
Molecular Insights into SARS-CoV2-Induced Alterations of the Gut/Brain Axis


Abstract
:1. Introduction
2. Results
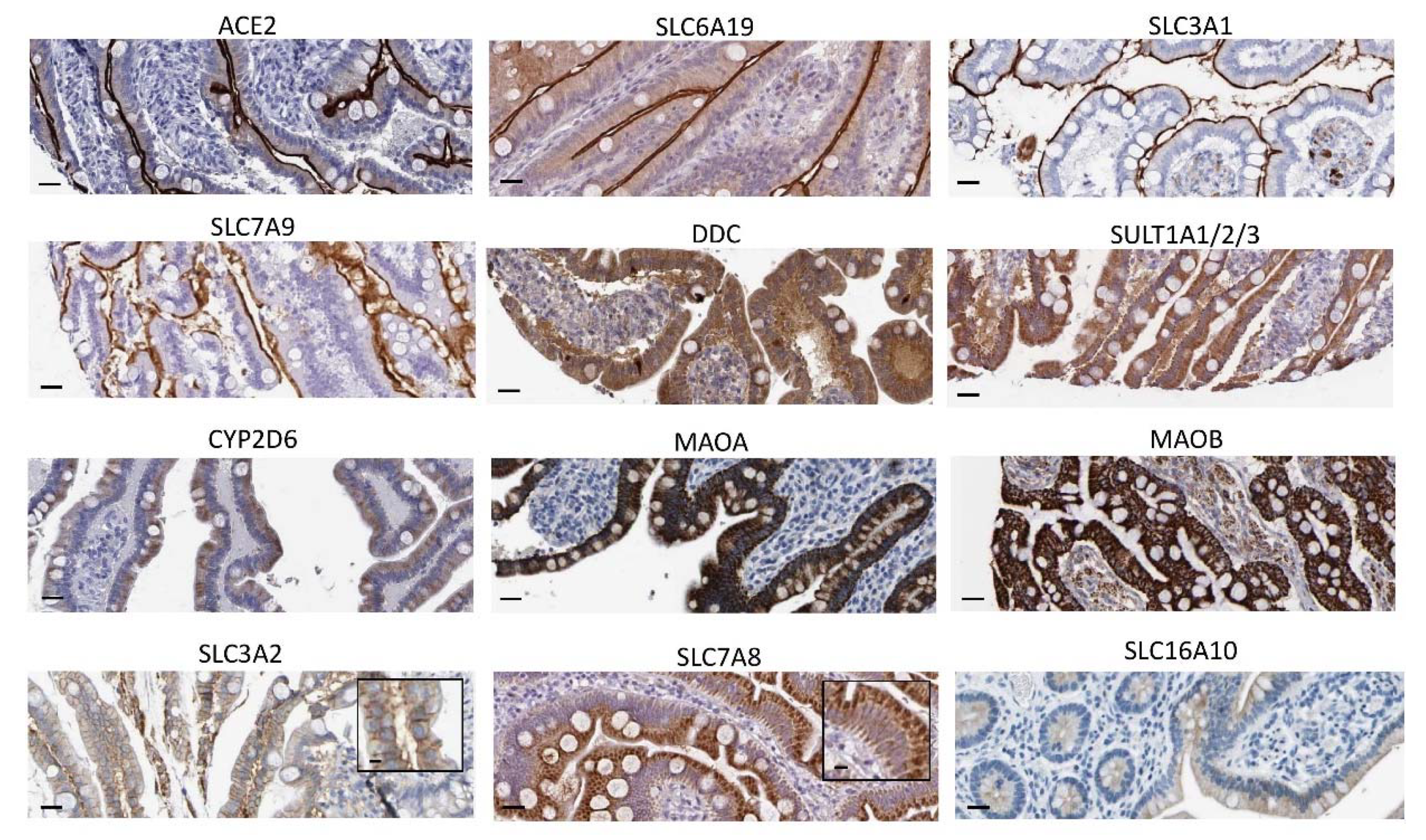
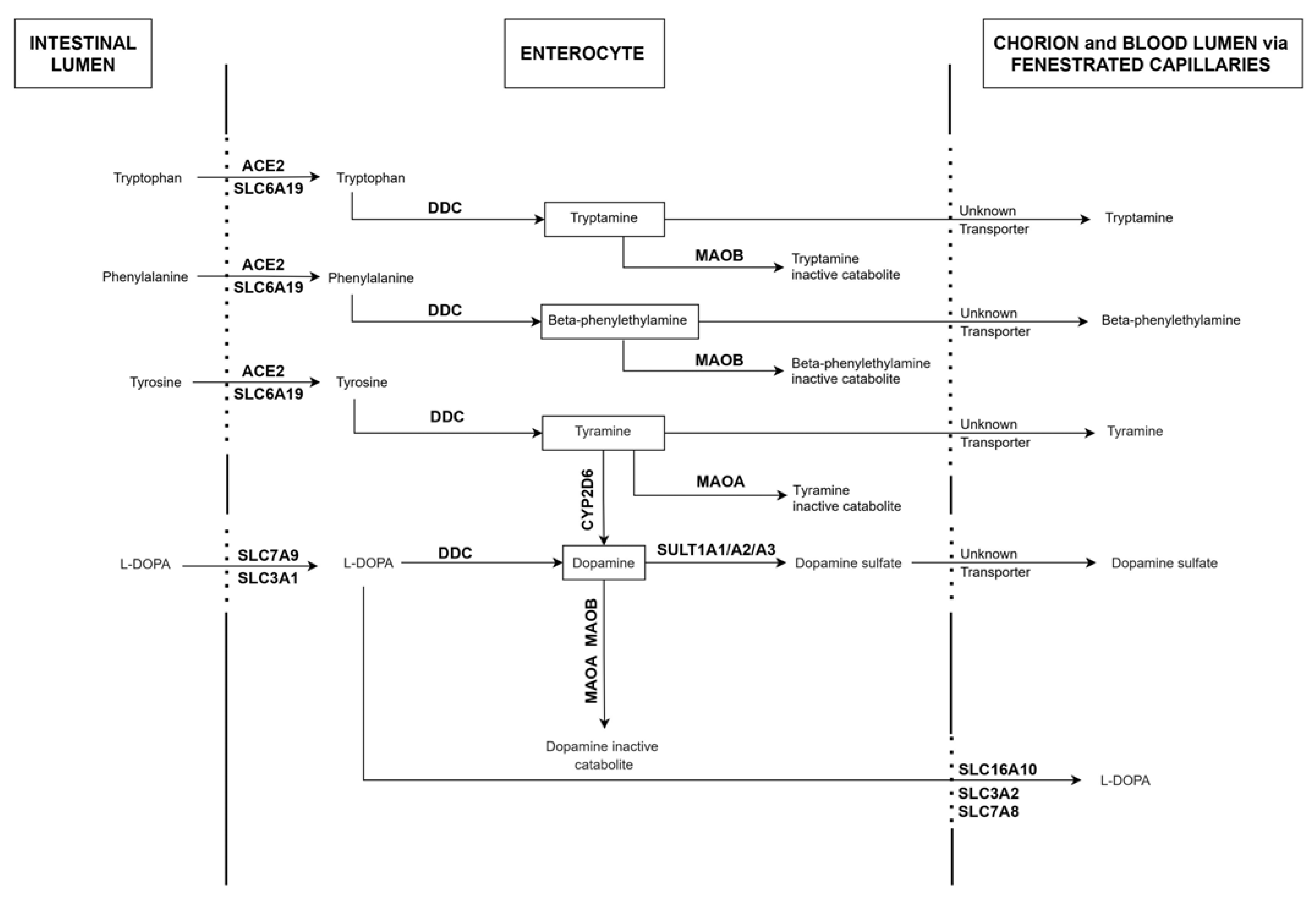
2.1. Expression Patterns of ACE2, DDC and Key Genes of the Dopamine/Trace Amines Synthetic Pathways in Enterocytes of the Human Small Intestine
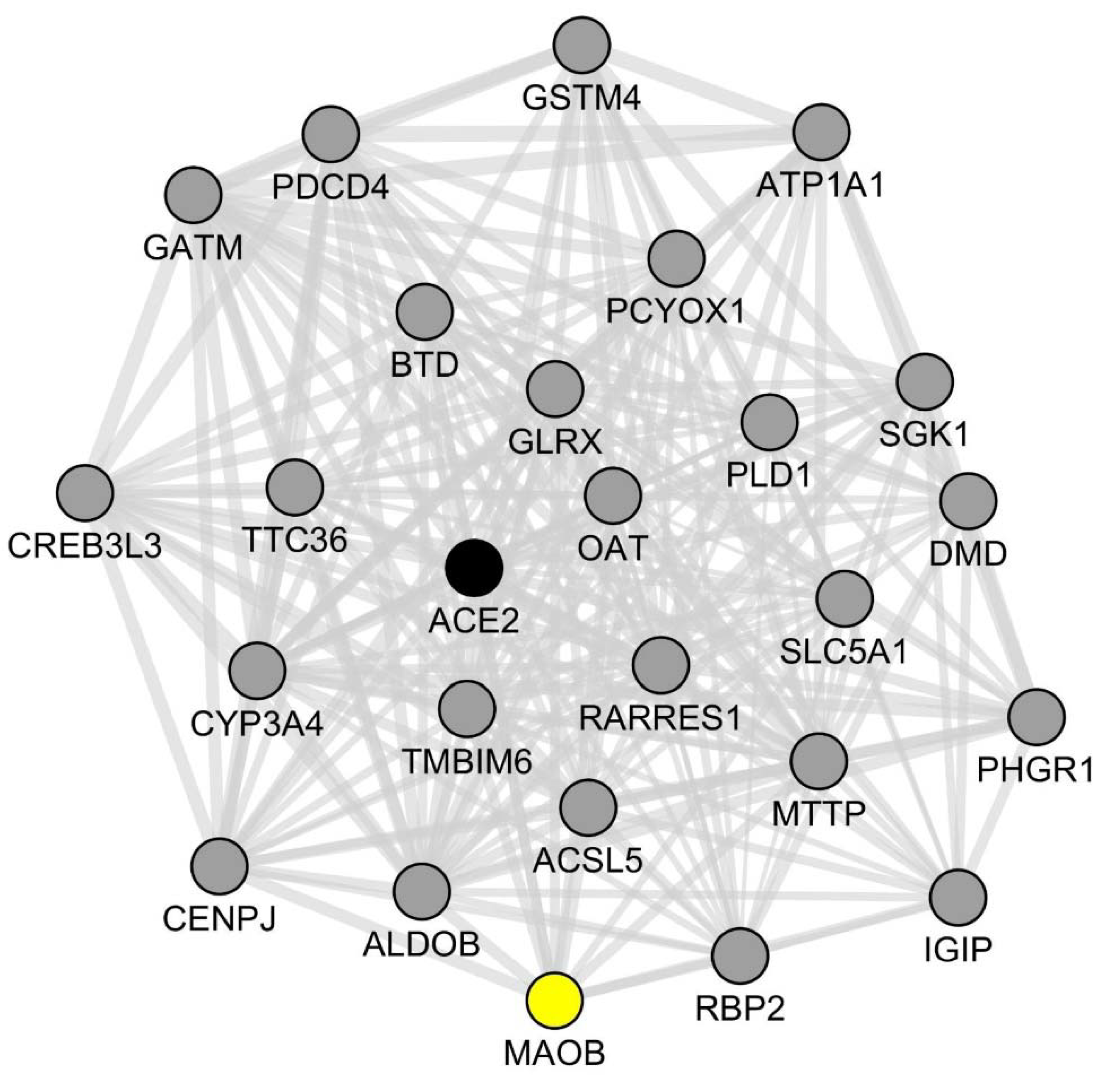
2.2. Assessment of Co-Expression Links between ACE2 and Key Genes of the Dopamine/Trace Amines Metabolic Pathways in SARS-CoV2-Infected Human Intestinal Organoids
3. Discussion
4. Materials and Methods
4.1. Mining of Human Expression Atlases
- ∗
- ∗
- Cytochrome P450 family 2 subfamily D member 6 (CYP2D6): an enzyme allowing the synthesis of dopamine from tyramine [43];
- ∗
- Solute carrier family 7 member 9 (SLC7A9) and solute carrier family 3 member 1 (SLC3A1): transporters allowing the cellular influx of L-DOPA [75];
- ∗
- Solute carrier family 6 member 10 (SLC16A10), solute carrier family 7 member 8 (SLC7A8) and solute carrier family 3 member 2 (SLC3A2): transporters allowing the cellular efflux of L-DOPA [75];
- ∗
- ∗
- ∗
4.2. Gene Co-Expression Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taquet, M.; Luciano, S.; Geddes, J.R.; Harrison, P.J. Bidirectional associations between COVID-19 and psychiatric disorder: Retrospective cohort studies of 62 354 COVID-19 cases in the USA. Lancet Psychiatry 2021, 8, 130–140. [Google Scholar] [CrossRef]
- Helms, J.; Kremer, S.; Merdji, H.; Clere-Jehl, R.; Schenck, M.; Kummerlen, C.; Collange, O.; Boulay, C.; Fafi-Kremer, S.; Ohana, M.; et al. Neurologic Features in Severe SARS-CoV-2 Infection. N. Engl. J. Med. 2020, 382, 2268–2270. [Google Scholar] [CrossRef]
- Pun, B.T.; Badenes, R.; Heras La Calle, G.; Orun, O.M.; Chen, W.; Raman, R.; Simpson, B.G.K.; Wilson-Linville, S.; Hinojal Olmedillo, B.; Vallejo de la Cueva, A.; et al. Prevalence and risk factors for delirium in critically ill patients with COVID-19 (COVID-D): A multicentre cohort study. Lancet Respir. Med. 2021, 9, 239–250. [Google Scholar] [CrossRef]
- Graham, E.L.; Clark, J.R.; Orban, Z.S.; Lim, P.H.; Szymanski, A.L.; Taylor, C.; DiBiase, R.M.; Jia, D.T.; Balabanov, R.; Ho, S.U.; et al. Persistent neurologic symptoms and cognitive dysfunction in non-hospitalized COVID-19 “long haulers”. Ann. Clin. Transl. Neurol. 2021, 8, 1073–1085. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Veldhuis, A.; Malhotra, T. Neuropsychiatric and Cognitive Sequelae of COVID-19. Front. Psychol. 2021, 12, 577529. [Google Scholar] [CrossRef]
- Taquet, M.; Geddes, J.R.; Husain, M.; Luciano, S.; Harrison, P.J. 6-month neurological and psychiatric outcomes in 236 379 survivors of COVID-19: A retrospective cohort study using electronic health records. Lancet Psychiatry 2021, 8, 416–427. [Google Scholar] [CrossRef]
- Marshall, M. COVID and the brain: Researchers zero in on how damage occurs. Nature 2021, 595, 484–485. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, M.; Garcia, G.; Tian, E.; Cui, Q.; Chen, X.; Sun, G.; Wang, J.; Arumugaswami, V.; Shi, Y. ApoE-Isoform-Dependent SARS-CoV-2 Neurotropism and Cellular Response. Cell Stem Cell 2021, 28, 331–342.e5. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Bai, W.Z.; Hashikawa, T. The neuroinvasive potential of SARS-CoV2 may play a role in the respiratory failure of COVID-19 patients. J. Med. Virol. 2020, 92, 552–555. [Google Scholar] [CrossRef] [PubMed]
