
Exploring Aβ Proteotoxicity and Therapeutic Candidates Using Drosophila melanogaster


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Direction of Protein Expression to Drosophila Neurons
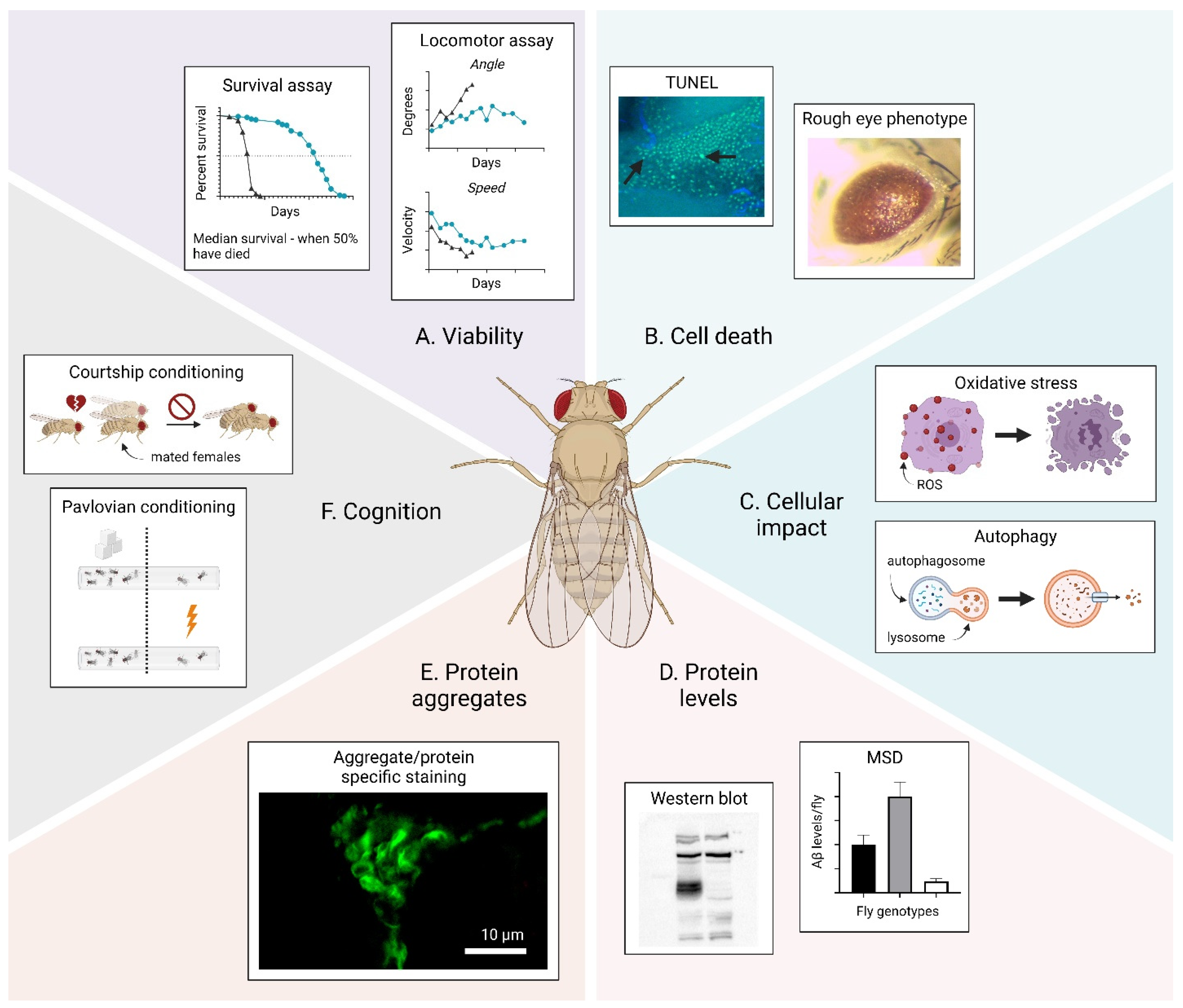
3. Methods to Study Proteotoxicity in Drosophila
4. Drosophila and Proteotoxicity of the Aβ Peptide
4.1. The History of Drosophila Aβ Models
4.2. Investigating Aβ Isoforms in Drosophila
4.3. The AβPP-BACE Fly
4.4. Relative Toxicity between Aβ Isoforms
5. Drosophila as Model Organism for Drug Screen against Aβ Proteotoxicity
5.1. Blocking Aβ Aggregation
5.2. Enhancing Aβ Aggregation
5.3. Increasing Protein Clearance
5.4. Proteins and Peptides as Drug Candidates
5.5. Targeting Inflammatory Processes
5.6. Preventing Oxidative Stress
5.7. Preventing Mitochondrial Dysfunction
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Takahashi, R.H.; Nagao, T.; Gouras, G.K. Plaque Formation and the Intraneuronal Accumulation of β-Amyloid in Alzheimer’s Disease. Pathol. Int. 2017, 67, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Alzheimer’s Association. 2016 Alzheimer’s Disease Facts and Figures. Alzheimers Dement. J. Alzheimers Assoc. 2016, 12, 459–509. [Google Scholar] [CrossRef] [PubMed]
- Roher, A.E.; Kokjohn, T.A.; Clarke, S.G.; Sierks, M.R.; Maarouf, C.L.; Serrano, G.E.; Sabbagh, M.S.; Beach, T.G. APP/Aβ Structural Diversity and Alzheimer’s Disease Pathogenesis. Neurochem. Int. 2017, 110, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Thal, D.R.; Walter, J.; Saido, T.C.; Fändrich, M. Neuropathology and Biochemistry of Aβ and Its Aggregates in Alzheimer’s Disease. Acta Neuropathol. 2015, 129, 167–182. [Google Scholar] [CrossRef]
- Brand, A.H.; Perrimon, N. Targeted Gene Expression as a Means of Altering Cell Fates and Generating Dominant Phenotypes. Dev. Camb. Engl. 1993, 118, 401–415. [Google Scholar]
- Yamaguchi, M.; Yoshida, H. Drosophila as a Model Organism. Adv. Exp. Med. Biol. 2018, 1076, 1–10. [Google Scholar] [CrossRef]
- Mhatre, S.D.; Paddock, B.E.; Saunders, A.J.; Marenda, D.R. Invertebrate Models of Alzheimer’s Disease. J. Alzheimers Dis. 2013, 33, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Rubin, G.M.; Spradling, A.C. Genetic Transformation of Drosophila with Transposable Element Vectors. Science 1982, 218, 348–353. [Google Scholar] [CrossRef]
- Osterwalder, T.; Yoon, K.S.; White, B.H.; Keshishian, H. A Conditional Tissue-Specific Transgene Expression System Using Inducible GAL4. Proc. Natl. Acad. Sci. USA 2001, 98, 12596–12601. [Google Scholar] [CrossRef]
- Berger, C.; Renner, S.; Lüer, K.; Technau, G.M. The Commonly Used Marker ELAV Is Transiently Expressed in Neuroblasts and Glial Cells in the Drosophila Embryonic CNS. Dev. Dyn. 2007, 236, 3562–3568. [Google Scholar] [CrossRef]
