
Changes in Locomotor Activity and Oxidative Stress-Related Factors after the Administration of an Amino Acid Mixture by Generation and Age



Abstract
1. Introduction
2. Results
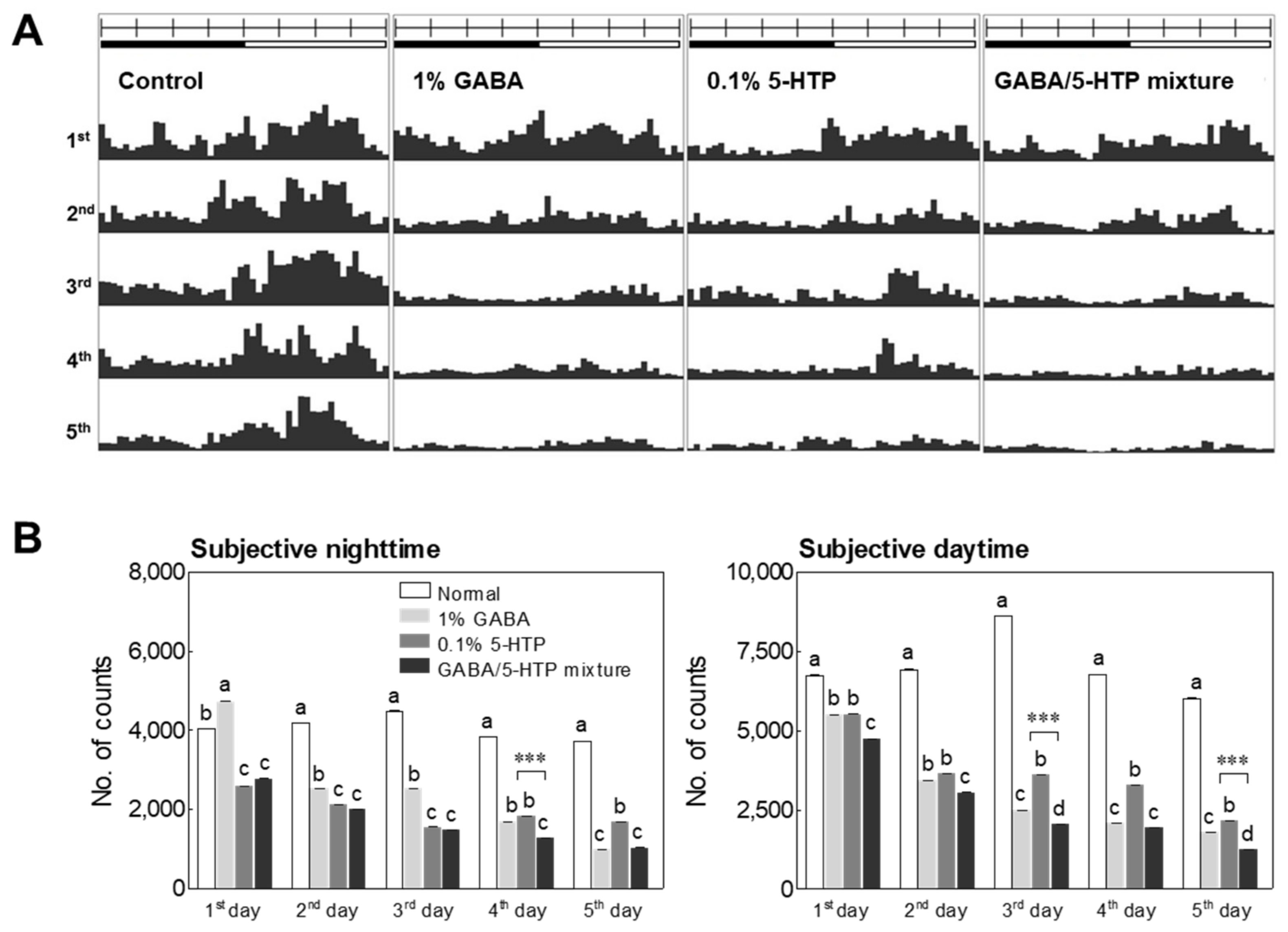
2.1. Locomotor Activity
2.2. Transcriptome Sequencing and Differential Expression Analysis
2.3. Gene Function Prediction
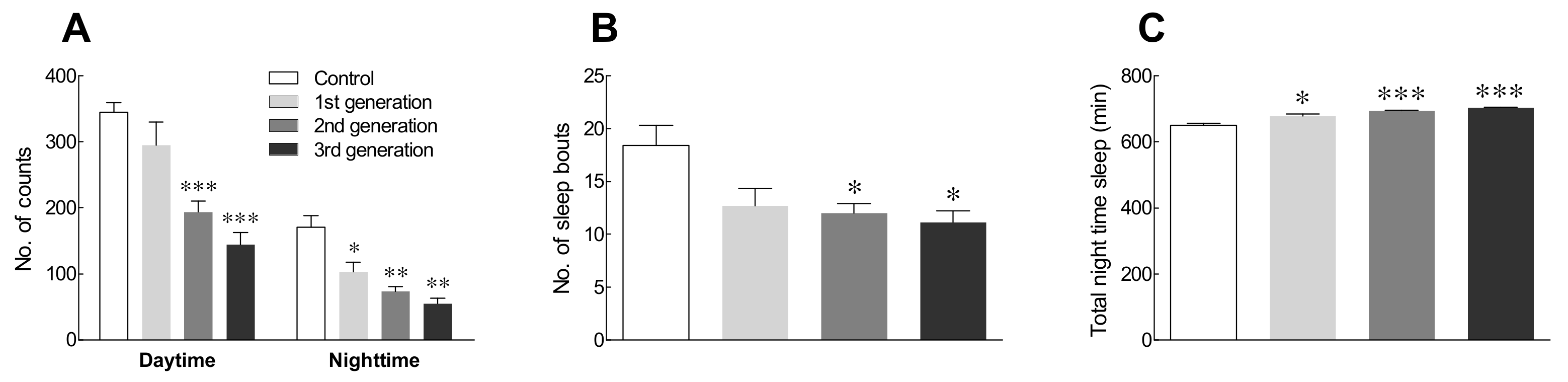
2.4. Locomotor Activity in Drosophila Flies by Generation
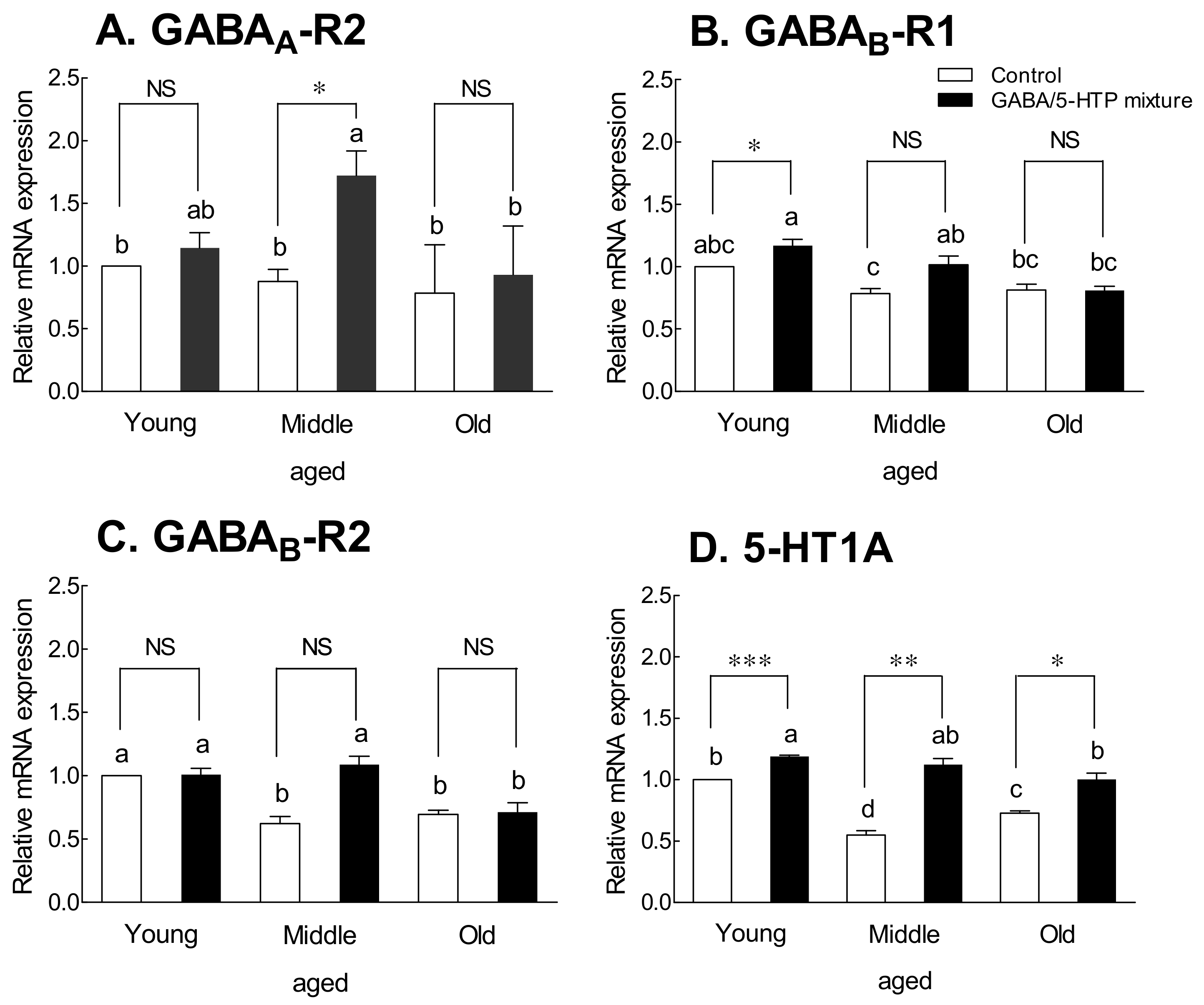
2.5. Receptor Expression in Drosophila Flies by Generation
2.6. ROS Production in Drosophila Flies by Generation
2.7. Age-Related Receptor Expression in Mice
2.8. Age-Related ROS Production in Mice
3. Discussion
4. Materials and Methods
4.1. Fly Stocks
4.2. Behavioral Assays
4.3. High-Throughput RNA Sequencing and DEG Analysis
4.4. mRNA Expression of Neurotransmitter Receptors
4.5. Measurement of MDA and H2O2
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Faludi, B.; Rozgonyi, R. Pharmacological and nonpharmacological treatment of insomnias with regard to sleep medicine. Ideggyogyaszati szemle 2018, 71, 149–159. [Google Scholar] [CrossRef]
- Pagel, J.F.; Pandi-Perumal, S.R.; Monti, J.M. Treating insomnia with medications. Sleep Sci. Pract. 2018, 2, 1–12. [Google Scholar] [CrossRef]
- Szmyd, B.; Rogut, M.; Białasiewicz, P.; Gabryelska, A. The impact of glucocorticoids and statins on sleep quality. Sleep Med. Rev. 2021, 55, 101380. [Google Scholar] [CrossRef]
- Hong, K.B.; Park, Y.; Suh, H.J. Sleep-promoting effects of a GABA/5-HTP mixture: Behavioral changes and neuromodulation in an invertebrate model. Life Sci. 2016, 150, 42–49. [Google Scholar] [CrossRef]
- Hong, K.B.; Park, Y.; Suh, H.J. Sleep-promoting effects of the GABA/5-HTP mixture in vertebrate models. Behav. Brain Res. 2016, 310, 36–41. [Google Scholar] [CrossRef]
- Hong, K.B.; Park, Y.; Suh, H.J. Two combined amino acids promote sleep activity in caffeine-induced sleepless model systems. Nutr. Res. Pract. 2018, 12, 208–214. [Google Scholar] [CrossRef]
- Dijk, D.J.; Groeger, J.; Deacon, S.; Stanley, N.P. 8.a.007 Association between individual differences in slow wave sleep, slow wave activity and sleep continuity in young, middle-aged and older men and women. Eur. Neuropsychopharmacol. 2006, 16, S538. [Google Scholar] [CrossRef]
