
Smoothelin-Like Protein 1 Regulates the Thyroid Hormone-Induced Homeostasis and Remodeling of C2C12 Cells via the Modulation of Myosin Phosphatase


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
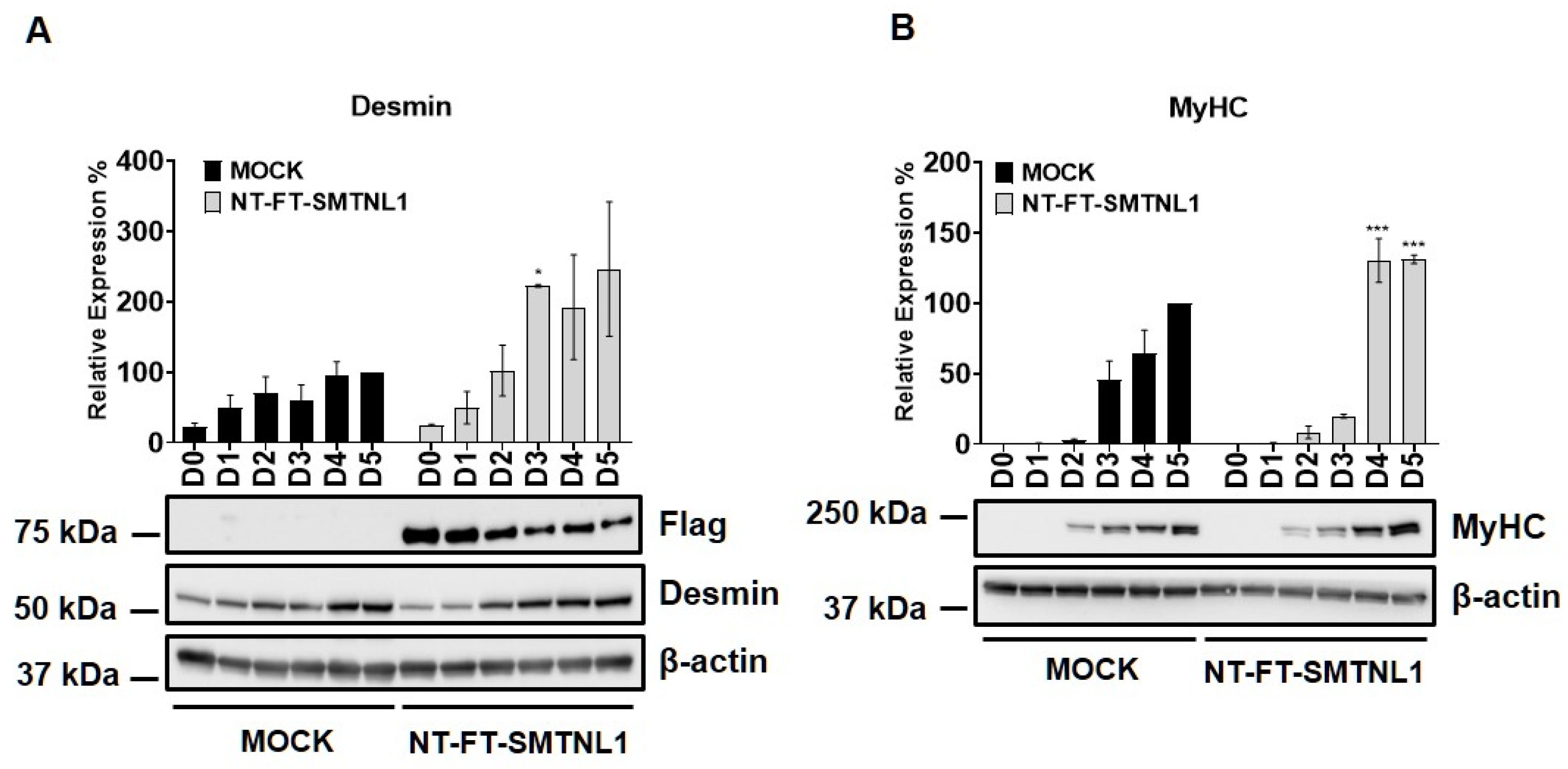
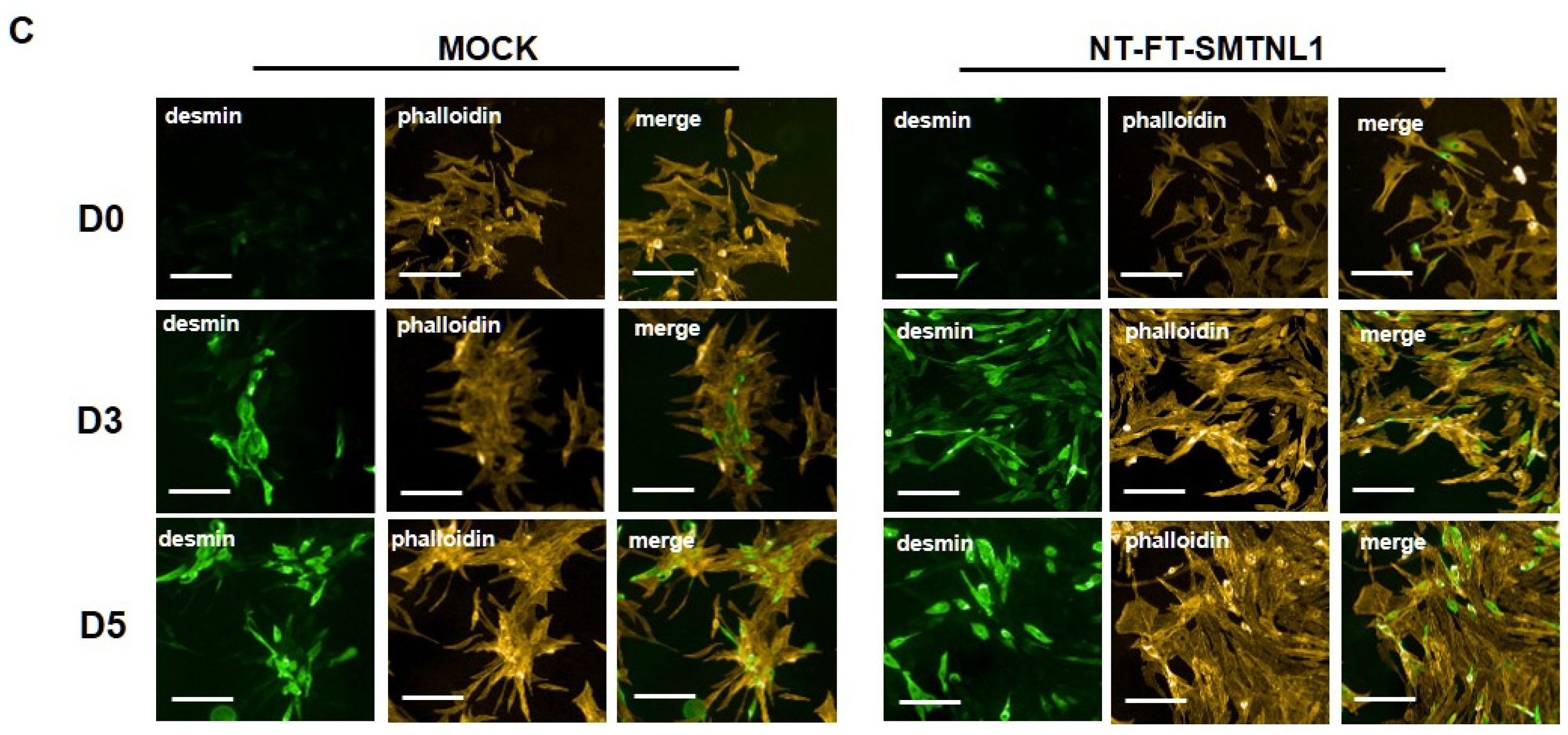
2.1. Overexpression of SMTNL1 Promotes C2C12 Mouse Myoblast Differentiation
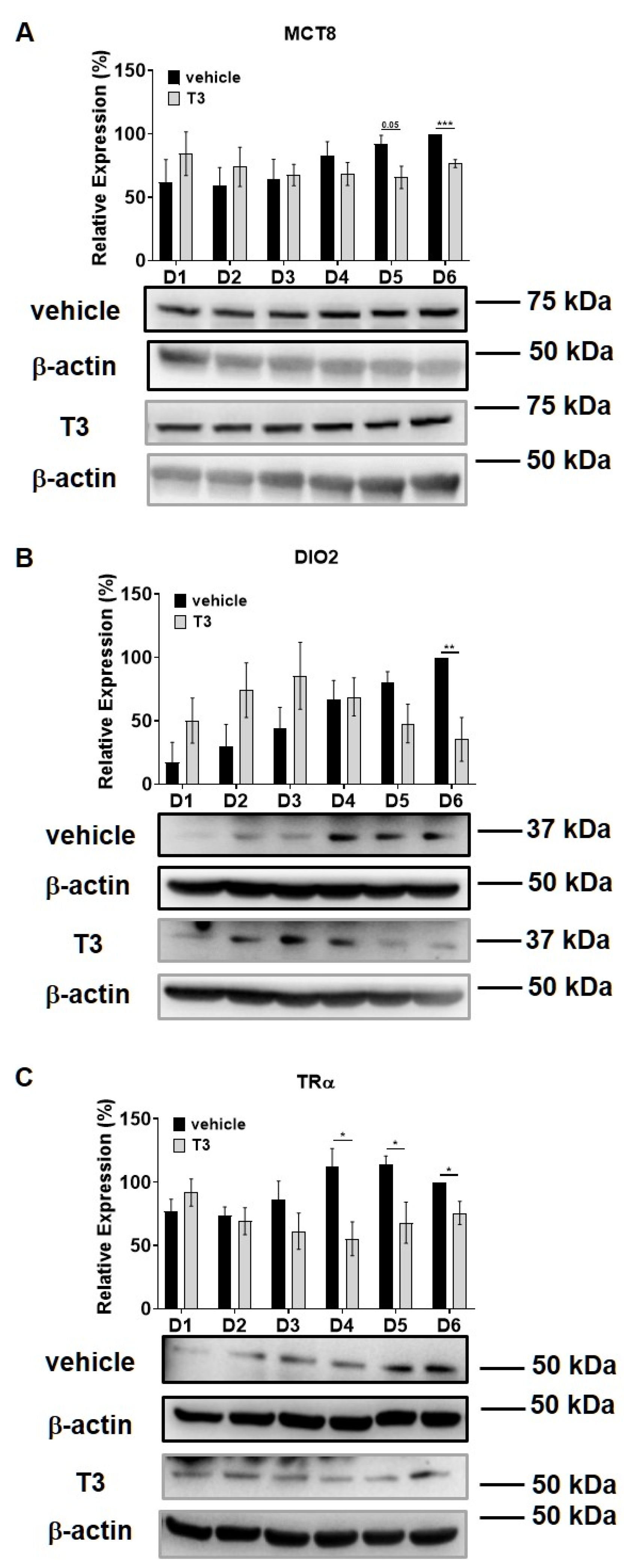
2.2. Myosin Phosphatase Expression during C2C12 Myogenesis in the Presence of Excess T3
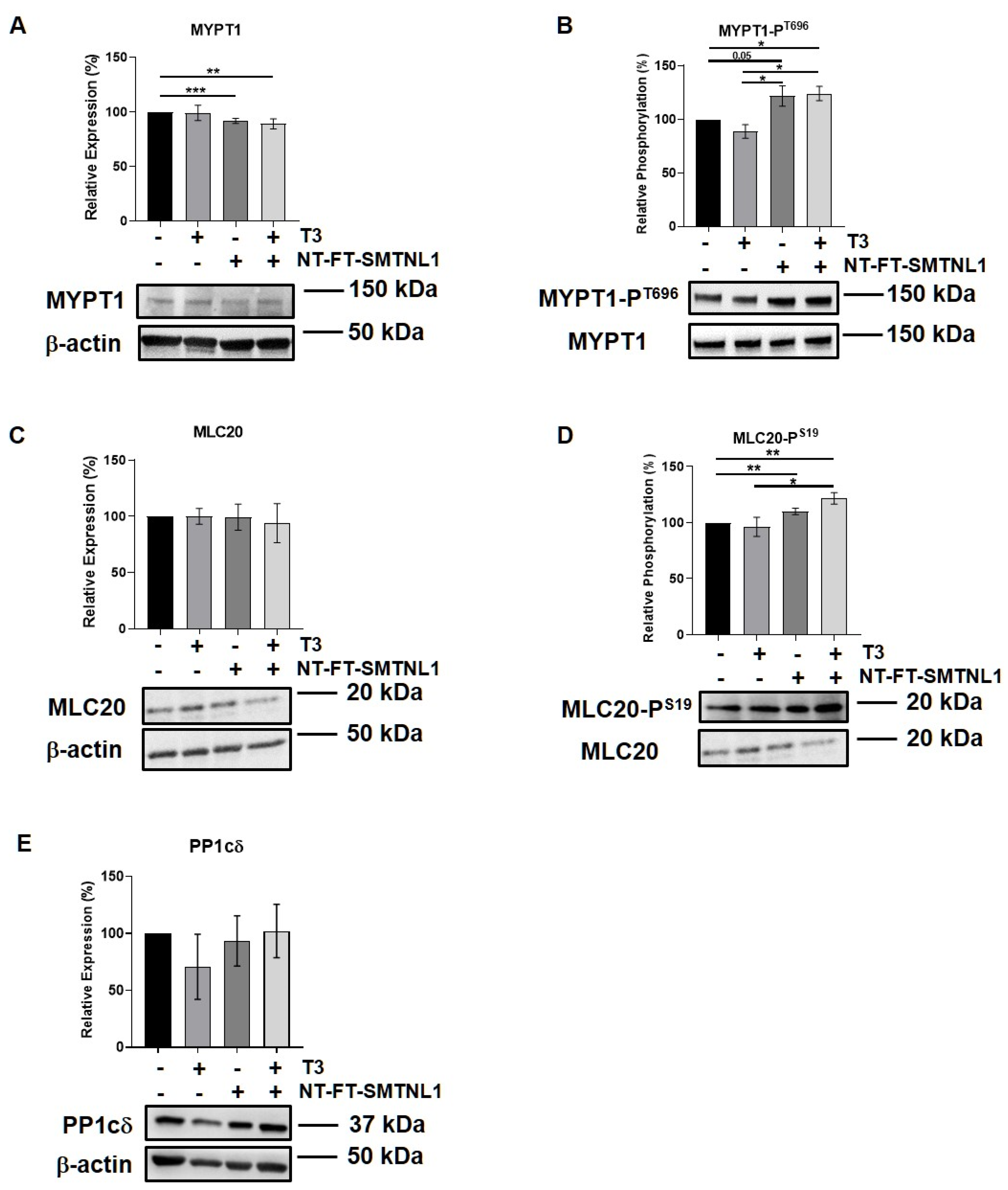
2.3. SMTNL1 Overexpression Induces the Phosphorylation of MYPT1 and MLC20 in Myoblasts
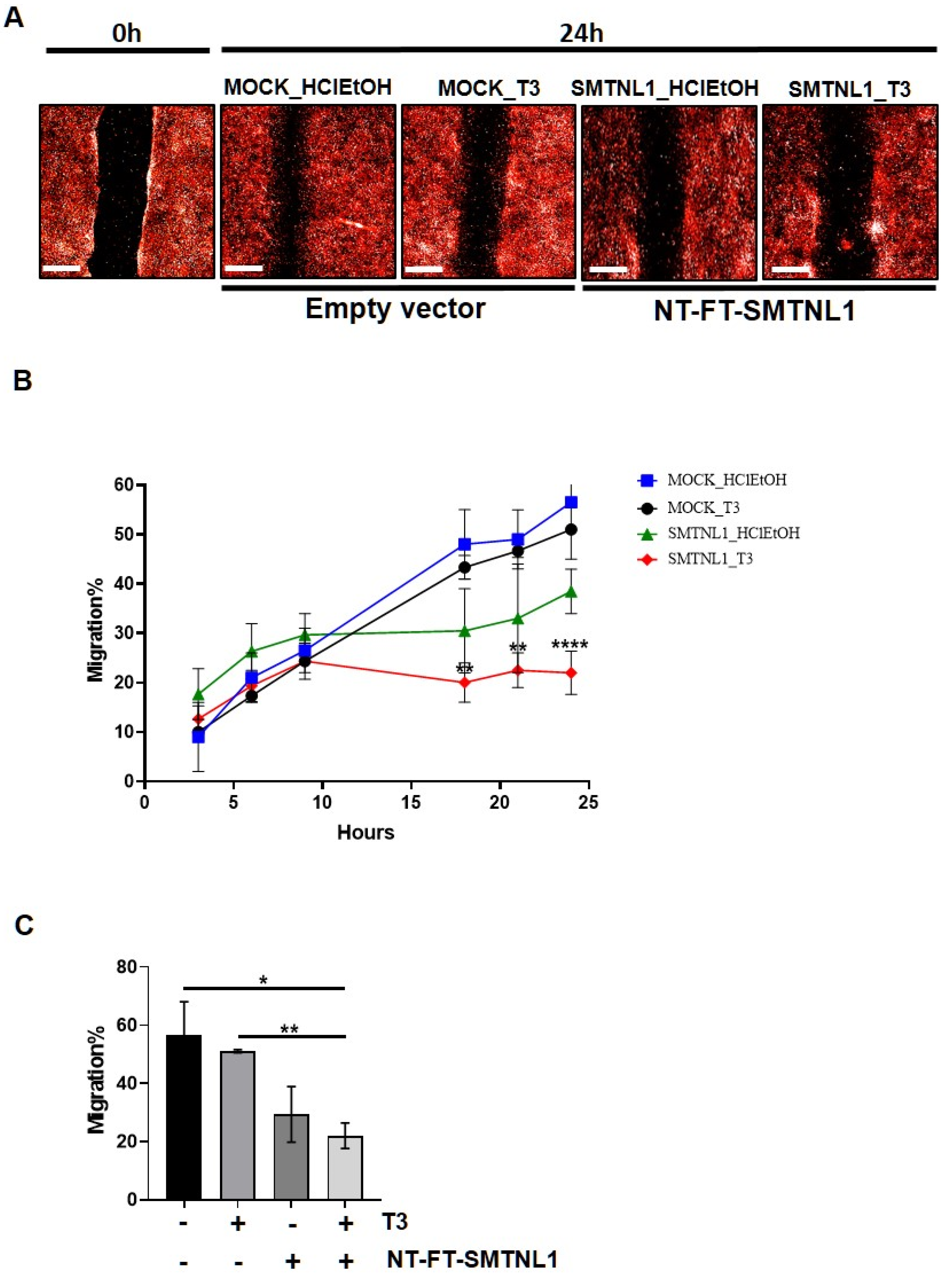
2.4. SMTNL1 Overexpression Inhibits the Migration of Myoblasts
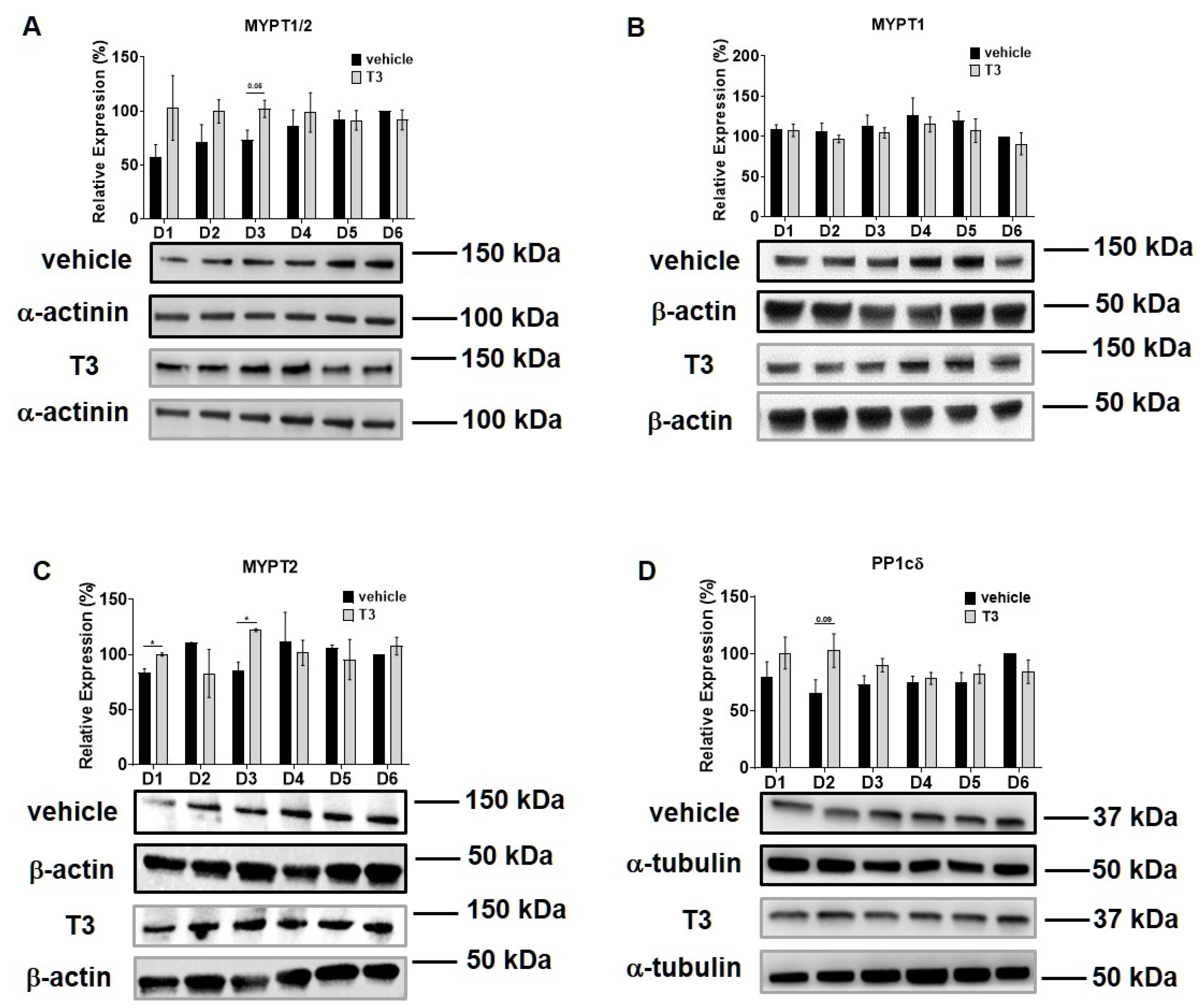
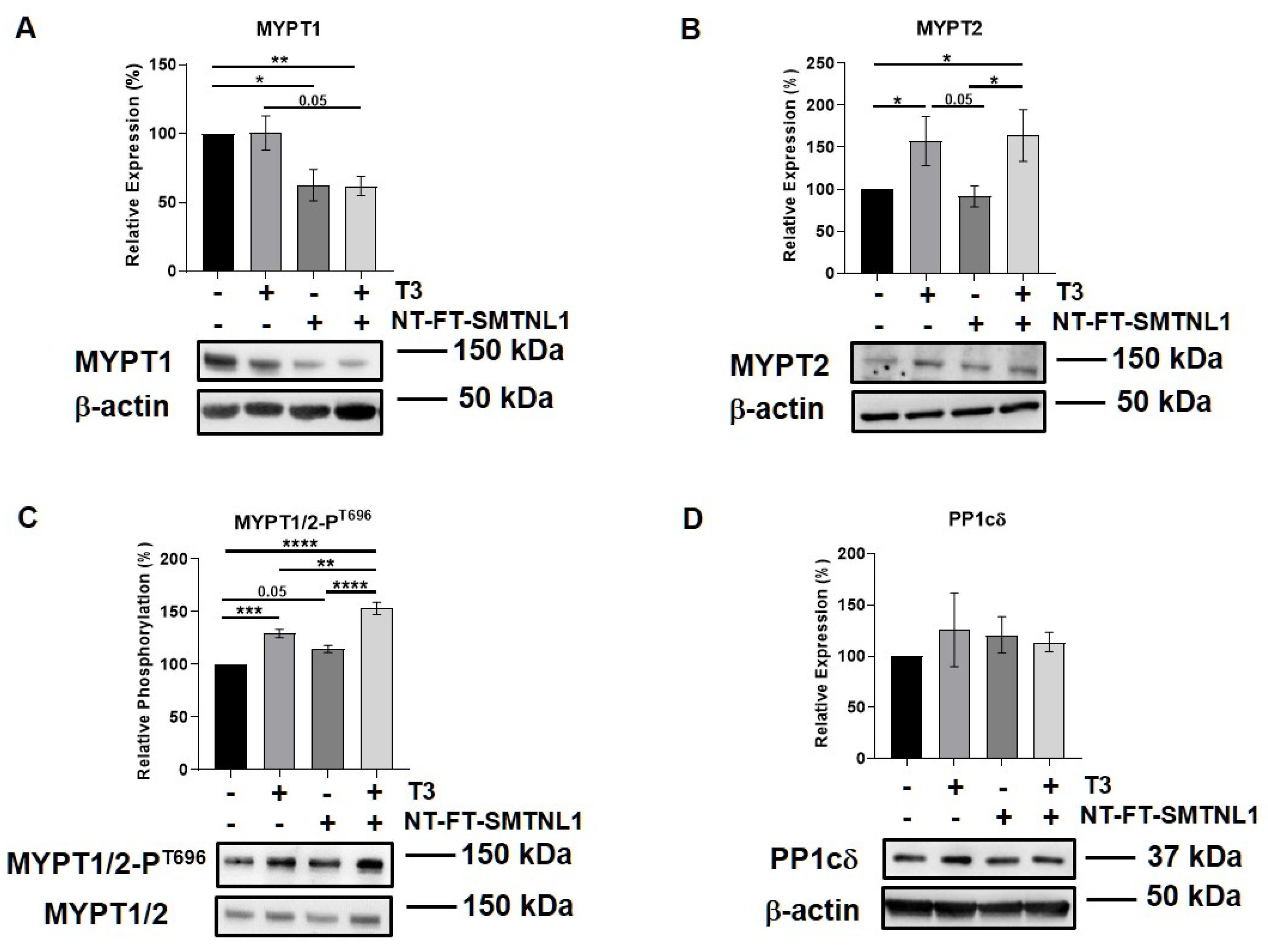
2.5. Differential Regulation of Myosin Phosphatase Target Subunit Isoforms by T3 Treatment and SMTNL1 Overexpression in Myotubes
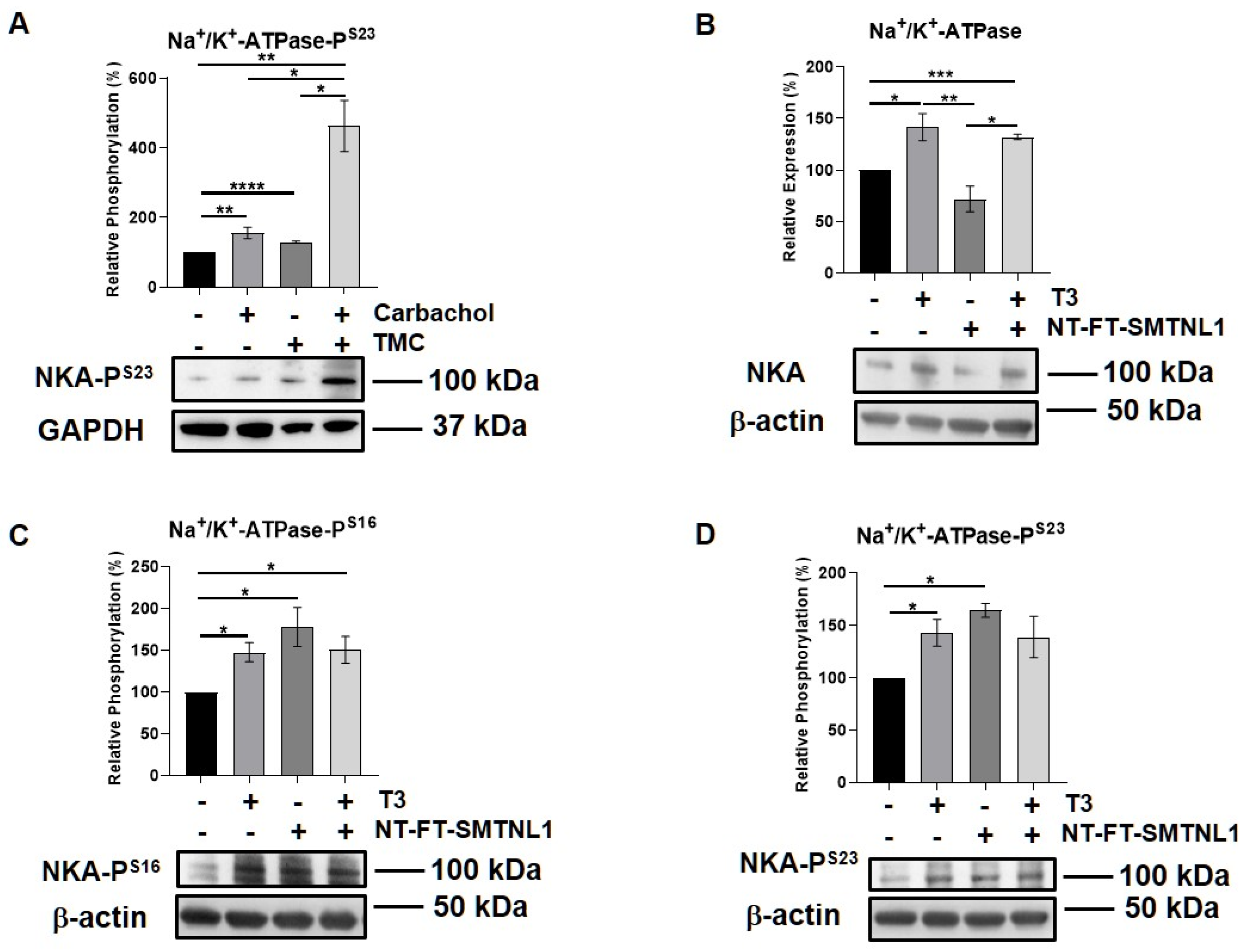
2.6. Effect of T3 Treatment and/or SMTNL1 Overexpression on the Expression and Activity of the Na+/K+-ATPase, an Alternative MP Substrate in Myotubes
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Antibodies
4.3. Cell Culture Maintenance
4.4. Thin Coating of Tissue Culture Plates
4.5. Transient Transfection and Differentiation
4.6. Immunofluorescence Staining
4.7. 3,3′,5-Triiodo-L-thyronine (T3) Hormone Treatment
4.8. Cell Lysis
4.9. Western Blot Analysis
4.10. Scratch Assay and High Content Screening (HCS) Analysis
4.11. Carbachol and Tautomycetin Treatment
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vale, C.; Neves, J.S.; von Hafe, M.; Borges-Canha, M.; Leite-Moreira, A. The Role of Thyroid Hormones in Heart Failure. Cardiovasc. Drugs Ther. 2019, 33, 179–188. [Google Scholar] [CrossRef]
- Goncalves, A.; Resende, E.S.; Fernandes, M.L.; da Costa, A.M. Effect of thyroid hormones on cardiovascular and muscle systems and on exercise tolerance: A brief review. Arq. Bras. Cardiol. 2006, 87, e45–e47. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bloise, F.F.; Cordeiro, A.; Ortiga-Carvalho, T.M. Role of thyroid hormone in skeletal muscle physiology. J. Endocrinol. 2018, 236, R57–R68. [Google Scholar] [CrossRef]
