
Utility of Platelet Endothelial Cell Adhesion Molecule 1 in the Platelet Activity Assessment in Mouse and Human Blood

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
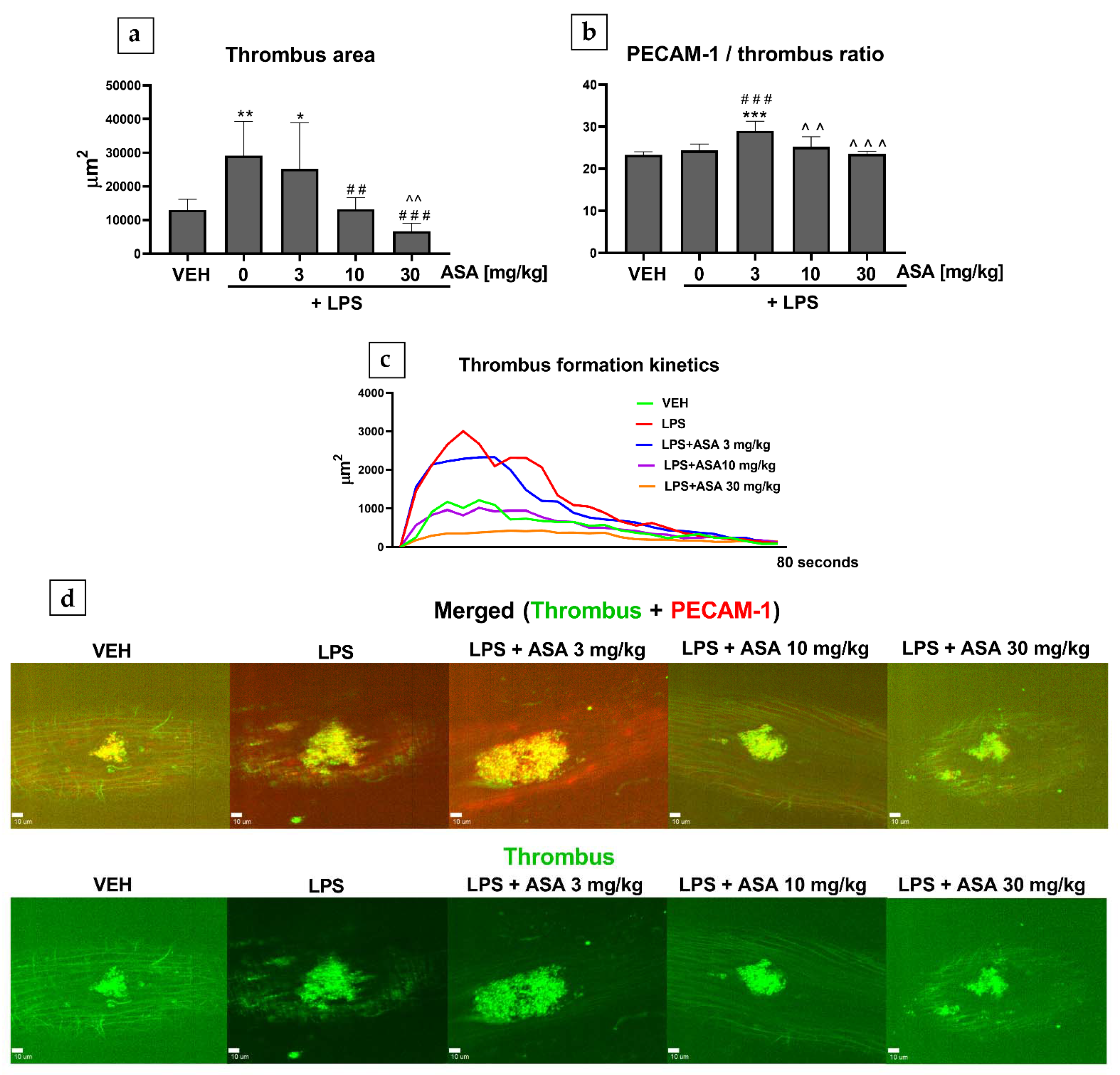
2.1. Laser-Induced Thrombosis in Mice (Intravital)
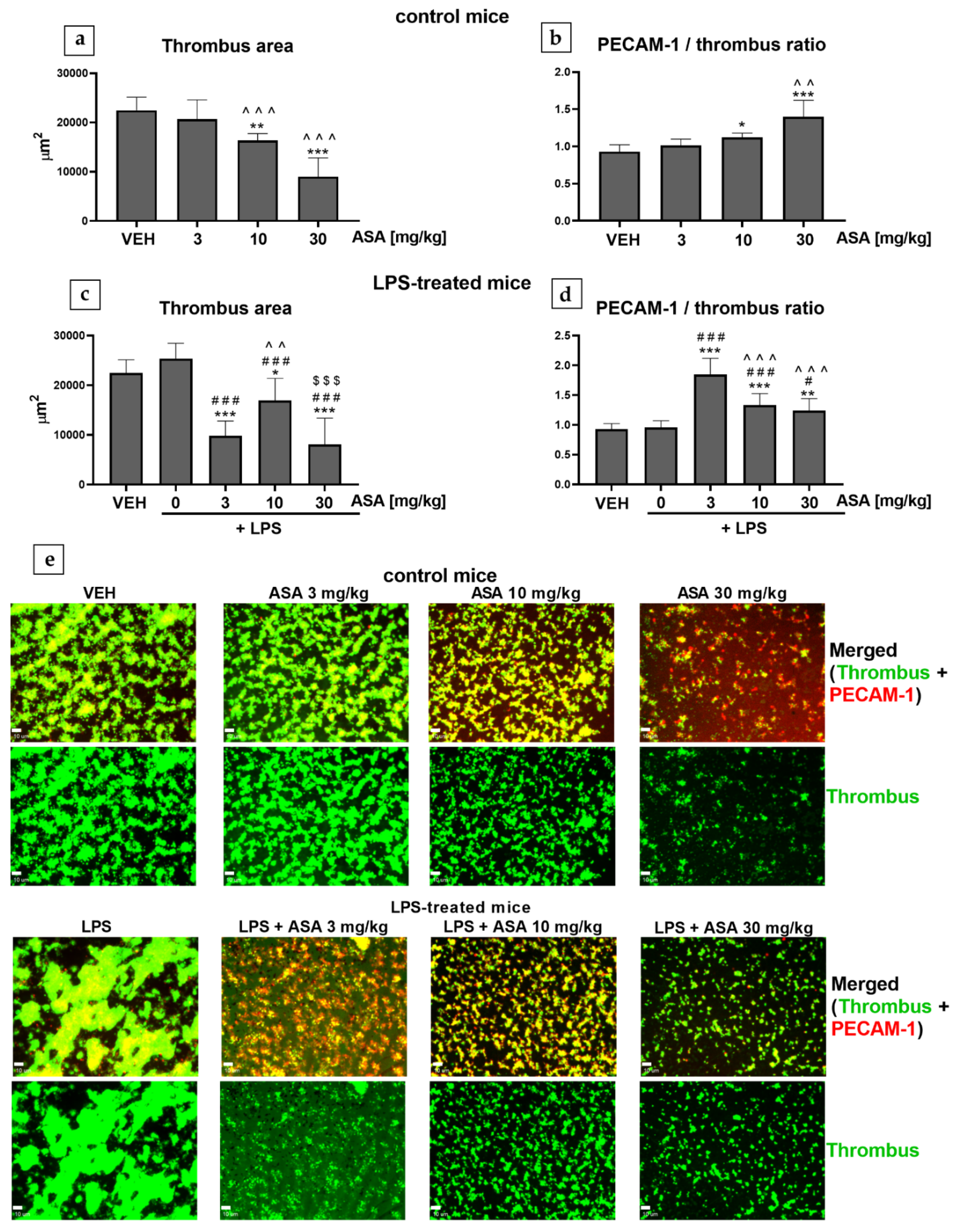
2.2. Thrombus Formation in the Flow Chamber (Mice, Ex Vivo)
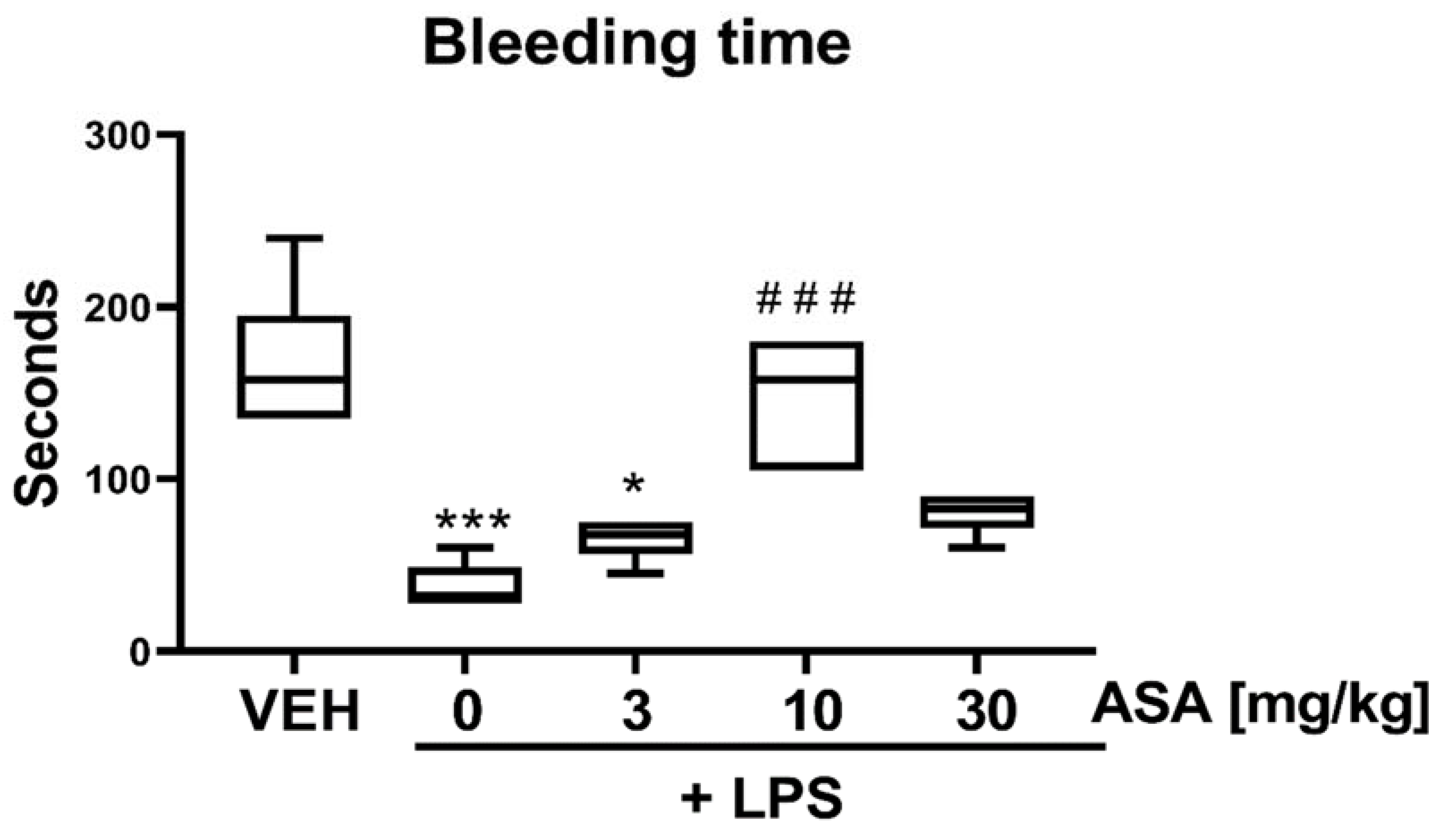
2.3. Bleeding Time (BT) in LPS-Treated Mice
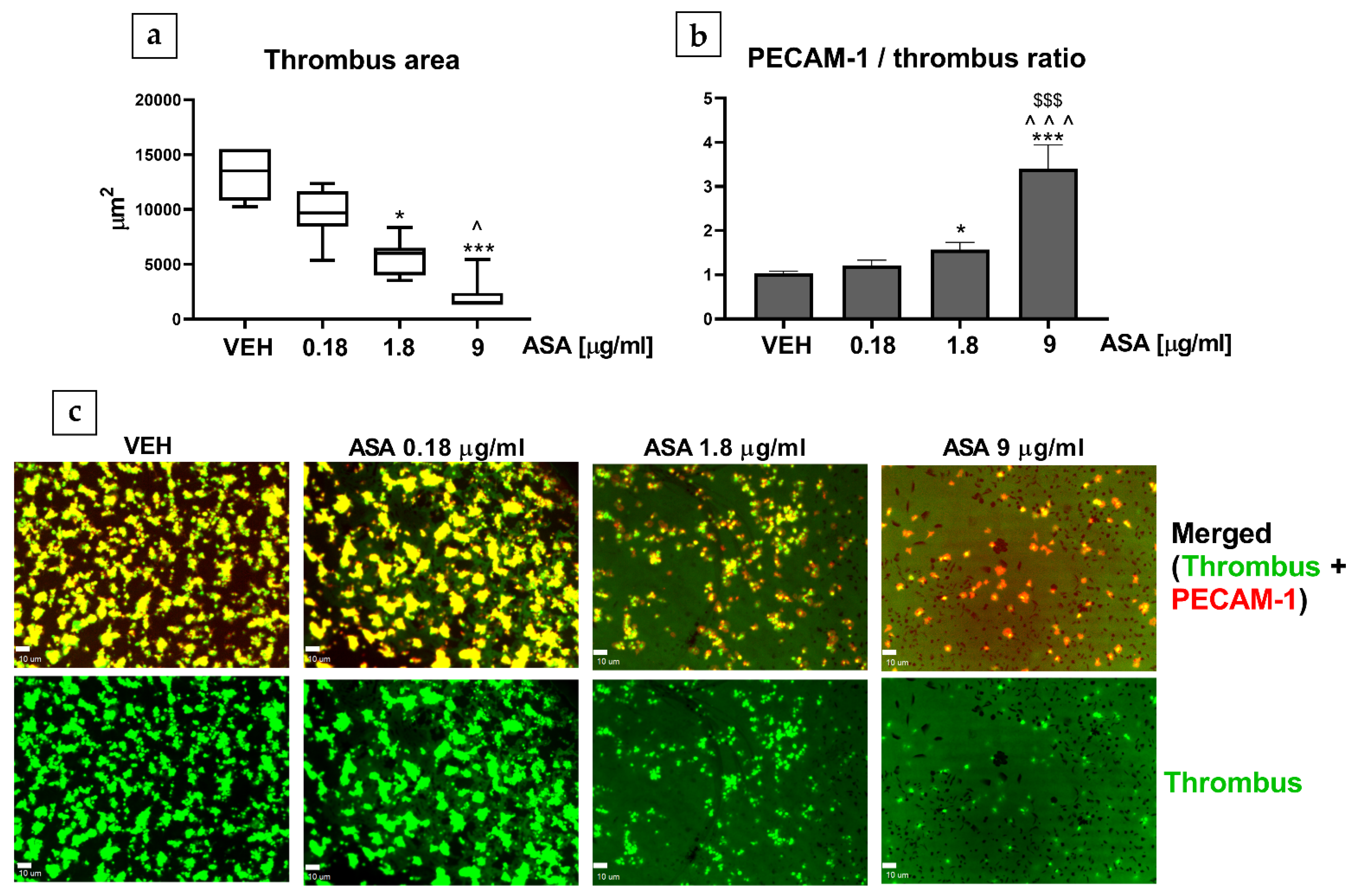
2.4. Thrombus Formation in the Flow Chamber (Human, In Vitro)
3. Discussion
3.1. Mouse Study
3.2. Human Study
4. Materials and Methods
4.1. Blood Collection for Human Study
4.2. Animals
4.3. Induction of Inflammation
4.4. ASA Administration
4.5. Microscopic Visualization
4.6. Flow Chamber Characteristics and Preparation of Collagen-Coated Surfaces
4.7. Model of Thrombus Formation in a Flow Chamber
4.8. Laser-Induced Thrombosis in Mouse Mesenteric Vein and Assessment of Thrombus Area and PECAM-1/Thrombus Ratio (Intravital)
4.9. Determination of BT in Mice