- Dijk, D.J.; Groeger, J.A.; Stanley, N.; Deacon, S. Age-related reduction in daytime sleep propensity and nocturnal slow wave sleep. Sleep 2010, 33, 211–223. [Google Scholar] [CrossRef]
- Jenni, O.G.; Carskadon, M.A. Spectral analysis of the sleep electroencephalogram during adolescence. Sleep 2004, 27, 774–783. [Google Scholar]
- Landolt, H.P.; Dijk, D.J.; Achermann, P.; Borbély, A.A. Effect of age on the sleep EEG: Slow-wave activity and spindle frequency activity in young and middle-aged men. Brain Res. 1996, 738, 205–212. [Google Scholar] [CrossRef]
- Reimund, E. The free radical flux theory of sleep. Med. Hypotheses 1994, 43, 231–233. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, S.X.; Gozal, D. Reactive oxygen species and the brain in sleep apnea. Respir. Physiol. Neurobiol. 2010, 174, 307–316. [Google Scholar] [CrossRef]
- Gabryelska, A.; Sochal, M.; Turkiewicz, S.; Białasiewicz, P. Relationship between HIF-1 and circadian clock proteins in obstructive sleep apnea patients-preliminary study. J. Clin. Med. 2020, 9, 1599. [Google Scholar] [CrossRef]
- Villafuerte, G.; Miguel-Puga, A.; Rodríguez, E.M.; Machado, S.; Manjarrez, E.; Arias-Carrión, O. Sleep deprivation and oxidative stress in animal models: A systematic review. Oxid Med. Cell. Longev. 2015, 2015, 234952. [Google Scholar] [CrossRef]
- Gallicchio, L.; Kalesan, B. Sleep duration and mortality: A systematic review and meta-analysis. J. Sleep Res. 2009, 18, 148–158. [Google Scholar] [CrossRef]
- Fang, X.S.; Hao, J.F.; Zhou, H.Y.; Zhu, L.X.; Wang, J.H.; Song, F.Q. Pharmacological studies on the sedative-hypnotic effect of Semen Ziziphi spinosae (Suanzaoren) and Radix et Rhizoma Salviae miltiorrhizae (Danshen) extracts and the synergistic effect of their combinations. Phytomedicine 2010, 17, 75–80. [Google Scholar] [CrossRef]
- Rashmi, D.; Zanan, R.; John, S.; Khandagale, K.; Nadaf, A. γ-aminobutyric acid (GABA): Biosynthesis, role, commercial production, and applications. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2018; Volume 57, pp. 413–452. [Google Scholar]
