
Kinases of the Focal Adhesion Complex Contribute to Cardiomyocyte Specification


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
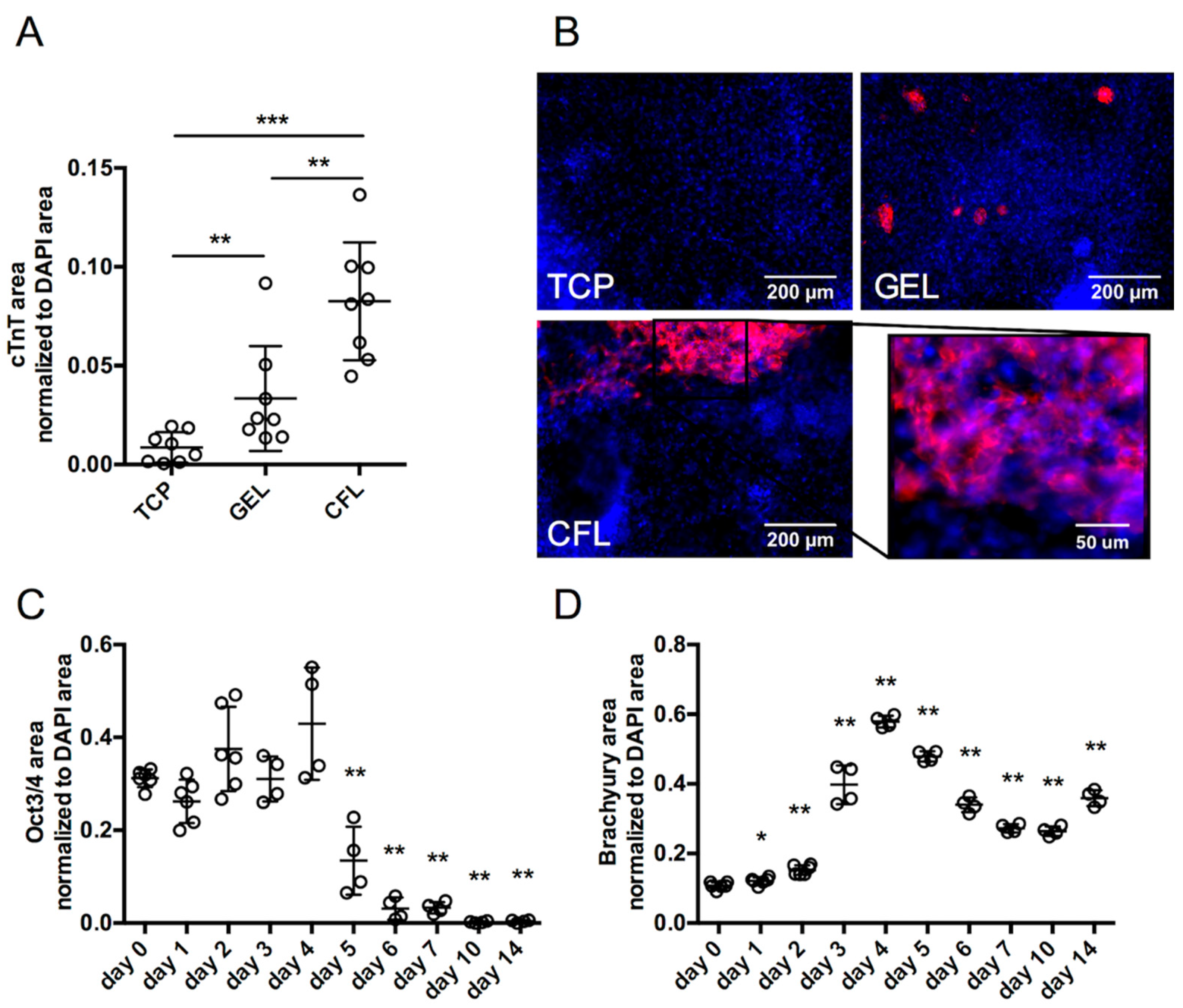
2.1. Engagement of Optimized ECM Formulation with miPSCs in 2D Induced Cardiomyocyte Differentiation
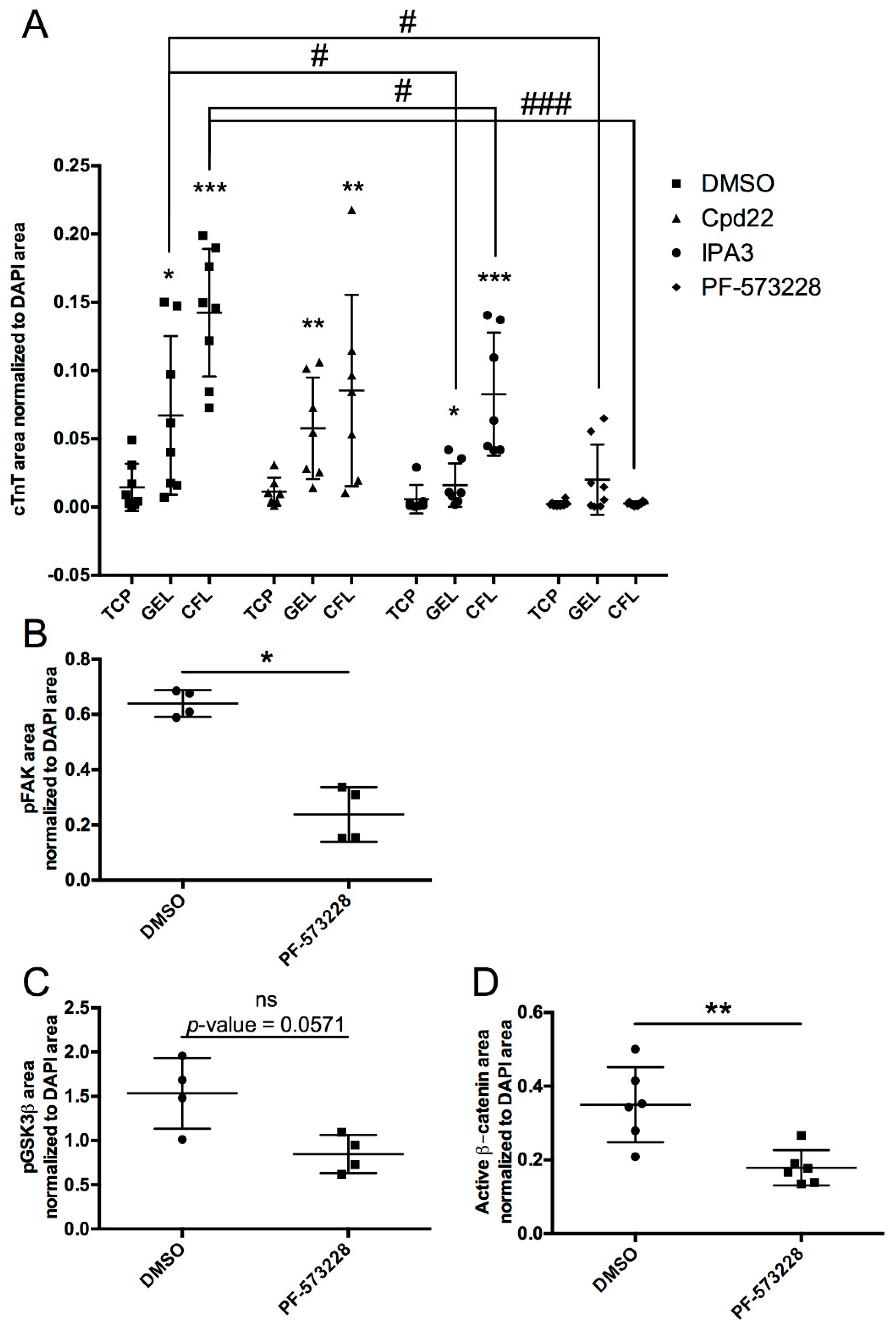
2.2. FAK and ILK Signaling Pathways Regulated Cardiomyocyte Differentiation of miPSCs upon ECM Engagement
2.3. ECM-Engagement with miPSC Included Characteristic Changes in ECM Composition during Cardiomyocyte Differentiation
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cardiomyocyte Differentiation upon ECM Engagement
4.3. Decellularization
4.4. Mass-Spectrometry Analysis
4.5. Immunofluorescence Staining
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BMP4 | bone morphogenetic protein 4 |
| CFL | collagen type I, fibronectin, laminin-111 |
| cTnT | cardiac troponin T |
| DI | deionized |
| DTT | dithiothreitol |
| ECM | extracellular matrix |
| EDTA | ethylenediaminetetraacetic acid |
| emPAI | exponentially modified protein abundance index |
| FAK | focal adhesion kinase |
| FBS | fetal bovine serum |
| GEL | gelatin |
| GMEM | Glasgow minimum essential medium |
| GSK3β | glycogen synthase kinase-3 beta |
| ILK | integrin-linked kinase |
| LC-MS2 | liquid chromatography with tandem mass spectrometry |
| LIF | leukemia inhibitory factor |
| miPSC | mouse induced pluripotent stem cell |
| PAK | p21 activated kinase |
| PBS | phosphate-buffered saline |
| SD | standard deviation |
| T | brachyury |
| TCP | tissue culture plastic |
References
- Go, A.S.; Mozaffarian, D.; Roger, V.L.; Benjamin, E.J.; Berry, J.D.; Blaha, M.J.; Dai, S.; Ford, E.S.; Fox, C.S.; Franco, S.; et al. Executive summary: Heart disease and stroke statistics—2014 update: A report from the American Heart Association. Circulation 2014, 129, 399–410. [Google Scholar] [CrossRef]