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Soriano Jerez, E.M.; Gibbins, J.M.; Hughes, C.E. Targeting platelet inhibition receptors for novel therapies: PECAM-1 and G6b-B. Platelets 2021, 32, 761–769. [Google Scholar] [CrossRef]
- Jones, C.I.; Garner, S.F.; Moraes, L.A.; Kaiser, W.J.; Rankin, A.; Ouwehand, W.H.; Goodall, A.H.; Gibbins, J.M. PECAM-1 expression and activity negatively regulate multiple platelet signaling pathways. FEBS Lett. 2009, 583, 3618–3624. [Google Scholar] [CrossRef] [Green Version]
- Falati, S.; Patil, S.; Gross, P.L.; Stapleton, M.; Merrill-Skoloff, G.; Barrett, N.E.; Pixton, K.L.; Weiler, H.; Cooley, B.; Newman, D.K.; et al. Platelet PECAM-1 inhibits thrombus formation in vivo. Blood 2006, 107, 535–541. [Google Scholar] [CrossRef] [Green Version]
- Marcinczyk, N.; Golaszewska, A.; Misztal, T.; Gromotowicz-Poplawska, A.; Rusak, T.; Chabielska, E. New approaches for the assessment of platelet activation status in thrombus under flow condition using confocal microscopy. Naunyn. Schmiedebergs. Arch. Pharmacol. 2020, 393, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Marcinczyk, N.; Gołaszewska, A.; Gromotowicz-Poplawska, A.; Misztal, T.; Strawa, J.; Tomczyk, M.; Kasacka, I.; Chabielska, E. Multidirectional effects of tormentil extract on hemostasis in experimental diabetes. Front. Pharmacol. 2021, 12, 682987. [Google Scholar] [CrossRef] [PubMed]
