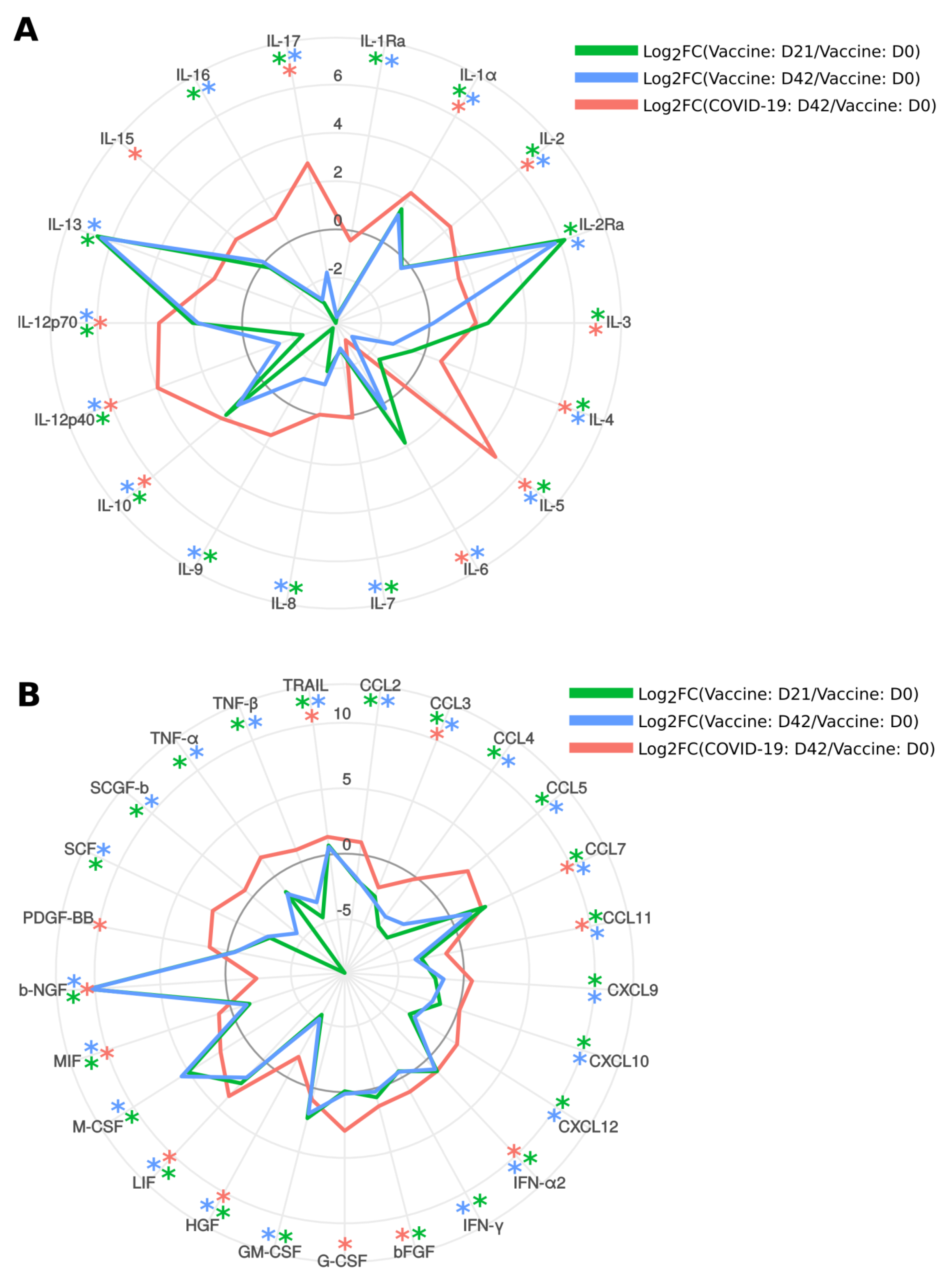
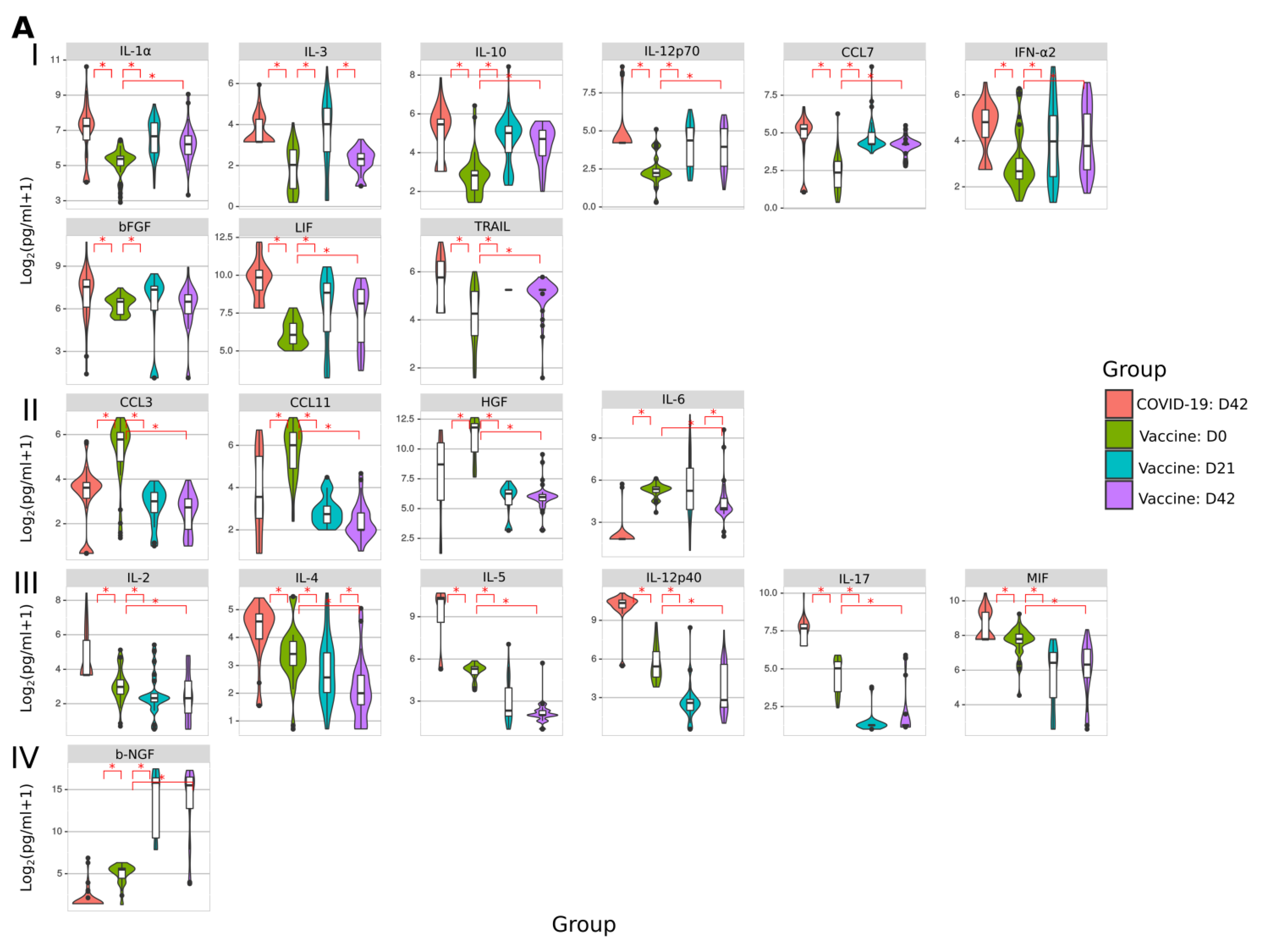
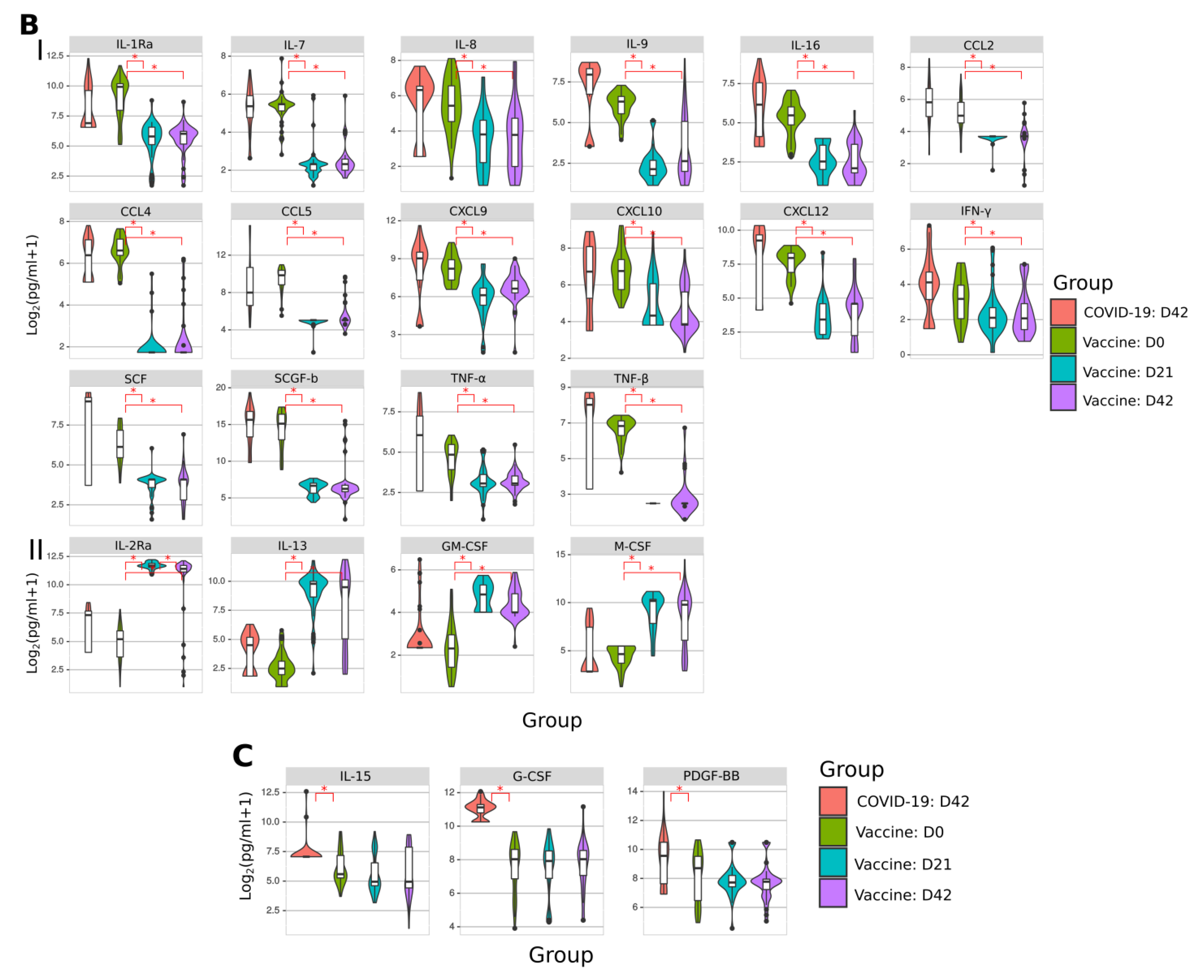
2.7. Cytokine Analysis
3. Discussion
4. Materials and Methods
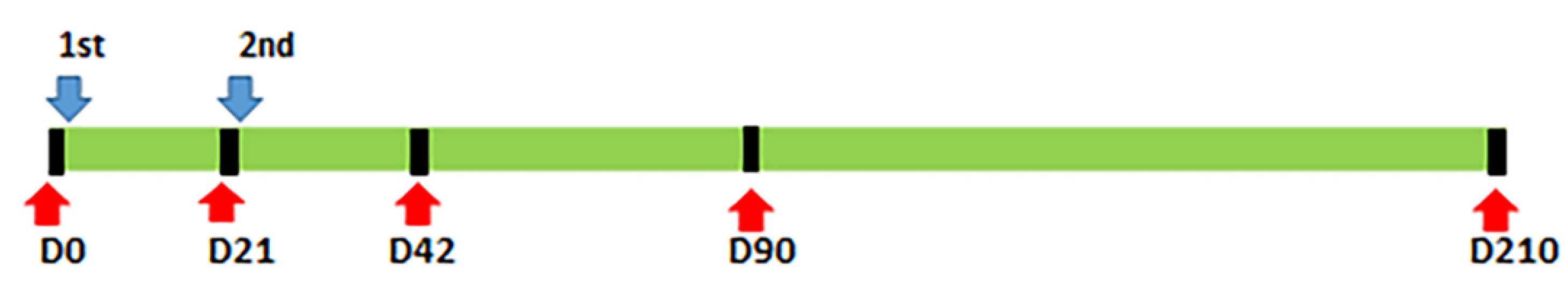
4.1. Subjects
4.2. Inclusion Criteria
4.3. Ethics Statement
4.4. COVID-19 ELISA
4.5. COVID-19 Peptide Reactivity
4.6. Peripheral Blood Mononuclear Cells (PBMCs) Isolation
4.7. ELISpot Analysis
4.8. Multiplex Analysis
4.9. Mapping of Reactive Peptides
4.10. Conservancy and Mutational Analysis of Reactive Peptides
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACE2 | angiotensin-converting enzyme 2 |
| COVID-19 | coronavirus disease 2019 |
| ELISA | enzyme-linked immunoassay |
| FP | Fusion Protein |
| GRCEM | Gamaleya National Research Centre for Epidemiology and Microbiology |
| HRs | Heptad Repeats |
| ICTV | International Committee on Taxonomy of Viruses |
| NTD | N-terminal Domain |
| RBD | Receptor Binding Domain |
| SARS-CoV-2 | severe acute respiratory syndrome coronavirus 2 |
References
- WHO. Director-General’s Opening Remarks at the Media Briefing on COVID-19—11 March 2020. Available online: https://www.who.int/director-general/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19---11-march-2020 (accessed on 12 January 2021).
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.-M.; Wang, W.; Song, Z.-G.; Hu, Y.; Tao, Z.-W.; Tian, J.-H.; Pei, Y.-Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 21 September 2021).
- Komissarov, A.B.; Safina, K.R.; Garushyants, S.K.; Fadeev, A.V.; Sergeeva, M.V.; Ivanova, A.A.; Danilenko, D.M.; Lioznov, D.; Shneider, O.V.; Shvyrev, N.; et al. Genomic epidemiology of the early stages of the SARS-CoV-2 outbreak in Russia. Nat. Commun. 2021, 12, 649. [Google Scholar] [CrossRef]
- Sanders, J.M.; Monogue, M.L.; Jodlowski, T.Z.; Cutrell, J.B. Pharmacologic Treatments for Coronavirus Disease 2019 (COVID-19): A Review. JAMA 2020, 323, 1824–1836. [Google Scholar] [CrossRef]
- Beigel, J.H.; Tomashek, K.M.; Dodd, L.E.; Mehta, A.K.; Zingman, B.S.; Kalil, A.C.; Hohmann, E.; Chu, H.Y.; Luetkemeyer, A.; Kline, S.; et al. Remdesivir for the Treatment of Covid-19—final Report. N. Engl. J. Med. 2020, 383, 1813–1836. [Google Scholar] [CrossRef]
- Lloyd, E.; Gandhi, T.; Petty, L. Monoclonal Antibodies for COVID-19. JAMA 2021, 325, 1015. [Google Scholar] [CrossRef]
- WHO Solidarity Trial Consortium Repurposed Antiviral Drugs for Covid-19—Interim WHO Solidarity Trial Results. N. Engl. J. Med. 2021, 384, 497–511. [CrossRef]
- Yao, X.; Ye, F.; Zhang, M.; Cui, C.; Huang, B.; Niu, P.; Liu, D. In Vitro Antiviral Activity and Projection of Optimized Dosing Design of Hydroxychloroquine for the Treatment of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2). Clin. Infect Dis. 2020, 71, 732–739. [Google Scholar] [CrossRef] [Green Version]