- De Melo, G.D.; Lazarini, F.; Levallois, S.; Hautefort, C.; Michel, V.; Larrous, F.; Verillaud, B.; Aparicio, C.; Wagner, S.; Gheusi, G.; et al. COVID-19-related anosmia is associated with viral persistence and inflammation in human olfactory epithelium and brain infection in hamsters. Sci. Transl. Med. 2021, 13, eabf8396. [Google Scholar] [CrossRef] [PubMed]
- Troyer, E.A.; Kohn, J.N.; Hong, S. Are we facing a crashing wave of neuropsychiatric sequelae of COVID-19? Neuropsychiatric symptoms and potential immunologic mechanisms. Brain. Behav. Immun. 2020, 87, 34–39. [Google Scholar] [CrossRef]
- Guo, Q.; Zheng, Y.; Shi, J.; Wang, J.; Li, G.; Li, C.; Fromson, J.A.; Xu, Y.; Liu, X.; Xu, H.; et al. Immediate psychological distress in quarantined patients with COVID-19 and its association with peripheral inflammation: A mixed-method study. Brain. Behav. Immun. 2020, 88, 17–27. [Google Scholar] [CrossRef]
- Thakur, K.T.; Miller, E.H.; Glendinning, M.D.; Al-Dalahmah, O.; Banu, M.A.; Boehme, A.K.; Boubour, A.L.; Bruce, S.S.; Chong, A.M.; Claassen, J.; et al. COVID-19 neuropathology at Columbia University Irving Medical Center/New York Presbyterian Hospital. Brain 2021, awab148, in press. [Google Scholar] [CrossRef]
- Qin, Y.; Wu, J.; Chen, T.; Li, J.; Zhang, G.; Wu, D.; Zhou, Y.; Zheng, N.; Cai, A.; Ning, Q.; et al. Long-term microstructure and cerebral blood flow changes in patients recovered from COVID-19 without neurological manifestations. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef]
- Sashindranath, M.; Nandurkar, H.H. Endothelial dysfunction in the brain: Setting the stage for stroke and other cerebrovascular complications of COVID-19. Stroke 2021, 52, 1895–1904. [Google Scholar] [CrossRef]
- Che Mohd Nassir, C.M.N.; Hashim, S.; Wong, K.K.; Abdul Halim, S.; Idris, N.S.; Jayabalan, N.; Guo, D.; Mustapha, M. COVID-19 Infection and Circulating Microparticles—Reviewing Evidence as Microthrombogenic Risk Factor for Cerebral Small Vessel Disease. Mol. Neurobiol. 2021, 58, 4188–4215. [Google Scholar] [CrossRef] [PubMed]
- Nataf, S. An alteration of the dopamine synthetic pathway is possibly involved in the pathophysiology of COVID-19. J. Med. Virol. 2020, 92, 1743–1744. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.Y.; Juorio, A.V. Aromatic l-amino acid decarboxylase: Biological characterization and functional role. Gen. Pharmacol. 1995, 26, 681–696. [Google Scholar] [CrossRef]
- Zang, R.; Castro, M.F.G.; McCune, B.T.; Zeng, Q.; Rothlauf, P.W.; Sonnek, N.M.; Liu, Z.; Brulois, K.F.; Wang, X.; Greenberg, H.B.; et al. TMPRSS2 and TMPRSS4 promote SARS-CoV-2 infection of human small intestinal enterocytes. Sci. Immunol. 2020, 5, eabc3582. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, H.B.; Lyu, J.R.; Lei, X.M.; Li, W.; Wu, G.; Lyu, J.; Dai, Z.M. Specific ACE2 expression in small intestinal enterocytes may cause gastrointestinal symptoms and injury after 2019-nCoV infection. Int. J. Infect. Dis. 2020, 96, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Hamming, I.; Timens, W.; Bulthuis, M.L.C.; Lely, A.T.; Navis, G.J.; van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Camargo, S.M.R.; Singer, D.; Makrides, V.; Huggel, K.; Pos, K.M.; Wagner, C.A.; Kuba, K.; Danilczyk, U.; Skovby, F.; Kleta, R.; et al. Tissue-specific amino acid transporter partners ACE2 and collectrin differentially interact with hartnup mutations. Gastroenterology 2009, 136, 872–882. [Google Scholar] [CrossRef] [Green Version]
- Martin, A.M.; Young, R.L.; Leong, L.; Rogers, G.B.; Spencer, N.J.; Jessup, C.F.; Keating, D.J. The diverse metabolic roles of peripheral serotonin. Endocrinology 2017, 158, 1049–1063. [Google Scholar] [CrossRef]
- Weng, J.; Li, Y.; Li, J.; Shen, L.; Zhu, L.; Liang, Y.; Lin, X.; Jiao, N.; Cheng, S.; Huang, Y.; et al. Gastrointestinal sequelae 90 days after discharge for COVID-19. Lancet Gastroenterol. Hepatol. 2021, 6, 344–346. [Google Scholar] [CrossRef]
- Livanos, A.E.; Jha, D.; Cossarini, F.; Gonzalez-Reiche, A.S.; Tokuyama, M.; Aydillo, T.; Parigi, T.L.; Ladinsky, M.S.; Ramos, I.; Dunleavy, K.; et al. Intestinal Host Response to SARS-CoV-2 Infection and COVID-19 Outcomes in Patients with Gastrointestinal Symptoms. Gastroenterology 2021, 160, 2435–2450.e34. [Google Scholar] [CrossRef]
- Gaebler, C.; Wang, Z.; Lorenzi, J.C.C.; Muecksch, F.; Finkin, S.; Tokuyama, M.; Cho, A.; Jankovic, M.; Schaefer-Babajew, D.; Oliveira, T.Y.; et al. Evolution of antibody immunity to SARS-CoV-2. Nature 2021, 591, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Perlot, T.; Rehman, A.; Trichereau, J.; Ishiguro, H.; Paolino, M.; Sigl, V.; Hanada, T.; Hanada, R.; Lipinski, S.; et al. ACE2 links amino acid malnutrition to microbial ecology and intestinal inflammation. Nature 2012, 487, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Valles-Colomer, M.; Falony, G.; Darzi, Y.; Tigchelaar, E.F.; Wang, J.; Tito, R.Y.; Schiweck, C.; Kurilshikov, A.; Joossens, M.; Wijmenga, C.; et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat. Microbiol. 2019, 4, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Nowak, J.K.; Lindstrøm, J.C.; Kalla, R.; Ricanek, P.; Halfvarson, J.; Satsangi, J. Age, Inflammation, and Disease Location Are Critical Determinants of Intestinal Expression of SARS-CoV-2 Receptor ACE2 and TMPRSS2 in Inflammatory Bowel Disease. Gastroenterology 2020, 159, 1151–1154.e2. [Google Scholar] [CrossRef]
- Potdar, A.A.; Dube, S.; Naito, T.; Li, K.; Botwin, G.; Haritunians, T.; Li, D.; Casero, D.; Yang, S.; Bilsborough, J.; et al. Altered Intestinal ACE2 Levels Are Associated with Inflammation, Severe Disease, and Response to Anti-Cytokine Therapy in Inflammatory Bowel Disease. Gastroenterology 2021, 160, 809–822.e7. [Google Scholar] [CrossRef]
- Verstockt, B.; Verstockt, S.; Abdu Rahiman, S.; Ke, B.-J.; Arnauts, K.; Cleynen, I.; Sabino, J.; Ferrante, M.; Matteoli, G.; Vermeire, S. Intestinal Receptor of SARS-CoV-2 in Inflamed IBD Tissue Seems Downregulated by HNF4A in Ileum and Upregulated by Interferon Regulating Factors in Colon. J. Crohns. Colitis 2021, 15, 485–498. [Google Scholar] [CrossRef]