- Deng, Y.; Wang, W.; Yu, P.; Xi, Z.; Xu, L.; Li, X.; He, N. Comparison of taurine, GABA, Glu, and Asp as scavengers of malondialdehyde in vitro and in vivo. Nanoscale Res. Lett. 2013, 8, 190. [Google Scholar] [CrossRef]
- Gutiérrez, A.; Khan, Z.U.; Miralles, C.P.; Mehta, A.K.; Ruano, D.; Araujo, F.; Vitorica, J.; De Blas, A.L. GABAA receptor subunit expression changes in the rat cerebellum and cerebral cortex during aging. Brain Res. Mol. Brain Res. 1997, 45, 59–70. [Google Scholar] [CrossRef]
- Trachte, G.J.; Uncini, T.; Hinz, M. Both stimulatory and inhibitory effects of dietary 5-hydroxytryptophan and tyrosine are found on urinary excretion of serotonin and dopamine in a large human population. Neuropsychiatr. Dis. Treat. 2009, 5, 227–235. [Google Scholar] [CrossRef][Green Version]
- Vivien-Roels, B.; Pévet, P. Melatonin: Presence and formation in invertebrates. Experientia 1993, 49, 642–647. [Google Scholar] [CrossRef]
- Chung, B.Y.; Kilman, V.L.; Keath, J.R.; Pitman, J.L.; Allada, R. The GABA(A) receptor RDL acts in peptidergic PDF neurons to promote sleep in Drosophila. Curr. Biol. 2009, 19, 386–390. [Google Scholar] [CrossRef]
- Hamasaka, Y.; Wegener, C.; Nassel, D.R. GABA modulates Drosophila circadian clock neurons via GABAB receptors and decreases in calcium. J. Neurobiol. 2005, 65, 225–240. [Google Scholar] [CrossRef]
- Yuan, Q.; Joiner, W.J.; Sehgal, A. A sleep-promoting role for the Drosophila serotonin receptor 1A. Curr. Biol. 2006, 16, 1051–1062. [Google Scholar] [CrossRef]
- Mehling, L.M.; Spottke, A.; Heidbreder, A.; Young, P.; Madea, B.; Hess, C.; Courts, C. Alterations in gene expression after gamma-hydroxybutyric acid intake-A pilot study. Int. J. Legal Med. 2017, 131, 1261–1270. [Google Scholar] [CrossRef] [PubMed]
- Grønli, J.; Clegern, W.C.; Schmidt, M.A.; Nemri, R.S.; Rempe, M.J.; Gallitano, A.L.; Wisor, J.P. Sleep homeostatic and waking behavioral phenotypes in Egr3-deficient mice associated with serotonin receptor 5-HT2 deficits. Sleep 2016, 39, 2189–2199. [Google Scholar] [CrossRef][Green Version]
- Moriya, S.; Tahara, Y.; Sasaki, H.; Ishigooka, J.; Shibata, S. Phase-delay in the light-dark cycle impairs clock gene expression and levels of serotonin, norepinephrine, and their metabolites in the mouse hippocampus and amygdala. Sleep Med. 2015, 16, 1352–1359. [Google Scholar] [CrossRef]