- Friesema, E.C.; Jansen, J.; Visser, T.J. Thyroid hormone transporters. Biochem. Soc. Trans. 2005, 33, 228–232. [Google Scholar] [CrossRef]
- Bianco, A.C.; Kim, B.W. Deiodinases: Implications of the local control of thyroid hormone action. J. Clin. Investig. 2006, 116, 2571–2579. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.J.; Goglia, F.; Leonard, J.L. Nongenomic actions of thyroid hormone. Nat. Rev. Endocrinol. 2016, 12, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Cakir, M.; Samanci, N.; Balci, N.; Balci, M.K. Musculoskeletal manifestations in patients with thyroid disease. Clin. Endocrinol. 2003, 59, 162–167. [Google Scholar] [CrossRef]
- Olson, B.R.; Klein, I.; Benner, R.; Burdett, R.; Trzepacz, P.; Levey, G.S. Hyperthyroid myopathy and the response to treatment. Thyroid 1991, 1, 137–141. [Google Scholar] [CrossRef]
- Short, K.R.; Nygren, J.; Barazzoni, R.; Levine, J.; Nair, K.S. T3 increases mitochondrial ATP production in oxidative muscle despite increased expression of UCP2 and -3. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E761–E769. [Google Scholar] [CrossRef]
- Chacko, S.; Conti, M.A.; Adelstein, R.S. Effect of phosphorylation of smooth muscle myosin on actin activation and Ca2+ regulation. Proc. Natl. Acad. Sci. USA 1977, 74, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Ikebe, M.; Hartshorne, D.J. The role of myosin phosphorylation in the contraction-relaxation cycle of smooth muscle. Experientia 1985, 41, 1006–1010. [Google Scholar] [CrossRef] [PubMed]
- Kiss, A.; Erdodi, F.; Lontay, B. Myosin phosphatase: Unexpected functions of a long-known enzyme. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 2–15. [Google Scholar] [CrossRef]
- Shimizu, H.; Ito, M.; Miyahara, M.; Ichikawa, K.; Okubo, S.; Konishi, T.; Naka, M.; Tanaka, T.; Hirano, K.; Hartshorne, D.J.; et al. Characterization of the myosin-binding subunit of smooth muscle myosin phosphatase. J. Biol. Chem. 1994, 269, 30407–30411. [Google Scholar] [CrossRef]
- Fujioka, M.; Takahashi, N.; Odai, H.; Araki, S.; Ichikawa, K.; Feng, J.; Nakamura, M.; Kaibuchi, K.; Hartshorne, D.J.; Nakano, T.; et al. A new isoform of human myosin phosphatase targeting/regulatory subunit (MYPT2): cDNA cloning, tissue expression, and chromosomal mapping. Genomics 1998, 49, 59–68. [Google Scholar] [CrossRef]