- Beristain-Covarrubias, N.; Perez-Toledo, M.; Thomas, M.R.; Henderson, I.R.; Watson, S.P.; Cunningham, A.F. Understanding infection-induced thrombosis: Lessons learned from animal models. Front. Immunol. 2019, 10, 2569. [Google Scholar] [CrossRef] [Green Version]
- Schrör, K. Aspirin and platelets: The antiplatelet action of aspirin and its role in thrombosis treatment and prophylaxis. Semin. Thromb. Hemost. 1997, 23, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Dubois, C.; Panicot-Dubois, L.; Gainor, J.F.; Furie, B.C.; Furie, B. Thrombin-initiated platelet activation in vivo is vWF independent during thrombus formation in a laser injury model. J. Clin. Investig. 2007, 117, 953–960. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, T.; Mogami, H.; Murakami, Y.; Nakamura, T.; Kanayama, N.; Konno, H.; Urano, T. Real-time analysis of platelet aggregation and procoagulant activity during thrombus formation in vivo. Pflügers Arch.-Eur. J. Physiol. 2008, 456, 1239–1251. [Google Scholar] [CrossRef] [Green Version]
- Gollomp, K.; Johnston, I.; Rauova, L.; Cines, D.B.; Kowalska, M.A.; Poncz, M. A special role for neutrophil extracellular traps (NETs) and neutrophils in the prothombotic nature of heparin-induced thrombocytopenia. Blood 2016, 128, 1023. [Google Scholar] [CrossRef]
- de Witt, S.M.; Swieringa, F.; Cavill, R.; Lamers, M.M.E.; van Kruchten, R.; Mastenbroek, T.; Baaten, C.; Coort, S.; Pugh, N.; Schulz, A.; et al. Identification of platelet function defects by multi-parameter assessment of thrombus formation. Nat. Commun. 2014, 5, 4257. [Google Scholar] [CrossRef] [Green Version]
- Clark, S.R.; Ma, A.C.; Tavener, S.A.; McDonald, B.; Goodarzi, Z.; Kelly, M.M.; Patel, K.D.; Chakrabarti, S.; McAvoy, E.; Sinclair, G.D.; et al. Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. Nat. Med. 2007, 13, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Frydman, G.H.; Le, A.; Ellett, F.; Jorgensen, J.; Fox, J.G.; Tompkins, R.G.; Irimia, D. Technical advance: Changes in neutrophil migration patterns upon contact with platelets in a microfluidic assay. J. Leukoc. Biol. 2017, 101, 797–806. [Google Scholar] [CrossRef] [Green Version]