- Ellis, M.C.; O’Neill, E.M.; Rubin, G.M. Expression of Drosophila Glass Protein and Evidence for Negative Regulation of Its Activity in Non-Neuronal Cells by Another DNA-Binding Protein. Development 1993, 119, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-Z.; Li, S.-L.; Zheng, H.Y.; Zhang, S.-P.; Xue, L. A Broad Expression Profile of the GMR-GAL4 Driver in Drosophila melanogaster. Genet. Mol. Res. 2012, 11, 1997–2002. [Google Scholar] [CrossRef] [PubMed]
- van Dam, E.; van Leeuwen, L.A.G.; dos Santos, E.; James, J.; Best, L.; Lennicke, C.; Vincent, A.J.; Marinos, G.; Foley, A.; Buricova, M.; et al. Sugar-Induced Obesity and Insulin Resistance Are Uncoupled from Shortened Survival in Drosophila. Cell Metab. 2020, 31, 710–725.e7. [Google Scholar] [CrossRef]
- Bergkvist, L.; Sandin, L.; Kågedal, K.; Brorsson, A.-C. AβPP Processing Results in Greater Toxicity per Amount of Aβ1-42 than Individually Expressed and Secreted Aβ1-42 in Drosophila melanogaster. Biol. Open 2016, 5, 1030–1039. [Google Scholar] [CrossRef] [PubMed]
- Crowther, D.C.; Kinghorn, K.J.; Miranda, E.; Page, R.; Curry, J.A.; Duthie, F.A.I.; Gubb, D.C.; Lomas, D.A. Intraneuronal Aβ, Non-Amyloid Aggregates and Neurodegeneration in a Drosophila Model of Alzheimer’s Disease. Neuroscience 2005, 132, 123–135. [Google Scholar] [CrossRef]
- Scaplen, K.M.; Mei, N.J.; Bounds, H.A.; Song, S.L.; Azanchi, R.; Kaun, K.R. Automated Real-Time Quantification of Group Locomotor Activity in Drosophila melanogaster. Sci. Rep. 2019, 9, 4427. [Google Scholar] [CrossRef]
- Kohlhoff, K.J.; Jahn, T.R.; Lomas, D.A.; Dobson, C.M.; Crowther, D.C.; Vendruscolo, M. The IFly Tracking System for an Automated Locomotor and Behavioural Analysis of Drosophila melanogaster. Integr. Biol. Quant. Biosci. Nano Macro 2011, 3, 755–760. [Google Scholar] [CrossRef]
- Jeon, Y.; Lee, S.; Shin, M.; Lee, J.H.; Suh, Y.S.; Hwang, S.; Yun, H.S.; Cho, K.S. Phenotypic Differences between Drosophila Alzheimer’s Disease Models Expressing Human Aβ42 in the Developing Eye and Brain. Anim. Cells Syst. 2017, 21, 160–168. [Google Scholar] [CrossRef]
- Moore, B.D.; Martin, J.; de Mena, L.; Sanchez, J.; Cruz, P.E.; Ceballos-Diaz, C.; Ladd, T.B.; Ran, Y.; Levites, Y.; Kukar, T.L.; et al. Short Aβ Peptides Attenuate Aβ42 Toxicity in Vivo. J. Exp. Med. 2018, 215, 283–301. [Google Scholar] [CrossRef]
- Bergkvist, L.; Du, Z.; Elovsson, G.; Appelqvist, H.; Itzhaki, L.S.; Kumita, J.R.; Kågedal, K.; Brorsson, A. Mapping Pathogenic Processes Contributing to Neurodegeneration in Drosophila Models of Alzheimer’s Disease. FEBS Open Bio 2020, 10, 338–350. [Google Scholar] [CrossRef]