- Barberio, B.; Zamani, M.; Black, C.J.; Savarino, E.V.; Ford, A.C. Prevalence of symptoms of anxiety and depression in patients with inflammatory bowel disease: A systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 2021, 6, 359–370. [Google Scholar] [CrossRef]
- Bernstein, C.N.; Hitchon, C.A.; Walld, R.; Bolton, J.M.; Sareen, J.; Walker, J.R.; Graff, L.A.; Patten, S.B.; Singer, A.; Lix, L.M.; et al. Increased Burden of Psychiatric Disorders in Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2019, 25, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Lamers, M.M.; Beumer, J.; Van Der Vaart, J.; Knoops, K.; Puschhof, J.; Breugem, T.I.; Ravelli, R.B.G.; Van Schayck, J.P.; Mykytyn, A.Z.; Duimel, H.Q.; et al. SARS-CoV-2 productively infects human gut enterocytes. Science 2020, 369, 50–54. [Google Scholar] [CrossRef]
- Wang, Y.; Song, W.; Wang, J.; Wang, T.; Xiong, X.; Qi, Z.; Fu, W.; Yang, X.; Chen, Y.-G. Single-cell transcriptome analysis reveals differential nutrient absorption functions in human intestine. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef]
- He, X.C.; Li, L. Atlas of the human intestine. J. Exp. Med. 2020, 217, e20191130. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.; Grishagin, I.; Wang, Y.; Zhao, T.; Greene, J.; Obenauer, J.C.; Ngan, D.; Nguyen, D.T.; Guha, R.; Jadhav, A.; et al. The NCATS BioPlanet—An integrated platform for exploring the universe of cellular signaling pathways for toxicology, systems biology, and chemical genomics. Front. Pharmacol. 2019, 10, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, D.S.; Swoboda, K.J.; Miles, J.M.; Coppack, S.W.; Aneman, A.; Holmes, C.; Lamensdorf, I.; Eisenhofer, G. Sources and physiological significance of plasma dopamine sulfate. J. Clin. Endocrinol. Metab. 1999, 84, 2523–2531. [Google Scholar] [CrossRef] [PubMed]
- Apaydin, H.; Ertan, S.; Özekmekçi, S. Broad bean (Vicia faba)—A natural source of L-dopa—Prolongs “on” periods in patients with Parkinson’s disease who have “on-off” fluctuations. Mov. Disord. 2000, 15, 164–166. [Google Scholar] [CrossRef]
- Pereira, A.; Maraschin, M. Banana (Musa spp.) from peel to pulp: Ethnopharmacology, source of bioactive compounds and its relevance for human health. J. Ethnopharmacol. 2015, 160, 149–163. [Google Scholar] [CrossRef]
- Cilia, R.; Laguna, J.; Cassani, E.; Cereda, E.; Pozzi, N.G.; Isaias, I.U.; Contin, M.; Barichella, M.; Pezzoli, G. Mucuna pruriens in Parkinson disease: A double-blind, randomized, controlled, crossover study. Neurology 2017, 89, 432–438. [Google Scholar] [CrossRef] [Green Version]
- Ramya, K.B.; Thaakur, S. Herbs containing L—Dopa: An update. Anc. Sci. Life 2007, 27, 50–55. [Google Scholar]
- Hiroi, T.; Imaoka, S.; Funae, Y. Dopamine formation from tyramine by CYP2D6. Biochem. Biophys. Res. Commun. 1998, 249, 838–843. [Google Scholar] [CrossRef]
- Pletscher, A.; Kunz, E.; Staebler, H.; Gey, K.F. The uptake of tryptamine by brain in vivo and its alteration by drugs. Biochem. Pharmacol. 1963, 12, 1065–1070. [Google Scholar] [CrossRef]
- Vitale, A.A.; Pomilio, A.B.; Cañellas, C.O.; Vitale, M.G.; Putz, E.M.; Ciprian-Ollivier, J. In vivo long-term kinetics of radiolabeled N,N-dimethyltryptamine and tryptamine. J. Nucl. Med. 2011, 52, 970–977. [Google Scholar] [CrossRef] [Green Version]
