
Pistil Mating Type and Morphology Are Mediated by the Brassinosteroid Inactivating Activity of the S-Locus Gene BAHD in Heterostylous Turnera Species

 , ,
, , 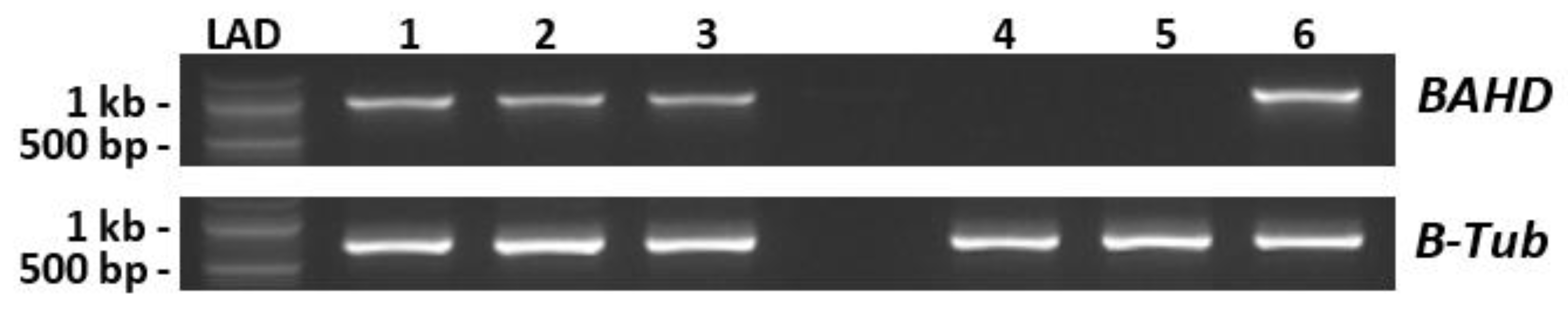{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
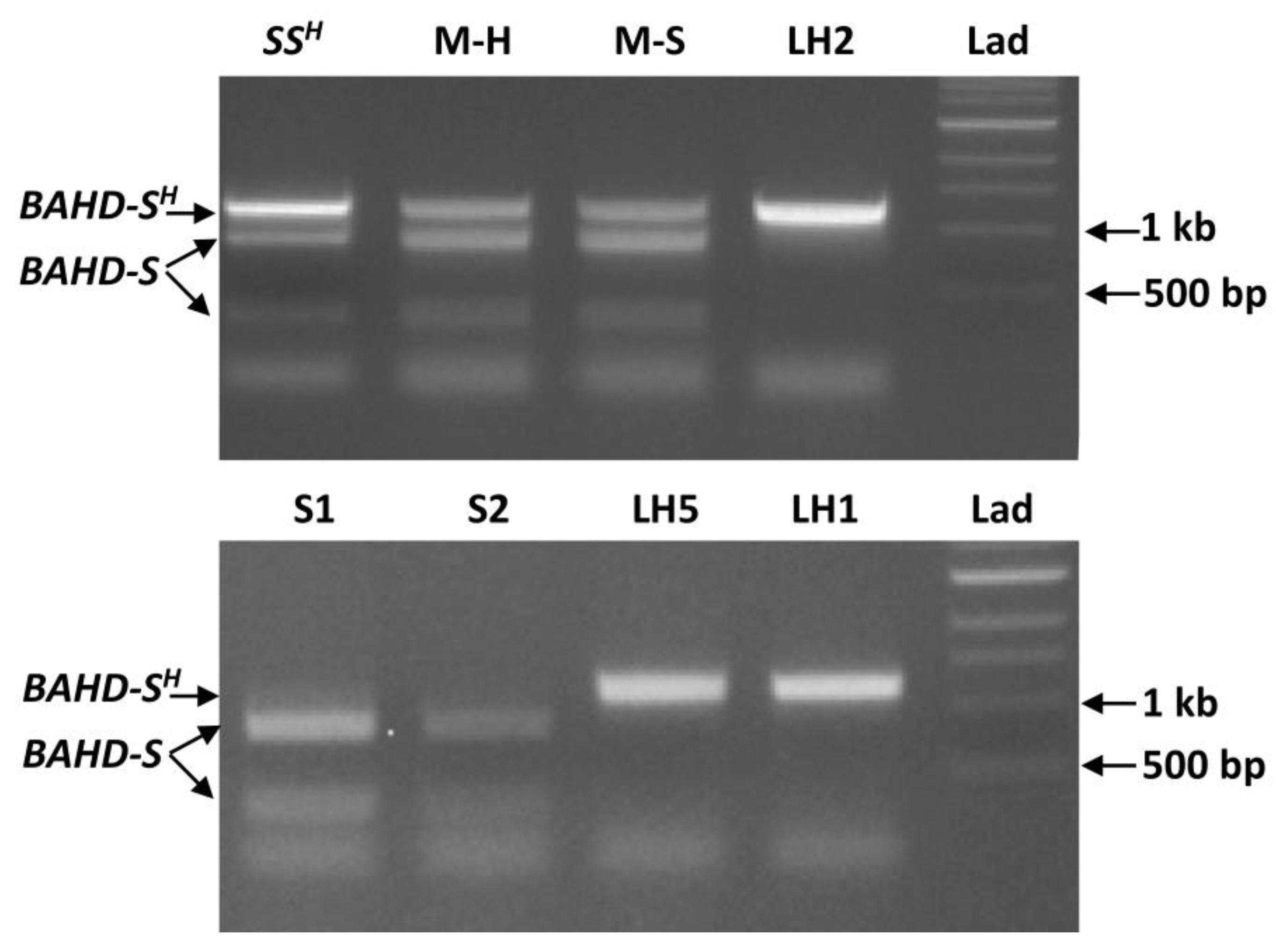{kind=link}
Abstract
1. Introduction
2. Results
2.1. Analysis of BAHD from Homostyle Species
2.2. S-Gene Expression in the Long-Homostyle Mutant Mhomo-H
2.3. BAHD Sequence Analysis
2.4. Does the Mutant BAHD in Mhomo-H Possess Brassinosteroid Activity?
2.5. Do BRs Act Directly on Pollen to Mediate Self-Incompatibility in Turnera?
2.6. Is the SH Allele in Mhomo-H Present as a Periclinal Chimera?
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Genomic PCR
4.3. RT-PCR
4.4. Purification and Mfe I Digestion of BAHD Amplicons
4.5. Plasmid Construction and Generation and Analysis of Transgenic Lines
4.6. Pollen Germination and Growth Assays
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Richards, A.J. Plant Breeding Systems, 2nd ed.; Chapman and Hall: London, UK, 1997. [Google Scholar]
- Igic, B.; Kohn, J.R. The distribution of plant mating systems: Study bias against obligately outcrossing species. Evolution 2006, 60, 1098–1103. [Google Scholar] [CrossRef]
- Bateson, W.; Gregory, R.P. On the inheritance of heterostylism in Primula. Proc. R. Soc. Lond. Ser. B 1905, 76, 581–586. [Google Scholar] [CrossRef]
- Barrett, S.C.H.; Cruzan, M.B. Incompatibility in heterostylous plants. In Genetic Control of Self-Incompatibility and Reproductive Development in Flowering Plants; Williams, E.G., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994; pp. 189–219. [Google Scholar] [CrossRef]
- Barrett, S.C.H. “A most complex marriage arrangement”, recent advances on heterostyly and unresolved questions. New Phytol. 2019, 224, 1051–1067. [Google Scholar] [CrossRef]
- Darwin, C. The Different Forms of Flowers on Plants of the Same Species; John Murray: London, UK, 1877. [Google Scholar]
- Ernst, A. Self-fertility in monomorphic. Primulas Genet. 1955, 27, 391–448. [Google Scholar] [CrossRef]
- Dowrick, V.P.J. Heterostyly and homostyly in Primula obconica. Heredity 1956, 10, 219–236. [Google Scholar]
- Lewis, D.; Jones, D.A. The genetics of heterostyly. In Evolution and Function of Heterostyly; Barrett, S.C.H., Ed.; Springer: Berlin/Heidelberg, Germany, 1992. [Google Scholar] [CrossRef]
