
Anti-Allergic and Anti-Inflammatory Effects of Neferine on RBL-2H3 Cells


 ,
, 
Abstract
:1. Introduction
2. Results
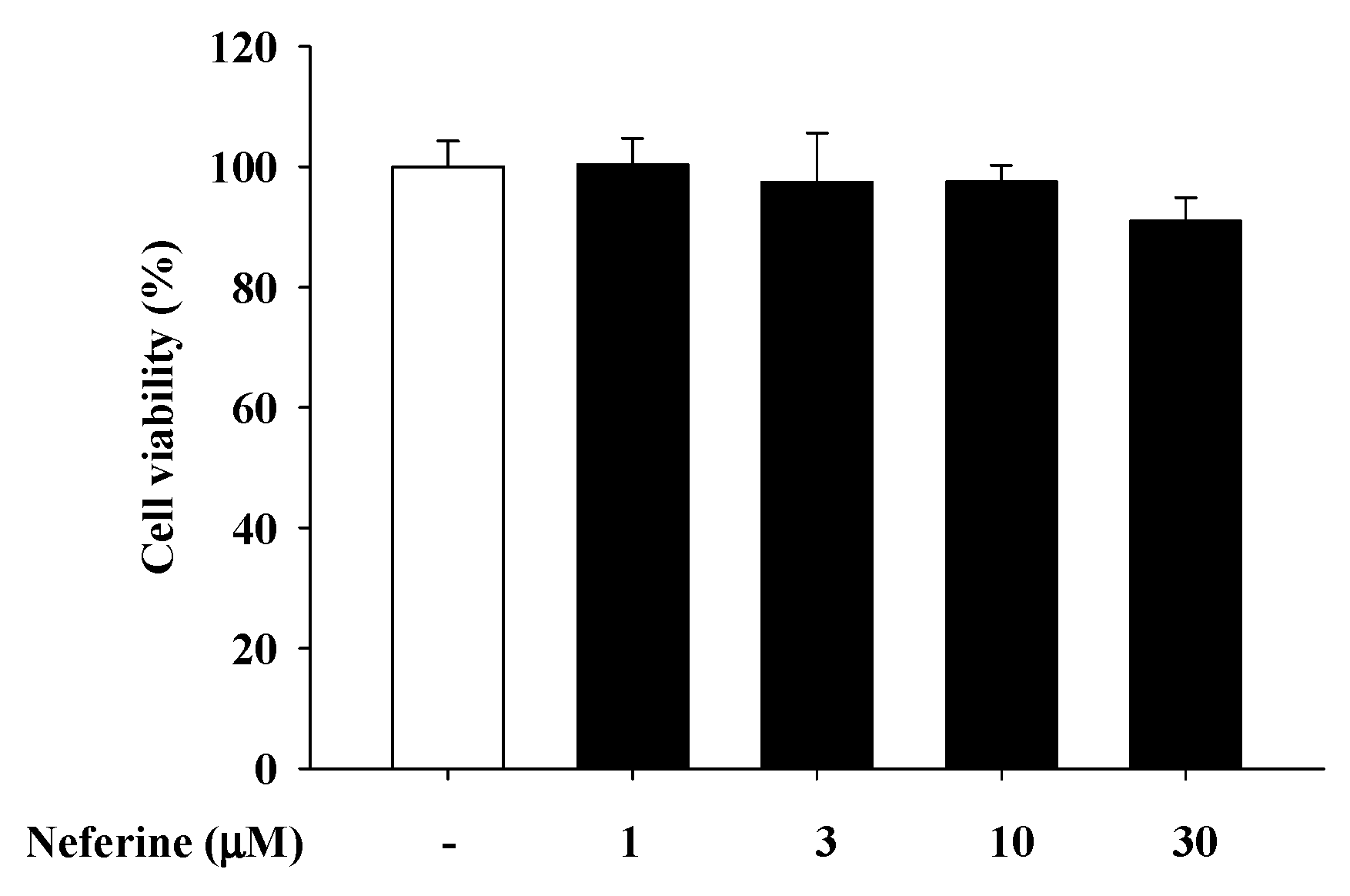
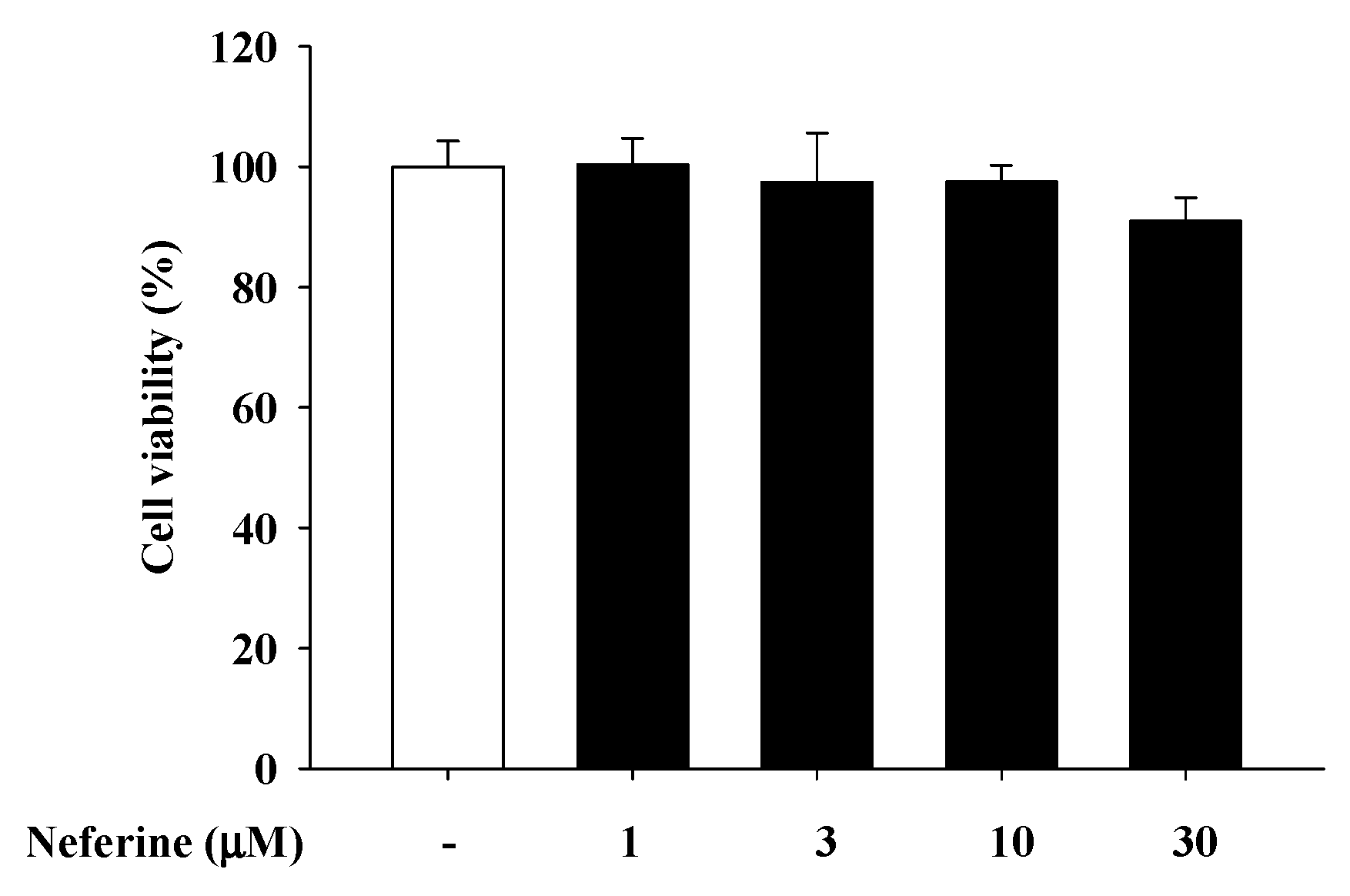
2.1. Neferine Cytotoxicity Test on Rat Mast CelLs (RBL-2H3)
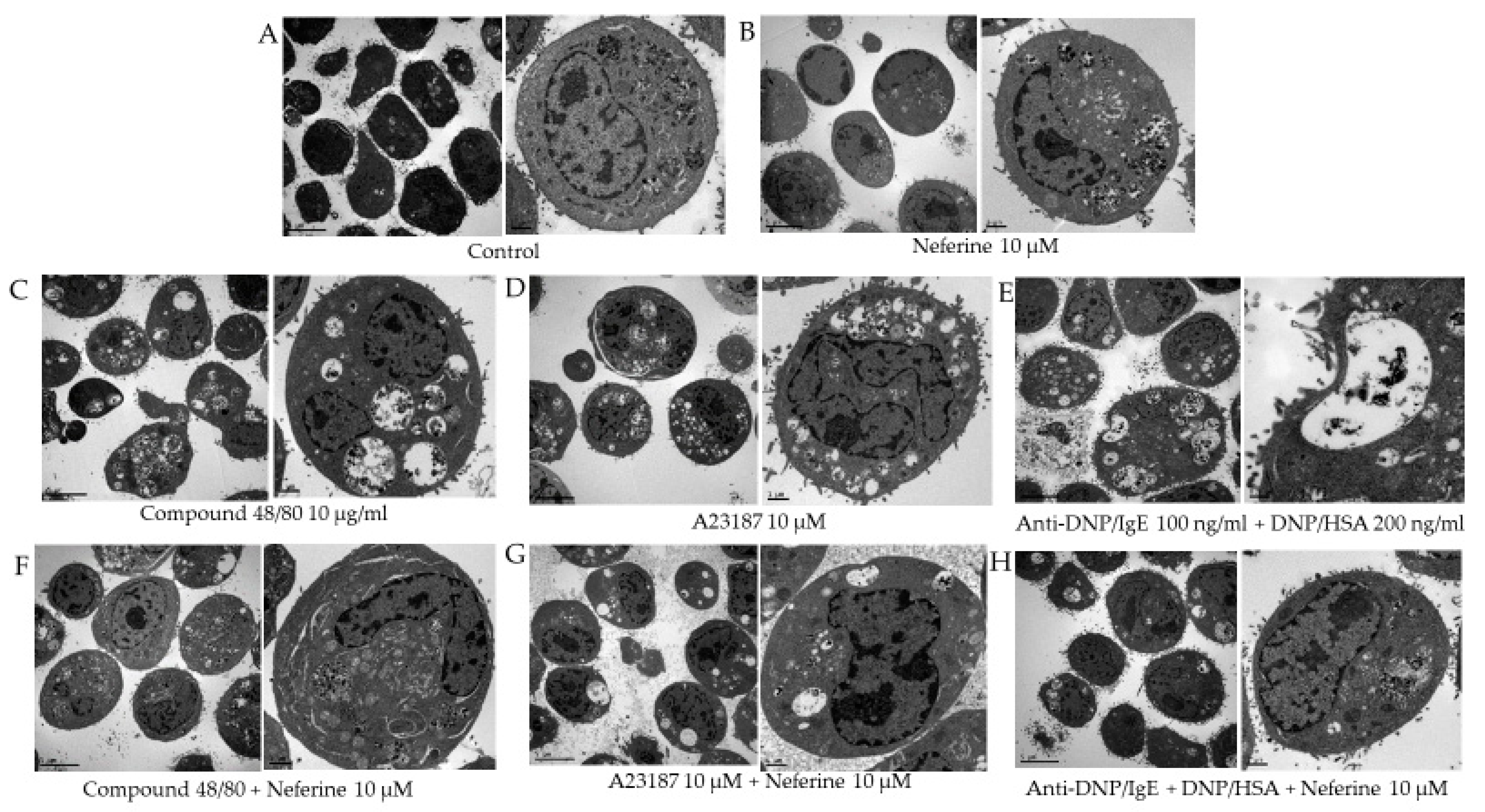
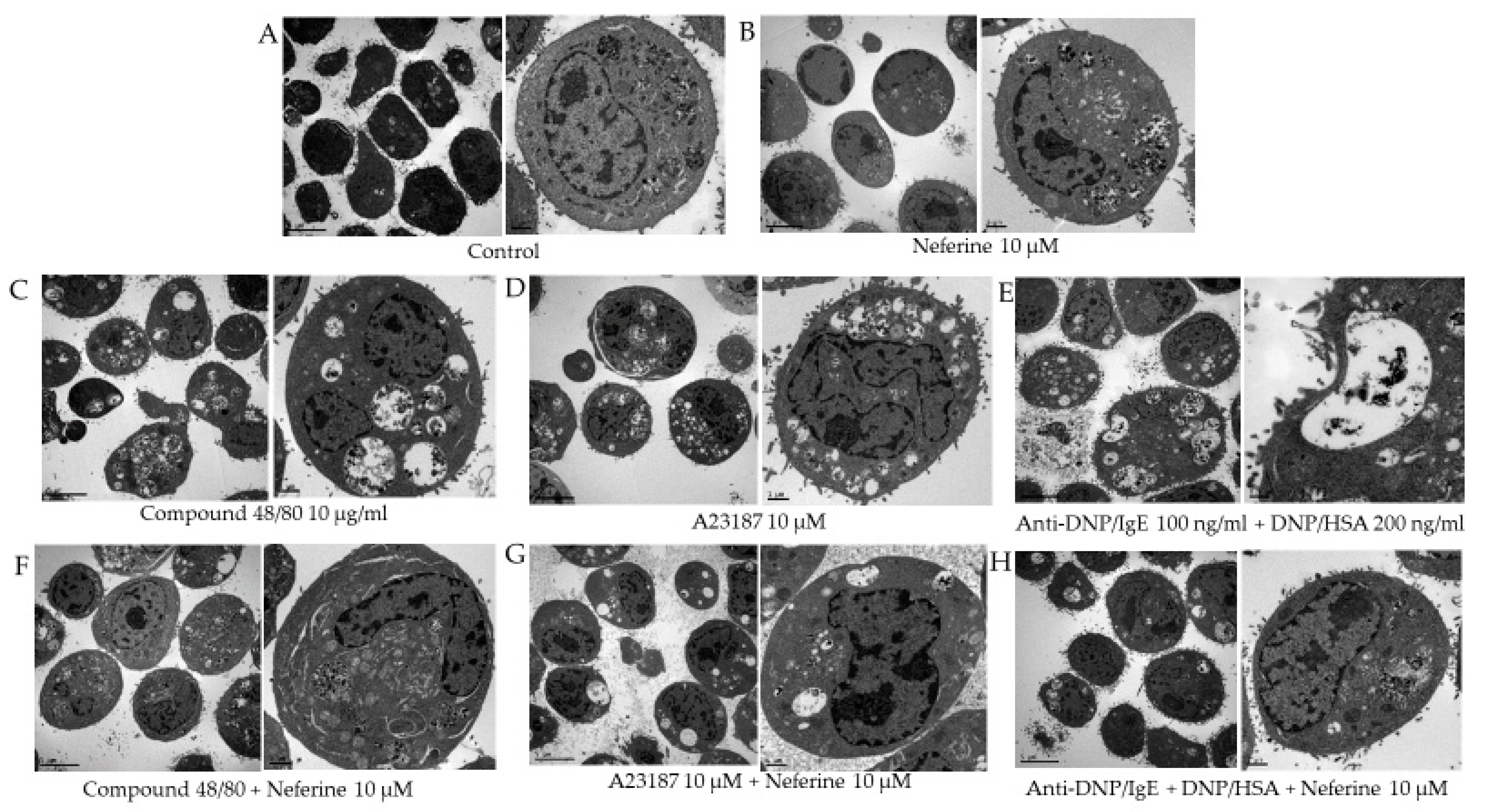
2.2. Neferine Reduces the Degranulation Effect of RBL-2H3 Stimulated by Different Activators
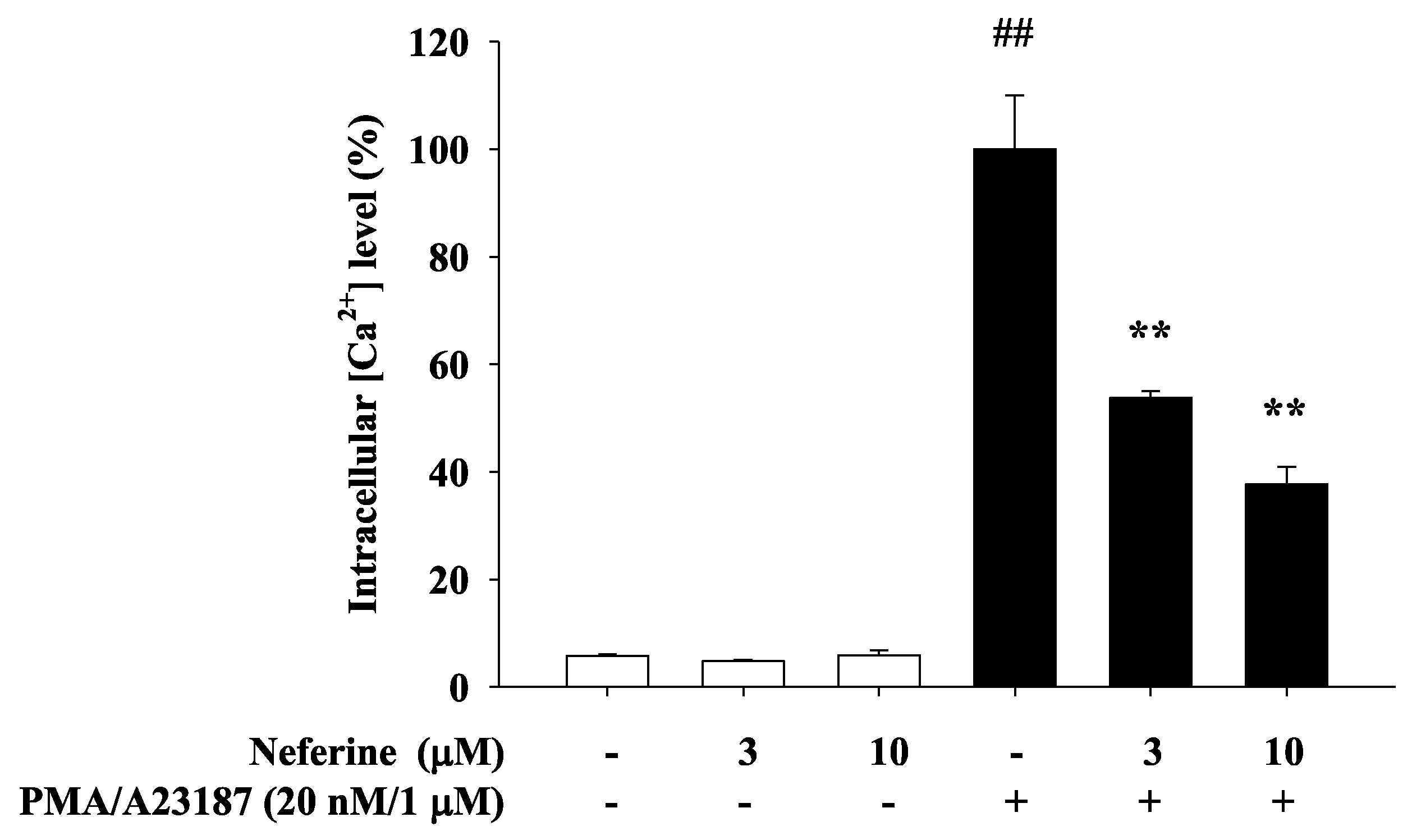
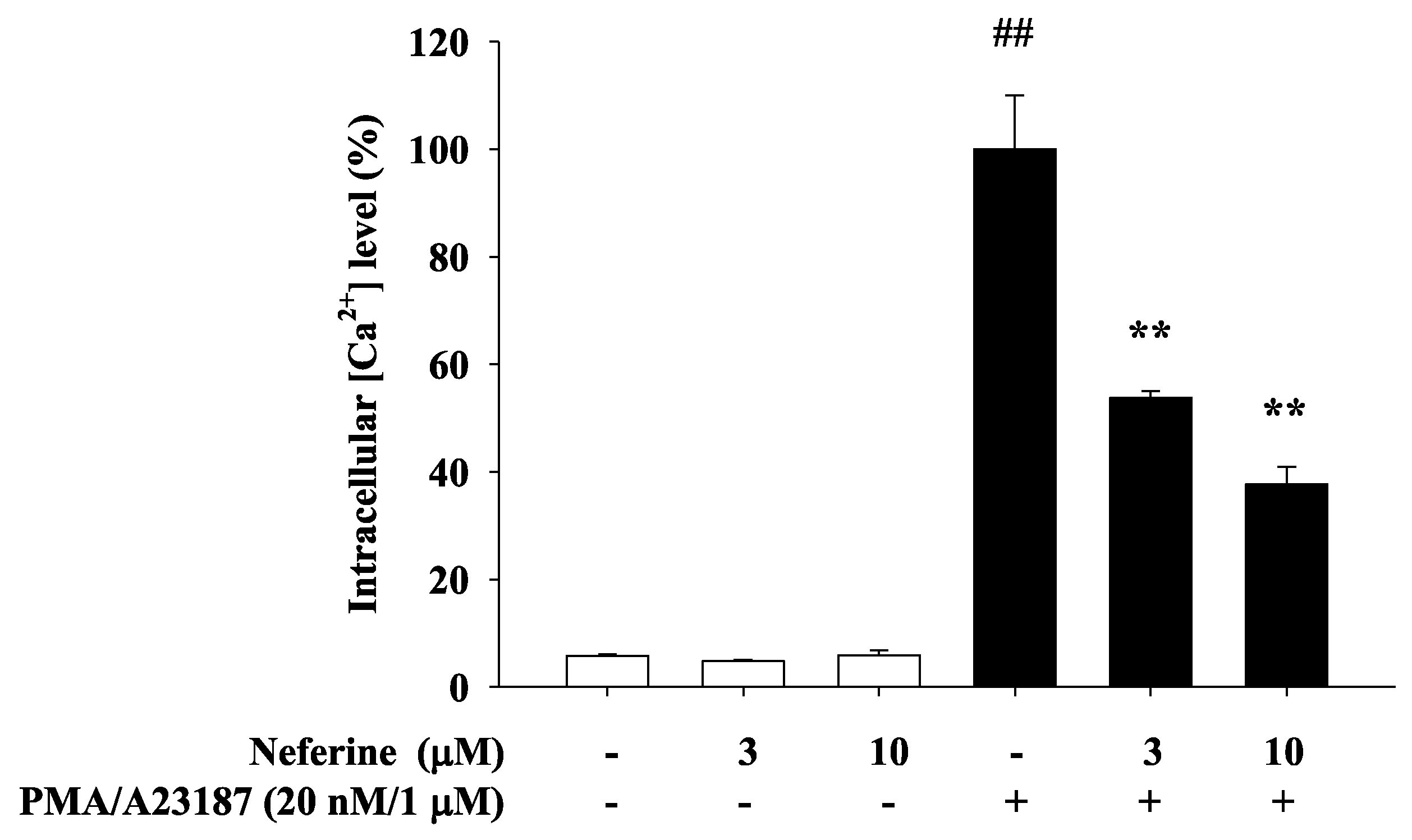
2.3. Neferine Inhibits PMA/A23187-Induced Intracellular Calcium Elevation in RBL-2H3
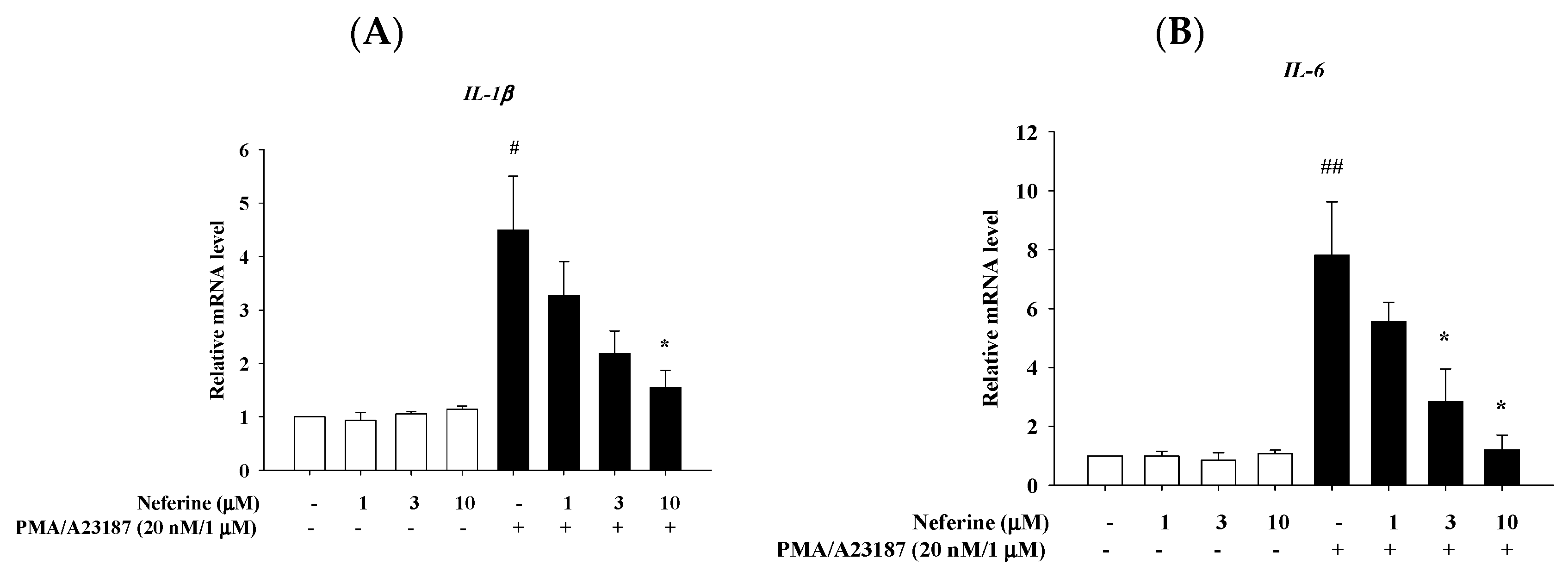
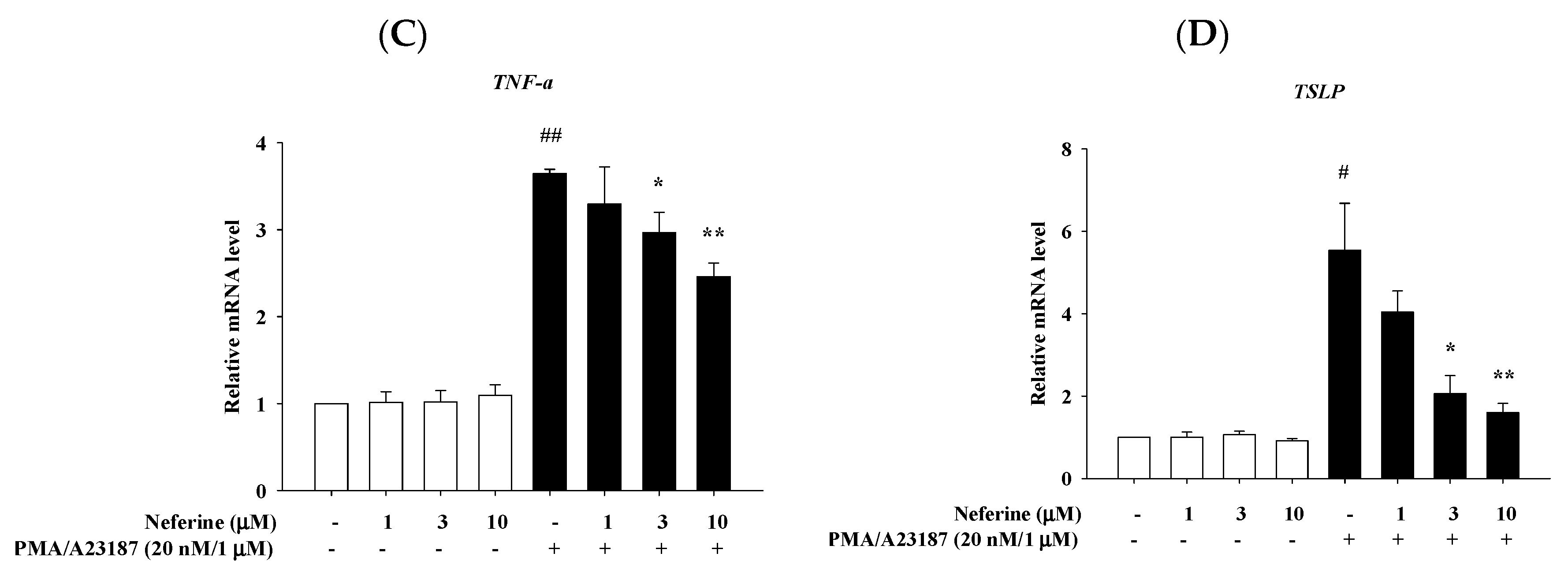
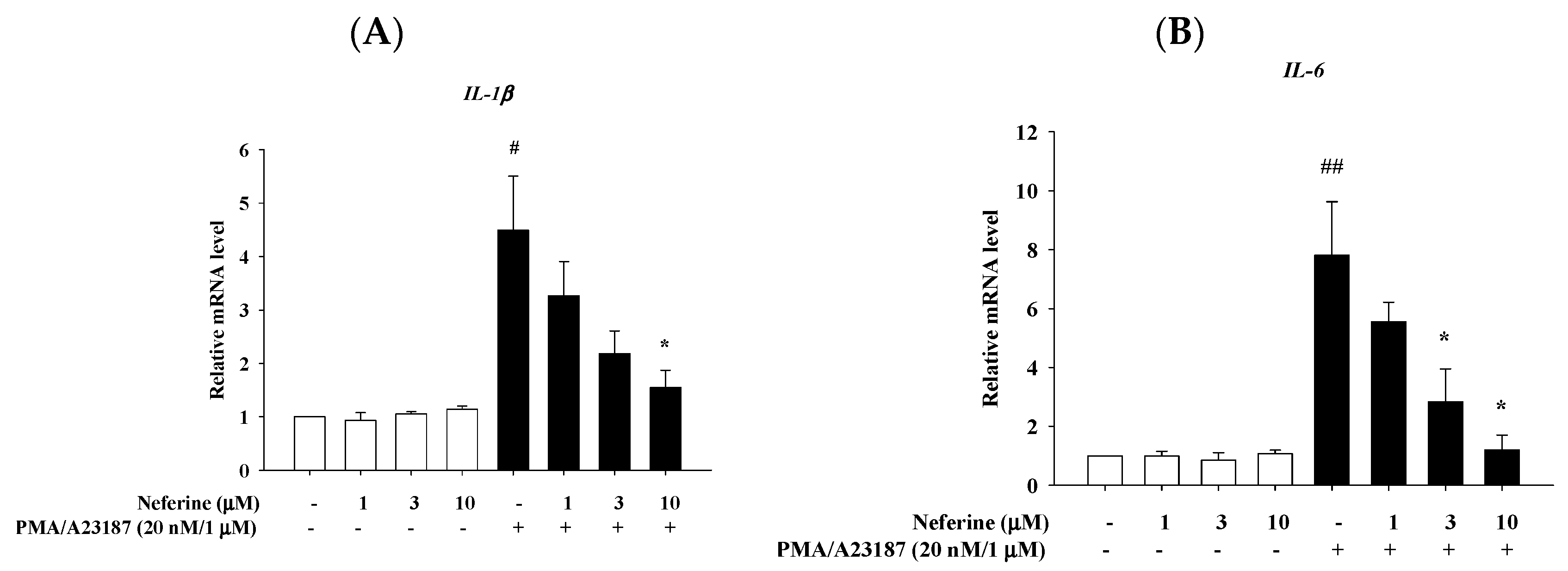
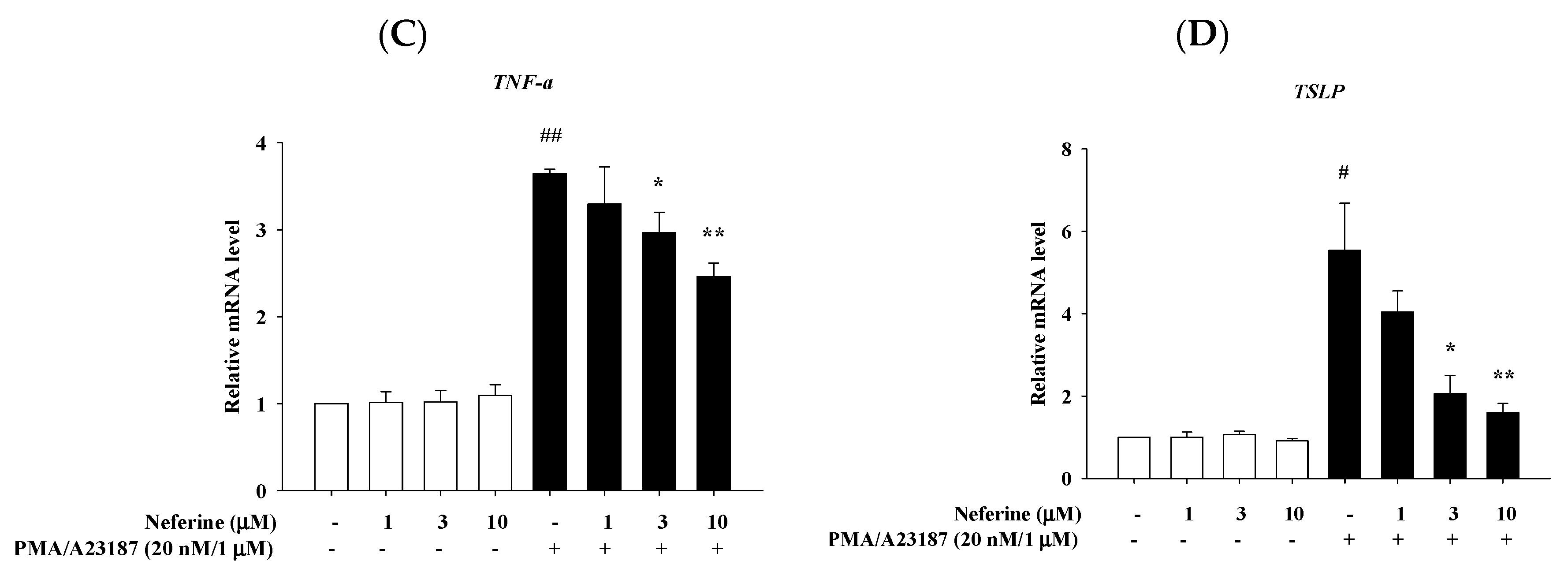
2.4. Neferine Reduced the mRNA Expression of Pro-Inflammatory Cytokines in PMA/A23187 Stimulated by RBL-2H3
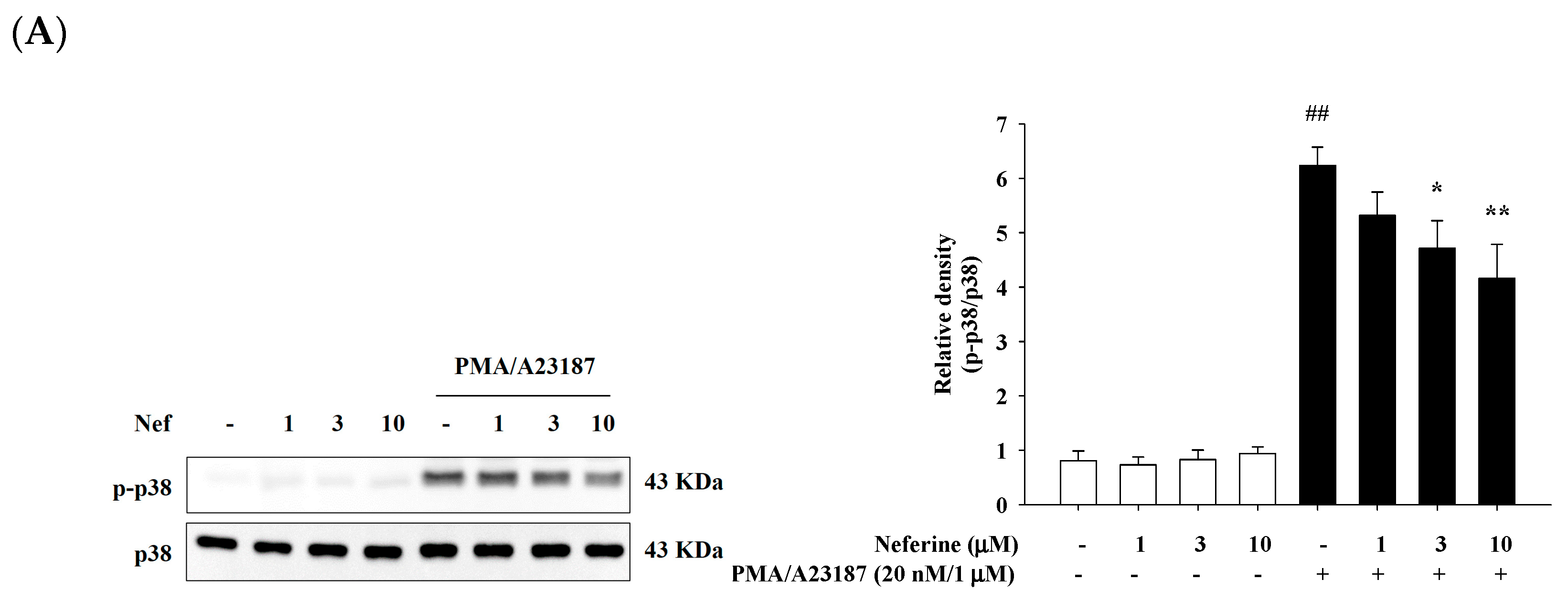
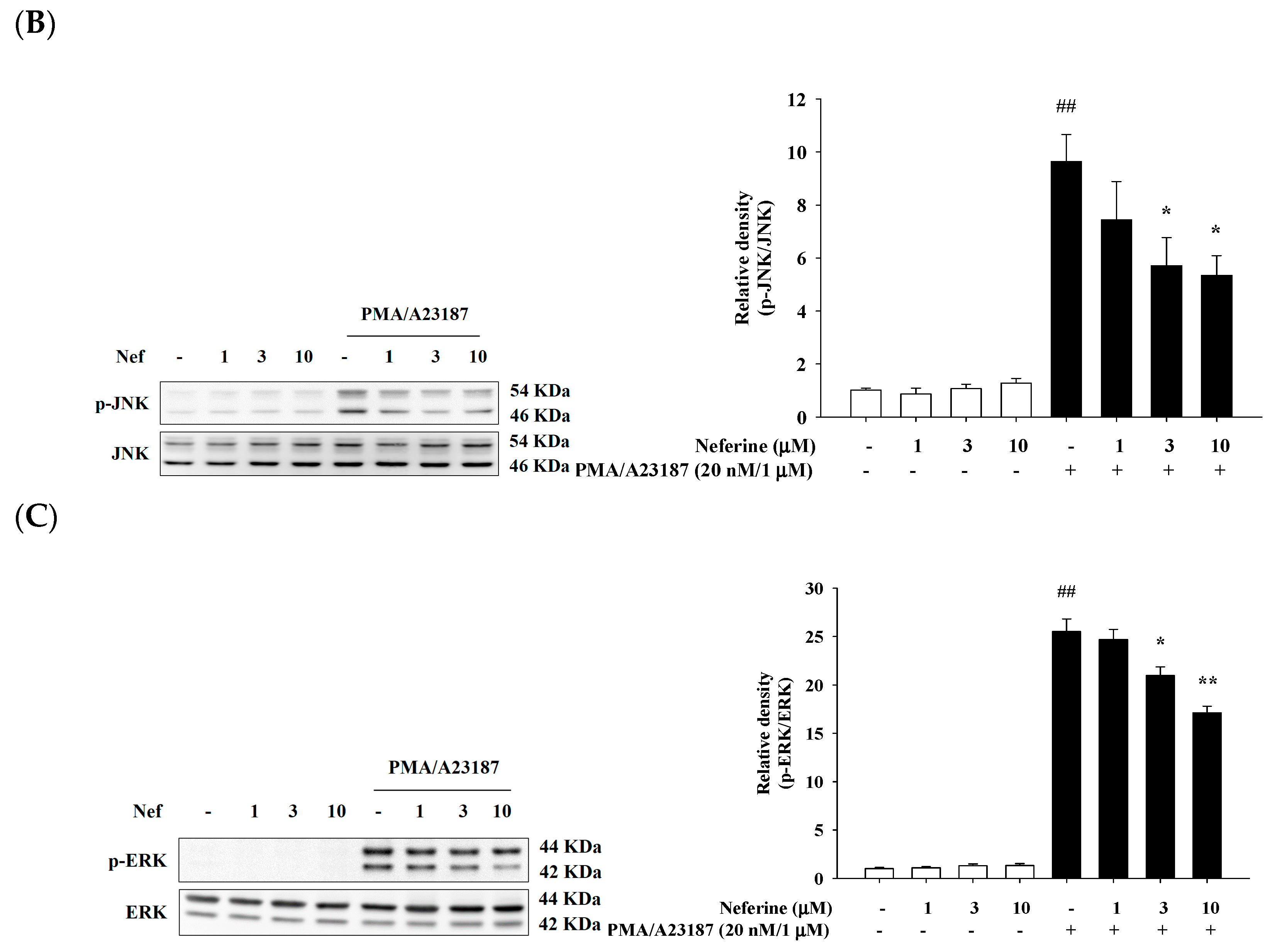
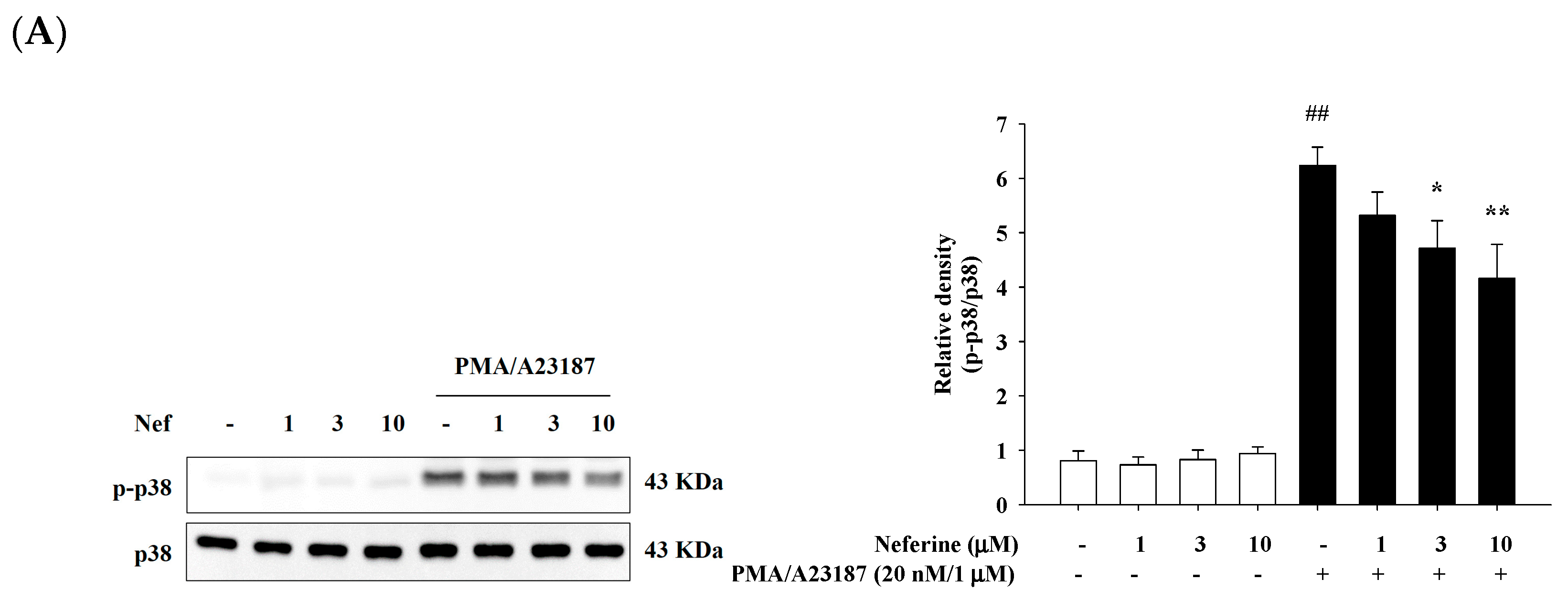
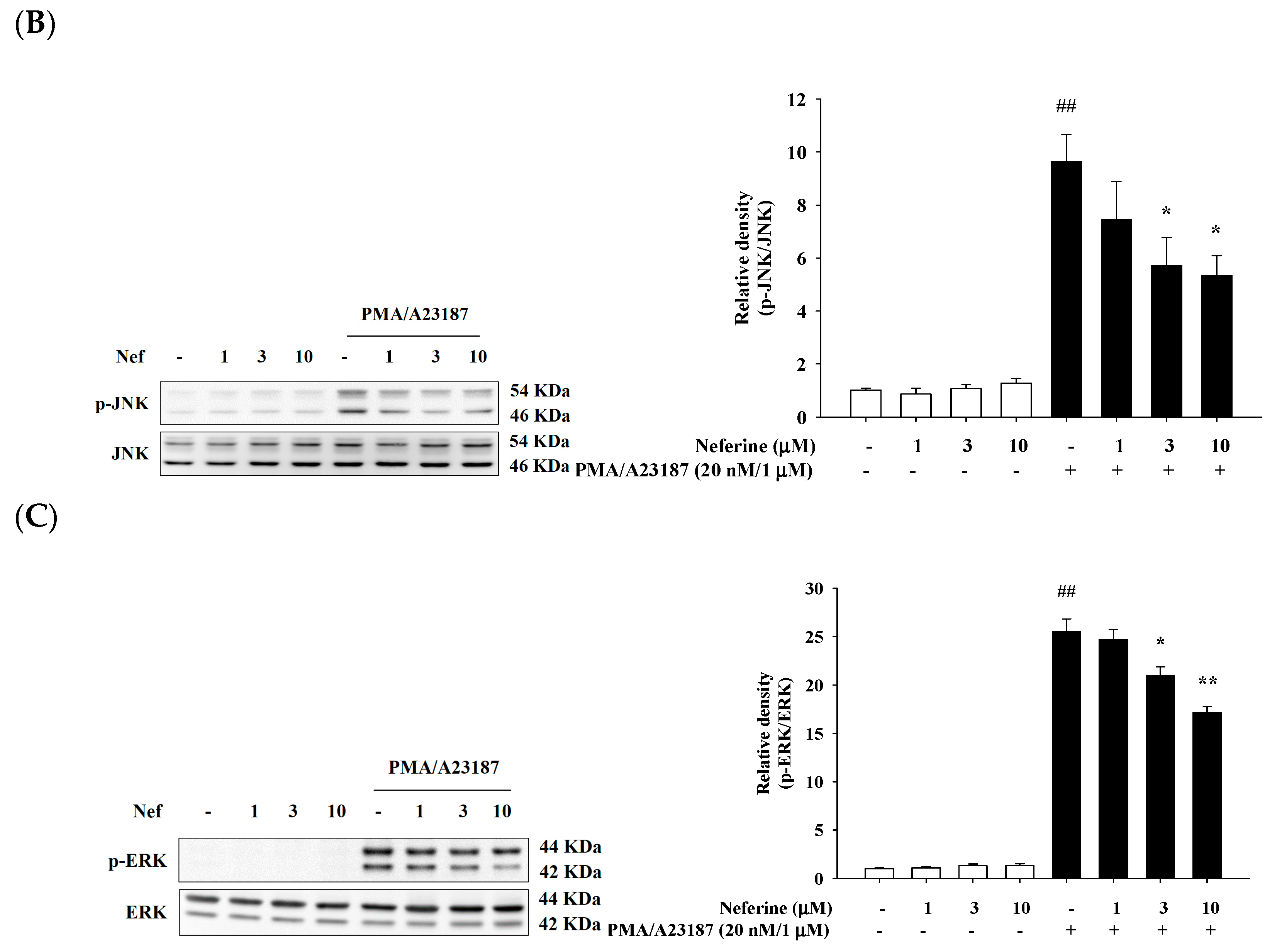
2.5. Neferine Inhibits PMA/A23187-Induced Phosphorylation of MAPK Pathway in RBL-2H3
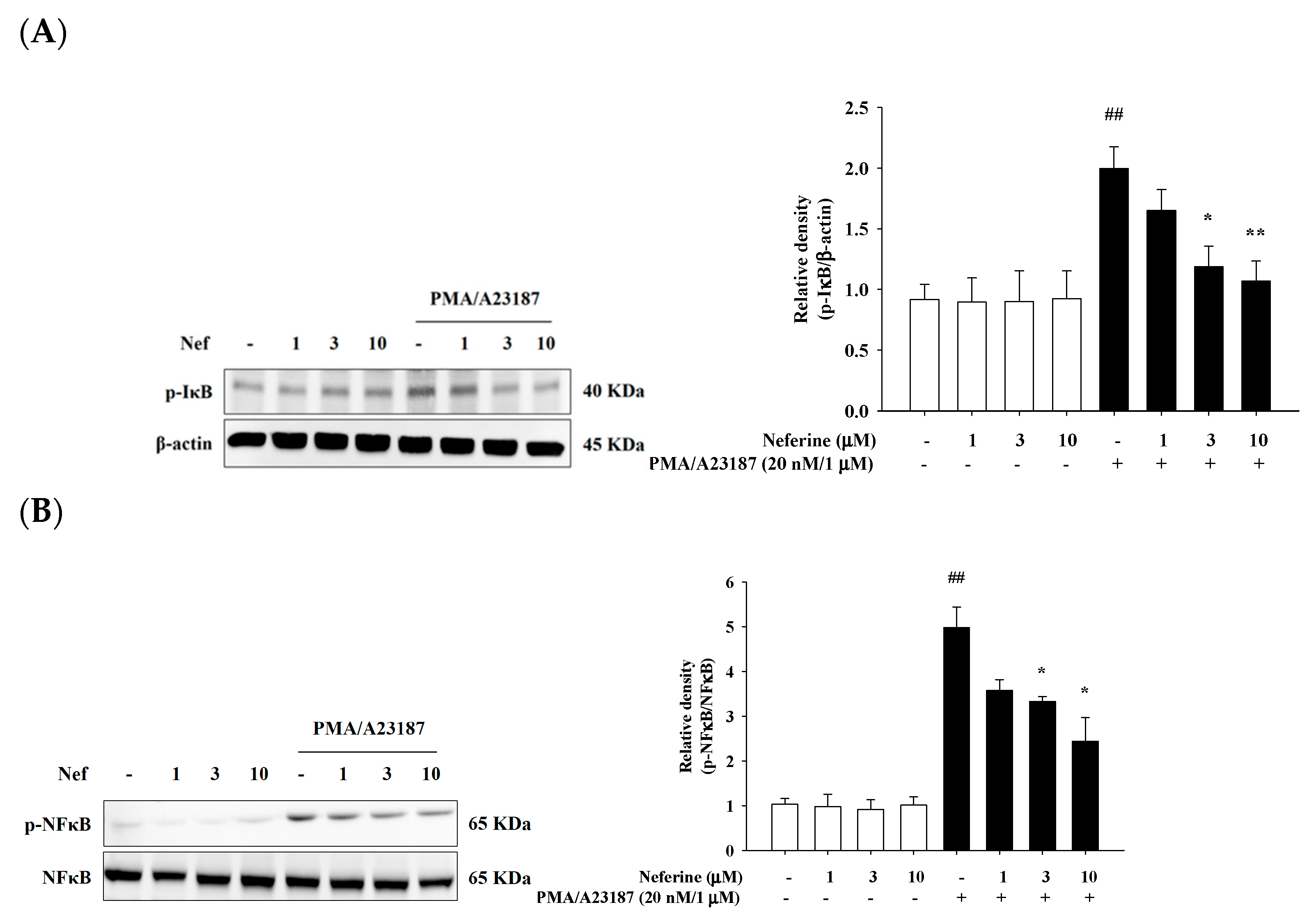
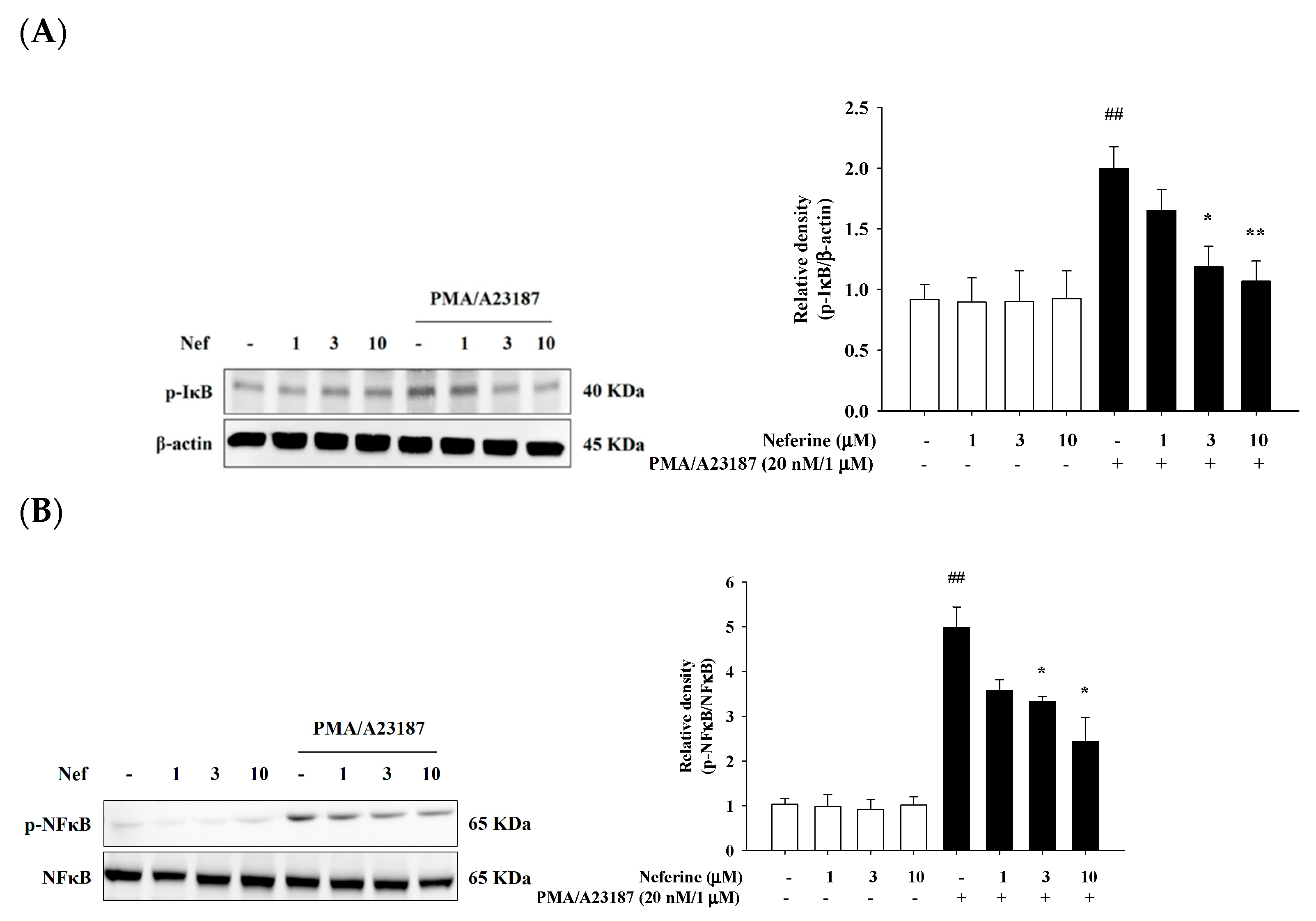
2.6. Neferine Inhibits PMA/A23187-Induced Phosphorylation of NF-κB Pathway in RBL-2H3
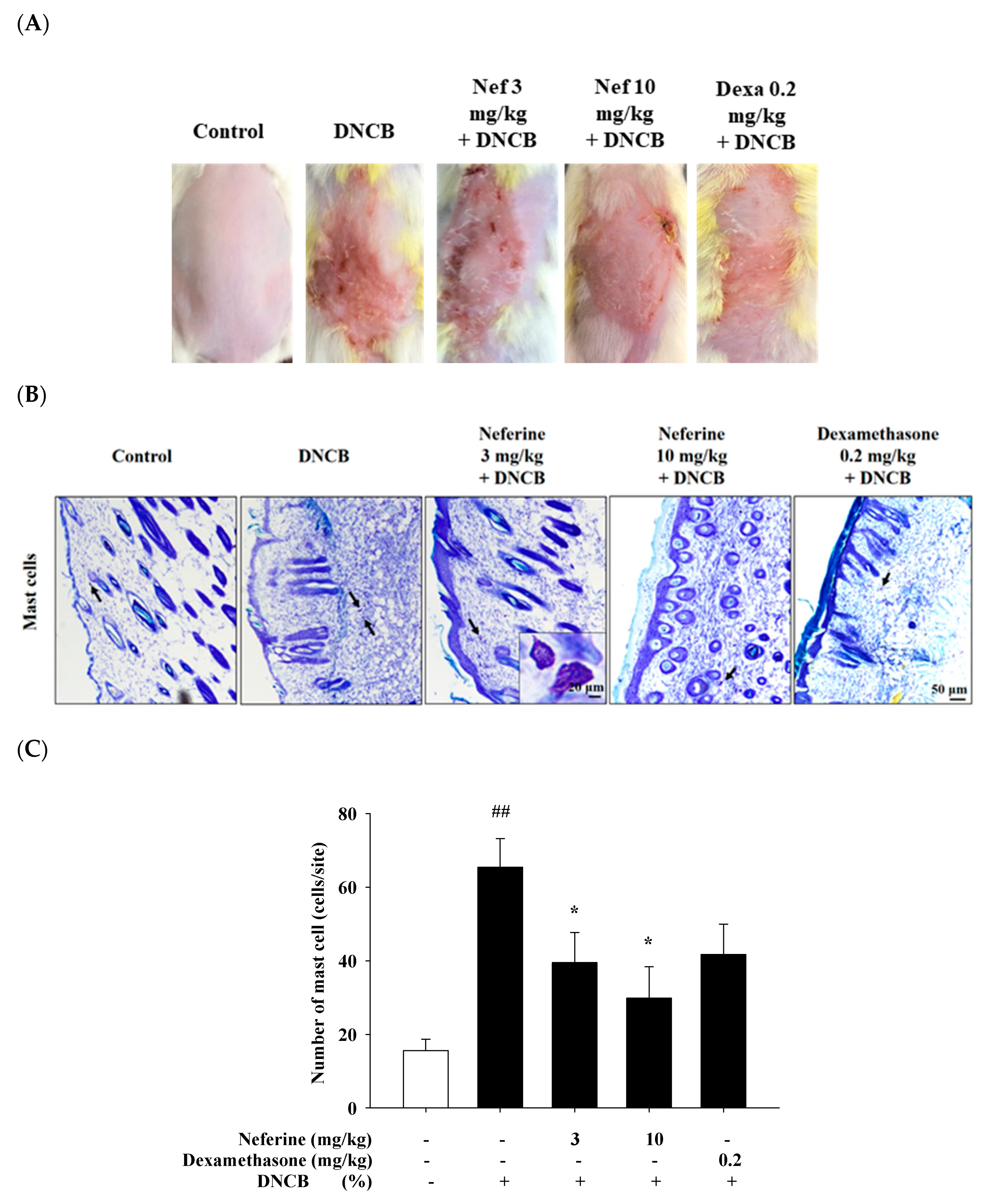
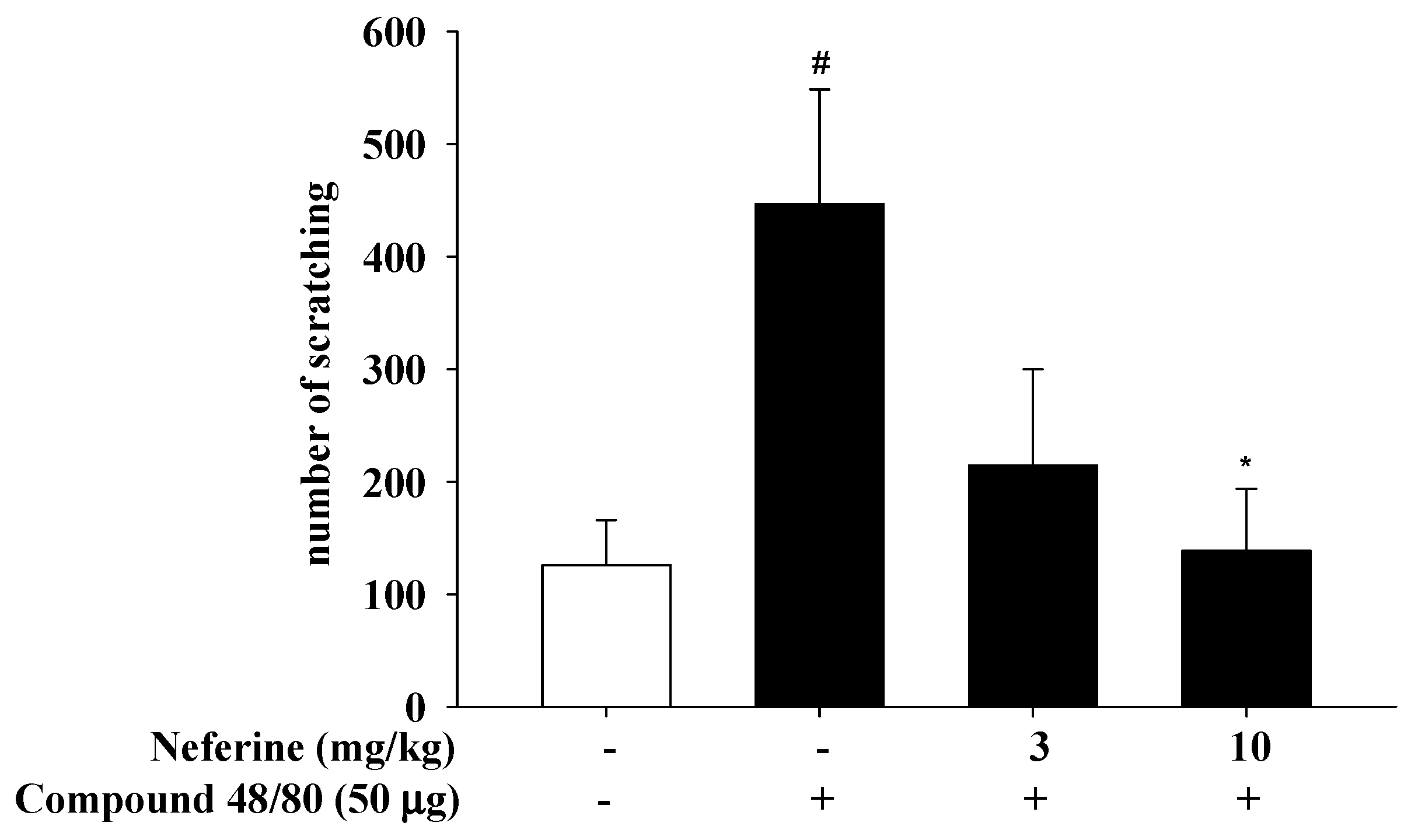
2.7. Neferine’s Effect on 2,4-Dinitrochlorobenzene (DNCB) Induced Atopic Dermatitis
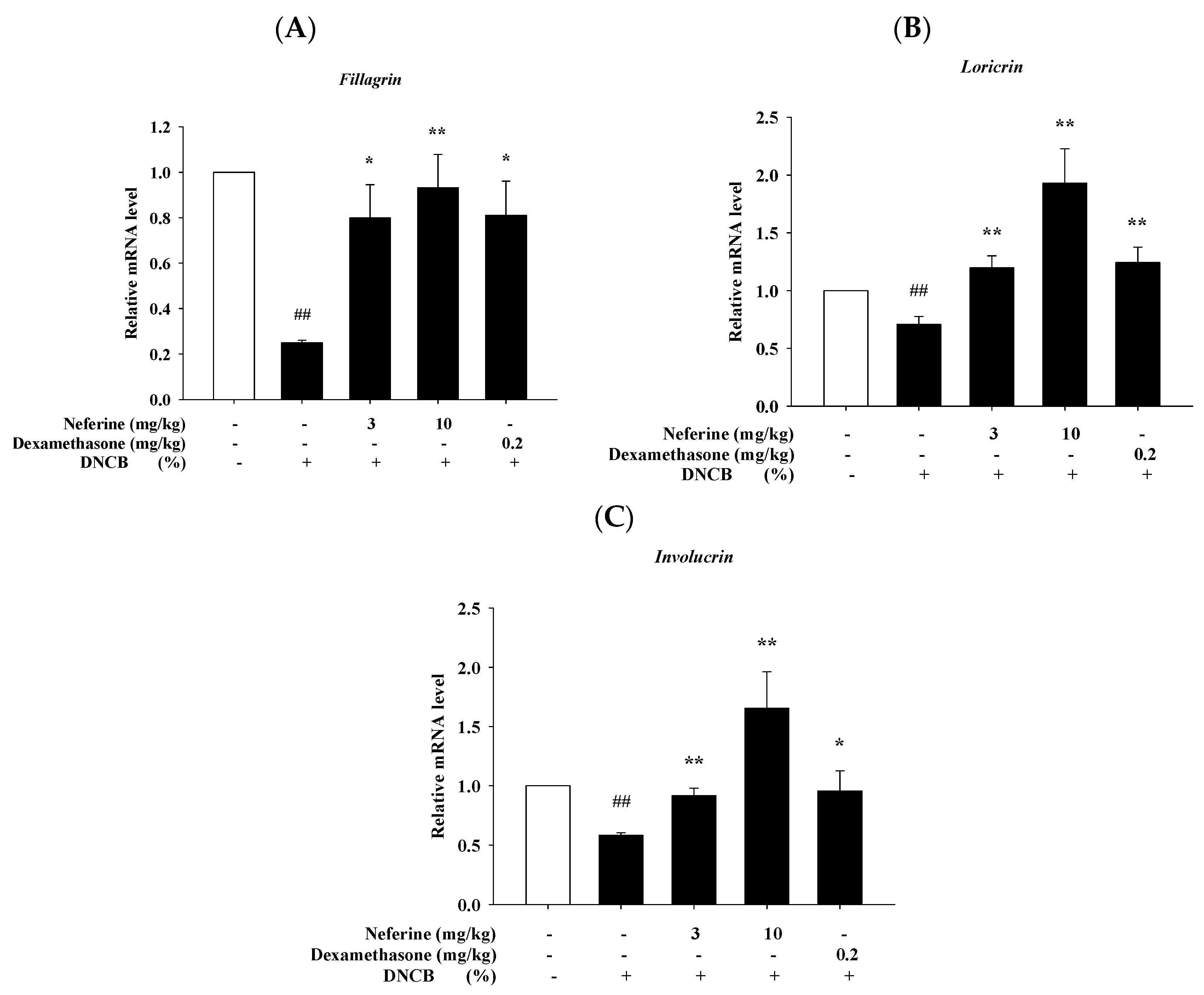
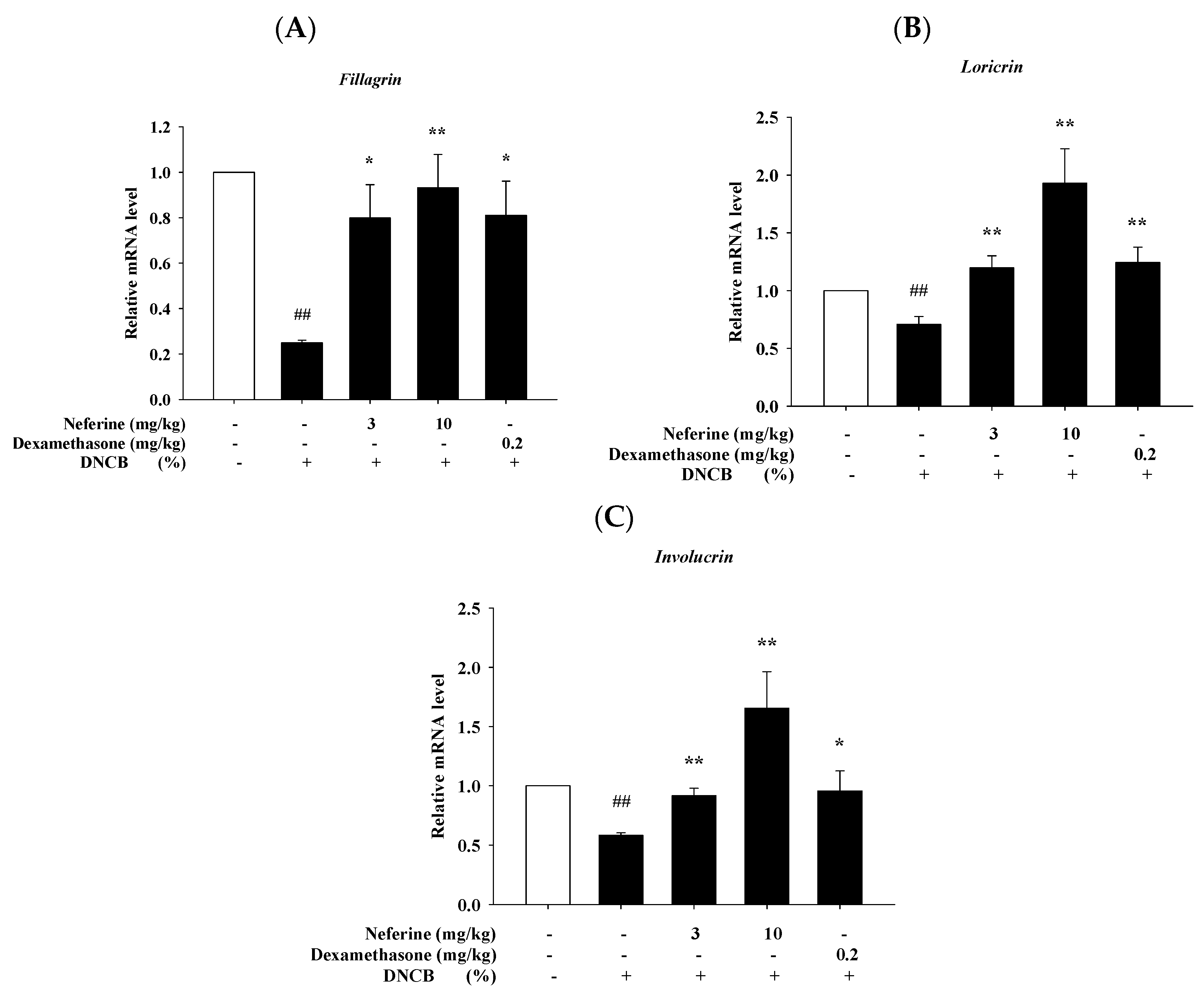
2.8. Neferine Improves the mRNA Expression of Barrier-Related Molecules after Treatment of DNCB
3. Discussion
4. Materials and Methods
4.1. Rat Basophilic Leukemia Cells
4.2. MTT Assay
4.3. Measurement of Intracellular Ca2+ Level
4.4. Quantitative Polymer Chain Reaction (qPCR)
4.5. Western Blot Assay
4.6. Dinitrochlorobenzene (2, 4-Dinitrochlorobenzene, DNCB) Induced Atopic Dermatitis-like Skin Inflammation in Mice
