
Mechanisms of DNA Methylation in Virus-Host Interaction in Hepatitis B Infection: Pathogenesis and Oncogenetic Properties


{kind=link}
Abstract
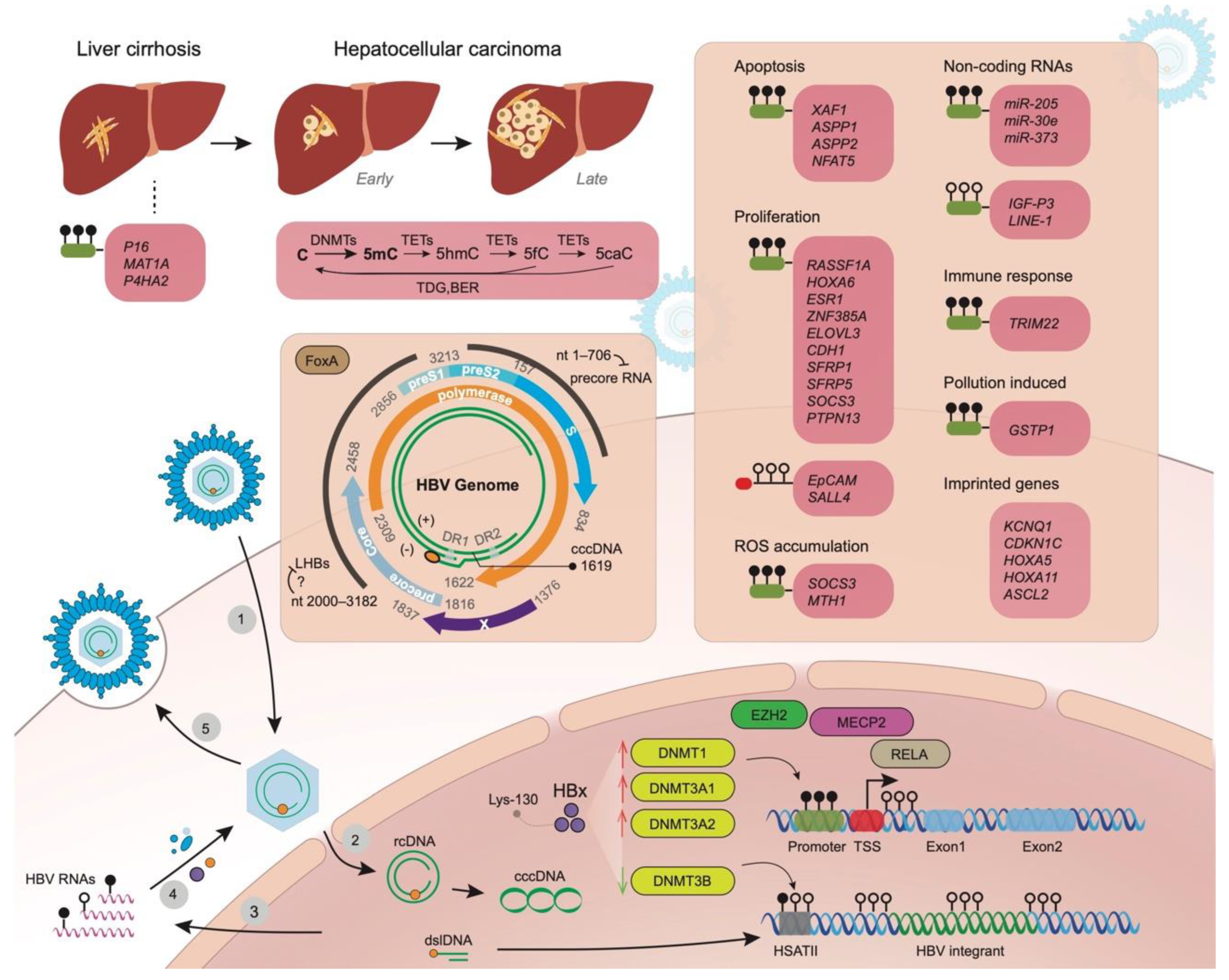
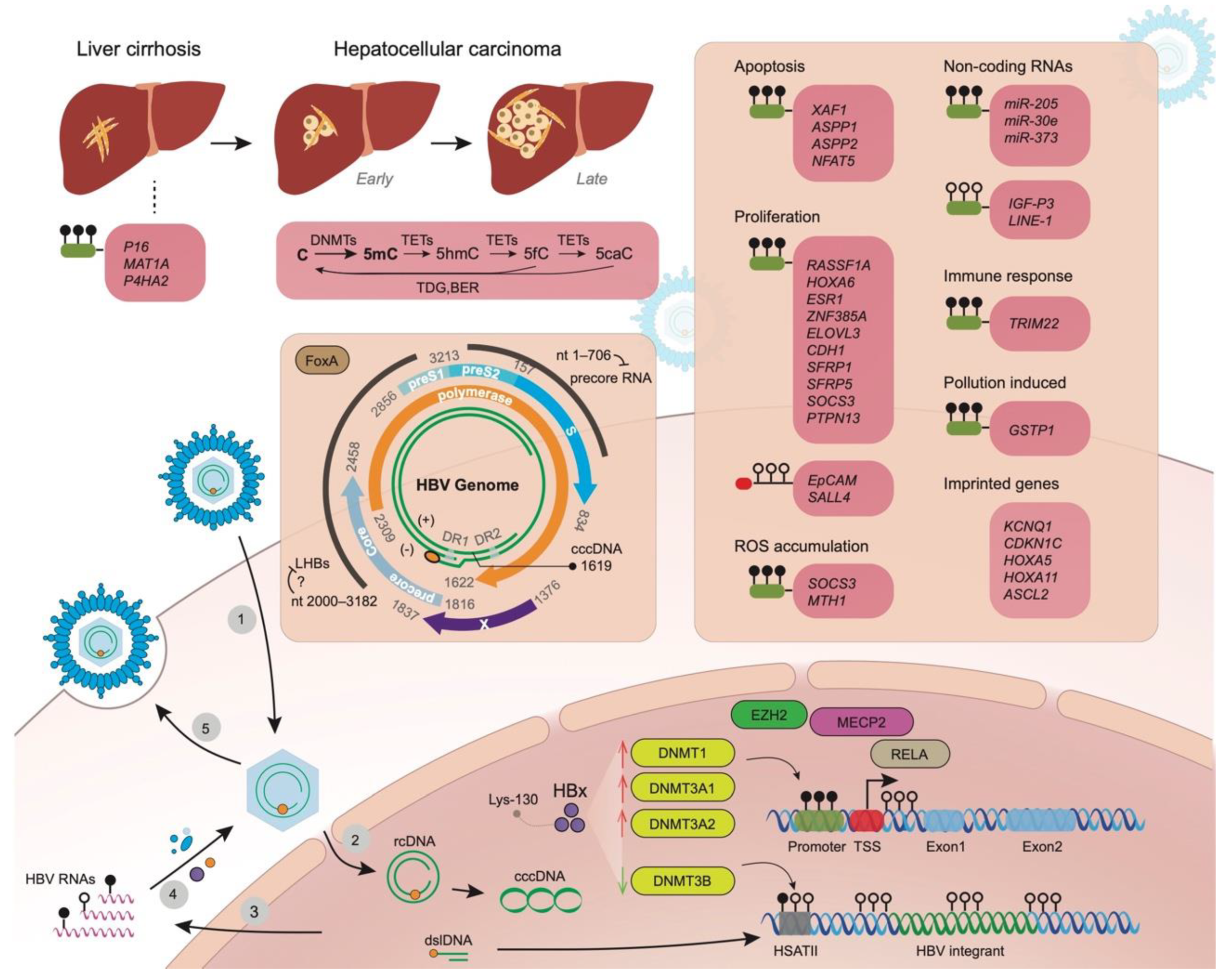
:1. Background
2. Aberrant Methylation of Promoter Regions of Genes in Liver Disease Progression Related to Tumorigenesis
3. Apoptosis, Proliferation, and Reactive Oxygen Species (ROS) Accumulation
4. Hypomethylation, Methylation Changes to Non-Coding Genes Other Than miRNAs and Imprinted Genes
5. Changes of Host Immune Status, Infected Hepatocytes, and Exposure to Environmental Pollutant
6. Viral Genome Methylation in HBV Replication, Modifications of cccDNA and HBV Integrants
7. Technical Challenges in Future Research
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brechot, C.; Pourcel, C.; Louise, A.; Rain, B.; Tiollais, P. Presence of integrated hepatitis B virus DNA sequences in cellular DNA of human hepatocellular carcinoma. Nature 1980, 286, 533–535. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liu, G.; Lin, Z.; Lin, Z.; Tian, N.; Lin, X.; Tan, J.; Huang, B.; Ji, X.; Pi, L.; et al. The association of lncRNA SNPs and SNPs-environment interactions based on GWAS with HBV-related HCC risk and progression. Mol. Genet. Genomic Med. 2021, 9, e1585. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Bi, X.; Huang, Y.; Zhao, J.; Li, Z.; Zhou, J.; Zhang, M.; Huang, Z.; Zhao, H.; Cai, J. Variants identified by hepatocellular carcinoma and chronic hepatitis B virus infection susceptibility GWAS associated with survival in HBV-related hepatocellular carcinoma. PLoS ONE 2014, 9, e101586. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Qian, J.; Yang, Y.; Zhao, W.; Dai, J.; Bei, J.X.; Foo, J.N.; McLaren, P.J.; Li, Z.; Yang, J.; et al. GWAS identifies novel susceptibility loci on 6p21.32 and 21q21.3 for hepatocellular carcinoma in chronic hepatitis B virus carriers. PLoS Genet. 2012, 8, e1002791. [Google Scholar] [CrossRef]
- Jiang, D.K.; Sun, J.; Cao, G.; Liu, Y.; Lin, D.; Gao, Y.Z.; Ren, W.H.; Long, X.D.; Zhang, H.; Ma, X.P.; et al. Genetic variants in STAT4 and HLA-DQ genes confer risk of hepatitis B virus-related hepatocellular carcinoma. Nat. Genet. 2013, 45, 72–75. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.Y.; Yu, M.W.; Lin, S.M.; Lee, S.D.; Chen, C.L.; Chen, D.S.; Chen, P.J. Genome-wide association analysis identifies a GLUL haplotype for familial hepatitis B virus-related hepatocellular carcinoma. Cancer 2017, 123, 3966–3976. [Google Scholar] [CrossRef]
- Zhao, Y.; Xue, F.; Sun, J.; Guo, S.; Zhang, H.; Qiu, B.; Geng, J.; Gu, J.; Zhou, X.; Wang, W.; et al. Genome-wide methylation profiling of the different stages of hepatitis B virus-related hepatocellular carcinoma development in plasma cell-free DNA reveals potential biomarkers for early detection and high-risk monitoring of hepatocellular carcinoma. Clin. Epigenet. 2014, 6, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhang, J.; Wang, R.; Guo, S.; Zhang, H.; Ma, Y.; Liu, Q.; Chu, H.; Xu, X.; Zhang, Y.; et al. Hypermethylation reduces the expression of PNPLA7 in hepatocellular carcinoma. Oncol. Lett. 2016, 12, 670–674. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Cui, Y.; Wang, W.; Gu, J.; Guo, S.; Ma, K.; Luo, X. Hypomethylation of the hsa-miR-191 locus causes high expression of hsa-mir-191 and promotes the epithelial-to-mesenchymal transition in hepatocellular carcinoma. Neoplasia 2011, 13, 841–853. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhang, J.; Ma, Y.; Chen, L.; Guo, S.; Zhang, X.; Ma, Y.; Wu, L.; Pei, X.; Liu, S.; et al. Association study of miR149 rs2292832 and miR608 rs4919510 and the risk of hepatocellular carcinoma in a largescale population. Mol. Med. Rep. 2014, 10, 2736–2744. [Google Scholar] [CrossRef]