- Lorrain, J.; Lechaire, I.; Gauffeny, C.; Masson, R.; Roome, N.; Herault, J.-P.; O’Connor, S.E.; Schaeffer, P.; Herbert, J.-M. Effects of SanOrg123781A, a synthetic hexadecasaccharide, in a mouse model of electrically induced carotid artery injury: Synergism with the antiplatelet agent clopidogrel. J. Pharmacol. Exp. Ther. 2004, 309, 235–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aktas, B.; Pozgajova, M.; Bergmeier, W.; Sunnarborg, S.; Offermanns, S.; Lee, D.; Wagner, D.D.; Nieswandt, B. Aspirin induces platelet receptor shedding via ADAM17 (TACE). J. Biol. Chem. 2005, 280, 39716–39722. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Sturgeon, S.; Mountford, J.; Jackson, S.; Hamilton, J. Safety and efficacy of targeting platelet proteinase-activated receptors in combination with existing anti-platelet drugs as antithrombotics in mice. Br. J. Pharmacol. 2012, 166, 2188–2197. [Google Scholar] [CrossRef] [Green Version]
- Wientjes, M.G.; Levy, G. Nonlinear pharmacokinetics of aspirin in rats. J. Pharmacol. Exp. Ther. 1988, 245, 809–815. [Google Scholar]