- Sarkissian, T.; Timmons, A.; Arya, R.; Abdelwahid, E.; White, K. Detecting Apoptosis in Drosophila Tissues and Cells. Methods 2014, 68, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Lőrincz, P.; Mauvezin, C.; Juhász, G. Exploring Autophagy in Drosophila. Cells 2017, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Denton, D.; Kumar, S. Autophagy-Dependent Cell Death. Cell Death Differ. 2019, 26, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, M.; Bollineni, R.C.; Hoffmann, R. Protein Carbonylation as a Major Hallmark of Oxidative Damage: Update of Analytical Strategies: PROTEIN CARBONYLATION: AN ANALYTICAL UPDATE. Mass Spectrom. Rev. 2014, 33, 79–97. [Google Scholar] [CrossRef] [PubMed]
- Ericson, B.L.; Carlson, D.J.; Carlson, K.A. Characterization of Nora Virus Structural Proteins via Western Blot Analysis. Scientifica 2016, 2016, 1–8. [Google Scholar] [CrossRef]
- Reindl, W.; Baldo, B.; Schulz, J.; Janack, I.; Lindner, I.; Kleinschmidt, M.; Sedaghat, Y.; Thiede, C.; Tillack, K.; Schmidt, C.; et al. Meso Scale Discovery-Based Assays for the Detection of Aggregated Huntingtin. PLoS ONE 2019, 14, e0213521. [Google Scholar] [CrossRef]
- Caesar, I.; Jonson, M.; Nilsson, K.P.R.; Thor, S.; Hammarström, P. Curcumin Promotes A-Beta Fibrillation and Reduces Neurotoxicity in Transgenic Drosophila. PLoS ONE 2012, 7, e31424. [Google Scholar] [CrossRef]
- Jonson, M.; Pokrzywa, M.; Starkenberg, A.; Hammarstrom, P.; Thor, S. Systematic Aβ Analysis in Drosophila Reveals High Toxicity for the 1-42, 3-42 and 11-42 Peptides, and Emphasizes N- and C-Terminal Residues. PLoS ONE 2015, 10, e0133272. [Google Scholar] [CrossRef]
- Klingstedt, T.; Ghetti, B.; Holton, J.L.; Ling, H.; Nilsson, K.P.R.; Goedert, M. Luminescent Conjugated Oligothiophenes Distinguish between α-Synuclein Assemblies of Parkinson’s Disease and Multiple System Atrophy. Acta Neuropathol. Commun. 2019, 7, 193. [Google Scholar] [CrossRef]
- Siniukova, V.A.; Sopova, J.V.; Galkina, S.A.; Galkin, A.P. Search for Functional Amyloid Structures in Chicken and Fruit Fly Female Reproductive Cells. Prion 2020, 14, 278–282. [Google Scholar] [CrossRef]
- Greeve, I. Age-Dependent Neurodegeneration and Alzheimer-Amyloid Plaque Formation in Transgenic Drosophila. J. Neurosci. 2004, 24, 3899–3906. [Google Scholar] [CrossRef]
- Mariano, V.; Achsel, T.; Bagni, C.; Kanellopoulos, A.K. Modelling Learning and Memory in Drosophila to Understand Intellectual Disabilities. Neuroscience 2020, 445, 12–30. [Google Scholar] [CrossRef]
- Iijima, K.; Liu, H.-P.; Chiang, A.-S.; Hearn, S.A.; Konsolaki, M.; Zhong, Y. Dissecting the Pathological Effects of Human Aβ 40 and Aβ 42 in Drosophila: A Potential Model for Alzheimer’s Disease. Proc. Natl. Acad. Sci. USA 2004, 101, 6623–6628. [Google Scholar] [CrossRef]