- Shih, Y.L.; Kuo, C.C.; Yan, M.D.; Lin, Y.W.; Hsieh, C.B.; Hsieh, T.Y. Quantitative methylation analysis reveals distinct association between PAX6 methylation and clinical characteristics with different viral infections in hepatocellular carcinoma. Clin. Epigenet. 2016, 8, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edamoto, Y.; Hara, A.; Biernat, W.; Terracciano, L.; Cathomas, G.; Riehle, H.M.; Matsuda, M.; Fujii, H.; Scoazec, J.Y.; Ohgaki, H. Alterations of RB1, p53 and Wnt pathways in hepatocellular carcinomas associated with hepatitis C, hepatitis B and alcoholic liver cirrhosis. Int. J. Cancer 2003, 106, 334–341. [Google Scholar] [CrossRef]
- Li, H.; Zhang, M.; Linghu, E.; Zhou, F.; Herman, J.G.; Hu, L.; Guo, M. Epigenetic silencing of TMEM176A activates ERK signaling in human hepatocellular carcinoma. Clin. Epigenet. 2018, 10, 137. [Google Scholar] [CrossRef] [PubMed]
- Hensel, K.O.; Rendon, J.C.; Navas, M.C.; Rots, M.G.; Postberg, J. Virus-host interplay in hepatitis B virus infection and epigenetic treatment strategies. FEBS J. 2017, 284, 3550–3572. [Google Scholar] [CrossRef] [Green Version]
- Ozen, C.; Yildiz, G.; Dagcan, A.T.; Cevik, D.; Ors, A.; Keles, U.; Topel, H.; Ozturk, M. Genetics and epigenetics of liver cancer. New Biotechnol. 2013, 30, 381–384. [Google Scholar] [CrossRef]
- Liu, J.; Jiang, J.; Mo, J.; Liu, D.; Cao, D.; Wang, H.; He, Y.; Wang, H. Global DNA 5-Hydroxymethylcytosine and 5-Formylcytosine Contents Are Decreased in the Early Stage of Hepatocellular Carcinoma. Hepatology 2019, 69, 196–208. [Google Scholar] [CrossRef]
- Park, I.Y.; Sohn, B.H.; Yu, E.; Suh, D.J.; Chung, Y.H.; Lee, J.H.; Surzycki, S.J.; Lee, Y.I. Aberrant epigenetic modifications in hepatocarcinogenesis induced by hepatitis B virus X protein. Gastroenterology 2007, 132, 1476–1494. [Google Scholar] [CrossRef]
- Huang, J.; Wang, Y.; Guo, Y.; Sun, S. Down-regulated microRNA-152 induces aberrant DNA methylation in hepatitis B virus-related hepatocellular carcinoma by targeting DNA methyltransferase 1. Hepatology 2010, 52, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Vivekanandan, P.; Daniel, H.D.; Kannangai, R.; Martinez-Murillo, F.; Torbenson, M. Hepatitis B virus replication induces methylation of both host and viral DNA. J. Virol. 2010, 84, 4321–4329. [Google Scholar] [CrossRef] [Green Version]
- Gunther, S.; Li, B.C.; Miska, S.; Kruger, D.H.; Meisel, H.; Will, H. A novel method for efficient amplification of whole hepatitis B virus genomes permits rapid functional analysis and reveals deletion mutants in immunosuppressed patients. J. Virol. 1995, 69, 5437–5444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benhenda, S.; Ducroux, A.; Rivière, L.; Sobhian, B.; Ward, M.D.; Dion, S.; Hantz, O.; Protzer, U.; Michel, M.L.; Benkirane, M.; et al. Methyltransferase PRMT1 is a binding partner of HBx and a negative regulator of hepatitis B virus transcription. J. Virol. 2013, 87, 4360–4371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, P.F.; Lee, T.C.; Lin, P.J.; Lee, P.H.; Jeng, Y.M.; Chen, C.H.; Liang, J.D.; Chiou, L.L.; Huang, G.T.; Lee, H.S. Differential DNA methylation associated with hepatitis B virus infection in hepatocellular carcinoma. Int. J. Cancer 2007, 121, 1257–1264. [Google Scholar] [CrossRef]
- Shim, Y.H.; Yoon, G.S.; Choi, H.J.; Chung, Y.H.; Yu, E. p16 Hypermethylation in the early stage of hepatitis B virus-associated hepatocarcinogenesis. Cancer Lett. 2003, 190, 213–219. [Google Scholar] [CrossRef]
- Kuramoto, J.; Arai, E.; Tian, Y.; Funahashi, N.; Hiramoto, M.; Nammo, T.; Nozaki, Y.; Takahashi, Y.; Ito, N.; Shibuya, A.; et al. Genome-wide DNA methylation analysis during non-alcoholic steatohepatitis-related multistage hepatocarcinogenesis: Comparison with hepatitis virus-related carcinogenesis. Carcinogenesis 2017, 38, 261–270. [Google Scholar] [CrossRef]