- Axfors, C.; Schmitt, A.M.; Janiaud, P.; van’t Hooft, J.; Abd-Elsalam, S.; Abdo, E.F.; Abella, B.S.; Akram, J.; Amaravadi, R.K.; Angus, D.C.; et al. Mortality outcomes with hydroxychloroquine and chloroquine in COVID-19 from an international collaborative meta-analysis of randomized trials. Nat. Commun. 2021, 12, 2349. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.-J.; Ni, Z.-Y.; Hu, Y.; Liang, W.-H.; Ou, C.-Q.; He, J.-X.; Liu, L.; Shan, H.; Lei, C.-L.; Hui, D.S.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- Young, B.E.; Ong, S.W.X.; Kalimuddin, S.; Low, J.G.; Tan, S.Y.; Loh, J. Epidemiologic Features and Clinical Course of Patients Infected With SARS-CoV-2 in Singapore. JAMA 2020, 323, 1488–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and safety of the mRNA-1273 SARS-CoV-2 vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Gruber, W.C. Safety and efficacy of the BNT162b2 mRNA Covid-19 vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Walsh, E.E.; Frenck, R.W.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based Covid-19 Vaccine Candidates. N. Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef]
- Capone, S.; Raggioli, A.; Gentile, M.; Battella, S.; Lahm, A.; Sommella, A.; Contino, A.M.; Urbanowicz, R.A.; Scala, R.; Barra, F.; et al. Immunogenicity of a new gorilla adenovirus vaccine candidate for COVID-19. Mol. Ther. 2021, 29, 2412–2423. [Google Scholar] [CrossRef] [PubMed]
- Ewer, K.J.; Barrett, J.R.; Belij-Rammerstorfer, S.; Sharpe, H.; Makinson, R.; Morter, R.; Flaxman, A.; Wright, D.; Bellamy, D.; Bittaye, M.; et al. T cell and antibody responses induced by a single dose of ChAdOx1 nCoV-19 (AZD1222) vaccine in a phase 1/2 clinical trial. Nat. Med. 2021, 27, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, M.N.; Minassian, A.M.; Ewer, K.J.; Flaxman, A.L.; Folegatti, P.M.; Owens, D.R. Safety and immunogenicity of ChAdOx1 nCoV-19 vaccine administered in a prime-boost regimen in young and old adults (COV002): A single-blind, randomised, controlled, phase 2/3 trial. Lancet 2020, 396, 1979–1993. [Google Scholar] [CrossRef]
- Logunov, D.Y.; Dolzhikova, L.V.; Shcheblyakov, D.V.; Tukhvatulin, A.I.; Zubkova, O.V.; Dzharullaeva, A.S.; Kovyrshina, K.V.; Nadezhda, L.L.; Grousova, D.M.; Erokhova, A.S. Safety and efficacy of a rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine: An interim analysis of a randomized controlled phase 3 trial in Russia. Lancet 2021, 397, 671–681. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, C.; Xu, X.F.; Xu, W.; Liu, S.W. Structural and functional properties of SARS-CoV-2 spike protein: Potential antivirus drug development for COVID-19. Acta Pharm. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef]
- Li, C.; Tian, X.; Jia, X.; Wan, J.; Lu, L.; Jiang, S.; Lan, F.; Lu, Y.; Wu, Y.; Ying, T. The impact of receptor-binding domain natural mutations on antibody recognition of SARS-CoV-2. Signal Transduct. Target. Ther. 2021, 6, 132. [Google Scholar] [CrossRef]
- Baraniuk, C. Covid-19: What do we know about airborne transmission of SARS-CoV-2? BMJ 2021, 373, n1030. [Google Scholar] [CrossRef]
