
Multi-Systemic Alterations by Chronic Exposure to a Low Dose of Bisphenol A in Drinking Water: Effects on Inflammation and NAD+-Dependent Deacetylase Sirtuin1 in Lactating and Weaned Rats

 ,
, 
 ,
,  , ,
, ,  ,
,  ,
,
Abstract
1. Introduction
2. Results
2.1. Experimental Groups and Body Weight
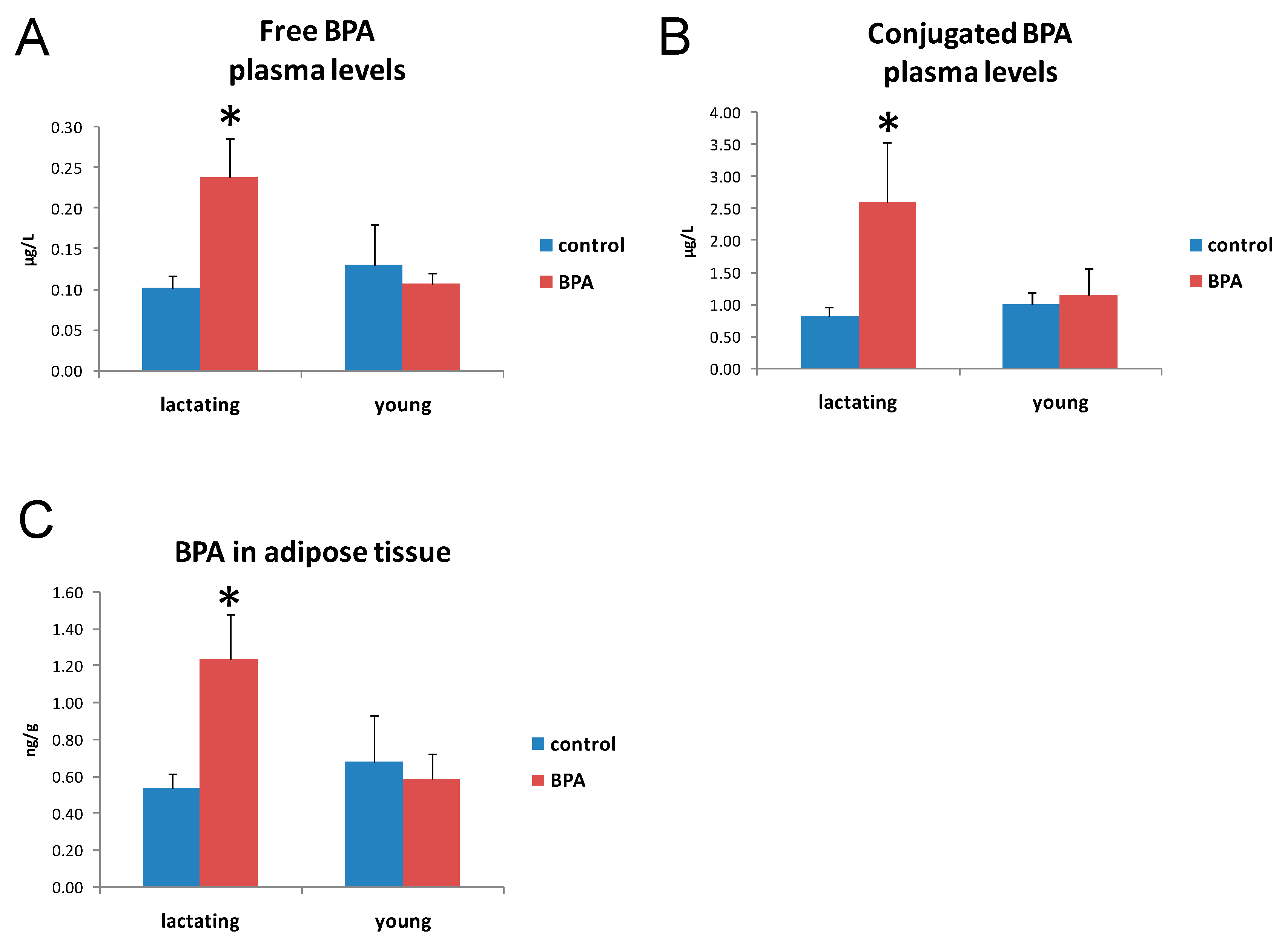
2.2. BPA Concentration in Plasma and Adipose Tissue
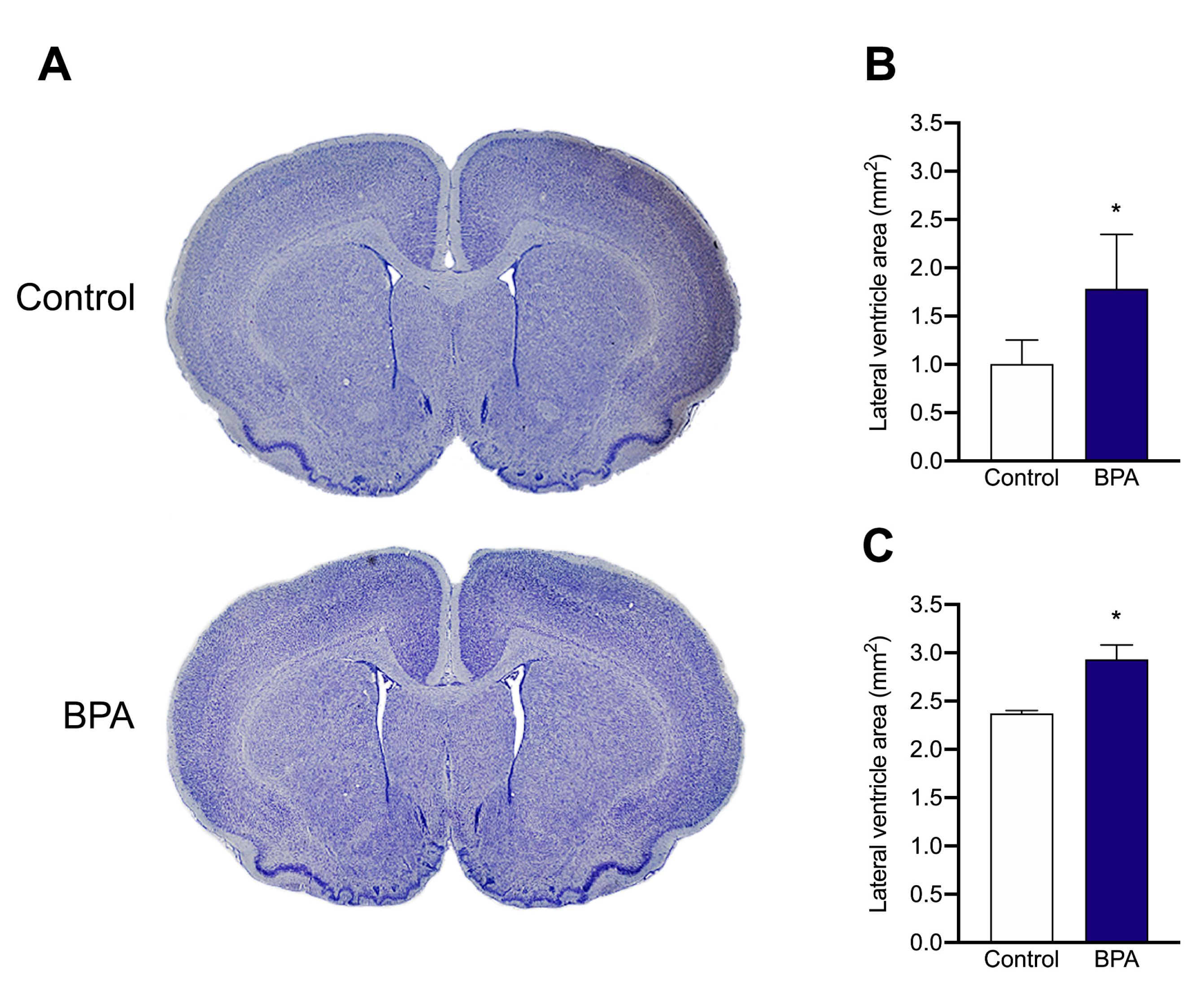
2.3. Nissl Staining and Measurement of Ventricle Size
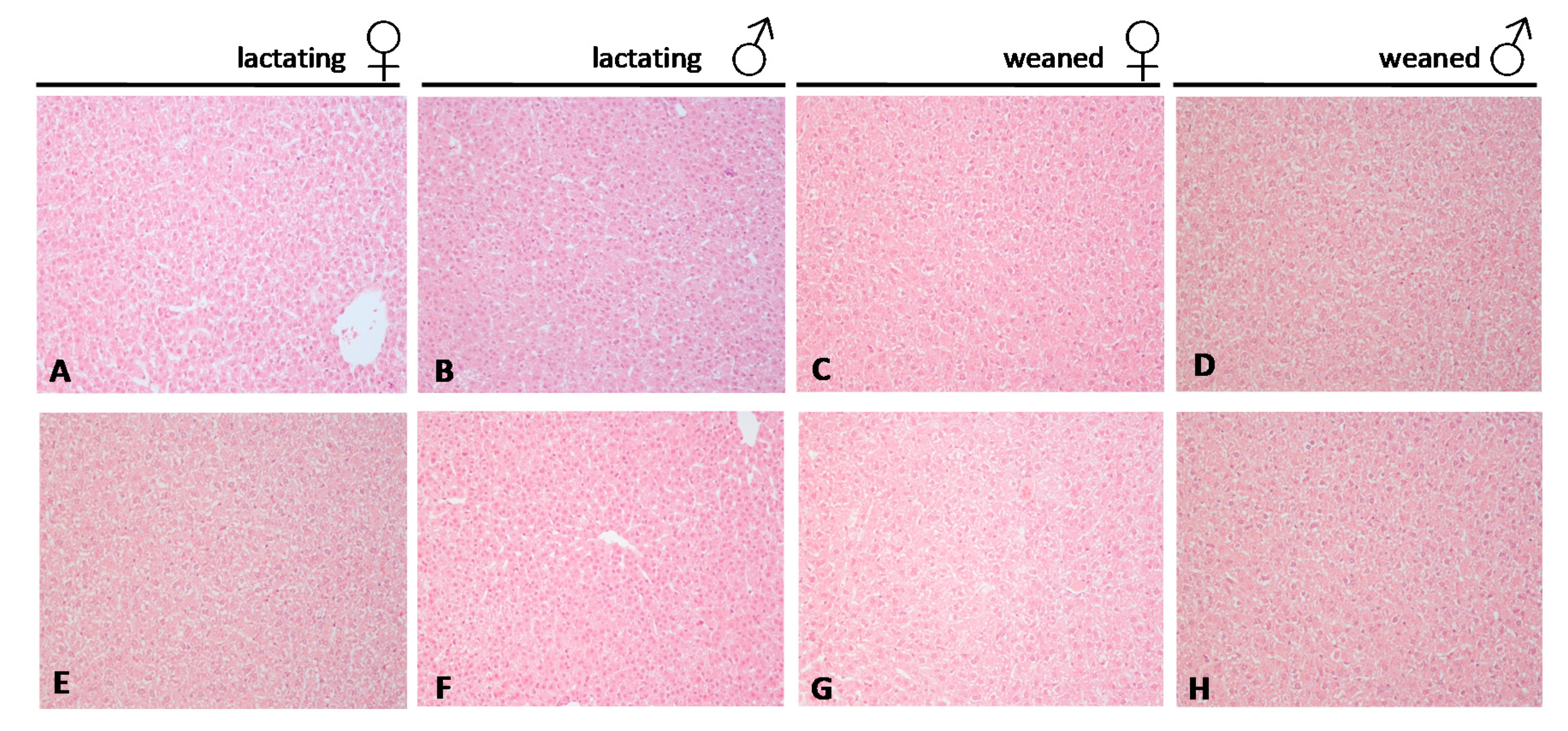
2.4. Histological Analysis of the Liver
2.5. Molecular Markers in Liver
2.5.1. Inflammation
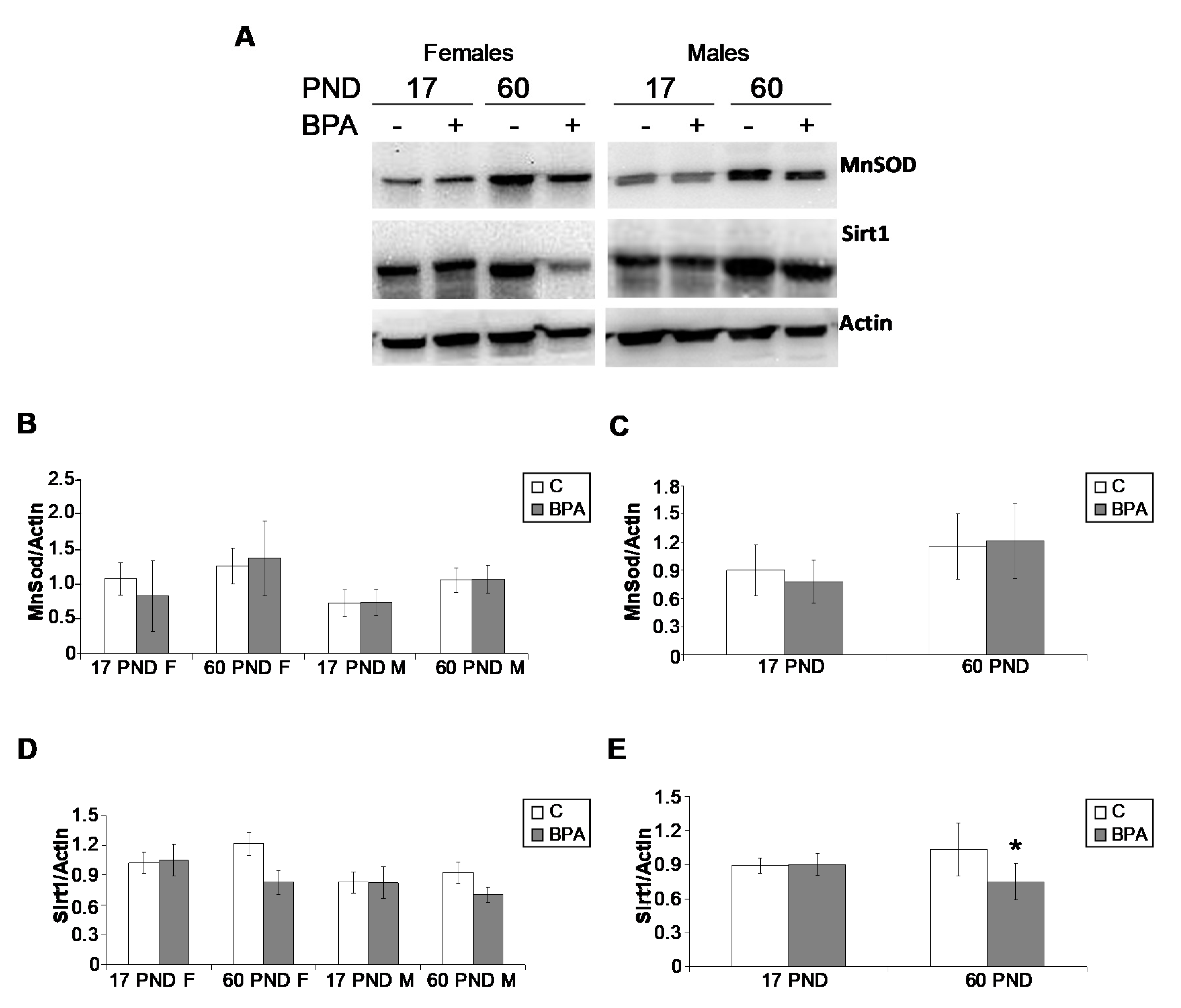
2.5.2. Oxidative Stress
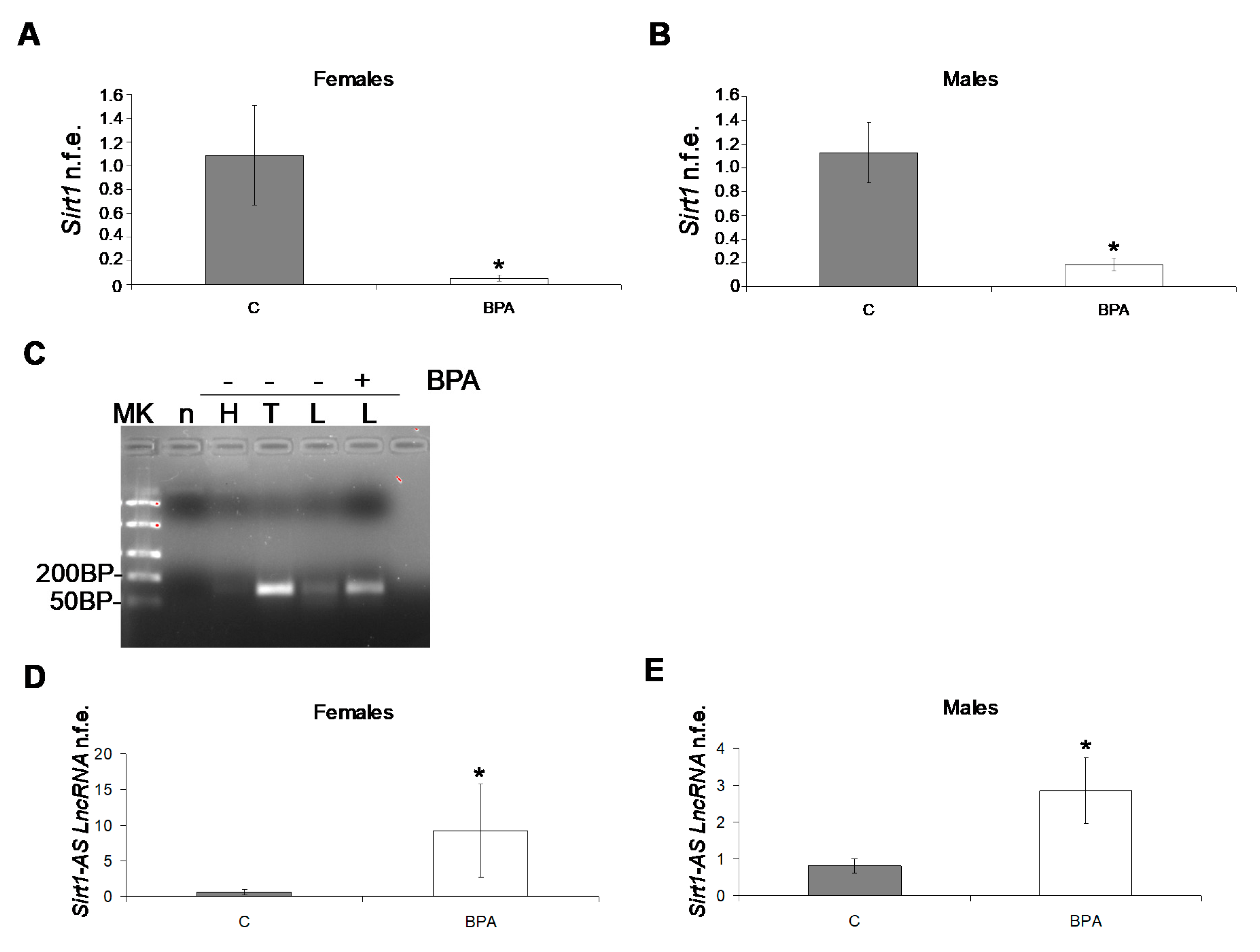
2.5.3. Epigenetics and Epigenetic Machinery
2.5.4. Lipid and Glucose Metabolism
3. Discussion
4. Materials and Methods
4.1. Chemicals and Antisera
4.2. Animals and BPA Exposure Protocol
4.3. Free and Conjugate Plasma BPA Measure
4.4. Adipose Tissue BPA Measure
4.5. Nissl Staining and Measurement of Ventricle Size
4.6. Evaluation of Liver Morphology
4.7. Protein Extraction and Western Blot
4.8. Total RNA Extraction, Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
4.9. Evaluation of Sirt1-AS LncRNA: Strand Specific PCR and Expression Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rubin, B.S. Bisphenol A: An endocrine disruptor with widespread exposure and multiple effects. J. Steroid. Biochem. Mol. Biol. 2011, 127, 27–34. [Google Scholar] [CrossRef]
- Corrales, J.; Kristofco, L.A.; Steele, W.B.; Yates, B.S.; Breed, C.S.; Williams, E.S.; Brooks, B.W. Global Assessment of Bisphenol A in the Environment: Review and Analysis of Its Occurrence and Bioaccumulation. Dose-Response Int. J. 2015, 13, 1–29. [Google Scholar] [CrossRef]
- Chianese, R.; Troisi, J.; Richards, S.; Scafuro, M.; Fasano, S.; Guida, M.; Pierantoni, R.; Meccariello, R. Bisphenol A in reproduction: Epigenetic effects. Curr. Med. Chem. 2018, 25, 748–770. [Google Scholar] [CrossRef]
- Murata, M.; Kang, J.H. Bisphenol A (BPA) and cell signaling pathways. Biotechnol. Adv. 2018, 36, 311–327. [Google Scholar] [CrossRef]
- Santoro, A.; Chianese, R.; Troisi, J.; Richards, S.; Nori, S.; Fasano, S.; Guida, M.; Plunk, E.; Viggiano, A.; Pierantoni, R.; et al. Neuro-toxic and reproductiveeffects of BPA. Curr. Neuropharmacol. 2019, 17, 1109–1132. [Google Scholar] [CrossRef]