- Vogel, J.W.; Iturria-Medina, Y.; Strandberg, O.T.; Smith, R.; Levitis, E.; Evans, A.C.; Hansson, O.; Alzheimer’s Disease Neuroimaging Initiative; Swedish BioFinder Study. Spread of Pathological Tau Proteins through Communicating Neurons in Human Alzheimer’s Disease. Nat. Commun. 2020, 11, 2612. [Google Scholar] [CrossRef]
- Fitzpatrick, A.W.P.; Falcon, B.; He, S.; Murzin, A.G.; Murshudov, G.; Garringer, H.J.; Crowther, R.A.; Ghetti, B.; Goedert, M.; Scheres, S.H.W. Cryo-EM Structures of Tau Filaments from Alzheimer’s Disease. Nature 2017, 547, 185–190. [Google Scholar] [CrossRef]
- Sardar Sinha, M.; Ansell-Schultz, A.; Civitelli, L.; Hildesjö, C.; Larsson, M.; Lannfelt, L.; Ingelsson, M.; Hallbeck, M. Alzheimer’s Disease Pathology Propagation by Exosomes Containing Toxic Amyloid-Beta Oligomers. Acta Neuropathol. 2018, 136, 41–56. [Google Scholar] [CrossRef]
- Chen, Y.-G. Research Progress in the Pathogenesis of Alzheimer’s Disease. Chin. Med. J. 2018, 131, 1618–1624. [Google Scholar] [CrossRef] [PubMed]
- Citron, M.; Teplow, D.B.; Selkoe, D.J. Generation of Amyloid Protein from Its Precursor Is Sequence Specific. Neuron 1995, 14, 661–670. [Google Scholar] [CrossRef]
- Fu, L.; Sun, Y.; Guo, Y.; Chen, Y.; Yu, B.; Zhang, H.; Wu, J.; Yu, X.; Kong, W.; Wu, H. Comparison of Neurotoxicity of Different Aggregated Forms of Aβ 40, Aβ 42 and Aβ 43 in Cell Cultures: Neurotoxicity of Different Amyloid-β Peptides. J. Pept. Sci. 2017, 23, 245–251. [Google Scholar] [CrossRef]
- Dahlgren, K.N.; Manelli, A.M.; Stine, W.B.; Baker, L.K.; Krafft, G.A.; LaDu, M.J. Oligomeric and Fibrillar Species of Amyloid-β Peptides Differentially Affect Neuronal Viability. J. Biol. Chem. 2002, 277, 32046–32053. [Google Scholar] [CrossRef] [PubMed]
- Finelli, A.; Kelkar, A.; Song, H.-J.; Yang, H.; Konsolaki, M. A Model for Studying Alzheimer’s Aβ42-Induced Toxicity in Drosophila melanogaster. Mol. Cell. Neurosci. 2004, 26, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Nilsberth, C.; Westlind-Danielsson, A.; Eckman, C.B.; Condron, M.M.; Axelman, K.; Forsell, C.; Stenh, C.; Luthman, J.; Teplow, D.B.; Younkin, S.G.; et al. The “Arctic” APP Mutation (E693G) Causes Alzheimer’s Disease by Enhanced Aβ Protofibril Formation. Nat. Neurosci. 2001, 4, 887–893. [Google Scholar] [CrossRef]
- Scholes, H.M.; Cryar, A.; Kerr, F.; Sutherland, D.; Gethings, L.A.; Vissers, J.P.C.; Lees, J.G.; Orengo, C.A.; Partridge, L.; Thalassinos, K. Dynamic Changes in the Brain Protein Interaction Network Correlates with Progression of Aβ42 Pathology in Drosophila. Sci. Rep. 2020, 10, 18517. [Google Scholar] [CrossRef] [PubMed]
- Sowade, R.F.; Jahn, T.R. Seed-Induced Acceleration of Amyloid-β Mediated Neurotoxicity in Vivo. Nat. Commun. 2017, 8, 512. [Google Scholar] [CrossRef] [PubMed]