- Reuters. COVID-19 TRACKER. 2021. Available online: https://graphics.reuters.com/world-coronavirus-tracker-and-maps/countries-and-territories/russia/ (accessed on 6 April 2021).
- CRC. The Race to Vaccinate the World. 2021. Available online: https://coronavirus.jhu.edu/vaccines/international (accessed on 8 June 2021).
- Statista. Number of Doses of the COVID-19 Vaccine Sputnik V Ordered from Russia or Agreed to Be produced Abroad as of 25 May 2021, by Country. 2021. Available online: https://www.statista.com/statistics/1123927/sputnik-v-exports-from-russia-by-country/ (accessed on 5 March 2021).
- Zhu, W.; Wang, C.; Wang, B.-Z. From Variation of Influenza Viral Proteins to Vaccine Development. Int. J. Mol. Sci. 2017, 18, 1554. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.T.; Burgers, W.A. SARS-CoV-2 evolution and vaccines: Cause for concern? Lancet Respir. Med. 2021, 9, 333–335. [Google Scholar] [CrossRef]
- Callaway, E. Delta coronavirus variant: Scientists brace for impact. Nature 2021, 595, 17–18. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, S. T--cell--inducing vaccines–what’s the future. Immunology 2012, 135, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Slota, M.; Lim, J.-B.; Dang, Y.; Disis, M.L. ELISpot for measuring human immune responses to vaccines. Expert Rev. Vaccines 2011, 10, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Et Biophys. Acta (BBA)—Bioenerg. 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Carrillo, J.L.; Contreras-Cordero, J.F.; Gutiérrez-Coronado, O.; Villalobos-Gutiérrez, P.T.; Ramos-Gracia, L.G.; Hernández-Reyes, V.E. Cytokine Profiling Plays a Crucial Role in Activating Immune System to Clear Infectious Pathogens; IntechOpen: London, UK, 2019. [Google Scholar]
- Furman, D.; Davis, M.M. New approaches to understanding the immune response to vaccination and infection. Vaccine 2015, 33, 5271–5281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudd, P.A.; Crawford, J.C.; Turner, J.S.; Souquette, A.; Reynolds, D.; Bender, D.; Bosanquet, J.P.; Anand, N.J.; Striker, D.A.; Martin, R.S.; et al. Distinct inflammatory profiles distinguish COVID-19 from influenza with limited contributions from cytokine storm. Sci. Adv. 2020, 6, eabe3024. [Google Scholar] [CrossRef]
- Skibinski, D.A.G.; Jones, L.A.; Zhu, Y.O.; Xue, L.W.; Au, B.; Lee, B.; Naim, A.N.M.; Lee, A.; Kaliaperumal, N.; Low, J.G.H.; et al. Induction of Human T-cell and Cytokine Responses Following Vaccination with a Novel Influenza Vaccine. Sci. Rep. 2018, 8, 18007. [Google Scholar] [CrossRef] [PubMed]
- Duan, K.; Liu, B.; Li, C.; Zhang, H.; Yu, T.; Qu, J.; Zhou, M.; Chen, L.; Meng, S.; Hu, Y.; et al. Effectiveness of convalescent plasma therapy in severe COVID-19 patients. Proc. Natl. Acad. Sci. USA 2020, 117, 9490–9496. [Google Scholar] [CrossRef] [Green Version]