- Oldendorf, W.H. Brain uptake of radiolabeled amino acids, amines, and hexoses after arterial injection. Am. J. Physiol. 1971, 221, 1629–1639. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Nwe, P.K.; Yang, Y.; Rosen, C.E.; Bielecka, A.A.; Kuchroo, M.; Cline, G.W.; Kruse, A.C.; Ring, A.M.; Crawford, J.M.; et al. A Forward Chemical Genetic Screen Reveals Gut Microbiota Metabolites That Modulate Host Physiology. Cell 2019, 177, 1217–1231.e18. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.D.; Canales, J.J.; Zucchi, R.; Espinoza, S.; Sukhanov, I.; Gainetdinov, R.R. Trace amine-associated receptor 1: A multimodal therapeutic target for neuropsychiatric diseases. Expert Opin. Ther. Targets 2018, 22, 513–526. [Google Scholar] [CrossRef]
- Burchett, S.A.; Hicks, T.P. The mysterious trace amines: Protean neuromodulators of synaptic transmission in mammalian brain. Prog. Neurobiol. 2006, 79, 223–246. [Google Scholar] [CrossRef] [PubMed]
- Gainetdinov, R.R.; Hoener, M.C.; Berry, M.D. Trace amines and their receptors. Pharmacol. Rev. 2018, 70, 549–620. [Google Scholar] [CrossRef] [Green Version]
- Marsden, C.A.; Curzon, G. The contribution of tryptamine to the behavioural effects of l-tryptophan in tranylcypromine-treated rats. Psychopharmacology 1978, 57, 71–76. [Google Scholar] [CrossRef]
- Bieck, P.R.; Antonin, K.H. Monoamine oxidase inhibition by tranylcypromine: Assessment in human volunteers. Eur. J. Clin. Pharmacol. 1982, 22, 301–308. [Google Scholar] [CrossRef]
- Grimsby, J.; Toth, M.; Chen, K.; Kumazawa, T.; Klaidman, L.; Adams, J.D.; Karoum, F.; Gal, J.; Shih, J.C. Increased stress response and p-phenylethylamine in maob-def icient mice. Nat. Genet. 1997, 17, 210. [Google Scholar] [CrossRef] [PubMed]
- Birkmayer, W.; Riederer, P.; Linauer, W.; Knoll, J. L-deprenyl plus l-phenylalanine in the treatment of depression. J. Neural Transm. 1984, 59, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Youdim, M.B.H.; Weinstock, M. Therapeutic Applications of Selective and Non-Selective Inhibitors of Monoamine Oxidase A and B that do not Cause Significant Tyramine Potentiation. Neurotoxicology 2004, 25, 243–250. [Google Scholar] [CrossRef]
- Finberg, J.P.M.; Gillman, K. Selective inhibitors of monoamine oxidase type B and the “cheese effect”. Int. Rev. Neurobiol. 2011, 100, 169–190. [Google Scholar] [PubMed]
- Rossi, Á.D.; de Araújo, J.L.F.; de Almeida, T.B.; Ribeiro-Alves, M.; de Almeida Velozo, C.; de Almeida, J.M.; de Carvalho Leitão, I.; Ferreira, S.N.; da Silva Oliveira, J.; Alves, H.J.; et al. Association between ACE2 and TMPRSS2 nasopharyngeal expression and COVID-19 respiratory distress. Sci. Rep. 2021, 11, 9658. [Google Scholar] [CrossRef]
- Sun, J.; Ye, F.; Wu, A.; Yang, R.; Pan, M.; Sheng, J.; Zhu, W.; Mao, L.; Wang, M.; Xia, Z.; et al. Comparative Transcriptome Analysis Reveals the Intensive Early Stage Responses of Host Cells to SARS-CoV-2 Infection. Front. Microbiol. 2020, 11, 593857. [Google Scholar] [CrossRef]