- Avila, M.A.; Berasain, C.; Torres, L.; Martin-Duce, A.; Corrales, F.J.; Yang, H.; Prieto, J.; Lu, S.C.; Caballeria, J.; Rodes, J.; et al. Reduced mRNA abundance of the main enzymes involved in methionine metabolism in human liver cirrhosis and hepatocellular carcinoma. J. Hepatol. 2000, 33, 907–914. [Google Scholar] [CrossRef] [Green Version]
- Bing, Y.; Zhu, S.; Yu, G.; Li, T.; Liu, W.; Li, C.; Wang, Y.; Qi, H.; Guo, T.; Yuan, Y.; et al. Glucocorticoid-induced S-adenosylmethionine enhances the interferon signaling pathway by restoring STAT1 protein methylation in hepatitis B virus-infected cells. J. Biol. Chem. 2014, 289, 32639–32655. [Google Scholar] [CrossRef] [Green Version]
- Dang, S.; Zhou, J.; Chen, Y.; Chen, P.; Ji, M.; Shi, B.; Yang, Q.; Hou, P. Dynamic expression of ZNF382 and its tumor-suppressor role in hepatitis B virus-related hepatocellular carcinogenesis. Oncogene 2019, 38, 4804–4819. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Dong, S.; Qiao, F.; Lu, S.; Song, Y.; Lao, Y.; Li, Y.; Zeng, T.; Hu, J.; Zhang, L.; et al. HBx-mediated miR-21 upregulation represses tumor-suppressor function of PDCD4 in hepatocellular carcinoma. Oncogene 2013, 32, 3296–3305. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Zhang, J.; Cui, M.; Liu, F.; You, X.; Du, Y.; Gao, Y.; Zhang, S.; Lu, Z.; Ye, L.; et al. Hepatitis B virus X protein inhibits tumor suppressor miR-205 through inducing hypermethylation of miR-205 promoter to enhance carcinogenesis. Neoplasia 2013, 15, 1282–1291. [Google Scholar] [CrossRef] [Green Version]
- Feng, G.X.; Li, J.; Yang, Z.; Zhang, S.Q.; Liu, Y.X.; Zhang, W.Y.; Ye, L.H.; Zhang, X.D. Hepatitis B virus X protein promotes the development of liver fibrosis and hepatoma through downregulation of miR-30e targeting P4HA2 mRNA. Oncogene 2017, 36, 6895–6905. [Google Scholar] [CrossRef] [Green Version]
- Boyault, S.; Rickman, D.S.; de Reyniès, A.; Balabaud, C.; Rebouissou, S.; Jeannot, E.; Hérault, A.; Saric, J.; Belghiti, J.; Franco, D.; et al. Transcriptome classification of HCC is related to gene alterations and to new therapeutic targets. Hepatology 2007, 45, 42–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Van Nhieu, J.T.; Prigent, S.; Srivatanakul, P.; Tiollais, P.; Buendia, M.A. Altered expression of E-cadherin in hepatocellular carcinoma: Correlations with genetic alterations, beta-catenin expression, and clinical features. Hepatology 2002, 36, 692–701. [Google Scholar] [CrossRef]
- Lee, J.O.; Kwun, H.J.; Jung, J.K.; Choi, K.H.; Min, D.S.; Jang, K.L. Hepatitis B virus X protein represses E-cadherin expression via activation of DNA methyltransferase 1. Oncogene 2005, 24, 6617–6625. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Lian, Z.; Han, S.; Waye, M.M.; Wang, H.; Wu, M.C.; Wu, K.; Ding, J.; Arbuthnot, P.; Kew, M.; et al. Downregulation of E-cadherin by hepatitis B virus X antigen in hepatocellullar carcinoma. Oncogene 2006, 25, 1008–1017. [Google Scholar] [CrossRef] [Green Version]
- Arzumanyan, A.; Friedman, T.; Kotei, E.; Ng, I.O.; Lian, Z.; Feitelson, M.A. Epigenetic repression of E-cadherin expression by hepatitis B virus x antigen in liver cancer. Oncogene 2012, 31, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Wu, L.M.; Zhou, L.; Chen, Q.X.; Xie, H.Y.; Feng, X.W.; Zheng, S.S. Predictive value of expression and promoter hypermethylation of XAF1 in hepatitis B virus-associated hepatocellular carcinoma treated with transplantation. Ann. Surg. Oncol. 2008, 15, 3494–3502. [Google Scholar] [CrossRef]
- Zhao, J.; Wu, G.; Bu, F.; Lu, B.; Liang, A.; Cao, L.; Tong, X.; Lu, X.; Wu, M.; Guo, Y. Epigenetic silence of ankyrin-repeat-containing, SH3-domain-containing, and proline-rich-region- containing protein 1 (ASPP1) and ASPP2 genes promotes tumor growth in hepatitis B virus-positive hepatocellular carcinoma. Hepatology 2010, 51, 142–153. [Google Scholar] [CrossRef]