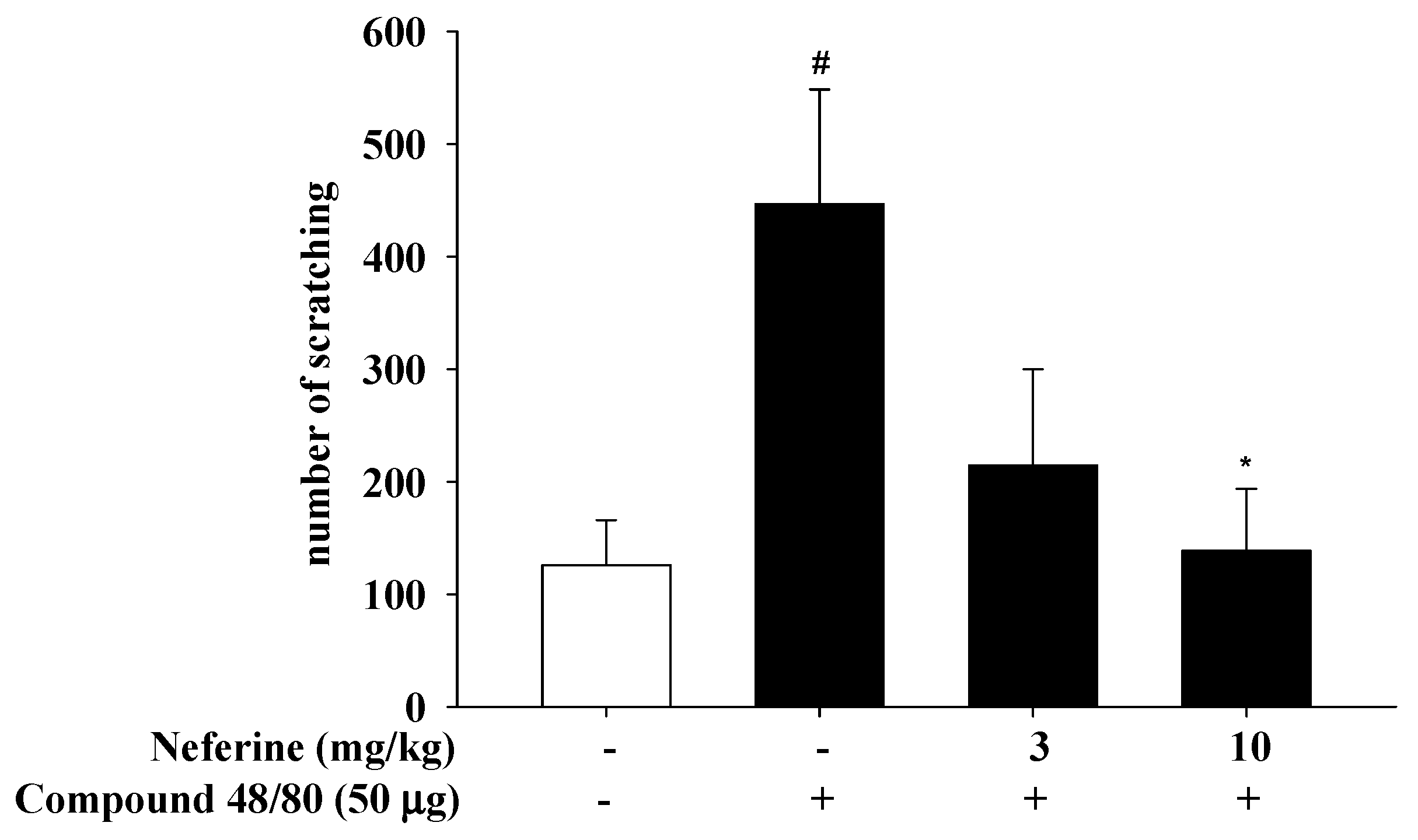
4.7. Compound 48/80-Induced Scratching Behavior in BALB/c Mice
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Gilfillan, A.M.; Beaven, M.A. Regulation of Mast Cell Responses in Health and Disease. Crit. Rev. Immunol. 2011, 31, 475–530. [Google Scholar] [CrossRef]
- Stone, K.D.; Prussin, C.; Metcalfe, D.D. IgE, mast cells, basophils, and eosinophils. J. Allergy Clin. Immunol. 2010, 125, S73–S80. [Google Scholar] [CrossRef] [PubMed]
- Beaven, M.A.; Rogers, J.; Moore, J.P.; Hesketh, T.R.; Smith, G.A.; Metcalfe, J.C. The mechanism of the calcium signal and correlation with histamine release in 2H3 cells. J. Biol. Chem. 1984, 259, 7129–7136. [Google Scholar] [CrossRef]
- Moon, T.C.; Befus, A.D.; Kulka, M. Mast cell mediators: Their differential release and the secretory pathways involved. Front. Immunol. 2014, 5, 569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, J.M.; Sok, D.E.; Kim, M.R. Anti-allergic action of aged black garlic extract in RBL-2H3 cells and passive cutaneous anaphylaxis reaction in mice. J. Med. Food 2014, 17, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Kono, R.; Nomura, S.; Okuno, Y.; Kagiya, T.; Nakamura, M.; Utsunomiya, H.; Ueno, M. Two Japanese pepper (Zanthoxylum piperitum) fruit-derived compounds attenuate IgE-mediated allergic response in vitro and in vivo via inhibition of mast cell degranulation. Eur. J. Pharm. 2020, 885, 173435. [Google Scholar] [CrossRef]
- McNeil, B.D.; Pundir, P.; Meeker, S.; Han, L.; Undem, B.J.; Kulka, M.; Dong, X. Identification of a mast-cell-specific receptor crucial for pseudo-allergic drug reactions. Nature 2015, 519, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Ogasawara, T.; Murakami, M.; Suzuki-Nishimura, T.; Uchida, M.K.; Kudo, I. Mouse bone marrow-derived mast cells undergo exocytosis, prostanoid generation, and cytokine expression in response to G protein-activating polybasic compounds after coculture with fibroblasts in the presence of c-kit ligand. J. Immunol. 1997, 158, 393–404. [Google Scholar]