- Skinner, J.A.; Saltiel, A.R. Cloning and identification of MYPT3: A prenylatable myosin targetting subunit of protein phosphatase 1. Biochem. J. 2001, 356 Pt 1, 257–267. [Google Scholar] [CrossRef]
- Tan, I.; Ng, C.H.; Lim, L.; Leung, T. Phosphorylation of a novel myosin binding subunit of protein phosphatase 1 reveals a conserved mechanism in the regulation of actin cytoskeleton. J. Biol. Chem. 2001, 276, 21209–21216. [Google Scholar] [CrossRef]
- Cao, W.S.; Mattagajasingh, S.N.; Xu, H.X.; Kim, K.H.; Fierlbeck, W.; Deng, J.; Lowenstein, C.J.; Ballermann, B.J. TIMAP, a novel CAAX box protein regulated by TGF-beta 1 and expressed in endothelial cells. Am. J. Physiol.-Cell Physiol. 2002, 283, C327–C337. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, F.; Hartshorne, D.J. Myosin phosphatase target subunit: Many roles in cell function. Biochem. Biophys. Res. Commun. 2008, 369, 149–156. [Google Scholar] [CrossRef]
- Arimura, T.; Suematsu, N.; Zhou, Y.B.; Nishimura, J.; Satoh, S.; Takeshita, A.; Kanaide, H.; Kimura, A. Identification, characterization, and functional analysis of heart-specific myosin light chain phosphatase small subunit. J. Biol. Chem. 2001, 276, 6073–6082. [Google Scholar] [CrossRef]
- Ichikawa, K.; Ito, M.; Hartshorne, D.J. Phosphorylation of the large subunit of myosin phosphatase and inhibition of phosphatase activity. J. Biol. Chem. 1996, 271, 4733–4740. [Google Scholar] [CrossRef]
- Muranyi, A.; Derkach, D.; Erdodi, F.; Kiss, A.; Ito, M.; Hartshorne, D.J. Phosphorylation of Thr695 and Thr850 on the myosin phosphatase target subunit: Inhibitory effects and occurrence in A7r5 cells. FEBS Lett. 2005, 579, 6611–6615. [Google Scholar] [CrossRef]
- Velasco, G.; Armstrong, C.; Morrice, N.; Frame, S.; Cohen, P. Phosphorylation of the regulatory subunit of smooth muscle protein phosphatase 1M at Thr850 induces its dissociation from myosin. FEBS Lett. 2002, 527, 101–104. [Google Scholar] [CrossRef]
- Borman, M.A.; MacDonald, J.A.; Haystead, T.A. Modulation of smooth muscle contractility by CHASM, a novel member of the smoothelin family of proteins. FEBS Lett. 2004, 573, 207–213. [Google Scholar] [CrossRef]