- Qin, X.; Li, C.; Guo, T.; Chen, J.; Wang, H.T.; Wang, Y.T.; Xiao, Y.S.; Li, J.; Liu, P.; Liu, Z.S.; et al. Upregulation of DARS2 by HBV promotes hepatocarcinogenesis through the miR-30e-5p/MAPK/NFAT5 pathway. J. Exp. Clin. Cancer Res. 2017, 36, 148. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, Y.; Shinjo, K.; Shimizu, Y.; Sano, T.; Yamao, K.; Gao, W.; Fujii, M.; Osada, H.; Sekido, Y.; Murakami, S.; et al. Hepatitis virus infection affects DNA methylation in mice with humanized livers. Gastroenterology 2014, 146, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Sia, D.; Villanueva, A.; Friedman, S.L.; Llovet, J.M. Liver Cancer Cell of Origin, Molecular Class, and Effects on Patient Prognosis. Gastroenterology 2017, 152, 745–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jhunjhunwala, S.; Jiang, Z.; Stawiski, E.W.; Gnad, F.; Liu, J.; Mayba, O.; Du, P.; Diao, J.; Johnson, S.; Wong, K.F.; et al. Diverse modes of genomic alteration in hepatocellular carcinoma. Genome Biol. 2014, 15, 436. [Google Scholar] [CrossRef] [Green Version]
- Xie, Q.; Chen, L.; Shan, X.; Shan, X.; Tang, J.; Zhou, F.; Chen, Q.; Quan, H.; Nie, D.; Zhang, W.; et al. Epigenetic silencing of SFRP1 and SFRP5 by hepatitis B virus X protein enhances hepatoma cell tumorigenicity through Wnt signaling pathway. Int. J. Cancer 2014, 135, 635–646. [Google Scholar] [CrossRef]
- Zhang, X.; You, Q.; Zhang, X.; Chen, X. SOCS3 Methylation Predicts a Poor Prognosis in HBV Infection-Related Hepatocellular Carcinoma. Int. J. Mol. Sci. 2015, 16, 22662–22675. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Li, J.; Feng, G.; Wang, Y.; Yang, G.; Liu, Y.; Zhang, S.; Feng, J.; Zhang, X. Hepatitis B virus X protein enhances hepatocarcinogenesis by depressing the targeting of NUSAP1 mRNA by miR-18b. Cancer Biol. Med. 2019, 16, 276–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munz, M.; Kieu, C.; Mack, B.; Schmitt, B.; Zeidler, R.; Gires, O. The carcinoma-associated antigen EpCAM upregulates c-myc and induces cell proliferation. Oncogene 2004, 23, 5748–5758. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Zhang, H.; Pascuzzi, P.E.; Andrisani, O. Hepatitis B virus X protein induces EpCAM expression via active DNA demethylation directed by RelA in complex with EZH2 and TET2. Oncogene 2016, 35, 715–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, H.; Cui, Z.; Zhang, H.; Mani, S.K.; Diab, A.; Lefrancois, L.; Fares, N.; Merle, P.; Andrisani, O. DNA demethylation induces SALL4 gene re-expression in subgroups of hepatocellular carcinoma associated with Hepatitis B or C virus infection. Oncogene 2017, 36, 2435–2445. [Google Scholar] [CrossRef] [Green Version]
- Pasini, D.; Bracken, A.P.; Hansen, J.B.; Capillo, M.; Helin, K. The polycomb group protein Suz12 is required for embryonic stem cell differentiation. Mol. Cell Biol. 2007, 27, 3769–3779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salerno, D.; Chiodo, L.; Alfano, V.; Floriot, O.; Cottone, G.; Paturel, A.; Pallocca, M.; Plissonnier, M.L.; Jeddari, S.; Belloni, L.; et al. Hepatitis B protein HBx binds the DLEU2 lncRNA to sustain cccDNA and host cancer-related gene transcription. Gut 2020, 69, 2016–2024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mani, S.K.; Zhang, H.; Diab, A.; Pascuzzi, P.E.; Lefrancois, L.; Fares, N.; Bancel, B.; Merle, P.; Andrisani, O. EpCAM-regulated intramembrane proteolysis induces a cancer stem cell-like gene signature in hepatitis B virus-infected hepatocytes. J. Hepatol. 2016, 65, 888–898. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Xing, Z.; Mani, S.K.; Bancel, B.; Durantel, D.; Zoulim, F.; Tran, E.J.; Merle, P.; Andrisani, O. RNA helicase DEAD box protein 5 regulates Polycomb repressive complex 2/Hox transcript antisense intergenic RNA function in hepatitis B virus infection and hepatocarcinogenesis. Hepatology 2016, 64, 1033–1048. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Diab, A.; Fan, H.; Mani, S.K.; Hullinger, R.; Merle, P.; Andrisani, O. PLK1 and HOTAIR Accelerate Proteasomal Degradation of SUZ12 and ZNF198 during Hepatitis B Virus-Induced Liver Carcinogenesis. Cancer Res. 2015, 75, 2363–2374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; Huang, P.; Mao, K.; He, C.; Xu, Q.; Zhang, M.; Liu, H.; Zhou, Z.; Zhou, Q.; Zhou, Q.; et al. Anti-oncogene PTPN13 inactivation by hepatitis B virus X protein counteracts IGF2BP1 to promote hepatocellular carcinoma progression. Oncogene 2020, 40, 28–45. [Google Scholar] [CrossRef] [PubMed]
- Moloney, J.N.; Cotter, T.G. ROS signalling in the biology of cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef]
- Yuan, K.; Lei, Y.; Chen, H.N.; Chen, Y.; Zhang, T.; Li, K.; Xie, N.; Wang, K.; Feng, X.; Pu, Q.; et al. HBV-induced ROS accumulation promotes hepatocarcinogenesis through Snail-mediated epigenetic silencing of SOCS3. Cell Death Differ. 2016, 23, 616–627. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.T.; Liu, W.; He, Y.; Wu, Y.L.; Chen, W.N.; Lin, X.J.; Lin, X. Hepatitis B Virus X Protein Increases 8-Oxo-7,8-Dihydro-2’-Deoxyguanosine (8-Oxodg) Level via Repressing MTH1/ MTH2 Expression in Hepatocytes. Cell Physiol. Biochem. 2018, 51, 80–96. [Google Scholar] [CrossRef]
- Tang, S.; Hu, W.; Hu, J.; Wu, S.; Li, J.; Luo, Y.; Cao, M.; Zhou, H.; Jiang, X. Hepatitis B virus X protein promotes P3 transcript expression of the insulin-like growth factor 2 gene via inducing hypomethylation of P3 promoter in hepatocellular carcinoma. Liver Int. 2015, 35, 608–619. [Google Scholar] [CrossRef]
- Gao, X.D.; Qu, J.H.; Chang, X.J.; Lu, Y.Y.; Bai, W.L.; Wang, H.; Xu, Z.X.; An, L.J.; Wang, C.P.; Zeng, Z.; et al. Hypomethylation of long interspersed nuclear element-1 promoter is associated with poor outcomes for curative resected hepatocellular carcinoma. Liver Int. 2014, 34, 136–146. [Google Scholar] [CrossRef] [Green Version]
- Lambert, M.P.; Ancey, P.B.; Esposti, D.D.; Cros, M.P.; Sklias, A.; Scoazec, J.Y.; Durantel, D.; Hernandez-Vargas, H.; Herceg, Z. Aberrant DNA methylation of imprinted loci in hepatocellular carcinoma and after in vitro exposure to common risk factors. Clin. Epigenet. 2015, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Qin, L.; Jiang, S.; Li, A.; Zhang, C.; Liu, G.; Sun, J.; Sun, H.; Zhao, Y.; Li, N.; et al. The signature of HBV-related liver disease in peripheral blood mononuclear cell DNA methylation. Clin. Epigenet. 2020, 12, 81. [Google Scholar] [CrossRef]
- Gindin, Y.; Gaggar, A.; Lok, A.S.; Janssen, H.L.A.; Ferrari, C.; Subramanian, G.M.; Jiang, Z.; Masur, H.; Emmanuel, B.; Poonia, B.; et al. DNA Methylation and Immune Cell Markers Demonstrate Evidence of Accelerated Aging in Patients with Chronic HBV or HCV, with or without HIV Co-Infection. Clin. Infect. Dis. 2020, 73, e184–e190. [Google Scholar] [CrossRef] [PubMed]
- Ozato, K.; Shin, D.M.; Chang, T.H.; Morse, H.C., III. TRIM family proteins and their emerging roles in innate immunity. Nat. Rev. Immunol. 2008, 8, 849–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, K.H.; Park, E.S.; Kim, D.H.; Cho, K.C.; Kim, K.P.; Park, Y.K.; Ahn, S.H.; Park, S.H.; Kim, K.H.; Kim, C.W.; et al. Suppression of interferon-mediated anti-HBV response by single CpG methylation in the 5’-UTR of TRIM22. Gut 2018, 67, 166–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.; Xu, H.; Wu, R.; Gao, N.; Wu, N.; Li, S.; Niu, J. Identification of key genes and pathways associated with different immune statuses of hepatitis B virus infection. J. Cell Mol. Med. 2019, 23, 7474–7489. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Zhao, B.; Zhang, J.; Martin, F.L.; Huang, Q.; Liu, L.; Shen, H. Association of environmental benzo[a]pyrene exposure and DNA methylation alterations in hepatocellular carcinoma: A Chinese case-control study. Sci. Total Environ. 2016, 541, 1243–1252. [Google Scholar] [CrossRef] [Green Version]