- Avena-Woods, C. Overview of atopic dermatitis. Am. J. Manag. Care 2017, 23, S115–S123. [Google Scholar]
- Xie, J.; Chen, M.H.; Ying, C.P.; Chen, M.Y. Neferine induces p38 MAPK/JNK1/2 activation to modulate melanoma proliferation, apoptosis, and oxidative stress. Ann. Transl. Med. 2020, 8, 1643. [Google Scholar] [CrossRef]
- Zhong, Y.; He, S.; Huang, K.; Liang, M. Neferine suppresses vascular endothelial inflammation by inhibiting the NF-κB signaling pathway. Arch. Biochem. Biophys. 2020, 696, 108595. [Google Scholar] [CrossRef]
- Tang, Y.-S.; Zhao, Y.-H.; Zhong, Y.; Li, X.-Z.; Pu, J.-X.; Luo, Y.-C.; Zhou, Q.-L. Neferine inhibits LPS-ATP-induced endothelial cell pyroptosis via regulation of ROS/NLRP3/Caspase-1 signaling pathway. Inflamm. Res. 2019, 68, 727–738. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, X.; Chang, Q.; Xu, J.; Huang, Y.; Guo, Q.; Zhang, S.; Wang, W.; Chen, X.; Wang, J. Neferine, a bisbenzylisoquinline alkaloid attenuates bleomycin-induced pulmonary fibrosis. Eur. J. Pharmacol. 2010, 627, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Marthandam Asokan, S.; Mariappan, R.; Muthusamy, S.; Velmurugan, B.K. Pharmacological benefits of neferine - A comprehensive review. Life Sci. 2018, 199, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Yeh, K.C.; Hung, C.F.; Lin, Y.F.; Chang, D.C.; Pai, M.S.; Wang, S.J. Neferine, a bisbenzylisoquinoline alkaloid of Nelumbo nucifera, inhibits glutamate release in rat cerebrocortical nerve terminals through 5-HT1A receptors. Eur. J. Pharmacol. 2020, 889, 173589. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Bai, H.; Shu, M.; Chen, M.; Khan, A.; Bai, Z. Antioxidative and antiphotoaging activities of neferine upon UV-A irradiation in human dermal fibroblasts. Biosci. Rep. 2018, 38, BSR20181414. [Google Scholar] [CrossRef]