- Wooldridge, A.A.; Fortner, C.N.; Lontay, B.; Akimoto, T.; Neppl, R.L.; Facemire, C.; Datto, M.B.; Kwon, A.; McCook, E.; Li, P.; et al. Deletion of the protein kinase A/protein kinase G target SMTNL1 promotes an exercise-adapted phenotype in vascular smooth muscle. J. Biol. Chem. 2008, 283, 11850–11859. [Google Scholar] [CrossRef] [PubMed]
- Hartshorne, D.J.; Ito, M.; Erdodi, F. Role of protein phosphatase type 1 in contractile functions: Myosin phosphatase. J. Biol. Chem. 2004, 279, 37211–37214. [Google Scholar] [CrossRef] [PubMed]
- Lontay, B.; Bodoor, K.; Weitzel, D.H.; Loiselle, D.; Fortner, C.; Lengyel, S.; Zheng, D.; Devente, J.; Hickner, R.; Haystead, T.A. Smoothelin-like 1 protein regulates myosin phosphatase-targeting subunit 1 expression during sexual development and pregnancy. J. Biol. Chem. 2010, 285, 29357–29366. [Google Scholar] [CrossRef] [PubMed]
- Major, E.; Győry, F.; Horváth, D.; Keller, I.; Tamás, I.; Uray, K.; Fülöp, P.; Lontay, B. Smoothelin-like protein 1 regulates development and metabolic transformation of skeletal muscle in hyperthyroidism. Front. Endocrinol 2021, in press. [Google Scholar]
- Lontay, B.; Bodoor, K.; Sipos, A.; Weitzel, D.H.; Loiselle, D.; Safi, R.; Zheng, D.; Devente, J.; Hickner, R.C.; McDonnell, D.P.; et al. Pregnancy and Smoothelin-like Protein 1 (SMTNL1) Deletion Promote the Switching of Skeletal Muscle to a Glycolytic Phenotype in Human and Mice. J. Biol. Chem. 2015, 290, 17985–17998. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Choudhary, S.K.; Milner, D.J.; Munir, M.I.; Kuisk, I.R.; Capetanaki, Y. Inhibition of desmin expression blocks myoblast fusion and interferes with the myogenic regulators MyoD and myogenin. J. Cell Biol. 1994, 124, 827–841. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Chen, H.; Guo, C.; Yuan, Z.; Zhou, X.; Zhang, Y.; Zhang, X.; Mo, D.; Chen, Y. Palmdelphin promotes myoblast differentiation and muscle regeneration. Sci. Rep. 2017, 7, 41608. [Google Scholar] [CrossRef]