- Logunov, D.Y.; Dolzhikova, I.V.; Zubkova, O.V.; Tukhvatullin, A.I.; Shcheblyakov, D.V.; Dzharullaeva, A.S. Safety and immunogenicity of a rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine in two formulations: Two open, non-randomized phase 1/2 studies from Russia. Lancet 2020, 396, 887–897. [Google Scholar] [CrossRef]
- Wong, S.K.; Li, W.; Moore, M.J.; Choe, H.; Farzan, M. A 193-amino acid fragment of the SARS coronavirus S protein efficiently binds angiotensin-converting enzyme 2. J. Biol. Chem. 2004, 279, 3197–3201. [Google Scholar] [CrossRef] [Green Version]
- Robbiani, D.F.; Gaebler, C.; Muecksch, F.; Lorenzi, J.C.; Wang, Z.; Cho, A.; Nussenzweig, M.C. Convergent antibody responses to SARS-CoV-2 in convalescent individuals. Nature 2020, 584, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Shan, C.; Duan, X.; Chen, Z.; Liu, P.; Song, J.; Yan, J. A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2. Nature 2020, 584, 120–124. [Google Scholar] [CrossRef]
- Brink, E.N.V.D.; ter Meulen, J.; Cox, F.; Jongeneelen, M.A.C.; Thijsse, A.; Throsby, M.; Marissen, W.E.; Rood, P.M.L.; Bakker, A.B.H.; Gelderblom, H.R.; et al. Molecular and Biological Characterization of Human Monoclonal Antibodies Binding to the Spike and Nucleocapsid Proteins of Severe Acute Respiratory Syndrome Coronavirus. J. Virol. 2005, 79, 1635–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Z.; Liu, L.; Du, L.; Zhang, C.; Jiang, S.; Li, T.; He, Y. Potent and persistent antibody responses against the receptor-binding domain of SARS-CoV spike protein in recovered patients. Virol. J. 2010, 7, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerutti, G.; Guo, Y.; Zhou, T.; Gorman, J.; Lee, M.; Rapp, M.; Reddem, E.R.; Yu, J.; Bahna, F.; Bimela, J.; et al. Potent SARS-CoV-2 neutralizing antibodies directed against spike N-terminal domain target a single supersite. Cell Host Microbe 2021, 29, 819–833. [Google Scholar] [CrossRef]
- Zhou, H.; Chen, Y.; Zhang, S.; Niu, P.; Qin, K.; Jia, W.; Wang, X. Structural definition of a neutralization epitope on the N-terminal domain of MERS-CoV spike glycoprotein. Nat. Commun. 2019, 10, 3068. [Google Scholar] [CrossRef]
- Tang, T.; Bidon, M.; Jaimes, J.A.; Whittaker, G.R.; Daniel, S. Coronavirus membrane fusion mechanism offers a potential target for antiviral development. Antivir. Res. 2020, 178, 104792. [Google Scholar] [CrossRef]
- Ou, X.; Zheng, W.; Shan, Y.; Mu, Z.; Dominguez, S.R.; Holmes, K.V.; Qian, Z. Identification of the Fusion Peptide-Containing Region in Betacoronavirus Spike Glycoproteins. J. Virol. 2016, 90, 5586–5600. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Zhu, Y.; Liu, M.; Lan, Q.; Xu, W.; Wu, Y.; Ying, T.; Liu, S.; Shi, Z.; Jiang, S.; et al. Fusion mechanism of 2019-nCoV and fusion inhibitors targeting HR1 domain in spike protein. Cell. Mol. Immunol. 2020, 17, 765–767. [Google Scholar] [CrossRef]
- Trigueiro-Louro, J.; Correia, V.; Figueiredo-Nunes, I.; Gíria, M.; Rebelo-de-Andrade, H. Unlocking COVID therapeutic targets: A structure-based rationale against SARS-CoV-2, SARS-CoV and MERS-CoV Spike. Comput. Struct. Biotechnol. J. 2020, 18, 2117–2131. [Google Scholar] [CrossRef]