- Tavares, R.S.; Escada-Rebelo, S.; Correia, M.; Mota, P.C.; Ramalho-Santos, J. The non-genomic effects of endocrine-disrupting chemicals on mammalian sperm. Reproduction 2016, 151, R1–R13. [Google Scholar] [CrossRef] [PubMed]
- Peretz, J.; Vrooman, L.; Ricke, W.A.; Hunt, P.A.; Ehrlich, S.; Hauser, R.; Padmanabhan, V.; Taylor, H.S.; Swan, S.H.; VandeVoort, C.A.; et al. Bisphenol A and Reproductive Health: Update of Experimental and Human Evidence. Environ. Health Perspect. 2007, 8, 775–778. [Google Scholar]
- Vandenberg, L.N.; Ehrlich, S.; Belcher, S.M.; BenJonathan, N.; Dolinoy, D.C.; Hugo, E.R.; Hunt, P.A.; Newbold, R.R.; Rubin, B.S.; Saili, K.S.; et al. Low dose effects of Bisphenol A: An integrated review of in vitro, laboratory animal and epidemiology studies. Endocr. Disrupt. 2013, 1, e2507. [Google Scholar] [CrossRef]
- Le Magueresse-Battistoni, B.; Multigner, L.; Beausoleil, C.; Rousselle, C. Effects of bisphenol A on metabolism and evidences of a mode of action mediated through endocrine disruption. Mol. Cell. Endocrinol. 2018, 475, 74–91. [Google Scholar] [CrossRef]
- D’Angelo, S.; Scafuro, M.; Meccariello, R. BPA and nutraceuticals, simultaneous effects on endocrine functions. Endocr. Metab. Immune. Disord. Drug Targets 2019, 19, 594–604. [Google Scholar] [CrossRef]
- Genuis, S.J.; Beesoon, S.; Birkholz, D.; Lobo, R.A. Human Excretion of Bisphenol A: Blood, Urine, and Sweat (BUS) Study. J. Environ. Public Health 2012, 2012, 185731. [Google Scholar] [CrossRef]
- Nunez, A.A.; Kannan, K.; Giesy, J.P.; Fang, J.; Clemens, L.G. Effects of bisphenol A on energy balance and accumulation in brown adipose tissue in rats. Chemosphere 2001, 42, 917–922. [Google Scholar] [CrossRef]
- Mercogliano, R.; Santonicola, S. Investigation on bisphenol A levels in human milk and dairy supply chain: A review. Food Chem. Toxicol. 2018, 114, 98–107. [Google Scholar] [CrossRef]
- Dualde, P.; Pardo, O.; Corpas-Burgos, F.; Kuligowski, J.; Gormaz, M.; Vento, M.; Pastor, A.; Yusà, V. Biomonitoring of bisphenols A, F, S in human milk and probabilistic risk assessment for breastfed infants. Sci. Total Environ. 2019, 668, 797–805. [Google Scholar] [CrossRef]
- Mørck, T.J.; Sorda, G.; Bechi, N.; Rasmussen, B.S.; Nielsen, J.B.; Ietta, F.; Rytting, E.; Mathiesen, L.; Paulesu, L.; Knudsen, L.E. Placental transport and in vitro effects of Bisphenol A. Reprod. Toxicol. 2010, 30, 131–137. [Google Scholar] [CrossRef]