- Kuba, K.; Imai, Y.; Rao, S.; Gao, H.; Guo, F.; Guan, B.; Huan, Y.; Yang, P.; Zhang, Y.; Deng, W.; et al. A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus-induced lung injury. Nat. Med. 2005, 11, 875–879. [Google Scholar] [CrossRef]
- Singer, D.; Camargo, S.M.R.; Ramadan, T.; Schäfer, M.; Mariotta, L.; Herzog, B.; Huggel, K.; Wolfer, D.; Werner, S.; Penninger, J.M.; et al. Defective intestinal amino acid absorption in Ace2 null mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G686–G695. [Google Scholar] [CrossRef]
- Alenina, N.; Bader, M. ACE2 in Brain Physiology and Pathophysiology: Evidence from Transgenic Animal Models. Neurochem. Res. 2019, 44, 1323–1329. [Google Scholar] [CrossRef]
- Tahmoush, A.J.; Alpers, D.H.; Feigin, R.D.; Armbrustmacher, V.; Prensky, A.L. Hartnup Disease: Clinical, Pathological, and Biochemical Observations. Arch. Neurol. 1976, 33, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Žigman, T.; Petković Ramadža, D.; Šimić, G.; Barić, I. Inborn Errors of Metabolism Associated with Autism Spectrum Disorders: Approaches to Intervention. Front. Neurosci. 2021, 15, 673600. [Google Scholar] [CrossRef] [PubMed]
- Conio, B.; Martino, M.; Magioncalda, P.; Escelsior, A.; Inglese, M.; Amore, M.; Northoff, G. Opposite effects of dopamine and serotonin on resting-state networks: Review and implications for psychiatric disorders. Mol. Psychiatry 2020, 25, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Rutigliano, G.; Zucchi, R. Molecular Variants in Human Trace Amine-Associated Receptors and Their Implications in Mental and Metabolic Disorders. Cell. Mol. Neurobiol. 2020, 40, 239–255. [Google Scholar] [CrossRef] [Green Version]
- Arjmand, S.; Behzadi, M.; Stephens, G.J.; Ezzatabadipour, S.; Seifaddini, R.; Arjmand, S.; Shabani, M. A Brain on a Roller Coaster: Can the Dopamine Reward System Act as a Protagonist to Subdue the Ups and Downs of Bipolar Disorder? Neuroscientist 2018, 24, 423–439. [Google Scholar] [CrossRef]
- Berlet, H.H.; Pscheidt, G.R.; Spaide, J.K.; Himwich, H.E. Variations of urinary creatinine and its correlation to tryptamine excretion in schizophrenic patients. Nature 1964, 203, 1198–1199. [Google Scholar] [CrossRef]
- O’reilly, R.L.; Davis, B.A. Phenylethylamine and schizophrenia. Prog. Neuropsychopharmacol. Biol. Psychiatry 1994, 18, 63–75. [Google Scholar] [CrossRef]
- McCutcheon, R.A.; Abi-Dargham, A.; Howes, O.D. Schizophrenia, Dopamine and the Striatum: From Biology to Symptoms. Trends Neurosci. 2019, 42, 205–220. [Google Scholar] [CrossRef] [Green Version]
- Acharya, S.; Kim, K.M. Roles of the functional interaction between brain cholinergic and dopaminergic systems in the pathogenesis and treatment of schizophrenia and Parkinson’s disease. Int. J. Mol. Sci. 2021, 22, 4299. [Google Scholar] [CrossRef]
- Thomas, T.; Stefanoni, D.; Reisz, J.A.; Nemkov, T.; Bertolone, L.; Francis, R.O.; Hudson, K.E.; Zimring, J.C.; Hansen, K.C.; Hod, E.A.; et al. COVID-19 infection alters kynurenine and fatty acid metabolism, correlating with IL-6 levels and renal status. JCI Insight 2020, 5, e140327. [Google Scholar] [CrossRef]
- Song, J.W.; Lam, S.M.; Fan, X.; Cao, W.J.; Wang, S.Y.; Tian, H.; Chua, G.H.; Zhang, C.; Meng, F.P.; Xu, Z.; et al. Omics-Driven Systems Interrogation of Metabolic Dysregulation in COVID-19 Pathogenesis. Cell Metab. 2020, 32, 188–202.e5. [Google Scholar] [CrossRef] [PubMed]