- Khan, A.; Bai, H.; Liu, E.; Chen, M.; Yu, C.; Wang, R.; Khan, A.; Bai, Z. Protective effect of neferine against UV-B-mediated oxidative damage in human epidermal keratinocytes. J. Dermatol. Treat. 2018, 29, 733–741. [Google Scholar] [CrossRef]
- Yang, C.-C.; Hung, Y.-L.; Ko, W.-C.; Tsai, Y.-J.; Chang, J.-F.; Liang, C.-W.; Chang, D.-C.; Hung, C.-F. Effect of Neferine on DNCB-Induced Atopic Dermatitis in HaCaT Cells and BALB/c Mice. Int. J. Mol. Sci. 2021, 22, 8237. [Google Scholar] [CrossRef]
- Bae, Y.; Lee, S.; Kim, S.H. Chrysin suppresses mast cell-mediated allergic inflammation: Involvement of calcium, caspase-1 and nuclear factor-κB. Toxicol. Appl. Pharm. 2011, 254, 56–64. [Google Scholar] [CrossRef]
- Passante, E.; Frankish, N. The RBL-2H3 cell line: Its provenance and suitability as a model for the mast cell. Inflamm. Res. 2009, 58, 737–745. [Google Scholar] [CrossRef]
- Falcone, F.H.; Wan, D.; Barwary, N.; Sagi-Eisenberg, R. RBL cells as models for in vitro studies of mast cells and basophils. Immunol. Rev. 2018, 282, 47–57. [Google Scholar] [CrossRef]
- Weatherly, L.M.; Nelson, A.J.; Shim, J.; Riitano, A.M.; Gerson, E.D.; Hart, A.J.; De Juan-Sanz, J.; Ryan, T.A.; Sher, R.; Hess, S.T.; et al. Antimicrobial agent triclosan disrupts mitochondrial structure, revealed by super-resolution microscopy, and inhibits mast cell signaling via calcium modulation. Toxicol. Appl. Pharmacol. 2018, 349, 39–54. [Google Scholar] [CrossRef]
- Verbsky, J.W.; McAllister, P.K.; Malone, D.G. Mast cell activation in human synovium explants by calcium ionophore A23187, compound 48/80, and rabbit IgG anti-human IgE, but not morphine sulfate. Inflamm. Res. 1996, 45, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Koch, G.; Habermann, B.; Mohr, C.; Just, I.; Aktories, K. ADP-ribosylation of rho proteins is inhibited by melittin, mast cell degranulating peptide and compound 48/80. Eur. J. Pharm. 1992, 226, 87–91. [Google Scholar] [CrossRef]