- Corbel, T.; Gayrard, V.; Puel, S.; Lacroix, M.Z.; Berrebi, A.; Gil, S.; Viguie, C.; Toutain, P.L.; Picard-Hagen, N. Bidirectional placental transfer of Bisphenol A and its main metabolite, Bisphenol A Glucuronide, in the isolated perfused human placenta. Reprod. Toxicol. 2014, 47, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Troisi, J.; Giugliano, L.; D’Antonio, A.; Viggiano, A.; Meccariello, R.; Scafuro, M.; Monda, M.; Colucci, A.; Scala, G.; Cofano, M.; et al. Placental Vascularization and Apoptosis in Rats Orally Exposed to Low Doses of Bisphenol A. Open J. Obstet. Gynecol. 2018, 8, 958–969. [Google Scholar] [CrossRef][Green Version]
- Roepke, T.A.; Yang, J.A.; Yasrebi, A.; Mamounis, K.J.; Oruc, E.; Zama, A.M.; Uzumcu, M. Regulation of arcuate genes by devel- opmental exposures to endocrine-disrupting compounds in female rats. Reprod. Toxicol. 2016, 62, 18–26. [Google Scholar] [CrossRef]
- Desai, M.; Ferrini, M.G.; Han, G.; Jellyman, J.K.; Ross, M.G. In vivo maternal and in vitro BPA exposure effects on hypothalamic neurogenesis and appetite regulators. Environ. Res. 2018, 164, 45–52. [Google Scholar] [CrossRef]
- Di Pietro, P.; D’Auria, R.; Viggiano, A.; Ciaglia, E.; Meccariello, R.; Dello Russo, R.; Puca, A.A.; Vecchione, C.; Nori, S.L.; Santoro, A. Bisphenol A induces DNA damage in cells exerting immunosurveillance functions at peripheral and central level by differentially modulating estrogen receptors expression. Chemosphere 2020, 254, 126819. [Google Scholar] [CrossRef]
- Eid, J.I.; Eissa, S.M.; El-Ghor, A.A. Bisphenol A induces oxidative stress and DNA damage in hepatic tissue of female rat offspring. JOBAZ 2015, 71, 10–19. [Google Scholar] [CrossRef]
- Hugo, E.R.; Brandebourg, T.D.; Woo, J.G.; Loftus, J.; Alexander, J.W.; Ben-Jonathan, N. Bisphenol A at environmentally relevant doses inhibits adiponectin release from human adipose tissue explants and adipocytes. Environ. Health Perspect. 2008, 116, 1642–1647. [Google Scholar] [CrossRef] [PubMed]
- Ben-Jonathan, N.; Hugo, E.R.; Brandebourg, T.D. Effects of bisphenol A on adipokine release from human adipose tissue: Implications for the metabolic syndrome. Mol. Cell. Endocrinol. 2009, 304, 49–54. [Google Scholar] [CrossRef]
- Huc, L.; Lemarié, A.; Guéraud, F.; Héliès-Toussaint, C. Low concentrations of bisphenol A induce lipid accumulation mediated by the production of reactive oxygen species in the mitochondria of HepG2 cells. Toxicol. In Vitro 2012, 26, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Ke, Z.H.; Pan, J.X.; Jin, L.Y.; Xu, H.Y.; Yu, T.T.; Ullah, K.; Rahman, T.U.; Ren, J.; Cheng, Y.; Dong, X.Y.; et al. Bisphenol A Exposure May Induce Hepatic Lipid Accumulation via Reprogramming the DNA Methylation Patterns of Genes Involved in Lipid Metabolism. Sci. Rep. 2016, 6, 31331. [Google Scholar] [CrossRef] [PubMed]