- Speretta, E.; Jahn, T.R.; Tartaglia, G.G.; Favrin, G.; Barros, T.P.; Imarisio, S.; Lomas, D.A.; Luheshi, L.M.; Crowther, D.C.; Dobson, C.M. Expression in Drosophila of Tandem Amyloid β Peptides Provides Insights into Links between Aggregation and Neurotoxicity. J. Biol. Chem. 2012, 287, 20748–20754. [Google Scholar] [CrossRef]
- Burnouf, S.; Gorsky, M.K.; Dols, J.; Grönke, S.; Partridge, L. Aβ43 Is Neurotoxic and Primes Aggregation of Aβ40 in vivo. Acta Neuropathol. 2015, 130, 35–47. [Google Scholar] [CrossRef]
- Sofola-Adesakin, O.; Khericha, M.; Snoeren, I.; Tsuda, L.; Partridge, L. PGluAβ Increases Accumulation of Aβ in Vivo and Exacerbates Its Toxicity. Acta Neuropathol. Commun. 2016, 4, 109. [Google Scholar] [CrossRef]
- Vivien Chiu, W.Y.; Koon, A.C.; Ki Ngo, J.C.; Edwin Chan, H.Y.; Lau, K.-F. GULP1/CED-6 Ameliorates Amyloid-β Toxicity in a Drosophila Model of Alzheimer’s Disease. Oncotarget 2017, 8, 99274–99283. [Google Scholar] [CrossRef][Green Version]
- Chakraborty, R.; Vepuri, V.; Mhatre, S.D.; Paddock, B.E.; Miller, S.; Michelson, S.J.; Delvadia, R.; Desai, A.; Vinokur, M.; Melicharek, D.J.; et al. Characterization of a Drosophila Alzheimer’s Disease Model: Pharmacological Rescue of Cognitive Defects. PLoS ONE 2011, 6, e20799. [Google Scholar] [CrossRef]
- McKoy, A.F.; Chen, J.; Schupbach, T.; Hecht, M.H. A Novel Inhibitor of Amyloid β (Aβ) Peptide Aggregation. J. Biol. Chem. 2012, 287, 38992–39000. [Google Scholar] [CrossRef]
- McKoy, A.F.; Chen, J.; Schupbach, T.; Hecht, M.H. Structure-Activity Relationships for a Series of Compounds That Inhibit Aggregation of the Alzheimer’s Peptide, Aβ 42. Chem. Biol. Drug Des. 2014, 84, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Uras, G.; Manca, A.; Zhang, P.; Markus, Z.; Mack, N.; Allen, S.; Bo, M.; Xu, S.; Xu, J.; Georgiou, M.; et al. In Vivo Evaluation of a Newly Synthesized Acetylcholinesterase Inhibitor in a Transgenic Drosophila Model of Alzheimer’s Disease. Front. Neurosci. 2021, 15, 691222. [Google Scholar] [CrossRef] [PubMed]
- Akinyemi, A.J.; Oboh, G.; Ogunsuyi, O.; Abolaji, A.O.; Udofia, A. Curcumin-Supplemented Diets Improve Antioxidant Enzymes and Alter Acetylcholinesterase Genes Expression Level in Drosophila melanogaster Model. Metab. Brain Dis. 2018, 33, 369–375. [Google Scholar] [CrossRef]
- Luheshi, L.M.; Hoyer, W.; de Barros, T.P.; van Dijk Härd, I.; Brorsson, A.-C.; Macao, B.; Persson, C.; Crowther, D.C.; Lomas, D.A.; Ståhl, S.; et al. Sequestration of the Aβ Peptide Prevents Toxicity and Promotes Degradation In Vivo. PLoS Biol. 2010, 8, e1000334. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, H.; Okamoto, Y.; Motoi, A.; Watanabe, T.; Katayama, S.; Kawahara, S.; Makabe, H.; Fujii, H.; Yonekura, S. Adzuki Bean (Vigna angularis) Extract Reduces Amyloid-β Aggregation and Delays Cognitive Impairment in Drosophila Models of Alzheimer’s Disease. Nutr. Res. Pract. 2019, 13, 64. [Google Scholar] [CrossRef]