- Kim, D.; Kobayashi, T.; Nagao, K. Research Techniques Made Simple: Mouse Models of Atopic Dermatitis. J. Investig. Dermatol. 2019, 139, 984–990.e981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Shin, J.U.; Lee, K.H. Atopic dermatitis and skin barrier dysfunction. Allergy Asthma Respir. Dis. 2013, 1, 20. [Google Scholar] [CrossRef]
- Nakamura, Y.; Oscherwitz, J.; Cease, K.B.; Chan, S.M.; Muñoz-Planillo, R.; Hasegawa, M.; Villaruz, A.E.; Cheung, G.Y.C.; McGavin, M.J.; Travers, J.B.; et al. Staphylococcus δ-toxin induces allergic skin disease by activating mast cells. Nature 2013, 503, 397–401. [Google Scholar] [CrossRef] [Green Version]
- Gilfillan, A.M.; Tkaczyk, C. Integrated signalling pathways for mast-cell activation. Nat. Rev. Immunol. 2006, 6, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Barsumian, E.L.; Isersky, C.; Petrino, M.G.; Siraganian, R.P. IgE-induced histamine release from rat basophilic leukemia cell lines: Isolation of releasing and nonreleasing clones. Eur. J. Immunol 1981, 11, 317–323. [Google Scholar] [CrossRef]
- Naal, R.M.Z.G.; Tabb, J.; Holowka, D.; Baird, B. In situ measurement of degranulation as a biosensor based on RBL-2H3 mast cells. Biosens. Bioelectron. 2004, 20, 791–796. [Google Scholar] [CrossRef]
- Sahid, M.N.A.; Kiyoi, T. Mast cell activation markers for in vitro study. J. Immunoass. Immunochem. 2020, 41, 778–816. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.T.; Beaven, M.A. Regulation of Ca2+ signaling with particular focus on mast cells. Crit. Rev. Immunol. 2009, 29, 155–186. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Maeyama, K.; Sagi-Eisenberg, R.; Beaven, M.A. Antigen and thapsigargin promote influx of Ca2+ in rat basophilic RBL-2H3 cells by ostensibly similar mechanisms that allow filling of inositol 1,4,5-trisphosphate-sensitive and mitochondrial Ca2+ stores. Biochem. J. 1994, 304 Pt 2, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Maeyama, K.; Sasaki, M.; Watanabe, T. Simultaneous determination of intracellular calcium concentration and histamine secretion in rat basophilic leukemia cells (RBL-2H3). Anal. Biochem. 1991, 194, 316–325. [Google Scholar] [CrossRef]