- Xu, Y.; Zhu, J.; Liu, Y.; Lou, Z.; Yua, F.; Liu, Y.; Cole, D.K.; Ni, L.; Su, N.; Qin, L.; et al. Characterization of the Heptad Repeat Regions, HR1 and HR2, and Design of a Fusion Core Structure Model of the Spike Protein from Severe Acute Respiratory Syndrome (SARS) Coronavirus. Biochemistry 2004, 43, 14064–14071. [Google Scholar]
- Montelongo-Jauregui, D.; Vila, T.; Sultan, A.S.; Jabra-Rizk, M.A. Convalescent serum therapy for COVID-19: A 19th century remedy for a 21st century disease. PLOS Pathog. 2020, 16, e1008735. [Google Scholar] [CrossRef]
- Kumar, N.; Kumar, S.; Patel, S. Convalescent Plasma Therapy In Critically Ill Patients With Late Stage Covid-19. Chest 2020, 158, A601. [Google Scholar] [CrossRef]
- Dispinseri, S.; Secchi, M.; Pirillo, M.F.; Tolazzi, M.; Borghi, M.; Brigatti, C.; De Angelis, M.L.; Baratella, M.; Bazzigaluppi, E.; Venturi, G.; et al. Neutralizing antibody responses to SARS-CoV-2 in symptomatic COVID-19 is persistent and critical for survival. Nat. Commun. 2021, 12, 2670. [Google Scholar] [CrossRef]
- He, Y.; Li, J.; Jiang, S. A single amino acid substitution (R441A) in the receptor-binding domain of SARS coronavirus spike protein disrupts the antigenic structure and binding activity. Biochem. Biophys. Res. Commun. 2006, 344, 106–113. [Google Scholar] [CrossRef]
- Mitsuki, Y.Y.; Ohnishi, K.; Takagi, H.; Oshima, M.; Yamamoto, T.; Mizukoshi, F. A single amino acid substitution in the S1 and S2 Spike protein domains determines the neutralization escape phenotype of SARS-CoV. Microbes Infect. 2008, 10, 908–915. [Google Scholar] [CrossRef]
- Korber, B.; Fischer, W.M.; Gnanakaran, S.; Yoon, H.; Theiler, J.; Abfalterer, W.; Foley, B.; Giorgi, E.E.; Bhattacharya, T.; Parker, M.D.; et al. Spike mutation pipeline reveals the emergence of a more transmissible form of SARS-CoV-2. BioRxiv 2020. [Google Scholar] [CrossRef]
- Galloway, S.E.; Paul, P.; MacCannell, D.R.; Johansson, M.A.; Brooks, J.T.; MacNeil, A.; Slayton, R.B.; Tong, S.; Silk, B.J.; Armstrong, G.L.; et al. Emergence of SARS-CoV-2 B.1.1.7 Lineage—United States, 29 December 2020–12 January 2021. Morb. Mortal. Wkly. Rep. 2021, 70, 95. [Google Scholar] [CrossRef] [PubMed]
- Rubin, R. COVID-19 Vaccines vs Variants—Determining How Much Immunity Is Enough. JAMA 2021, 325, 1241. [Google Scholar] [CrossRef]
- Agrawal, L.; Poullikkas, T.; Eisenhower, S.; Monsanto, C.; Bakku, R.K.; Chen, M.H.; Kalra, R.S. Bioinformatics-Based Analysis of SARS-CoV-2 Core Proteins for Potential Therapeutic Targets. Antibodies 2021, 10, 3. [Google Scholar] [CrossRef]
- Lei, K.; Zhang, X. Conservation analysis of SARS-CoV-2 spike suggests complicated viral adaptation history from bat to human. Evol. Med. Public Health 2020, 2020, 290–303. [Google Scholar] [CrossRef] [PubMed]
- GISAID. Tracking of Variants. 2021. Available online: https://www.gisaid.org/hcov19-variants/ (accessed on 24 April 2021).