- Zhong, S.; Tang, M.W.; Yeo, W.; Liu, C.; Lo, Y.M.; Johnson, P.J. Silencing of GSTP1 gene by CpG island DNA hypermethylation in HBV-associated hepatocellular carcinomas. Clin. Cancer Res. 2002, 8, 1087–1092. [Google Scholar]
- Niu, D.; Zhang, J.; Ren, Y.; Feng, H.; Chen, W.N. HBx genotype D represses GSTP1 expression and increases the oxidative level and apoptosis in HepG2 cells. Mol. Oncol. 2009, 3, 67–76. [Google Scholar] [CrossRef]
- Lambert, M.P.; Paliwal, A.; Vaissière, T.; Chemin, I.; Zoulim, F.; Tommasino, M.; Hainaut, P.; Sylla, B.; Scoazec, J.Y.; Tost, J.; et al. Aberrant DNA methylation distinguishes hepatocellular carcinoma associated with HBV and HCV infection and alcohol intake. J. Hepatol. 2011, 54, 705–715. [Google Scholar] [CrossRef]
- Tian, M.; Zhao, B.; Martin, F.L.; Morais, C.L.M.; Liu, L.; Huang, Q.; Zhang, J.; Shen, H. Gene-environment interactions between GSTs polymorphisms and targeted epigenetic alterations in hepatocellular carcinoma following organochlorine pesticides (OCPs) exposure. Environ. Int. 2020, 134, 105313. [Google Scholar] [CrossRef]
- Lee, C.S.; Friedman, J.R.; Fulmer, J.T.; Kaestner, K.H. The initiation of liver development is dependent on Foxa transcription factors. Nature 2005, 435, 944–947. [Google Scholar] [CrossRef]
- Vivekanandan, P.; Kannangai, R.; Ray, S.C.; Thomas, D.L.; Torbenson, M. Comprehensive genetic and epigenetic analysis of occult hepatitis B from liver tissue samples. Clin. Infect. Dis. 2008, 46, 1227–1236. [Google Scholar] [CrossRef] [Green Version]
- Vivekanandan, P.; Thomas, D.; Torbenson, M. Methylation regulates hepatitis B viral protein expression. J. Infect. Dis. 2009, 199, 1286–1291. [Google Scholar] [CrossRef] [Green Version]
- Uchida, T.; Imamura, M.; Hayes, C.N.; Hiraga, N.; Kan, H.; Tsuge, M.; Abe-Chayama, H.; Zhang, Y.; Makokha, G.N.; Aikata, H.; et al. Persistent Loss of Hepatitis B Virus Markers in Serum without Cellular Immunity by Combination of Peginterferon and Entecavir Therapy in Humanized Mice. Antimicrob. Agents Chemother. 2017, 61, e00725-17. [Google Scholar] [CrossRef] [Green Version]
- Raney, A.K.; Zhang, P.; McLachlan, A. Regulation of transcription from the hepatitis B virus large surface antigen promoter by hepatocyte nuclear factor 3. J. Virol. 1995, 69, 3265–3272. [Google Scholar] [CrossRef] [Green Version]
- Oropeza, C.E.; Tarnow, G.; Taha, T.Y.; Shalaby, R.E.; Hyde, M.V.; Maienschein-Cline, M.; Green, S.J.; McLachlan, A. Relative DNA methylation and demethylation efficiencies during postnatal liver development regulate hepatitis B virus biosynthesis. J. Virol. 2020, 95, e02148-20. [Google Scholar] [CrossRef]
- McFadden, V.C.; Shalaby, R.E.; Iram, S.; Oropeza, C.E.; Landolfi, J.A.; Lyubimov, A.V.; Maienschein-Cline, M.; Green, S.J.; Kaestner, K.H.; McLachlan, A. Hepatic deficiency of the pioneer transcription factor FoxA restricts hepatitis B virus biosynthesis by the developmental regulation of viral DNA methylation. PLoS Pathog. 2017, 13, e1006239. [Google Scholar] [CrossRef]