- Dentice, M.; Marsili, A.; Ambrosio, R.; Guardiola, O.; Sibilio, A.; Paik, J.H.; Minchiotti, G.; DePinho, R.A.; Fenzi, G.; Larsen, P.R.; et al. The FoxO3/type 2 deiodinase pathway is required for normal mouse myogenesis and muscle regeneration. J. Clin. Investig. 2010, 120, 4021–4030. [Google Scholar] [CrossRef]
- Milanesi, A.; Lee, J.W.; Kim, N.H.; Liu, Y.Y.; Yang, A.; Sedrakyan, S.; Kahng, A.; Cervantes, V.; Tripuraneni, N.; Cheng, S.Y.; et al. Thyroid Hormone Receptor alpha Plays an Essential Role in Male Skeletal Muscle Myoblast Proliferation, Differentiation, and Response to Injury. Endocrinology 2016, 157, 4–15. [Google Scholar] [CrossRef]
- Wu, Y.; Erdodi, F.; Muranyi, A.; Nullmeyer, K.D.; Lynch, R.M.; Hartshorne, D.J. Myosin phosphatase and myosin phosphorylation in differentiating C2C12 cells. J. Muscle Res. Cell Motil. 2003, 24, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Lontay, B.; Kiss, A.; Gergely, P.; Hartshorne, D.J.; Erdodi, F. Okadaic acid induces phosphorylation and translocation of myosin phosphatase target subunit 1 influencing myosin phosphorylation, stress fiber assembly and cell migration in HepG2 cells. Cell Signal. 2005, 17, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Lehka, L.; Redowicz, M.J. Mechanisms regulating myoblast fusion: A multilevel interplay. Semin. Cell Dev. Biol. 2020, 104, 81–92. [Google Scholar] [CrossRef]
- Lontay, B.; Pal, B.; Serfozo, Z.; Koszeghy, A.; Szucs, G.; Rusznak, Z.; Erdodi, F. Protein phosphatase-1M and Rho-kinase affect exocytosis from cortical synaptosomes and influence neurotransmission at a glutamatergic giant synapse of the rat auditory system. J. Neurochem. 2012, 123, 84–99. [Google Scholar] [CrossRef]
- Pirkmajer, S.; Chibalin, A.V. Na,K-ATPase regulation in skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E1–E31. [Google Scholar] [CrossRef]
- Riis, A.L.; Jorgensen, J.O.; Moller, N.; Weeke, J.; Clausen, T. Hyperthyroidism and cation pumps in human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E1265–E1269. [Google Scholar] [CrossRef]
- Soltoff, S.P.; Asara, J.M.; Hedden, L. Regulation and identification of Na,K-ATPase alpha1 subunit phosphorylation in rat parotid acinar cells. J. Biol. Chem. 2010, 285, 36330–36338. [Google Scholar] [CrossRef]