- Legeay, S.; Faure, S. Is bisphenol A an environmental obesogen? Fundam. Clin. Pharmacol. 2017, 6, 594–609. [Google Scholar] [CrossRef] [PubMed]
- Chianese, R.; Viggiano, A.; Urbanek, K.; Cappetta, D.; Troisi, J.; Scafuro, M.; Guida, M.; Esposito, G.; Ciuffreda, L.P.; Rossi, F.; et al. Chronic exposure to low dose of bisphenol A impacts the first round of spermatogenesis via SIRT1 modulation. Sci. Rep. 2018, 8, 2961. [Google Scholar] [CrossRef]
- Zhang, T.; Kraus, W.L. SIRT1-dependent Regulation of Chromatin and Transcription: Linking NAD+ Metabolism and Signaling to the Control of Cellular Functions. Biochim. Biophys. Acta 2010, 1804, 1666–1675. [Google Scholar] [CrossRef]
- D’Angelo, S.; Mele, E.; Di Filippo, F.; Viggiano, A.; Meccariello, R. Sirt1 activity in the brain: Simultaneous effects on energy homeostasis and reproduction. Int. J. Environ. Res. Public Health 2021, 18, 1243. [Google Scholar] [CrossRef]
- Jing, H.; Lin, H. Sirtuins in Epigenetic Regulation. Chem. Rev. 2015, 115, 2350–2375. [Google Scholar] [CrossRef]
- Elibol, B.; Kilic, U. High Levels of SIRT1 Expression as a Protective Mechanism Against Disease-Related Conditions. Front. Endocrinol. 2018, 9, 614. [Google Scholar] [CrossRef]
- Scisciola, L.; Sarno, F.; Carafa, V.; Cosconati, S.; Di Maro, S.; Ciuffreda, L.; De Angelis, A.; Stiuso, P.; Feoli, A.; Sbardella, G.; et al. Two novel SIRT1 activators, SCIC2 and SCIC2.1, enhance SIRT1-mediated effects in stress response and senescence. Epigenetics 2020, 15, 664–683. [Google Scholar] [CrossRef] [PubMed]
- Piegari, E.; Cozzolino, A.; Ciuffreda, L.P.; Cappetta, D.; De Angelis, A.; Urbanek, K.; Rossi, F.; Berrino, L. Cardioprotective effects of miR-34a silencing in a rat model of doxorubicin toxicity. Sci. Rep. 2020, 10, 12250. [Google Scholar] [CrossRef] [PubMed]
- Piegari, E.; Russo, R.; Cappetta, D.; Esposito, G.; Urbanek, K.; Dell’Aversana, C.; Altucci, L.; Berrino, L.; Rossi, F.; De Angelis, A. MicroRNA-34a regulates doxorubicin-induced cardiotoxicity in rat. Oncotarget 2016, 7, 62312–62326. [Google Scholar] [CrossRef]