- Richard, K.; Hume, R.; Kaptein, E.; Stanley, E.L.; Visser, T.J.; Coughtrie, M.W.H. Sulfation of Thyroid Hormone and Dopamine during Human Development: Ontogeny of Phenol Sulfotransferases and Arylsulfatase in Liver, Lung, and Brain 1. J. Clin. Endocrinol. Metab. 2001, 86, 2734–2742. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tong, Q.; Ma, S.R.; Zhao, Z.X.; Bin Pan, L.; Cong, L.; Han, P.; Peng, R.; Yu, H.; Lin, Y.; et al. Oral berberine improves brain dopa/dopamine levels to ameliorate Parkinson’s disease by regulating gut microbiota. Signal. Transduct. Target. Ther. 2021, 6, 77. [Google Scholar] [CrossRef] [PubMed]
- Camargo, S.M.R.; Vuille-Dit-Bille, R.N.; Mariotta, L.; Ramadan, T.; Huggel, K.; Singer, D.; Götze, O.; Verrey, F. The molecular mechanism of intestinal levodopa absorption and its possible implications for the treatment of Parkinson’s disease. J. Pharmacol. Exp. Ther. 2014, 351, 114–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowsher, R.R.; Henry, D.P. Decarboxylation of p-Tyrosine: A Potential Source of p-Tyramine in Mammalian Tissues. J. Neurochem. 1983, 40, 992–1002. [Google Scholar] [CrossRef] [PubMed]
- Youdim, M.B.H.; Riederer, P.F. A review of the mechanisms and role of monoamine oxidase inhibitors in Parkinson’s. Neurology 2004, 63, S32–S35. [Google Scholar] [CrossRef]
- Tsugeno, Y.; Ito, A. A key amino acid responsible for substrate selectivity of monoamine oxidase A and B. J. Biol. Chem. 1997, 272, 14033–14036. [Google Scholar] [CrossRef] [Green Version]
- Murphy, D.L.; Karoum, F.; Pickar, D.; Cohen, R.M.; Lipper, S.; Mellow, A.M.; Tariot, P.N.; Sunderland, T. Differential trace amine alterations in individuals receiving acetylenic inhibitors of MAO-A (clorgyline) or MAO-B (selegiline and pargyline). J. Neural Transm. Suppl. 1998, 52, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Godefroy, D.; Rostène, W.; Anouar, Y.; Goazigo, A.R. Le Tyrosine-hydroxylase immunoreactivity in the mouse transparent brain and adrenal glands. J. Neural Transm. 2019, 126, 367–375. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, S.; Arakawa, T.; Fukuda, S.; Furuno, M.; Hasegawa, A.; Hori, F.; Ishikawa-Kato, S.; Kaida, K.; Kaiho, A.; Kanamori-Katayama, M.; et al. FANTOM5 CAGE profiles of human and mouse samples. Sci. Data 2017, 4, 170112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonsdale, J.; Thomas, J.; Salvatore, M.; Phillips, R.; Lo, E.; Shad, S.; Hasz, R.; Walters, G.; Garcia, F.; Young, N.; et al. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software Environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Franz, M.; Rodriguez, H.; Lopes, C.; Zuberi, K.; Montojo, J.; Bader, G.D.; Morris, Q. GeneMANIA update 2018. Nucleic Acids Res. 2018, 46, W60–W64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nataf, S.; Barritault, M.; Pays, L. A unique TGFB1-driven genomic program links astrocytosis, low-grade inflammation and partial demyelination in spinal cord periplaques from progressive multiple sclerosis patients. Int. J. Mol. Sci. 2017, 18, 97. [Google Scholar] [CrossRef] [PubMed]