- Schwartz, C.; Moran, T.; Saunders, S.P.; Kaszlikowska, A.; Floudas, A.; Bom, J.; Nunez, G.; Iwakura, Y.; O’Neill, L.; Irvine, A.D.; et al. Spontaneous atopic dermatitis in mice with a defective skin barrier is independent of ILC2 and mediated by IL-1β. Allergy 2019, 74, 1920–1933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kritas, S.K.; Saggini, A.; Varvara, G.; Murmura, G.; Caraffa, A.; Antinolfi, P.; Toniato, E.; Pantalone, A.; Neri, G.; Frydas, S.; et al. Impact of Mast Cells on the Skin. Int. J. Immunopathol. Pharmacol. 2013, 26, 855–859. [Google Scholar] [CrossRef] [PubMed]
- Zarubin, T.; Han, J. Activation and signaling of the p38 MAP kinase pathway. Cell Res. 2005, 15, 11–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czarnowicki, T.; Krueger, J.G.; Guttman-Yassky, E. Skin barrier and immune dysregulation in atopic dermatitis: An evolving story with important clinical implications. J. Allergy Clin. Immunol. Pract. 2014, 2, 371–379, quiz 380–371. [Google Scholar] [CrossRef]
- Furue, M. T helper type 2 signatures in atopic dermatitis. J. Cutan. Immunol. Allergy 2018, 1, 93–99. [Google Scholar] [CrossRef]
- Brunner, P.M.; Guttman-Yassky, E.; Leung, D.Y.M. The immunology of atopic dermatitis and its reversibility with broad-spectrum and targeted therapies. J. Allergy Clin. Immunol. 2017, 139, S65–S76. [Google Scholar] [CrossRef] [Green Version]
- Ilves, T.; Tiitu, V.; Suttle, M.M.; Saarinen, J.V.; Harvima, I.T. Epidermal Expression of Filaggrin/Profilaggrin Is Decreased in Atopic Dermatitis: Reverse Association With Mast Cell Tryptase and IL-6 but Not With Clinical Severity. Dermatitis 2015, 26, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Wu, Q.; Jang, Y.P.; Choung, S.Y. Pinus densiflora bark extract ameliorates 2,4-dinitrochlorobenzene-induced atopic dermatitis in NC/Nga mice by regulating Th1/Th2 balance and skin barrier function. Phytother. Res. 2018, 32, 1135–1143. [Google Scholar] [CrossRef]