- De Angelis, A.; Piegari, E.; Cappetta, D.; Russo, R.; Esposito, G.; Ciuffreda, L.P.; Ferraiolo, F.A.; Frati, C.; Fagnoni, F.; Berrino, L.; et al. SIRT1 activation rescues doxorubicin-induced loss of functional competence of human cardiac progenitor cells. Int. J. Cardiol. 2015, 189, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Li, X. SIRT1 and energy metabolism. Acta Biochim. Biophys. Sin. 2013, 45, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Ding, R.B.; Bao, J.; Deng, C.X. Emerging roles of SIRT1 in fatty liver diseases. Int. J. Biol. Sci. 2017, 13, 852–867. [Google Scholar] [CrossRef]
- Wang, Y.; Pang, W.J.; Wei, N.; Xiong, Y.; Wu, W.J.; Zhao, C.Z.; Shen, Q.W.; Yang, G.S. Identification, stability and expression of Sirt1 antisense long non-coding RNA. Gene 2014, 539, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.Q.; Wang, Y.; Xiong, Y.; Chen, X.C.; Ma, M.L.; Cai, R.; Gao, Y.; Sun, Y.M.; Yang, G.S.; Pang, W.J. Sirt1 AS lncRNA interacts with its mRNA to inhibit muscle formation by attenuating function of miR-34a. Sci. Rep. 2016, 6, 21865. [Google Scholar] [CrossRef]
- Liu, J.; Wu, W.; Jin, J. A novel mutation in SIRT1-AS leading to a decreased risk of HCC. Oncol. Rep. 2015, 34, 2343–2350. [Google Scholar] [CrossRef] [PubMed]
- Calafat, A.M.; Ye, X.; Wong, L.Y.; Reidy, J.A.; Needham, L.L. Exposure of the U.S. population to bisphenol A and 4-tertiaryoctylphenol: 2003–2004. Environ. Health Perspect. 2008, 116, 39–44. [Google Scholar] [CrossRef]
- Trasande, L.; Attina, T.M.; Blustein, J. Association between urinary bisphenol A concentration and obesity prevalence in children and adolescents. JAMA 2012, 308, 1113–1121. [Google Scholar] [CrossRef]
- Do, M.T.; Chang, V.C.; Mendez, M.A.; de Groh, M. Urinary bisphenol A and obesity in adults: Results from the Canadian Health Measures Survey. Health Promot. Chronic Dis. Prev. Can. 2017, 37, 403–412. [Google Scholar] [CrossRef]
- Chevalier, N.; Fénichel, P. Bisphenol A: Targeting metabolic tissues. Rev. Endocr. Metab. Disord. 2015, 16, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Hunt, P.A.; Gore, A.C. Endocrine disruptors and the future of toxicology testing-lessons from CLARITY-BPA. Nat. Rev. Endocrinol. 2019, 15, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Commission Implementing Regulation (EU) No 321/2011 of 1 April 2011 Amending Regulation (EU) No 10/2011 as Regards the Restriction of Use of Bisphenol A in Plastic Infant Feeding Bottles Text with EEA Relevance. 2011. Available online: http://data.europa.eu/eli/reg_impl/2011/321/oj (accessed on 8 August 2021).