- Raney, A.K.; McLachlan, A. Characterization of the hepatitis B virus major surface antigen promoter hepatocyte nuclear factor 3 binding site. J. Gen. Virol. 1997, 78, 3029–3038. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Inoue, J.; Ninomiya, M.; Kakazu, E.; Iwata, T.; Takai, S.; Sano, A.; Kogure, T.; Shimosegawa, T.; Masamune, A. Effect of viral DNA methylation on expression of hepatitis B virus proteins depends on the virus genotype. Virus Genes 2020, 56, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, C.; Cohen, D.; Dubois, A.; Martinez, M.G.; Petitjean, K.; Corlu, A.; Testoni, B.; Hernandez-Vargas, H.; Chemin, I. Cas9-targeted nanopore sequencing reveals epigenetic heterogeneity after de novo assembly of native full-length hepatitis B virus genomes. Microb. Genom. 2021, 7, 000507. [Google Scholar] [CrossRef]
- Zhang, Y.; Mao, R.; Guo, H.; Zhang, J. Detection of HBV cccDNA Methylation from Clinical Samples by Bisulfite Sequencing and Methylation-Specific PCR. Methods Mol. Biol. 2017, 1540, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Jeong, H.; Lee, S.Y.; Kim, S.S.; Jang, K.L. Hepatitis B Virus X Protein Stimulates Virus Replication Via DNA Methylation of the C-1619 in Covalently Closed Circular DNA. Mol. Cells 2019, 42, 67–78. [Google Scholar] [CrossRef]
- Kostyushev, D.; Brezgin, S.; Kostyusheva, A.; Zarifyan, D.; Goptar, I.; Chulanov, V. Orthologous CRISPR/Cas9 systems for specific and efficient degradation of covalently closed circular DNA of hepatitis B virus. Cell Mol. Life Sci. 2019, 76, 1779–1794. [Google Scholar] [CrossRef]
- van Breugel, P.C.; Robert, E.I.; Mueller, H.; Decorsière, A.; Zoulim, F.; Hantz, O.; Strubin, M. Hepatitis B virus X protein stimulates gene expression selectively from extrachromosomal DNA templates. Hepatology 2012, 56, 2116–2124. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.H.; Robinson, W.S. Integrated hepatitis B virus DNA sequences specifying the major viral core polypeptide are methylated in PLC/PRF/5 cells. Proc. Natl. Acad. Sci. USA 1983, 80, 2534–2538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowyer, S.M.; Dusheiko, G.M.; Schoub, B.D.; Kew, M.C. Expression of the hepatitis B virus genome in chronic hepatitis B carriers and patients with hepatocellular carcinoma. Proc. Natl. Acad. Sci. USA 1987, 84, 847–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyoshi, E.; Fujii, J.; Hayashi, N.; Ueda, K.; Towata, T.; Fusamoto, H.; Kamada, T.; Taniguchi, N. Enhancement of hepatitis-B surface-antigen expression by 5-azacytidine in a hepatitis-B-virus-transfected cell line. Int. J. Cancer 1992, 52, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Yamamoto, H.; Oikawa, R.; Toyota, M.; Yamamoto, M.; Kokudo, N.; Tanaka, S.; Arii, S.; Yotsuyanagi, H.; Koike, K.; et al. DNA methylation at hepatitis B viral integrants is associated with methylation at flanking human genomic sequences. Genome Res. 2015, 25, 328–337. [Google Scholar] [CrossRef] [Green Version]
- Hama, N.; Totoki, Y.; Miura, F.; Tatsuno, K.; Saito-Adachi, M.; Nakamura, H.; Arai, Y.; Hosoda, F.; Urushidate, T.; Ohashi, S.; et al. Epigenetic landscape influences the liver cancer genome architecture. Nat. Commun. 2018, 9, 1643. [Google Scholar] [CrossRef]
- Zhang, H.; Dong, P.; Guo, S.; Tao, C.; Chen, W.; Zhao, W.; Wang, J.; Cheung, R.; Villanueva, A.; Fan, J.; et al. Hypomethylation in HBV integration regions aids non-invasive surveillance to hepatocellular carcinoma by low-pass genome-wide bisulfite sequencing. BMC Med. 2020, 18, 200. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, K.; Protzer, U.; Zeng, C. HBV Integration Induces Complex Interactions between Host and Viral Genomic Functions at the Insertion Site. J. Clin. Transl. Hepatol. 2021, 9, 399–408. [Google Scholar] [CrossRef]
- Zhao, K.; Liu, A.; Xia, Y. Insights into Hepatitis B Virus DNA Integration-55 Years after Virus Discovery. Innovation 2020, 1, 100034. [Google Scholar] [CrossRef]
- Frommer, M.; McDonald, L.E.; Millar, D.S.; Collis, C.M.; Watt, F.; Grigg, G.W.; Molloy, P.L.; Paul, C.L. A genomic sequencing protocol that yields a positive display of 5-methylcytosine residues in individual DNA strands. Proc. Natl. Acad. Sci. USA 1992, 89, 1827–1831. [Google Scholar] [CrossRef] [Green Version]