- Grifoni, A.; Sidney, J.; Zhang, Y.; Scheuermann, R.H.; Peters, B.; Sette, A. A sequence homology and bioinformatic approach can predict candidate targets for immune responses to SARS-CoV-2. Cell Host Microbe 2020, 27, 671–680.e672. [Google Scholar] [CrossRef] [PubMed]
- Shrock, E.; Fujimura, E.; Kula, T.; Timms, R.T.; Lee, I.H.; Leng, Y.; Elledge, S.J. MGH COVID-19 Collection & Processing Team Viral epitope profiling of COVID-19 patients reveals cross-reactivity and correlates of severity. Science 2020, 370, 4250. [Google Scholar]
- Schmidt, M.E.; Varga, S.M. The CD8 T Cell Response to Respiratory Virus Infections. Front. Immunol. 2018, 9, 678. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Mentzer, A.J.; Liu, G.; Yao, X.; Yin, Z.; Dong, D.; Dong, T. Broad and strong memory CD4+ and CD8+ T cells induced by SARS-CoV-2 in UK convalescent individuals following COVID-19. Nat. Immunol. 2020, 21, 1336–1345. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T cell responses to SARS-CoV-2 coronavirus in humans with COVID-19 disease and unexposed individuals. Cell 2020, 181, 1489–1501.e1415. [Google Scholar] [CrossRef]
- Ni, L.; Ye, F.; Cheng, M.L.; Feng, Y.; Deng, Y.Q.; Zhao, H.; Dong, C. Detection of SARS-CoV-2-specific humoral and cellular immunity in COVID-19 convalescent individuals. Immunity 2020, 52, 971–977.r973. [Google Scholar] [CrossRef]
- Goel, R.R.; Painter, M.M.; Apostolidis, S.A.; Mathew, D.; Meng, W.; Rosenfeld, A.M.; Lundgreen, K.A.; Reynaldi, A.; Khoury, D.S.; Pattekar, A.; et al. mRNA Vaccination Induces Durable Immune Memory to SARS-CoV-2 with Continued Evolution to Variants of Concern. Biorxiv 2021. [Google Scholar] [CrossRef]
- Urbanowicz, R.A.; Tsoleridis, T.; Jackson, H.J.; Cusin, L.; Duncan, J.D.; Chappell, J.G.; Ollivere, B.J. Two doses of the SARS-CoV-2 BNT162b2 vaccine enhances antibody responses to variants in individuals with prior SARS-CoV-2 infection. Sci. Transl. Med. 2021, 13, 253. [Google Scholar] [CrossRef]
- González, S.; Olszevicki, S.; Salazar, M.; Calabria, A.; Regairaz, L.; Marín, L.; Campos, P.; Varela, T.; Martínez, V.V.G.; Ceriani, L.; et al. Effectiveness of the first component of Gam-COVID-Vac (Sputnik V) on reduction of SARS-CoV-2 confirmed infections, hospitalisations and mortality in patients aged 60-79: A retrospective cohort study in Argentina. EClinicalMedicine 2021, 40, 101126. [Google Scholar] [CrossRef]
- Rossi, A.H.; Ojeda, D.S.; Varese, A.; Sanchez, L.; Ledesma, M.M.G.L.; Mazzitelli, I.; Juliá, A.A.; Rouco, S.O.; Pallarés, H.M.; Navarro, G.S.C.; et al. Sputnik V Vaccine Elicits Seroconversion and Neutralizing Capacity to SARS CoV-2 after a Single Dose. Cell Rep. Med. 2021, 2, 100359. [Google Scholar] [CrossRef] [PubMed]
- Claro, F.; Silva, D.; Rodriguez, M.; Rangel, R.; de Waard, J.H. IgG Antibody response to the Sputnik V vaccine: Previous SARS-CoV-2 seropositive individuals might need just one vaccine dose. Int. J. Infect. Dis. 2021, 111, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Waltuch, T.; Gill, P.; Zinns, L.E.; Whitney, R.; Tokarski, J.; Tsung, J.W.; Sanders, J.E. Features of COVID-19 post-infectious cytokine release syndrome in children presenting to the emergency department. Am. J. Emerg. Med. 2020, 38, 2246.e3–2246.e6. [Google Scholar] [CrossRef]