- Qian, Y.; Cao, Y.; Deng, B.; Yang, G.; Li, J.; Xu, R.; Zhang, D.; Huang, J.; Rao, Y. Sleep homeostasis regulated by 5HT2b receptor in a small subset of neurons in the dorsal fan-shaped body of drosophila. Elife 2017, 6, 26519. [Google Scholar] [CrossRef] [PubMed]
- Homyk, T., Jr.; Szidonya, J.; Suzuki, D.T. Behavioral mutants of Drosophila melanogaster. III. Isolation and mapping of mutations by direct visual observations of behavioral phenotypes. Mol. Gen. Genet. 1980, 177, 553–565. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, F.; Singh, P.; Sharma, A. A Drosophila systems model of pentylenetetrazole induced locomotor plasticity responsive to antiepileptic drugs. BMC Syst. Biol. 2009, 3, 11. [Google Scholar] [CrossRef]
- Trotta, N.; Rodesch, C.K.; Fergestad, T.; Broadie, K. Cellular bases of activity-dependent paralysis in Drosophila stress-sensitive mutants. J. Neurobiol. 2004, 60, 328–347. [Google Scholar] [CrossRef]
- Kim, E.; Goraksha-Hicks, P.; Li, L.; Neufeld, T.P.; Guan, K.L. Regulation of TORC1 by Rag GTPases in nutrient response. Nat. Cell Biol. 2008, 10, 935–945. [Google Scholar] [CrossRef]
- Semrau, S.; Holtzer, L.; González-Gaitán, M.; Schmidt, T. Quantification of biological interactions with particle image cross-correlation spectroscopy (PICCS). Biophys. J. 2011, 100, 1810–1818. [Google Scholar] [CrossRef]
- Stuart, L.M.; Boulais, J.; Charriere, G.M.; Hennessy, E.J.; Brunet, S.; Jutras, I.; Goyette, G.; Rondeau, C.; Letarte, S.; Huang, H.; et al. A systems biology analysis of the Drosophila phagosome. Nature 2007, 445, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Robinson, J.E.; Joiner, W.J. SLEEPLESS is a bifunctional regulator of excitability and cholinergic synaptic transmission. Curr. Biol. 2014, 24, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Roman, G. Presynaptic inhibition of gamma lobe neurons is required for olfactory learning in Drosophila. Curr. Biol. 2013, 23, 2519–2527. [Google Scholar] [CrossRef]
- Ninan, P.T. The functional anatomy, neurochemistry, and pharmacology of anxiety. J. Clin. Psychiatry 1999, 60 (Suppl. 22), 12–17. [Google Scholar]