- Almeria, M.; Cejudo, J.C.; Sotoca, J.; Deus, J.; Krupinski, J. Cognitive profile following COVID-19 infection: Clinical predictors leading to neuropsychological impairment. Brain Behav. Immun. Heal. 2020, 9, 100163. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Rank Reported for the Small Intestine among 61 Human Tissues |
|---|---|
| ACE2 | 1 |
| SLC6A19 | 1 |
| SLC7A9 | 1 |
| SLC3A1 | 2 |
| SLC3A2 | 20 |
| SLC7A8 | 30 |
| SLC16A10 | 13 |
| DDC | 1 |
| MAOA | 1 |
| MAOB | 14 |
| CYP2D6 | 2 |
| SULT1A1 | 5 |
| SULT1A2 | 1 |
| SULT1A3 | 3 |
| TH | not detected |
| Cell Type and Intestinal Segment | Genes of Interest with Reported Presence in the Molecular Signatures |
|---|---|
| Enterocytes | |
| ileum | ACE2, SLC6A19, SLC7A9, SLC3A1, DDC, MAOA, MAOB, CYP2D6, SULT1A1, SULT1A2, SULT1A3 |
| colon rectum | none none |
| Enteroendocrine cells ileum colon rectum | ACE2, SLC6A19, SLC7A9, SLC3A1, MAOA, SULT1A2 none none |
| Paneth cells | |
| ileum | SLC6A19, SLC7A9, SLC3A1, DDC, MAOA, CYP2D6, SULT1A1, SULT1A2 |
| colon rectum | none none |
| Goblet cells ileum colon rectum | SLC7A9, DDC, MAOA, SULT1A1, SULT1A2 none none |
| Stem cells ileum colon rectum | DDC, MAOA, SLC3A1, none none |
| Transit amplifying cells illeum colon rectum | DDC, MAOA none none |
| DDC | MAOA | MAOB | SULT1A1 | SLC7A9 | SLC3A1 | SLC6A19 | SLC3A2 |
|---|---|---|---|---|---|---|---|
| 0.84 | 0.86 | 0.96 | 0.92 | 0.95 | 0.87 | 0.88 | 0.9 |
| 0.035 | 0.025 | 0.001 | 0.007 | 0.003 | 0.02 | 0.017 | 0.013 |
| Molecular Processes Putatively Targeted by SARS-CoV2 in Human Enterocytes | Molecules with Predictable Altered Blood Levels |
|---|---|
| Influx of L-DOPA (SLC3A1, SLC7A9) Efflux of L-DOPA (SLC16A10, SLC3A2, SLC7A8) Conversion of L-DOPA into Dopamine (DDC) Catabolism of Dopamine (MAO-A, MAO-B) Sulfation of Dopamine (SULT1A1, SULT1A2, SULT1A3) | L-DOPA Dopamine Dopamine sulfate |
| Influx of Tyrosine (ACE2, SLC6A19) Conversion of Tyrosine into tyramine (DDC) Catabolism of Tyramine (MAO-A) | Tyramine, Tyrosine Dopamine Dopamine sulfate |
| Influx of Phenylalanine (ACE2, SLC6A19) Conversion of Phenylalanine into β-PEA (DDC) Catabolism of β-PEA (MAO-B) | β-PEA, Phenylalanine |
| Influx of Tryptophan (ACE2, SLC6A19) Conversion of Tryptophan into Tryptamine (DDC) Catabolism of Tryptamine (MAO-B) | Tryptamine, Tryptophan |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nataf, S.; Pays, L. Molecular Insights into SARS-CoV2-Induced Alterations of the Gut/Brain Axis. Int. J. Mol. Sci. 2021, 22, 10440. https://doi.org/10.3390/ijms221910440
Nataf S, Pays L. Molecular Insights into SARS-CoV2-Induced Alterations of the Gut/Brain Axis. International Journal of Molecular Sciences. 2021; 22(19):10440. https://doi.org/10.3390/ijms221910440
Chicago/Turabian StyleNataf, Serge, and Laurent Pays. 2021. "Molecular Insights into SARS-CoV2-Induced Alterations of the Gut/Brain Axis" International Journal of Molecular Sciences 22, no. 19: 10440. https://doi.org/10.3390/ijms221910440
APA StyleNataf, S., & Pays, L. (2021). Molecular Insights into SARS-CoV2-Induced Alterations of the Gut/Brain Axis. International Journal of Molecular Sciences, 22(19), 10440. https://doi.org/10.3390/ijms221910440