- Kruppa, A.J.; Ott, S.; Chandraratna, D.S.; Irving, J.A.; Page, R.M.; Speretta, E.; Seto, T.; Camargo, L.M.; Marciniak, S.J.; Lomas, D.A.; et al. Suppression of Aβ Toxicity by Puromycin-Sensitive Aminopeptidase Is Independent of Its Proteolytic Activity. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2013, 1832, 2115–2126. [Google Scholar] [CrossRef]
- Helmfors, L.; Boman, A.; Civitelli, L.; Nath, S.; Sandin, L.; Janefjord, C.; McCann, H.; Zetterberg, H.; Blennow, K.; Halliday, G.; et al. Protective Properties of Lysozyme on β-Amyloid Pathology: Implications for Alzheimer Disease. Neurobiol. Dis. 2015, 83, 122–133. [Google Scholar] [CrossRef]
- Sandin, L.; Bergkvist, L.; Nath, S.; Kielkopf, C.; Janefjord, C.; Helmfors, L.; Zetterberg, H.; Blennow, K.; Li, H.; Nilsberth, C.; et al. Beneficial Effects of Increased Lysozyme Levels in Alzheimer’s Disease Modelled in Drosophila melanogaster. FEBS J. 2016, 283, 3508–3522. [Google Scholar] [CrossRef]
- Hermansson, E.; Schultz, S.; Crowther, D.; Linse, S.; Winblad, B.; Westermark, G.; Johansson, J.; Presto, J. The Chaperone Domain BRICHOS Prevents Amyloid β-Peptide CNS Toxicity in Drosophila melanogaster. Dis. Model. Mech. 2014, 7, 659–665. [Google Scholar] [CrossRef]
- Poska, H.; Haslbeck, M.; Kurudenkandy, F.R.; Hermansson, E.; Chen, G.; Kostallas, G.; Abelein, A.; Biverstål, H.; Crux, S.; Fisahn, A.; et al. Dementia-Related Bri2 BRICHOS Is a Versatile Molecular Chaperone That Efficiently Inhibits Aβ42 Toxicity in Drosophila. Biochem. J. 2016, 473, 3683–3704. [Google Scholar] [CrossRef]
- Cohen, S.I.A.; Arosio, P.; Presto, J.; Kurudenkandy, F.R.; Biverstål, H.; Dolfe, L.; Dunning, C.; Yang, X.; Frohm, B.; Vendruscolo, M.; et al. A Molecular Chaperone Breaks the Catalytic Cycle That Generates Toxic Aβ Oligomers. Nat. Struct. Mol. Biol. 2015, 22, 207–213. [Google Scholar] [CrossRef]
- Kinney, J.W.; Bemiller, S.M.; Murtishaw, A.S.; Leisgang, A.M.; Salazar, A.M.; Lamb, B.T. Inflammation as a Central Mechanism in Alzheimer’s Disease. Alzheimers Dement. Transl. Res. Clin. Interv. 2018, 4, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, R.; Francioso, A.; d’Erme, M.; Trovato, M.; Mancini, P.; Piacentini, L.; Casale, A.; Wessjohann, L.; Gazzino, R.; Costantino, P.; et al. Anti-Inflammatory Activity of A Polyphenolic Extract from Arabidopsis Thaliana in In Vitro and In Vivo Models of Alzheimer’s Disease. Int. J. Mol. Sci. 2019, 20, 708. [Google Scholar] [CrossRef]