- Mather, K. The genetical architecture of heterostyly in Primula sinensis. Evolution 1950, 4, 340–352. [Google Scholar] [CrossRef]
- Kappel, C.; Huu, C.N.; Lenhard, M. A short story gets longer: Recent insights into the molecular basis of heterostyly. J. Exp. Bot. 2017, 68, 5719–5730. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.D.; Russo, G.; Schlapbach, R.; Huu, C.N.; Lenhard, M.; Conti, E. The draft genome of Primula veris yields insights into the molecular basis of heterostyly. Genome Biol. 2015, 16, 12. [Google Scholar] [CrossRef]
- Li, J.; Cocker, J.M.; Wright, J.; Webster, M.A.; McMullan, M.; Dyer, S.; Swarbreck, D.; Caccamo, M.; Oosterhout, C.V.; Gilmartin, P.M. Genetic architecture and evolution of the S locus supergene in Primula vulgaris. Nat. Plants 2016, 2, 16188. [Google Scholar] [CrossRef]
- Huu, C.N.; Kappel, C.; Keller, B.; Sicard, A.; Takebayashi, Y.; Breuninger, H.; Nowak, M.D.; Bäurle, I.; Himmelbach, A.; Burkart, M.; et al. Presence versus absence of CYP734A50 underlies the style-length dimorphism in primroses. eLife 2016, 5, e17956. [Google Scholar] [CrossRef] [PubMed]
- Huu, C.N.; Keller, B.; Conti, E.; Kappel, C.; Lenhard, M. Supergene evolution via stepwise duplications and neofunctionalization of a floral-organ identity gene. Proc. Natl. Acad. Sci. USA 2020, 117, 23149–23157. [Google Scholar] [CrossRef]
- Burrows, B.A.; McCubbin, A.G. Sequencing the genomic regions flanking S-linked PvGLO sequences confirm the presence of two GLO loci, one of which lies adjacent to the style-length determinant gene CYP734A50. Plant Reprod. 2017, 30, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Yasui, Y.; Mori, M.; Aii, J.; Abe, T.; Masumoto, D.; Sato, S.; Hayashi, Y.; Ohnishi, O.; Ota, T. S-locus early flowering 3 is exclusively present in the genomes of short-styled buckwheat plants that exhibit heteromorphic self-incompatibility. PLoS ONE 2012, 7, e31264. [Google Scholar] [CrossRef] [PubMed]
- Yasui, Y.; Hirakawa, H.; Ueno, M.; Matsui, K.; Katsube-Tanaka, T.; Yang, S.J.; Aii, J.; Sato, S.; Mori, M. Assembly of the draft genome of buckwheat and its applications in identifying agronomically useful genes. DNA Res. 2016, 23, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Shore, J.S.; Hamam, H.J.; Chafe, P.D.J.; Labonne, J.D.J.; Henning, P.M.; McCubbin, A.G. The long and short of the S-locus in Turnera (Passifloraceae). New Phytol. 2019, 224, 1316–1329. [Google Scholar] [CrossRef]