- Ambrosio, R.; De Stefano, M.A.; Di Girolamo, D.; Salvatore, D. Thyroid hormone signaling and deiodinase actions in muscle stem/progenitor cells. Mol. Cell Endocrinol. 2017, 459, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Dentice, M.; Ambrosio, R.; Damiano, V.; Sibilio, A.; Luongo, C.; Guardiola, O.; Yennek, S.; Zordan, P.; Minchiotti, G.; Colao, A.; et al. Intracellular inactivation of thyroid hormone is a survival mechanism for muscle stem cell proliferation and lineage progression. Cell Metab. 2014, 20, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Di Cosmo, C.; Liao, X.H.; Ye, H.; Ferrara, A.M.; Weiss, R.E.; Refetoff, S.; Dumitrescu, A.M. Mct8-deficient mice have increased energy expenditure and reduced fat mass that is abrogated by normalization of serum T3 levels. Endocrinology 2013, 154, 4885–4895. [Google Scholar] [CrossRef] [PubMed]
- Milanesi, A.; Lee, J.W.; Yang, A.; Liu, Y.Y.; Sedrakyan, S.; Cheng, S.Y.; Perin, L.; Brent, G.A. Thyroid Hormone Receptor Alpha is Essential to Maintain the Satellite Cell Niche During Skeletal Muscle Injury and Sarcopenia of Aging. Thyroid 2017, 27, 1316–1322. [Google Scholar] [CrossRef] [PubMed]
- Lontay, B.; Serfozo, Z.; Gergely, P.; Ito, M.; Hartshorne, D.J.; Erdodi, F. Localization of myosin phosphatase target subunit 1 in rat brain and in primary cultures of neuronal cells. J. Comp. Neurol. 2004, 478, 72–87. [Google Scholar] [CrossRef]
- Brown, D.M.; Parr, T.; Brameld, J.M. Myosin heavy chain mRNA isoforms are expressed in two distinct cohorts during C2C12 myogenesis. J. Muscle Res. Cell Motil. 2012, 32, 383–390. [Google Scholar] [CrossRef]
- Izumo, S.; Nadal-Ginard, B.; Mahdavi, V. All members of the MHC multigene family respond to thyroid hormone in a highly tissue-specific manner. Science 1986, 231, 597–600. [Google Scholar] [CrossRef]
- Vicente-Manzanares, M.; Horwitz, A.R. Myosin light chain mono- and di-phosphorylation differentially regulate adhesion and polarity in migrating cells. Biochem. Biophys. Res. Commun. 2010, 402, 537–542. [Google Scholar] [CrossRef]
- Tullio, A.N.; Accili, D.; Ferrans, V.J.; Yu, Z.X.; Takeda, K.; Grinberg, A.; Westphal, H.; Preston, Y.A.; Adelstein, R.S. Nonmuscle myosin II-B is required for normal development of the mouse heart. Proc. Natl. Acad. Sci. USA 1997, 94, 12407–12412. [Google Scholar] [CrossRef]