- Jung, J.P.; Hu, D.; Domian, I.J.; Ogle, B.M. An integrated statistical model for enhanced murine cardiomyocyte differentiation via optimized engagement of 3D extracellular matrices. Sci. Rep. 2015, 5, 18705. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Kupfer, M.E.; Jung, J.P.; Yang, L.; Zhang, P.; Da Sie, Y.; Tran, Q.; Ajeti, V.; Freeman, B.T.; Fast, V.G.; et al. Myocardial Tissue Engineering With Cells Derived From Human-Induced Pluripotent Stem Cells and a Native-Like, High-Resolution, 3-Dimensionally Printed Scaffold. Circ. Res. 2017, 120, 1318–1325. [Google Scholar] [CrossRef] [Green Version]
- Kupfer, M.E.; Lin, W.H.; Ravikumar, V.; Qiu, K.; Wang, L.; Gao, L.; Bhuiyan, D.B.; Lenz, M.; Ai, J.; Mahutga, R.R.; et al. In Situ Expansion, Differentiation, and Electromechanical Coupling of Human Cardiac Muscle in a 3D Bioprinted, Chambered Organoid. Circ. Res. 2020, 127, 207–224. [Google Scholar] [CrossRef]
- Finklea, F.B.; Tian, Y.; Kerscher, P.; Seeto, W.J.; Ellis, M.E.; Lipke, E.A. Engineered cardiac tissue microsphere production through direct differentiation of hydrogel-encapsulated human pluripotent stem cells. Biomaterials 2021, 274, 120818. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Luo, X.; Leighton, J. Extracellular Matrix and Integrins in Embryonic Stem Cell Differentiation. Biochem. Insights 2015, 8 (Suppl. S2), 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, K.H.; Hou, L.; Huang, N.F. Role of extracellular matrix signaling cues in modulating cell fate commitment for cardiovascular tissue engineering. Adv. Health Mater. 2014, 3, 628–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.P.; Bache-Wiig, M.K.; Provenzano, P.P.; Ogle, B.M. Heterogeneous Differentiation of Human Mesenchymal Stem Cells in 3D Extracellular Matrix Composites. Biores. Open Access 2016, 5, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Santiago, J.A.; Pogemiller, R.; Ogle, B.M. Heterogeneous differentiation of human mesenchymal stem cells in response to extended culture in extracellular matrices. Tissue Eng. Part A 2009, 15, 3911–3922. [Google Scholar] [CrossRef] [Green Version]
- Tan, T.W.; Huang, Y.L.; Chang, J.T.; Lin, J.J.; Fong, Y.C.; Kuo, C.C.; Tsai, C.H.; Chen, Y.J.; Hsu, H.C.; Cho, D.Y.; et al. CCN3 increases BMP-4 expression and bone mineralization in osteoblasts. J. Cell Physiol. 2012, 227, 2531–2541. [Google Scholar] [CrossRef]