- Matzke, C.M.; Shore, J.S.; Neff, M.M.; McCubbin, A.G. The Turnera style S-locus gene TsBAHD possesses brassinosteroid-inactivating activity when expressed in Arabidopsis thaliana. Plants 2020, 9, 1566. [Google Scholar] [CrossRef]
- Roh, H.; Jeong, C.W.; Fujioka, S.; Kim, Y.K.; Lee, S.; Ahn, J.H.; Choi, Y.D.; Lee, J.S. Genetic evidence for the reduction of brassinosteroid levels by a BAHD acyltransferase-like protein in Arabidopsis. Plant Physiol. 2012, 159, 696–709. [Google Scholar] [CrossRef] [PubMed]
- Tamari, F.; Khosravi, D.; Hilliker, A.J.; Shore, J.S. Inheritance of spontaneous mutant homostyles in Turnera subulata X krapovickasii and in autotetraploid T. scabra (Turneraceae). Heredity 2005, 94, 207–216. [Google Scholar] [CrossRef]
- Unno, H.; Ichimaida, F.; Suzuki, H.; Takahashi, S.; Tanaka, Y.; Saito, A.; Nishino, T.; Kusunoki, M.; Nakayama, T. Structural and mutational studies of anthocyanin malonyltransferases establish the features of BAHD enzyme catalysis. J. Biol. Chem. 2007, 282, 15812–15822. [Google Scholar] [CrossRef]
- Tuominen, L.K.; Johnson, V.E.; Tsai, C.-J. Differential phylogenetic expansions in BAHD acyltransferases across five angiosperm taxa and evidence of divergent expression among Populus paralogues. BMC Genom. 2011, 12, 236. [Google Scholar] [CrossRef]
- Wang, M.; Liu, X.; Wang, R.; Wanchun, L.; Rodermel, S.; Yu, F. Overexpression of a putative Arabidopsis BAHD acyltransferase causes dwarfism that can be rescued by brassinosteroid. J. Exp. Bot. 2012, 63, 5787–5801. [Google Scholar] [CrossRef]
- Li, J.; Nagpal, P.; Vitart, V.; McMorris, T.C.; Chory, J. A role for brassinosteroids in light-dependent development of Arabidopsis. Science 1996, 272, 398–401. [Google Scholar] [CrossRef]
- Szekeres, M.; Nemeth, K.; Kalman, Z.; Mathur, J.; Kauschmann, A.; Altmann, T.; Redei, G.P.; Nagy, F.; Schell, J.; Koncz, C. Brassinosteroids rescue the deficiency of CYP90, a cytochrome P450, controlling cell elongation and de-etiolation in Arabidopsis. Cell 1996, 85, 171–182. [Google Scholar] [CrossRef]