- Zvibel, I.; Atias, D.; Phillips, A.; Halpern, Z.; Oren, R. Thyroid hormones induce activation of rat hepatic stellate cells through increased expression of p75 neurotrophin receptor and direct activation of Rho. Lab. Investig. 2010, 90, 674–684. [Google Scholar] [CrossRef]
- Kondo, A.; Tokuda, H.; Kato, K.; Matsushima-Nishiwaki, R.; Kuroyanagi, G.; Mizutani, J.; Kozawa, O.; Otsuka, T. Rho-kinase negatively regulates thyroid hormone-stimulated osteocalcin synthesis in osteoblasts. Biochimie 2013, 95, 719–724. [Google Scholar] [CrossRef]
- Pajouhi, N.; Owji, M.; Naghibalhossaini, F.; Omrani, G.H.; Varedi, M. Modulation by thyroid hormone of myosin light chain phosphorylation and aquaporin 5 protein expression in intact lung. J. Physiol. Biochem. 2015, 71, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Horvath, D.; Sipos, A.; Major, E.; Konya, Z.; Batori, R.; Dedinszki, D.; Szoll Si, A.; Tamas, I.; Ivan, J.; Kiss, A.; et al. Myosin phosphatase accelerates cutaneous wound healing by regulating migration and differentiation of epidermal keratinocytes via Akt signaling pathway in human and murine skin. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3268–3280. [Google Scholar] [CrossRef]
- Stull, J.T.; Kamm, K.E.; Taylor, D.A. Calcium control of smooth muscle contractility. Am. J. Med. Sci. 1988, 296, 241–245. [Google Scholar] [CrossRef]
- Ragolia, L.; Cherpalis, B.; Srinivasan, M.; Begum, N. Role of serine/threonine protein phosphatases in insulin regulation of Na+/K+-ATPase activity in cultured rat skeletal muscle cells. J. Biol. Chem. 1997, 272, 23653–23658. [Google Scholar] [CrossRef][Green Version]
- Lei, J.; Mariash, C.N.; Ingbar, D.H. 3,3’,5-Triiodo-L-thyronine up-regulation of Na,K-ATPase activity and cell surface expression in alveolar epithelial cells is Src kinase- and phosphoinositide 3-kinase-dependent. J. Biol. Chem. 2004, 279, 47589–47600. [Google Scholar] [CrossRef]
- Kjeldsen, K.; Norgaard, A.; Gotzsche, C.O.; Thomassen, A.; Clausen, T. Effect of thyroid function on number of Na-K pumps in human skeletal muscle. Lancet 1984, 2, 8–10. [Google Scholar] [CrossRef]