- Cheng, P.; Andersen, P.; Hassel, D.; Kaynak, B.L.; Limphong, P.; Juergensen, L.; Kwon, C.; Srivastava, D. Fibronectin mediates mesendodermal cell fate decisions. Development 2013, 140, 2587–2596. [Google Scholar] [CrossRef] [Green Version]
- Lian, X.; Hsiao, C.; Wilson, G.; Zhu, K.; Hazeltine, L.B.; Azarin, S.M.; Raval, K.K.; Zhang, J.; Kamp, T.J.; Palecek, S.P. Robust cardiomyocyte differentiation from human pluripotent stem cells via temporal modulation of canonical Wnt signaling. Proc. Natl. Acad. Sci. USA 2012, 109, E1848–E1857. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Chen, G.; Kuan, S.F.; Zhang, D.H.; Schlaepfer, D.D.; Hu, J. FAK/PYK2 promotes the Wnt/beta-catenin pathway and intestinal tumorigenesis by phosphorylating GSK3beta. Elife 2015, 4, e10072. [Google Scholar] [CrossRef] [PubMed]
- Lindsley, R.C.; Gill, J.G.; Kyba, M.; Murphy, T.L.; Murphy, K.M. Canonical Wnt signaling is required for development of embryonic stem cell-derived mesoderm. Development 2006, 133, 3787–3796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gattazzo, F.; Urciuolo, A.; Bonaldo, P. Extracellular matrix: A dynamic microenvironment for stem cell niche. Biochim. Biophys Acta 2014, 1840, 2506–2519. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Tang, Y.; Zhou, Y.; Zhang, J. Deciphering Role of Wnt Signalling in Cardiac Mesoderm and Cardiomyocyte Differentiation from Human iPSCs: Four-dimensional control of Wnt pathway for hiPSC-CMs differentiation. Sci. Rep. 2019, 9, 19389. [Google Scholar] [CrossRef]
- Lian, X.; Zhang, J.; Azarin, S.M.; Zhu, K.; Hazeltine, L.B.; Bao, X.; Hsiao, C.; Kamp, T.J.; Palecek, S.P. Directed cardiomyocyte differentiation from human pluripotent stem cells by modulating Wnt/beta-catenin signaling under fully defined conditions. Nat. Protoc. 2013, 8, 162–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acconcia, F.; Barnes, C.J.; Singh, R.R.; Talukder, A.H.; Kumar, R. Phosphorylation-dependent regulation of nuclear localization and functions of integrin-linked kinase. Proc. Natl. Acad. Sci. USA 2007, 104, 6782–6787. [Google Scholar] [CrossRef] [Green Version]
- Bokoch, G.M. Biology of the p21-activated kinases. Annu. Rev. Biochem. 2003, 72, 743–781. [Google Scholar] [CrossRef]