- Holdsworth, S.R.; Gan, P.Y. Cytokines: Names and Numbers You Should Care About. Clin. J. Am. Soc. Nephrol. 2015, 10, 2243–2254. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Soh, W.T.; Kishikawa, J.I.; Hirose, M.; Nakayama, E.E.; Li, S.; Arase, H. An infectivity-enhancing site on the SARS-CoV-2 spike protein targeted by antibodies. Cell 2021, 184, 3452–3466.e18. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Fan, J.; Yu, F.; Feng, B.; Lou, B.; Zou, Q.; Liang, T. Viral load dynamics and disease severity in patients infected with SARS-CoV-2 in Zhejiang province, China, January-March 2020: Retrospective cohort study. BMJ 2020, 369, m1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawley, S.; Hawley, G. Flow Cytometry Protocols, 2nd ed.; Methods in Molecular Biology: 2004; © Humana Press Inc.: Totowa, NJ, USA, 2004; Volume 263, pp. 1–425. [Google Scholar]
- Dessau, R.B.; Pipper, C.B. [“R”--project for statistical computing]. Ugeskr. Laeger 2008, 170, 328–330. [Google Scholar] [PubMed]
- Wickham, H. ggplot2. Wiley Interdiscip. Rev. Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Tang, Y.; Horikoshi, M.; Li, W. ggfortify: Unified Interface to Visualize Statistical Results of Popular R Packages. R J. 2016, 8, 474–485. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptides (n) | Peptide Name | Strains with 100% Identity (n) | Conservation (%) when 52 Strains Were Included |
|---|---|---|---|
| 12 | S16, S17, S18, S19, S35, S47, S48, S59, S62, S65, S68, S72 | 52 | 100.0 |
| 4 | S45, S69, S70, S71 | 51 | 98.1 |
| 6 | S11, S21, S23, S24, S46, S74 | 50 | 96.2 |
| 2 | S25, S66 | 49 | 94.20 |
| 2 | S39, S56 | 48 | 92.30 |
| 1 | S3 | 47 | 90.4 |
| 1 | S.15 | 44 | 84.6 |
| 5 | S10, S28, S30, S42 | 43 | 82.7 |
| 1 | S.9 | 41 | 78.8 |
| 1 | S27 | 39 | 75.00 |
| 1 | S29 | 32 | 61.50 |
| 1 | S37 | 3 | 5.80 |
| All Reactive Peptides | Vaccinated and COVID-19 | COVID-19 | Vaccinated | |||||
|---|---|---|---|---|---|---|---|---|
| Groups | Conservation (%) | Peptides (n) | Peptides | n | Peptides | n | Peptides | n |
| 1 | 100 | 12 | S17, S18, S47, S59, S68, S72 | 6 | S19, S62 | 2 | S16, S35, S48, S65 | 4 |
| 2 | >90 < 100 | 15 | S23, S25, S45, S46, S69, S70, S74 | 7 | S3, S71 | 2 | S11, S21, S24, S39, S56, S66 | 6 |
| 3 | >80 < 90 | 5 | S10, S15, S28 | 3 | S30 | 1 | S42 | 1 |
| 4 | >70 < 80 | 2 | S9 | 1 | 0 | S27 | 1 | |
| 5 | >60 < 70 | 1 | 0 | S29 | 1 | 0 | ||
| 6 | <60 | 1 | 0 | 0 | S37 | 1 | ||
| Vaccinated | No. of Mutated Positions | Convalescent | No. of Mutated Positions | Vaccinated and Convalescent | No. of Mutated Positions |
|---|---|---|---|---|---|
| S11 | 2 | S3 | 2 | S9 | 7 |
| S21 | 1 | S29 | 5 | S10 | 3 |
| S24 | 2 | S30 | 2 | S15 | 2 |
| S27 | 2 | S71 | 1 | S23 | 2 |
| S37 | 4 | S25 | 2 | ||
| S39 | 3 | S28 | 2 | ||
| S42 | 2 | S45 | 1 | ||
| S56 | 1 | S46 | 2 | ||
| S66 | 2 | S69 | 1 | ||
| S70 | 1 | ||||
| S74 | 2 |
| Clinical Characteristics | Values |
|---|---|
| Vaccinated age (years) | 43.3 ± 16.4 |
| Vaccinated sex (M/F) | 17/23 |
| Convalescent COVID-19 age (years) | 39.1 ± 13.2 |
| Convalescent COVID-19 age (M/F) | 15/25 |
| Control age (years) | 47.1 ± 13.8 |
| Control sex (M/F) | 5/7 |
| Peptide | Aa Sequence | Position | Peptide | Aa Sequence | Position | Peptide | Aa Sequence | Position |
|---|---|---|---|---|---|---|---|---|
| S1 | MFVFLVLLPLVSSQCVNLTT | 1–20 | S25 | QIAPGQTGKIADYNYKLPDD | 409–428 | S50 | IKQYGDCLGDIAARDLICAQ | 834–853 |
| S2 | LTTRTQLPPAYTNSFTRGVY | 18–37 | S26 | PDDFTGCVIAWNSNNLDSKV | 426–445 | S51 | CAQKFNGLTVLPPLLTDEMI | 851–870 |
| S3 | GVYYPDKVFRSSVLHSTQDL | 35–54 | S27 | SKVGGNYNYLYRLFRKSNLK | 443–462 | S52 | EMIAQYTSALLAGTITSGWT | 868–887 |
| S4 | QDLFLPFFSNVTWFHAIHVS | 52–71 | S28 | NLKPFERDISTEIYQAGSTP | 460–479 | S53 | GWTFGAGAALQIPFAMQMAY | 885–904 |
| S5 | HVSGTNGTKRFDNPVLPFND | 69–88 | S29 | STPCNGVEGFNCYFPLQSYG | 477–496 | S54 | MAYRFNGIGVTQNVLYENQK | 902–921 |