- Kim; Seong; Choung. Fermented Morinda citrifolia (Noni) Alleviates DNCB-Induced Atopic Dermatitis in NC/Nga Mice through Modulating Immune Balance and Skin Barrier Function. Nutrients 2020, 12, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki, N.; Igeta, K.; Kim, J.F.; Nagao, M.; Shiraishi, N.; Nakamura, N.; Nagai, H. Involvement of unique mechanisms in the induction of scratching behavior in BALB/c mice by compound 48/80. Eur. J. Pharm. 2002, 448, 175–183. [Google Scholar] [CrossRef]
- Čelakovská, J. Atopic march, food allergy and food hypersensitivity in children and adolescents suffering from atopic dermatitis. Food Agric. Immunol. 2015, 26, 590–600. [Google Scholar] [CrossRef]
- Čelakovská, J.; Bukač, J.; Cermákova, E.; Vaňková, R.; Skalská, H.; Krejsek, J.; Andrýs, C. Analysis of Results of Specific IgE in 100 Atopic Dermatitis Patients with the Use of Multiplex Examination ALEX2—Allergy Explorer. Int. J. Mol. Sci. 2021, 22, 5286. [Google Scholar] [CrossRef] [PubMed]
- Su, I.C.; Hung, C.F.; Lin, C.N.; Huang, S.K.; Wang, S.J. Cycloheterophyllin Inhibits the Release of Glutamate from Nerve Terminals of the Rat Hippocampus. Chem. Res. Toxicol. 2019, 32, 1591–1598. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primers | Sequence (5′-3′) |
|---|---|---|
| IL-1β | Forward | CAG CTT TCG ACA GTG AGG AGA |
| Reverse | TTG TCG AGA TGC TGC TGT GA | |
| IL-6 | Forward | ACA AGT CCG GAG AGG AGA CT |
| Reverse | TTG CCA TTG CAC AAC TCT TTT C | |
| TNF-α | Forward | ATGGGCTCCCTCTCATCAGT |
| Reverse | GAAATGGCAAATCGGCTGAC | |
| TSLP | Forward | TCA GGC AAC AGC ATG GTT CT |
| Reverse | AAG TTA GTG CCA GCC GTA CC | |
| GAPDH | Forward | TTC ACC ACC ATG GAG AAG GC |
| Reverse | GGC ATG GAC TGT GGT CAT GA |
| Genes | Primers | Sequence (5′-3′) |