- Choe, S.; Dilkes, B.P.; Fujioka, S.; Takatsuto, S.; Sakurai, A.; Feldmann, K.A. The DWF4 gene of Arabidopsis encodes a Cytochrome P450 that mediates multiple 22-hydroxylation steps in brassinosteroid biosynthesis. Plant Cell 1998, 10, 231–243. [Google Scholar]
- Neff, M.M.; Nguyen, S.M.; Malancharuvil, E.J.; Fujioka, S.; Noguchi, T.; Seto, H.; Tsubuki, M.; Honda, T.; Takatsuto, S.; Yoshida, S.; et al. BAS1: A gene regulating brassinosteroid levels and light responsiveness in Arabidopsis. Proc. Natl. Acad. Sci. USA 1999, 96, 15316–15323. [Google Scholar] [CrossRef] [PubMed]
- Vogler, F.; Schmalzl, C.; Englhart, M.; Bircheneder, M.; Sprunck, S. Brassinosteroids promote Arabidopsis pollen germination and growth. Plant Reprod. 2014, 27, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Henning, P.M.; Shore, J.S.; McCubbin, A.G. Transcriptome and network analyses of heterostyly in Turnera subulata provide mechanistic insights: Are S-Loci a red-light for pistil elongation? Plants 2020, 9, 713. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Kushiro, T.; Yokota, T.; Kamiya, Y.; Bishop, G.J.; Yamaguchi, S. The last reaction producing brassinolide is catalyzed by cytochrome P-450s, CYP85A3 in tomato and CYP85A2 in Arabidopsis. J. Biol. Chem. 2005, 280, 17873–17879. [Google Scholar] [CrossRef] [PubMed]
- Tamari, F.; Athanasiou, A.; Shore, J.S. Pollen tube growth and inhibition in distylous and homostylous Turnera and Piriqueta (Turneraceae). Can. J. Bot. 2001, 79, 578–591. [Google Scholar]
- Freeling, M. Plant transposable elements and insertion sequences. Ann. Rev. Plant Physiol. 1984, 35, 277–298. [Google Scholar] [CrossRef]
- López, A.; Aveliano, F.; Shore, J.S. Inferences on the origins of polyploid Turnera species (Passifloraceae) based on molecular data. Botany 2013, 91, 167–175. [Google Scholar] [CrossRef]
- Labonne, J.D.J.; Goultiaeva, A.; Shore, J.S. High-resolution mapping of the S-locus in Turnera leads to the discovery of three genes tightly associated with the S-alleles. Mol. Genet. Genom. 2009, 281, 673–685. [Google Scholar] [CrossRef]
- Mable, B.K.; Schierup, M.H.; Charlesworth, D. Estimating the number, frequency, and dominance of S-alleles in a natural population of Arabidopsis lyrata (Brassicaceae) with sporophytic control of self-incompatibility. Heredity 2003, 90, 422–431. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hogenboom, N.G. Incompatibility and incongruity, two different mechanisms for the non-functioning of intimate partner relationships. Proc. R. Soc. Lond. Ser. B 1975, 188, 361–375. [Google Scholar]