- Herman, J.G.; Graff, J.R.; Myohanen, S.; Nelkin, B.D.; Baylin, S.B. Methylation-specific PCR: A novel PCR assay for methylation status of CpG islands. Proc. Natl. Acad. Sci. USA 1996, 93, 9821–9826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalgo, M.L.; Jones, P.A. Rapid quantitation of methylation differences at specific sites using methylation-sensitive single nucleotide primer extension (Ms-SNuPE). Nucleic Acids Res. 1997, 25, 2529–2531. [Google Scholar] [CrossRef] [Green Version]
- Meissner, A.; Gnirke, A.; Bell, G.W.; Ramsahoye, B.; Lander, E.S.; Jaenisch, R. Reduced representation bisulfite sequencing for comparative high-resolution DNA methylation analysis. Nucleic Acids Res. 2005, 33, 5868–5877. [Google Scholar] [CrossRef] [Green Version]
- Lister, R.; Pelizzola, M.; Dowen, R.H.; Hawkins, R.D.; Hon, G.; Tonti-Filippini, J.; Nery, J.R.; Lee, L.; Ye, Z.; Ngo, Q.M.; et al. Human DNA methylomes at base resolution show widespread epigenomic differences. Nature 2009, 462, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, A.; Kapranov, P.; Kaminsky, Z.; Flanagan, J.; Assadzadeh, A.; Yau, P.; Virtanen, C.; Winegarden, N.; Cheng, J.; Gingeras, T.; et al. Microarray-based DNA methylation profiling: Technology and applications. Nucleic Acids Res. 2006, 34, 528–542. [Google Scholar] [CrossRef]
- Xie, S.; Leung, A.W.-S.; Zheng, Z.; Zhang, D.; Xiao, C.; Luo, R.; Luo, M.; Zhang, S. Applications and potentials of nanopore sequencing in the (epi)genome and (epi)transcriptome era. Innovation 2021, 2, 100153. [Google Scholar] [CrossRef]
- Zhang, H.; Cai, R.; Dai, J.; Sun, W. EMeth: An EM algorithm for cell type decomposition based on DNA methylation data. Sci. Rep. 2021, 11, 5717. [Google Scholar] [CrossRef]
- Scherer, M.; Nazarov, P.V.; Toth, R.; Sahay, S.; Kaoma, T.; Maurer, V.; Vedeneev, N.; Plass, C.; Lengauer, T.; Walter, J.; et al. Reference-free deconvolution, visualization and interpretation of complex DNA methylation data using DecompPipeline, MeDeCom and FactorViz. Nat. Protoc. 2020, 15, 3240–3263. [Google Scholar] [CrossRef]
- Gervin, K.; Salas, L.A.; Bakulski, K.M.; van Zelm, M.C.; Koestler, D.C.; Wiencke, J.K.; Duijts, L.; Moll, H.A.; Kelsey, K.T.; Kobor, M.S.; et al. Systematic evaluation and validation of reference and library selection methods for deconvolution of cord blood DNA methylation data. Clin. Epigenet. 2019, 11, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, P.; Guo, H.; Ren, Y.; Hou, Y.; Dong, J.; Li, R.; Lian, Y.; Fan, X.; Hu, B.; Gao, Y.; et al. Single-cell DNA methylome sequencing of human preimplantation embryos. Nat. Genet. 2018, 50, 12–19. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.; Guo, S.; Schrodi, S.J. Mechanisms of DNA Methylation in Virus-Host Interaction in Hepatitis B Infection: Pathogenesis and Oncogenetic Properties. Int. J. Mol. Sci. 2021, 22, 9858. https://doi.org/10.3390/ijms22189858
Zhang D, Guo S, Schrodi SJ. Mechanisms of DNA Methylation in Virus-Host Interaction in Hepatitis B Infection: Pathogenesis and Oncogenetic Properties. International Journal of Molecular Sciences. 2021; 22(18):9858. https://doi.org/10.3390/ijms22189858
Chicago/Turabian StyleZhang, Dake, Shicheng Guo, and Steven J. Schrodi. 2021. "Mechanisms of DNA Methylation in Virus-Host Interaction in Hepatitis B Infection: Pathogenesis and Oncogenetic Properties" International Journal of Molecular Sciences 22, no. 18: 9858. https://doi.org/10.3390/ijms22189858
APA StyleZhang, D., Guo, S., & Schrodi, S. J. (2021). Mechanisms of DNA Methylation in Virus-Host Interaction in Hepatitis B Infection: Pathogenesis and Oncogenetic Properties. International Journal of Molecular Sciences, 22(18), 9858. https://doi.org/10.3390/ijms22189858