- Dekkers, B.G.; Spanjer, A.I.; van der Schuyt, R.D.; Kuik, W.J.; Zaagsma, J.; Meurs, H. Focal adhesion kinase regulates collagen I-induced airway smooth muscle phenotype switching. J. Pharmacol. Exp. Ther. 2013, 346, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Seong, J.; Tajik, A.; Sun, J.; Guan, J.L.; Humphries, M.J.; Craig, S.E.; Shekaran, A.; Garcia, A.J.; Lu, S.; Lin, M.Z.; et al. Distinct biophysical mechanisms of focal adhesion kinase mechanoactivation by different extracellular matrix proteins. Proc. Natl. Acad. Sci. USA 2013, 110, 19372–19377. [Google Scholar] [CrossRef] [Green Version]
- Delcommenne, M.; Tan, C.; Gray, V.; Rue, L.; Woodgett, J.; Dedhar, S. Phosphoinositide-3-OH kinase-dependent regulation of glycogen synthase kinase 3 and protein kinase B/AKT by the integrin-linked kinase. Proc. Natl. Acad. Sci. USA 1998, 95, 11211–11216. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Wang, Y.; Huang, B.; Liang, J.; Ding, Y.; Xu, A.; Wu, W. A Rac1/PAK1 cascade controls beta-catenin activation in colon cancer cells. Oncogene 2012, 31, 1001–1012. [Google Scholar] [CrossRef] [Green Version]
- van Laake, L.W.; van Donselaar, E.G.; Monshouwer-Kloots, J.; Schreurs, C.; Passier, R.; Humbel, B.M.; Doevendans, P.A.; Sonnenberg, A.; Verkleij, A.J.; Mummery, C.L. Extracellular matrix formation after transplantation of human embryonic stem cell-derived cardiomyocytes. Cell Mol. Life Sci. 2010, 67, 277–290. [Google Scholar] [CrossRef] [Green Version]
- Sasse, P.; Malan, D.; Fleischmann, M.; Roell, W.; Gustafsson, E.; Bostani, T.; Fan, Y.; Kolbe, T.; Breitbach, M.; Addicks, K.; et al. Perlecan is critical for heart stability. Cardiovasc. Res. 2008, 80, 435–444. [Google Scholar] [CrossRef] [Green Version]
- Hopf, M.; Gohring, W.; Kohfeldt, E.; Yamada, Y.; Timpl, R. Recombinant domain IV of perlecan binds to nidogens, laminin-nidogen complex, fibronectin, fibulin-2 and heparin. Eur. J. Biochem. 1999, 259, 917–925. [Google Scholar] [CrossRef]