- Lin, Z.; Eaves, D.J.; Sanchez-Moran, E.; Franklin, F.C.; Franklin-Tong, V.E. The Papaver rhoeas S determinants confer self-incompatibility to Arabidopsis thaliana in planta. Science 2015, 6, 684–687. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Li, J. Regulation of brassinosteroid homeostasis in higher plants. Front. Plant Sci. 2020, 11, 1480. [Google Scholar] [CrossRef]
- Labonne, J.D.J.; Shore, J.S. Positional cloning of the s haplotype determining the floral and incompatibility phenotype of the long-styled morph of distylous Turnera subulata. Mol. Gen. Genom. 2011, 285, 101–111. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matzke, C.M.; Hamam, H.J.; Henning, P.M.; Dougherty, K.; Shore, J.S.; Neff, M.M.; McCubbin, A.G. Pistil Mating Type and Morphology Are Mediated by the Brassinosteroid Inactivating Activity of the S-Locus Gene BAHD in Heterostylous Turnera Species. Int. J. Mol. Sci. 2021, 22, 10603. https://doi.org/10.3390/ijms221910603
Matzke CM, Hamam HJ, Henning PM, Dougherty K, Shore JS, Neff MM, McCubbin AG. Pistil Mating Type and Morphology Are Mediated by the Brassinosteroid Inactivating Activity of the S-Locus Gene BAHD in Heterostylous Turnera Species. International Journal of Molecular Sciences. 2021; 22(19):10603. https://doi.org/10.3390/ijms221910603
Chicago/Turabian StyleMatzke, Courtney M., Hasan J. Hamam, Paige M. Henning, Kyra Dougherty, Joel S. Shore, Michael M. Neff, and Andrew G. McCubbin. 2021. "Pistil Mating Type and Morphology Are Mediated by the Brassinosteroid Inactivating Activity of the S-Locus Gene BAHD in Heterostylous Turnera Species" International Journal of Molecular Sciences 22, no. 19: 10603. https://doi.org/10.3390/ijms221910603
APA StyleMatzke, C. M., Hamam, H. J., Henning, P. M., Dougherty, K., Shore, J. S., Neff, M. M., & McCubbin, A. G. (2021). Pistil Mating Type and Morphology Are Mediated by the Brassinosteroid Inactivating Activity of the S-Locus Gene BAHD in Heterostylous Turnera Species. International Journal of Molecular Sciences, 22(19), 10603. https://doi.org/10.3390/ijms221910603