- Andra, S.S.; Charisiadis, P.; Arora, M.; van Vliet-Ostaptchouk, J.V.; Makris, K.C. Biomonitoring of human exposures to chlorinated derivatives and structural analogs of bisphenol A. Environ. Int. 2015, 85, 352–379. [Google Scholar] [CrossRef]
- Ullah, A.; Pirzada, M.; Jahan, S.; Ullah, H.; Shaheen, G.; Rehman, H.; Siddiqui, M.F.; Butt, M.A. Bisphenol A and its analogs bisphenol B, bisphenol F, and bisphenol S: Comparative in vitro and in vivo studies on the sperms and testicular tissues of rats. Chemosphere 2018, 209, 508–516. [Google Scholar] [CrossRef]
- Rochester, J.R.; Bolden, A.L. Bisphenol S and F: A Systematic.Review and Comparison of the Hormonal Activity of Bisphenol A Substitutes. Environ. Health Perspect. 2015, 123, 643–650. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Food Contact Materials, Enzymes, Flavourings and Processing Aids (CEF). Scientific Opinion on the risks to public health related to the presence of bisphenol A (BPA) in foodstuffs. EFSA J. 2015, 13, 3978. [Google Scholar] [CrossRef]
- EFSA. A statement on the developmental immunotoxicity ofbisphenol A (BPA): Answer to the question from the Dutch Minis- try of Health, Welfare and Sport. EFSA J. 2016, 14, 4580. [Google Scholar]
- Nachman, R.M.; Hartle, J.C.; Lees, P.S.; Groopman, J.D. Early Life Metabolism of Bisphenol A: A Systematic Review of the Literature. Curr. Environ. Health Rep. 2014, 1, 90–100. [Google Scholar] [CrossRef]
- Negri-Cesi, P. Bisphenol A Interaction with Brain Development and Functions. Dose Response 2015, 13, 1559325815590394. [Google Scholar] [CrossRef]
- Tumani, H.; Huss, A.; Bachhuber, F. The cerebrospinal fluid and barriers-anatomic and physiologic considerations. Handb. Clin. Neurol. 2017, 146, 21–32. [Google Scholar] [PubMed]
- Ghersi-Egea, J.F.; Strazielle, N.; Catala, M.; Silva-Vargas, V.; Doetsch, F.; Engelhardt, B. Molecular anatomy and functions of the choroidal blood-cerebrospinal fluid barrier in health and disease. Acta Neuropathol. 2018, 135, 337–361. [Google Scholar] [CrossRef]
- Santoro, A.; Spinelli, C.C.; Martucciello, S.; Nori, S.L.; Capunzo, M.; Puca, A.A.; Ciaglia, E. Innate immunity and cellular senescence: The good and the bad in the developmental and aged brain. J. Leukoc. Biol. 2018, 103, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Scelsi, C.L.; Rahim, T.A.; Morris, J.A.; Kramer, G.J.; Gilbert, B.C.; Forseen, S.E. The Lateral Ventricles: A Detailed Review of Anatomy, Development, and Anatomic Variations. Am. J. Neuroradiol. 2020, 41, 566–572. [Google Scholar] [CrossRef]
- Paquette, N.; Shi, J.; Wang, Y.; Lao, Y.; Ceschin, R.; Nelson, M.D.; Panigrahy, A.; Lepore, N. Ventricular shape and relative position abnormalities in preterm neonates. Neuroimage Clin. 2017, 15, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yan, C.; Zhang, J.; Lin, R.; Lin, Q.; Yang, L.; Ren, F.; Zhang, J.; Ji, M.; Li, Y. SIRT1 inhibits TNF-α-induced apoptosis of vascular adventitial fibroblasts partly through the deacetylation of FoxO1. Apoptosis 2013, 18, 689–701. [Google Scholar] [CrossRef]
- Tang, L.; Chen, Q.; Meng, Z.; Sun, L.; Zhu, L.; Liu, J.; Hu, J.; Ni, Z.; Wang, X. Suppression of Sirtuin-1 Increases IL-6 Expression by Activation of the Akt Pathway During Allergic Asthma. Cell Physiol. Biochem. 2017, 43, 1950–1960. [Google Scholar] [CrossRef]
- Kauppinen, A.; Suuronen, T.; Ojala, J.; Kaarniranta, K.; Salminen, A. Antagonistic crosstalk between NF-κB and SIRT1 in the regulation of inflammation and metabolic disorders. Cell Signal. 2013, 25, 1939–1948. [Google Scholar] [CrossRef] [PubMed]
- Khorkova, O.; Myers, A.J.; Hsiao, J.; Wahlestedt, C. Natural antisense transcripts. Hum. Mol. Genet. 2014, 23, R54–R63. [Google Scholar] [CrossRef]
- Qian, W.; Cai, X.; Qian, Q. Sirt1 antisense long non-coding RNA attenuates pulmonary fibrosis through sirt1-mediated epithelial-mesenchymal transition. Aging 2020, 12, 4322–4336. [Google Scholar] [CrossRef] [PubMed]
- Mokhberian, N.; Hashemi, S.M.; Jajarmi, V.; Eftekhary, M.; Koochaki, A.; Ghanbarian, H. Sirt1 antisense transcript is down-regulated in human tumors. Mol. Biol. Rep. 2019, 46, 2299–2305. [Google Scholar] [CrossRef]