- Logvinenko, N.S.; Dulubova, I.; Fedosova, N.; Larsson, S.H.; Nairn, A.C.; Esmann, M.; Greengard, P.; Aperia, A. Phosphorylation by protein kinase C of serine-23 of the alpha-1 subunit of rat Na+,K(+)-ATPase affects its conformational equilibrium. Proc. Natl. Acad. Sci. USA 1996, 93, 9132–9137. [Google Scholar] [CrossRef] [PubMed]
- Feraille, E.; Beguin, P.; Carranza, M.L.; Gonin, S.; Rousselot, M.; Martin, P.Y.; Favre, H.; Geering, K. Is phosphorylation of the alpha1 subunit at Ser-16 involved in the control of Na,K-ATPase activity by phorbol ester-activated protein kinase C? Mol. Biol. Cell 2000, 11, 39–50. [Google Scholar] [CrossRef]
- Vasilets, L.A.; Postina, R.; Kirichenko, S.N. Mutations of Ser-23 of the alpha1 subunit of the rat Na+/K+-ATPase to negatively charged amino acid residues mimic the functional effect of PKC-mediated phosphorylation. FEBS Lett. 1999, 455, 8–12. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Major, E.; Keller, I.; Horváth, D.; Tamás, I.; Erdődi, F.; Lontay, B. Smoothelin-Like Protein 1 Regulates the Thyroid Hormone-Induced Homeostasis and Remodeling of C2C12 Cells via the Modulation of Myosin Phosphatase. Int. J. Mol. Sci. 2021, 22, 10293. https://doi.org/10.3390/ijms221910293
Major E, Keller I, Horváth D, Tamás I, Erdődi F, Lontay B. Smoothelin-Like Protein 1 Regulates the Thyroid Hormone-Induced Homeostasis and Remodeling of C2C12 Cells via the Modulation of Myosin Phosphatase. International Journal of Molecular Sciences. 2021; 22(19):10293. https://doi.org/10.3390/ijms221910293
Chicago/Turabian StyleMajor, Evelin, Ilka Keller, Dániel Horváth, István Tamás, Ferenc Erdődi, and Beáta Lontay. 2021. "Smoothelin-Like Protein 1 Regulates the Thyroid Hormone-Induced Homeostasis and Remodeling of C2C12 Cells via the Modulation of Myosin Phosphatase" International Journal of Molecular Sciences 22, no. 19: 10293. https://doi.org/10.3390/ijms221910293
APA StyleMajor, E., Keller, I., Horváth, D., Tamás, I., Erdődi, F., & Lontay, B. (2021). Smoothelin-Like Protein 1 Regulates the Thyroid Hormone-Induced Homeostasis and Remodeling of C2C12 Cells via the Modulation of Myosin Phosphatase. International Journal of Molecular Sciences, 22(19), 10293. https://doi.org/10.3390/ijms221910293