- Bliwise, D.L. Sleep in normal aging and dementia. Sleep 1993, 16, 40–81. [Google Scholar] [CrossRef]
- Edwards, B.A.; O’Driscoll, D.M.; Ali, A.; Jordan, A.S.; Trinder, J.; Malhotra, A. Aging and sleep: Physiology and pathophysiology. Semin. Respir. Crit. Care. Med 2010, 31, 618–633. [Google Scholar] [CrossRef] [PubMed]
- Mander, B.A.; Winer, J.R.; Walker, M.P. Sleep and human aging. Neuron 2017, 94, 19–36. [Google Scholar] [CrossRef]
- Morrow, A.L.; Pace, J.R.; Purdy, R.H.; Paul, S.M. Characterization of steroid interactions with gamma-aminobutyric acid receptor-gated chloride ion channels: Evidence for multiple steroid recognition sites. Mol. Pharmacol. 1990, 37, 263–270. [Google Scholar] [PubMed]
- Burchinsky, S.G. Neurotransmitter receptors in the central nervous system and aging: Pharmacological aspect (review). Exp. Gerontol. 1984, 19, 227–239. [Google Scholar] [CrossRef]
- Rissman, R.A.; De Blas, A.L.; Armstrong, D.M. GABA(A) receptors in aging and Alzheimer’s disease. J. Neurochem. 2007, 103, 1285–1292. [Google Scholar] [CrossRef]
- Mhatre, M.C.; Ticku, M.K. Aging related alterations in GABAA receptor subunit mRNA levels in Fischer rats. Brain Res. Mol. Brain Res. 1992, 14, 71–78. [Google Scholar] [CrossRef]
- Bruni, O.; Ferri, R.; Miano, S.; Verrillo, E. L -5-Hydroxytryptophan treatment of sleep terrors in children. Eur. J. Pediatr. 2004, 163, 402–407. [Google Scholar] [CrossRef]
- Touret, M.; Sarda, N.; Gharib, A.; Geffard, M.; Jouvet, M. The role of 5-hydroxytryptophan (5-HTP) in the regulation of the sleep/wake cycle in parachlorophenylalanine (p-CPA) pretreated rat: A multiple approach study. Exp. Brain Res. 1991, 86, 117–124. [Google Scholar] [CrossRef]
- Christen, S.; Peterhans, E.; Stocker, R. Antioxidant activities of some tryptophan metabolites: Possible implication for inflammatory diseases. Proc. Natl. Acad. Sci. USA 1990, 87, 2506–2510. [Google Scholar] [CrossRef]
- Keithahn, C.; Lerchl, A. 5-hydroxytryptophan is a more potent in vitro hydroxyl radical scavenger than melatonin or vitamin C. J. Pineal Res. 2005, 38, 62–66. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Demchak, B.; Hull, T.; Reich, M.; Liefeld, T.; Smoot, M.; Ideker, T.; Mesirov, J.P. Cytoscape: The network visualization tool for GenomeSpace workflows. F1000Research 2014, 3, 151. [Google Scholar] [CrossRef] [PubMed]