- Kong, Y.; Li, K.; Fu, T.; Wan, C.; Zhang, D.; Song, H.; Zhang, Y.; Liu, N.; Gan, Z.; Yuan, L. Quercetin Ameliorates Aβ Toxicity in Drosophila AD Model by Modulating Cell Cycle-Related Protein Expression. Oncotarget 2016, 7, 67716–67731. [Google Scholar] [CrossRef] [PubMed]
- Beg, T.; Jyoti, S.; Naz, F.; Rahul; Ali, F.; Ali, S.K.; Reyad, A.M.; Siddique, Y.H. Protective Effect of Kaempferol on the Transgenic Drosophila Model of Alzheimer’s Disease. CNS Neurol. Disord. Drug Targets 2018, 17, 421–429. [Google Scholar] [CrossRef]
- Ali, F.; Rahul; Jyoti, S.; Naz, F.; Ashafaq, M.; Shahid, M.; Siddique, Y.H. Therapeutic Potential of Luteolin in Transgenic Drosophila Model of Alzheimer’s Disease. Neurosci. Lett. 2019, 692, 90–99. [Google Scholar] [CrossRef]
- Ma, W.-W.; Tao, Y.; Wang, Y.-Y.; Peng, I.-F. Effects of Gardenia Jasminoides Extracts on Cognition and Innate Immune Response in an Adult Drosophila Model of Alzheimer’s Disease. Chin. J. Nat. Med. 2017, 15, 899–904. [Google Scholar] [CrossRef]
- Siddique, Y.H.; Ali, F. Protective Effect of Nordihydroguaiaretic Acid (NDGA) on the Transgenic Drosophila Model of Alzheimer’s Disease. Chem. Biol. Interact. 2017, 269, 59–66. [Google Scholar] [CrossRef]
- Lee, J.; Kim, Y.; Liu, T.; Hwang, Y.J.; Hyeon, S.J.; Im, H.; Lee, K.; Alvarez, V.E.; McKee, A.C.; Um, S.-J.; et al. SIRT3 Deregulation Is Linked to Mitochondrial Dysfunction in Alzheimer’s Disease. Aging Cell 2018, 17, e12679. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, P.F.; Hutter-Paier, B.; Havas, D.; Windisch, M.; Winblad, B. Development of GMP-1 a Molecular Chaperone Network Modulator Protecting Mitochondrial Function and Its Assessment in Fly and Mice Models of Alzheimer’s Disease. J. Cell. Mol. Med. 2018, 22, 3464–3474. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elovsson, G.; Bergkvist, L.; Brorsson, A.-C. Exploring Aβ Proteotoxicity and Therapeutic Candidates Using Drosophila melanogaster. Int. J. Mol. Sci. 2021, 22, 10448. https://doi.org/10.3390/ijms221910448
Elovsson G, Bergkvist L, Brorsson A-C. Exploring Aβ Proteotoxicity and Therapeutic Candidates Using Drosophila melanogaster. International Journal of Molecular Sciences. 2021; 22(19):10448. https://doi.org/10.3390/ijms221910448
Chicago/Turabian StyleElovsson, Greta, Liza Bergkvist, and Ann-Christin Brorsson. 2021. "Exploring Aβ Proteotoxicity and Therapeutic Candidates Using Drosophila melanogaster" International Journal of Molecular Sciences 22, no. 19: 10448. https://doi.org/10.3390/ijms221910448
APA StyleElovsson, G., Bergkvist, L., & Brorsson, A.-C. (2021). Exploring Aβ Proteotoxicity and Therapeutic Candidates Using Drosophila melanogaster. International Journal of Molecular Sciences, 22(19), 10448. https://doi.org/10.3390/ijms221910448