| S6 | FNDGVYFASTEKSNIIRGWI | 86–105 | S30 | SYGFQPTNGVGYQPYRVVVL | 494–513 | S55 | NQKLIANQFNSAIGKIQDSL | 919–938 |
| S8 | VNNATNVVIKVCEFQFCNDP | 120–139 | S31 | VVLSFELLHAPATVCGPKKS | 511–530 | S56 | DSLSSTASALGKLQDVVNQN | 936–955 |
| S9 | NDPFLGVYYHKNNKSWMESE | 137–156 | S32 | KKSTNLVKNKCVNFNFNGLT | 528–547 | S57 | NQNAQALNTLVKQLSSNFGA | 953–972 |
| S10 | ESEFRVYSSANNCTFEYVSQ | 154–173 | S33 | GLTGTGVLTESNKKFLPFQQ | 545–564 | S58 | FGAISSVLNDILSRLDKVEA | 970–989 |
| S11 | VSQPFLMDLEGKQGNFKNLR | 171–190 | S34 | FQQFGRDIADTTDAVRDPQT | 562–581 | S59 | VEAEVQIDRLITGRLQSLQT | 987–1006 |
| S12 | NLREFVFKNIDGYFKIYSKH | 188–207 | S35 | PQTLEILDITPCSFGGVSVI | 579–598 | S60 | LQTYVTQQLIRAAEIRASAN | 1004–1023 |
| S13 | SKHTPINLVRDLPQGFSALE | 205–224 | S36 | SVITPGTNTSNQVAVLYQDV | 596–615 | S61 | SANLAATKMSECVLGQSKRV | 1021–1040 |
| S14 | ALEPLVDLPIGINITRFQTL | 222–241 | S37 | QDVNCTEVPVAIHADQLTPT | 613–632 | S62 | KRVDFCGKGYHLMSFPQSAP | 1038–1057 |
| S15 | QTLLALHRSYLTPGDSSSGW | 239–258 | S38 | TPTWRVYSTGSNVFQTRAGC | 630–649 | S63 | SAPHGVVFLHVTYVPAQEKN | 1055–1074 |
| S16 | SGWTAGAAAYYVGYLQPRTF | 256–275 | S39 | AGCLIGAEHVNNSYECDIPI | 647–666 | S64 | EKNFTTAPAICHDGKAHFPR | 1072–1091 |
| S17 | RTFLLKYNENGTITDAVDCA | 273–292 | S41 | PRRARSVASQSIIAYTMSLG | 681–700 | S65 | FPREGVFVSNGTHWFVTQRN | 1089–1108 |
| S18 | DCALDPLSETKCTLKSFTVE | 290–309 | S42 | SLGAENSVAYSNNSIAIPTN | 698–717 | S66 | QRNFYEPQIITTDNTFVSGN | 1106–1125 |
| S19 | TVEKGIYQTSNFRVQPTESI | 307–326 | S43 | PTNFTISVTTEILPVSMTKT | 715–734 | S68 | PLQPELDSFKEELDKYFKNH | 1140–1159 |
| S20 | ESIVRFPNITNLCPFGEVFN | 324–343 | S45 | CSNLLLQYGSFCTQLNRALT | 749–768 | S69 | KNHTSPDVDLGDISGINASV | 1157–1176 |
| S21 | VFNATRFASVYAWNRKRISN | 341–360 | S46 | ALTGIAVEQDKNTQEVFAQV | 766–785 | S70 | ASVVNIQKEIDRLNEVAKNL | 1174–1193 |
| S22 | ISNCVADYSVLYNSASFSTF | 358–377 | S47 | AQVKQIYKTPPIKDFGGFNF | 783–802 | S71 | KNLNESLIDLQELGKYEQYI | 1191–1210 |
| S23 | STFKCYGVSPTKLNDLCFTN | 375–394 | S48 | FNFSQILPDPSKPSKRSFIE | 800–819 | S72 | QYIKWPWYIWLGFIAGLIAI | 1208–1227 |
| S24 | FTNVYADSFVIRGDEVRQIA | 392–411 | S49 | FIEDLLFNKVTLADAGFIKQ | 817–836 | S74 | SCLKGCCSCGSCCKFDEDDS | 1242–1261 |
| S75 | CKFDEDDSEPVLKGVKLHYT | 1254–1273 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martynova, E.; Hamza, S.; Garanina, E.E.; Kabwe, E.; Markelova, M.; Shakirova, V.; Khaertynova, I.M.; Kaushal, N.; Baranwal, M.; Rizvanov, A.A.; et al. Long Term Immune Response Produced by the SputnikV Vaccine. Int. J. Mol. Sci. 2021, 22, 11211. https://doi.org/10.3390/ijms222011211
Martynova E, Hamza S, Garanina EE, Kabwe E, Markelova M, Shakirova V, Khaertynova IM, Kaushal N, Baranwal M, Rizvanov AA, et al. Long Term Immune Response Produced by the SputnikV Vaccine. International Journal of Molecular Sciences. 2021; 22(20):11211. https://doi.org/10.3390/ijms222011211
Chicago/Turabian StyleMartynova, Ekaterina, Shaimaa Hamza, Ekaterina E. Garanina, Emmanuel Kabwe, Maria Markelova, Venera Shakirova, Ilsiyar M. Khaertynova, Neha Kaushal, Manoj Baranwal, Albert A. Rizvanov, and et al. 2021. "Long Term Immune Response Produced by the SputnikV Vaccine" International Journal of Molecular Sciences 22, no. 20: 11211. https://doi.org/10.3390/ijms222011211
APA StyleMartynova, E., Hamza, S., Garanina, E. E., Kabwe, E., Markelova, M., Shakirova, V., Khaertynova, I. M., Kaushal, N., Baranwal, M., Rizvanov, A. A., Urbanowicz, R. A., & Khaiboullina, S. F. (2021). Long Term Immune Response Produced by the SputnikV Vaccine. International Journal of Molecular Sciences, 22(20), 11211. https://doi.org/10.3390/ijms222011211