- Montojo, J.; Zuberi, K.; Rodriguez, H.; Bader, G.D.; Morris, Q. GeneMANIA: Fast gene network construction and function prediction for Cytoscape. F1000Research 2014, 3, 153. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Differentially Expressed Genes in Drosophila after GABA/5-HTP Treatment | |||
|---|---|---|---|
| Categories | Number of Transcripts | ||
| Total | 29,904 | ||
| Filtered gene | 27,191 | ||
| |fold|≥ 4 | 101 | Up | 56 |
| Down | 45 | ||
| |fold|≥ 2 | 646 | Up | 342 |
| Down | 304 | ||
| |Fold Change| ≥ 4 (DAVID Functional Analysis) | |||
| GO Term | Gene Count | p-Value | GO Accession |
| nucleolus | 7 | 2.07 × 10−0.5 | GO:0005730 |
| nuclear lumen | 7 | 8.30 × 10−3 | GO:0031981 |
| intracellular organelle lumen | 8 | 1.27 × 10−2 | GO:0070013 |
| organelle lumen | 8 | 1.27 × 10−2 | GO:0043233 |
| membrane-enclosed lumen | 8 | 1.46 × 10−2 | GO:0031974 |
| |Fold Change| ≥ 2 (DAVID Functional Analysis) | |||
| GO Term | Gene Count | p-Value | GO Accession |
| cell cortex | 7 | 0.01 | GO:0005938 |
| nucleolus | 8 | 0.04 | GO:0005730 |
| |Fold Change| ≥ 4 (DAVID Functional Analysis) | |||
| GO Term | Gene Count | p-Value | GO Accession |
| phagocytosis, engulfment | 4 | 0.01 | GO:0006911 |
| phagocytosis | 4 | 0.01 | GO:0006909 |
| endocytosis | 4 | 0.03 | GO:0006897 |
| membrane invagination | 4 | 0.03 | GO:0010324 |
| membrane organization | 4 | 0.04 | GO:0016044 |
| |Fold Change| ≥ 2 (DAVID Functional Analysis) | |||
| GO Term | Gene Count | p-Value | GO Accession |
| cellular ion homeostasis | 7 | 2.21 × 10−4 | GO:0006873 |
| ion homeostasis | 7 | 4.51 × 10−4 | GO:0050801 |
| regulation of membrane potential | 5 | 4.75 × 10−4 | GO:0042391 |
| cellular chemical homeostasis | 7 | 5.03 × 10−4 | GO:0055082 |
| chemical homeostasis | 7 | 0.00 | GO:0048878 |
| cellular homeostasis | 8 | 0.01 | GO:0019725 |
| homeostatic process | 8 | 0.05 | GO:0042592 |
| response to abiotic stimulus | 11 | 0.01 | GO:0009628 |
| visual behavior | 3 | 0.04 | GO:0007632 |
| positive regulation of cellular component organization | 6 | 1.42 × 10−4 | GO:0051130 |
| regulation of cellular component biogenesis | 6 | 0.02 | GO:0044087 |