|---|---|---|
| IL-1β | Forward | TGG ACC TTC CAG GAT GAG GAC A |
| Reverse | GTT CAT CTC GGA GCC TGT AGT G | |
| IL-6 | Forward | AGT TGC CTT CTT GGG ACT GA |
| Reverse | TCC ACG ATT TCC CAG AGA AC | |
| TNF-α | Forward | GGT GCC TAT GTC TCA GCC TCT TTT |
| Reverse | GCC ATA GAA CTG ATG AGA GGG AG | |
| TSLP | Forward | AGC TTG TCT CCT GAA AAT CGA G |
| Reverse | AGG TTT GAT TCA GGC AGA TGT T | |
| Fillagrin | Forward | ATG TCC GCT CTC CTG GAA AG |
| Reverse | TGG ATT CTT CAA GAC TGC CTG TA | |
| Involucrin | Forward | ATG TCC CAT CAA CAC ACA CTG |
| Reverse | TGG AGT TGG TTG CTT TGC TTG | |
| Loricrin | Forward | CTC CTG TGG GTT GTG GAA AGA |
| Reverse | TGG AAC CAC CTC CAT AGG AAC | |
| GAPDH | Forward | ACC CAG AAG ACT GTG GAT GG |
| Reverse | CAC ATT GGG GGT AGG AAC AC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, K.-M.; Hung, Y.-L.; Wang, S.-J.; Tsai, Y.-J.; Wu, N.-L.; Liang, C.-W.; Chang, D.-C.; Hung, C.-F. Anti-Allergic and Anti-Inflammatory Effects of Neferine on RBL-2H3 Cells. Int. J. Mol. Sci. 2021, 22, 10994. https://doi.org/10.3390/ijms222010994
Chiu K-M, Hung Y-L, Wang S-J, Tsai Y-J, Wu N-L, Liang C-W, Chang D-C, Hung C-F. Anti-Allergic and Anti-Inflammatory Effects of Neferine on RBL-2H3 Cells. International Journal of Molecular Sciences. 2021; 22(20):10994. https://doi.org/10.3390/ijms222010994
Chicago/Turabian StyleChiu, Kuan-Ming, Yen-Ling Hung, Su-Jane Wang, Yi-Ju Tsai, Nan-Lin Wu, Cher-Wei Liang, Der-Chen Chang, and Chi-Feng Hung. 2021. "Anti-Allergic and Anti-Inflammatory Effects of Neferine on RBL-2H3 Cells" International Journal of Molecular Sciences 22, no. 20: 10994. https://doi.org/10.3390/ijms222010994
APA StyleChiu, K.-M., Hung, Y.-L., Wang, S.-J., Tsai, Y.-J., Wu, N.-L., Liang, C.-W., Chang, D.-C., & Hung, C.-F. (2021). Anti-Allergic and Anti-Inflammatory Effects of Neferine on RBL-2H3 Cells. International Journal of Molecular Sciences, 22(20), 10994. https://doi.org/10.3390/ijms222010994