- Beetz, N.; Rommel, C.; Schnick, T.; Neumann, E.; Lother, A.; Monroy-Ordonez, E.B.; Zeeb, M.; Preissl, S.; Gilsbach, R.; Melchior-Becker, A.; et al. Ablation of biglycan attenuates cardiac hypertrophy and fibrosis after left ventricular pressure overload. J. Mol. Cell Cardiol. 2016, 101, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Scilabra, S.D.; Pigoni, M.; Pravata, V.; Schatzl, T.; Muller, S.A.; Troeberg, L.; Lichtenthaler, S.F. Increased TIMP-3 expression alters the cellular secretome through dual inhibition of the metalloprotease ADAM10 and ligand-binding of the LRP-1 receptor. Sci. Rep. 2018, 8, 14697. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, A.D.; Schulz, R. Glycogen synthase kinase-3beta is activated by matrix metalloproteinase-2 mediated proteolysis in cardiomyoblasts. Cardiovasc. Res. 2009, 83, 698–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, Z.; Sharma, D.; Jia, W.; Radke, D.; Kamp, T.; Zhao, F. Engineering stem cell cardiac patch with microvascular features representative of native myocardium. Theranostics 2019, 9, 2143–2157. [Google Scholar] [CrossRef]
- Van Laake, L.W.; Qian, L.; Cheng, P.; Huang, Y.; Hsiao, E.C.; Conklin, B.R.; Srivastava, D. Reporter-based isolation of induced pluripotent stem cell- and embryonic stem cell-derived cardiac progenitors reveals limited gene expression variance. Circ. Res. 2010, 107, 340–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robert, S.; Flowers, M.; Ogle, B.M. Kinases of the Focal Adhesion Complex Contribute to Cardiomyocyte Specification. Int. J. Mol. Sci. 2021, 22, 10430. https://doi.org/10.3390/ijms221910430
Robert S, Flowers M, Ogle BM. Kinases of the Focal Adhesion Complex Contribute to Cardiomyocyte Specification. International Journal of Molecular Sciences. 2021; 22(19):10430. https://doi.org/10.3390/ijms221910430
Chicago/Turabian StyleRobert, Sacha, Marcus Flowers, and Brenda M. Ogle. 2021. "Kinases of the Focal Adhesion Complex Contribute to Cardiomyocyte Specification" International Journal of Molecular Sciences 22, no. 19: 10430. https://doi.org/10.3390/ijms221910430
APA StyleRobert, S., Flowers, M., & Ogle, B. M. (2021). Kinases of the Focal Adhesion Complex Contribute to Cardiomyocyte Specification. International Journal of Molecular Sciences, 22(19), 10430. https://doi.org/10.3390/ijms221910430