| glutamine family amino acid catabolic process | 3 | 0.01 | GO:0009065 |
| glutamate metabolic process | 3 | 0.02 | GO:0006536 |
| cellular amino acid catabolic process | 4 | 0.03 | GO:0009063 |
| amine catabolic process | 4 | 0.03 | GO:0009310 |
| glycerol metabolic process | 3 | 0.05 | GO:0006071 |
| alditol metabolic process | 3 | 0.05 | GO:0019400 |
| response to abiotic stimulus | 11 | 0.01 | GO:0009628 |
| phosphate metabolic process | 21 | 0.01 | GO:0006796 |
| phosphorus metabolic process | 21 | 0.01 | GO:0006793 |
| phosphorylation | 15 | 0.05 | GO:0016310 |
| regulation of cytoskeleton organization | 5 | 0.02 | GO:0051493 |
| regulation of organelle organization | 7 | 0.02 | GO:0033043 |
| membrane organization | 13 | 0.03 | GO:0016044 |
| membrane invagination | 11 | 0.03 | GO:0010324 |
| endocytosis | 11 | 0.03 | GO:0006897 |
| transmission of nerve impulse | 12 | 0.00 | GO:0019226 |
| synaptic transmission | 10 | 0.01 | GO:0007268 |
| cell–cell signaling | 10 | 0.03 | GO:0007267 |
| synaptic vesicle endocytosis | 4 | 0.03 | GO:0048488 |
| |Fold Change| ≥ 4 (DAVID Functional Analysis) | |||
| GO Term | Gene Count | p-Value | GO Accession |
| ATPase activity, coupled to transmembrane movement of substances | 3 | 0.04 | GO:0042626 |
| ATPase activity, coupled to movement of substances | 3 | 0.04 | GO:0043492 |
| hydrolase activity, acting on acid anhydrides, catalyzing transmembrane movement of substances | 3 | 0.04 | GO:0016820 |
| primary active transmembrane transporter activity | 3 | 0.05 | GO:0015399 |
| P–P-bond-hydrolysis-driven transmembrane transporter activity | 3 | 0.05 | GO:0015405 |
| hydrogen ion transmembrane transporter activity | 3 | 0.02 | GO:0015078 |
| monovalent inorganic cation transmembrane transporter activity | 3 | 0.02 | GO:0015077 |
| inorganic cation transmembrane transporter activity | 3 | 0.03 | GO:0022890 |
| |Fold Change| ≥ 2 (DAVID Functional Analysis) | |||
| GO Term | Gene Count | p-Value | GO Accession |
| transition metal ion binding | 42 | 0.01 | GO:0046914 |
| metal ion binding | 49 | 0.02 | GO:0046872 |
| cation binding | 50 | 0.02 | GO:0043169 |
| ion binding | 50 | 0.03 | GO:0043167 |
| cytoskeletal protein binding | 14 | 7.58 × 10−4 | GO:0008092 |