- Ma, Y.; Xia, W.; Wang, D.Q.; Wan, Y.J.; Xu, B.; Chen, X.; Li, Y.Y.; Xu, S.Q. Hepatic DNA methylation modifications in early development of rats resulting from perinatal BPA exposure contribute to insulin resistance in adulthood. Diabetologia 2013, 56, 2059–2067. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, H.; Zou, J.; Mai, H.; Su, D.; Feng, X.; Feng, D. Bisphenol A exposure induces cholesterol synthesis and hepatic steatosis in C57BL/6 mice by down-regulating the DNA methylation levels of SREBP-2. Food Chem. Toxicol. 2019, 133, 110786. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Yan, B.; Liao, D.; Huang, S.; Qiu, Y. Acetylation of HDAC1 and degradation of SIRT1 form a positive feedback loop to regulate p53 acetylation during heat-shock stress. Cell Death. Dis. 2015, 6, e1747. [Google Scholar] [CrossRef] [PubMed]
- Erion, D.M.; Yonemitsu, S.; Nie, Y.; Nagai, Y.; Gillum, M.P.; Hsiao, J.J.; Iwasaki, T.; Stark, R.; Weismann, D.; Yu, X.X.; et al. SirT1 knockdown in liver decreases basal hepatic glucose production and increases hepatic insulin responsiveness in diabetic rats. Proc. Natl. Acad. Sci. USA 2009, 106, 11288–11293. [Google Scholar] [CrossRef]
- Li, X.; Zhang, S.; Blander, G.; Tse, J.G.; Krieger, M.; Guarente, L. SIRT1 deacetylates and positively regulates the nuclear receptor LXR. Mol. Cell 2007, 28, 91–106. [Google Scholar] [CrossRef]
- Errico, S.; Portaccio, M.; Nicolucci, C.; Meccariello, R.; Chianese, R.; Scafuro, M.; Lepore, M.; Diano, N. Complementary experimental approaches for liver analysis in BPA exposed rats. J. Pharm. Biomed. Anal. 2019, 165, 207–212. [Google Scholar] [CrossRef]
- Kosarac, I.; Kubwabo, C.; Lalonde, K.; Foster, W. A novel method for the quantitative determination of free and conjugated bisphenol A in human maternal and umbilical cord blood serum using a two-step solid phase extraction and gas chromatography/tandem mass spectrometry. J. Chromatog. B 2012, 898, 90–94. [Google Scholar] [CrossRef]
- Guida, M.; Troisi, J.; Ciccone, C.; Granozio, G.; Cosimato, C.; Sardo, A.D.S.; Ferrara, C.; Guida, M.; Nappi, C.; Zullo, F. Bisphenol A and congenital developmental defects in humans. Mutat. Res. /Fundam. Mol. Mech. Mutagenesis 2015, 774, 33–39. [Google Scholar] [CrossRef]
- Landolfi, A.; Troisi, J.; Savanelli, M.C.; Vitale, C.; Barone, P.; Amboni, M. Bisphenol A glucuronidation in patients with Parkinson’s disease. Neurotoxicology 2017, 63, 90–96. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press: San Diego, CA, USA, 2007. [Google Scholar]
- Annunziata, C.; Lama, A.; Pirozzi, C.; Cavaliere, G.; Trinchese, G.; Di Guida, F.; Nitrato Izzo, A.; Cimmino, F.; Paciello, O.; De Biase, D.; et al. Palmitoylethanolamide counteracts hepatic metabolic inflexibility modulating mitochondrial function and efficiency in diet-induced obese mice. FASEB J. 2020, 34, 350–364. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Menke, A.L.; Driessen, A.; Koek, G.H.; Lindeman, J.H.; Stoop, R.; Havekes, L.M.; Kleemann, R.; van den Hoek, A.M. Establishment of a General NAFLD Scoring System for Rodent Models and Comparison to Human Liver Pathology. PLoS ONE 2014, 9, e115922. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosenbrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n. of Subjects | Weight (g) Mean ± SE | ||||
|---|---|---|---|---|---|
| Treatment | Males | Females | Males | Females | |
| Lactating | Control | 6 | 5 | 34 ± 4 | 35 ± 4 |
| BPA | 7 | 6 | 29 ± 4 | 27 ± 4 | |
| Weaned | Control | 9 | 7 | 220 ± 14 | 155 ± 10 |
| BPA | 10 | 9 | 232 ± 11 | 150 ± 12 | |
| Macrovesicular Steatosis | Microvesicular Steatosis | Hypertrophy | Inflammation | ||
|---|---|---|---|---|---|
| Lactating | Control | 0.09 ± 0.10 | * 0.45 ± 0.22 (p < 0.05) | 0.00 ± 0.00 | 0.18 ± 0.13 |
| BPA | 0.00 ± 0.00 | 1.47 ± 0.26 | 0.33 ± 0.13 | 0.13 ± 0.09 | |
| Young | Control | 0.00 ± 0.00 | 1.00 ± 0.35 | 0.00 ± 0.00 | 0.33 ± 0.18 |
| BPA | 0.00 ± 0.00 | 1.53 ± 0.23 | 0.26 ± 0.11 | 0.00 ± 0.00 |
| 17 PND | 60 PND | |||
|---|---|---|---|---|
| mRNA | Control (n = 4–6) | BPA (n = 4–6) | Control (n = 4–6) | BPA (n = 4–6) |
| Tnfα | 1.031 ± 0.29 | 16.76 ± 1.071 * (p = 0.042) | 1.026 ± 0.27 | 2.54 ± 1.09 * (p = 0.0043) |
| Il-6 | 1.079 ± 0.62 | 14.99 ± 5.6 * (p = 0.041) | 1.094 ± 0.48 | 1.071 ± 0.98 (p = 0.87) |
| Nf-kb | 1.15 ± 0,30 | 10,68 ± 4.1 * (p = 0.043) | 1.19 ± 0.4 | 2.31 ± 0.93 * (p = 0.012) |