| actin cytoskeleton organization | 10 | 0.00 | GO:0030036 |
| actin filament-based process | 10 | 0.00 | GO:0030029 |
| actin binding | 9 | 0.00 | GO:0003779 |
| cytoskeleton organization | 17 | 0.02 | GO:0007010 |
| nucleotide binding | 37 | 0.02 | GO:0000166 |
| ATP binding | 24 | 0.05 | GO:0005524 |
| adenyl ribonucleotide binding | 24 | 0.05 | GO:0032559 |
| cytoskeletal protein binding | 14 | 7.58 × 10−4 | GO:0008092 |
| microtubule binding | 6 | 0.01 | GO:0008017 |
| tubulin binding | 6 | 0.01 | GO:0015631 |
| hydrolase activity, acting on acid anhydrides, catalyzing transmembrane movement of substances | 9 | 0.01 | GO:0016820 |
| primary active transmembrane transporter activity | 9 | 0.02 | GO:0015399 |
| P–P-bond-hydrolysis-driven transmembrane transporter activity | 9 | 0.02 | GO:0015405 |
| ATPase activity, coupled to movement of substances | 8 | 0.03 | GO:0043492 |
| ATPase activity, coupled to transmembrane movement of substances | 8 | 0.03 | GO:0042626 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, Y.; Hong, K.-B.; Kim, S.; Suh, H.J.; Jo, K. Changes in Locomotor Activity and Oxidative Stress-Related Factors after the Administration of an Amino Acid Mixture by Generation and Age. Int. J. Mol. Sci. 2021, 22, 9822. https://doi.org/10.3390/ijms22189822
Ahn Y, Hong K-B, Kim S, Suh HJ, Jo K. Changes in Locomotor Activity and Oxidative Stress-Related Factors after the Administration of an Amino Acid Mixture by Generation and Age. International Journal of Molecular Sciences. 2021; 22(18):9822. https://doi.org/10.3390/ijms22189822
Chicago/Turabian StyleAhn, Yejin, Ki-Bae Hong, Suhyeon Kim, Hyung Joo Suh, and Kyungae Jo. 2021. "Changes in Locomotor Activity and Oxidative Stress-Related Factors after the Administration of an Amino Acid Mixture by Generation and Age" International Journal of Molecular Sciences 22, no. 18: 9822. https://doi.org/10.3390/ijms22189822
APA StyleAhn, Y., Hong, K.-B., Kim, S., Suh, H. J., & Jo, K. (2021). Changes in Locomotor Activity and Oxidative Stress-Related Factors after the Administration of an Amino Acid Mixture by Generation and Age. International Journal of Molecular Sciences, 22(18), 9822. https://doi.org/10.3390/ijms22189822