| mRNA | 60 PND Females | 60 PND Males | ||
|---|---|---|---|---|
| Control (n = 3) | BPA (n = 3) | Control (n = 3) | BPA (n = 3) | |
| Dnmt1 | 1.110 ± 0.65 | 0.416 ± 0.36 (p = 0.181) | 1.164 ± 0.73 | 0.834 ± 0.42 (p = 0.359) |
| Dnmt3a | 1.145 ± 0.768 | 0.601 ± 0.57 (p = 0.387) | 1.039 ± 0.52 | 1.049 ± 0.35 (p = 0.985) |
| Hdac1 | 1.177 ± 0.25 | 4.477 ± 0.75* (p = 0.002) | 1.112 ± 0.35 | 1.823 ± 0.15 * (p = 0.019) |
| Hdac2 | 1.16 ± 0.64 | 0.866 ± 0.18 (p = 0.483) | 1.05 ± 0.41 | 1.56 ± 0.22 (p = 0.332) |
| mRNA | Control (n = 4–6) | BPA (n = 4–6) |
|---|---|---|
| G6Pase | 1.047 ± 0.331 | 1.251 ± 0.82 (p = 0.606) |
| Star | 1.105 ± 0.299 | 0.809 ± 0.151 (p = 0.426) |
| LXRa | 1.077 ± 0.41 | 1.025 ± 0.38 (p = 0.828) |
| Histological Feature | Score | ||||
|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | ||
| Steatosis | macrovesicular steatosis | <5% | 5–33% | 33–66% | >66% |
| microvesicular steatosis | <5% | 5–33% | 33–66% | >66% | |
| hypertrophy | <5% | 5–33% | 33–66% | >66% | |
| Inflammation | number of inflammatory foci/field | <0.5 | 0.5–1.0 | 1.0–2.0 | >2.0 |
| Target | Forward 5′-3′ | Reverse 5′-3′ | ||
|---|---|---|---|---|
| Inflammation | Tnfα | ACTGATGAGAGGGAGCCCAT | CTGTGCCTCAGCCTCTTCTC | qRT-PCR |
| Il-6 | AGTCTCCTCTCCGGACTTGT | AGAGACTTCCAGCCAGTTGC | qRT-PCR | |
| Nf-kb | TCACTGAGCTCCCGATCAGA | CATACGCTGACCCTAGCCTG | qRT-PCR | |
| Epigenetics | Sirt1 | CACCAGAAAGAACTTCACCACCAG | ACCATCAAGCCGCCTACTAATCTG | qRT-PCR |
| Sirt1-AS lncRNA | TCACATCATGCAAATGGAAAACTAA | TAGGACCATTACTGCCAGAGGA | Strand specific PCR | |
| Sirt1-AS lncRNA | TCACATCATGCAAATGGAAAACTAA | TAGGACCATTACTGCCAGAGGA | qRT-PCR | |
| Sirt1-AS lncRNA | -- | TAGGACCATTACTGCCAGAGGA | RT | |
| Dnmt1 | CTTTTTGGGTGACGGCAACTC | GCTAAGGACGATGATGAGACGC | qRT-PCR | |
| Dnmt3a | CAGCGTCACACAGAAGCATATCC | GGTCCTCACTTTGCTGAACTTGG | qRT-PCR | |
| Hdac1 | ATTGGAAGGGCTGATGTG | AATGCTAATGTTGGGAGG | qRT-PCR | |
| Hdac2 | GCTATCCGCTTGTCTGATGCTC | CAGTTGCCCTTGATTGTGAGATTC | qRT-PCR | |
| Metabolism | LXRa | AGGGCTCCAGGAAGAGATGT | AACTCCGTTGCAGAGTCAGG | qRT-PCR |
| G6Pase | CGTCACCTGTGAGACTGGAC | ACGACATTCAAGCACCGGAA | qRT-PCR | |
| Star | GCTGGCGAACTCTATCTGGGT | GGGAGATGCCTGAGCAAAGC | qRT-PCR | |
| Housekeeping | Actin-β | AGATGACCCAGATCATGTTTGAGA | ACCAGAGGCATACAGGGACAA | qRT-PCR |
| Gapdh | TGGAGTCTACTGGCGTCTT | TGTCATATTTCTCGTGGTTG | qRT-PCR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santoro, A.; Scafuro, M.; Troisi, J.; Piegari, G.; Di Pietro, P.; Mele, E.; Cappetta, D.; Marino, M.; De Angelis, A.; Vecchione, C.; et al. Multi-Systemic Alterations by Chronic Exposure to a Low Dose of Bisphenol A in Drinking Water: Effects on Inflammation and NAD+-Dependent Deacetylase Sirtuin1 in Lactating and Weaned Rats. Int. J. Mol. Sci. 2021, 22, 9666. https://doi.org/10.3390/ijms22189666
Santoro A, Scafuro M, Troisi J, Piegari G, Di Pietro P, Mele E, Cappetta D, Marino M, De Angelis A, Vecchione C, et al. Multi-Systemic Alterations by Chronic Exposure to a Low Dose of Bisphenol A in Drinking Water: Effects on Inflammation and NAD+-Dependent Deacetylase Sirtuin1 in Lactating and Weaned Rats. International Journal of Molecular Sciences. 2021; 22(18):9666. https://doi.org/10.3390/ijms22189666
Chicago/Turabian StyleSantoro, Antonietta, Marika Scafuro, Jacopo Troisi, Giuseppe Piegari, Paola Di Pietro, Elena Mele, Donato Cappetta, Marianna Marino, Antonella De Angelis, Carmine Vecchione, and et al. 2021. "Multi-Systemic Alterations by Chronic Exposure to a Low Dose of Bisphenol A in Drinking Water: Effects on Inflammation and NAD+-Dependent Deacetylase Sirtuin1 in Lactating and Weaned Rats" International Journal of Molecular Sciences 22, no. 18: 9666. https://doi.org/10.3390/ijms22189666
APA StyleSantoro, A., Scafuro, M., Troisi, J., Piegari, G., Di Pietro, P., Mele, E., Cappetta, D., Marino, M., De Angelis, A., Vecchione, C., Paciello, O., Fasano, S., Pierantoni, R., Viggiano, A., & Meccariello, R. (2021). Multi-Systemic Alterations by Chronic Exposure to a Low Dose of Bisphenol A in Drinking Water: Effects on Inflammation and NAD+-Dependent Deacetylase Sirtuin1 in Lactating and Weaned Rats. International Journal of Molecular Sciences, 22(18), 9666. https://doi.org/10.3390/ijms22189666