- Fu, C.J.; Melethil, S.; Mason, W.D. The pharmacokinetics of aspirin in rats and the effect of buffer. J. Pharmacokinet. Biopharm. 1991, 19, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Nagelschmitz, J.; Blunck, M.; Kraetzschmar, J.; Ludwig, M.; Wensing, G.; Hohlfeld, T. Pharmacokinetics and pharmacodynamics of acetylsalicylic acid after intravenous and oral administration to healthy volunteers. Clin. Pharmacol. 2014, 6, 51–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Droebner, K.; Haasbach, E.; Dudek, S.E.; Scheuch, G.; Nocker, K.; Canisius, S.; Ehrhardt, C.; von Degenfeld, G.; Ludwig, S.; Planz, O. Pharmacodynamics, pharmacokinetics, and antiviral activity of BAY 81-8781, a novel NF-κB inhibiting anti-influenza drug. Front. Microbiol. 2017, 8, 2130. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, A.; Guichard, J.; Massé, J.M.; Debili, N.; Cramer, E.M. Of mice and men: Comparison of the ultrastructure of megakaryocytes and platelets. Exp. Hematol. 2001, 29, 1295–1302. [Google Scholar] [CrossRef]
- Wang, X. Lipopolysaccharide augments venous and arterial thrombosis in the mouse. Thromb. Res. 2008, 123, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Rumbaut, R.E.; Bellera, R.V.; Randhawa, J.K.; Shrimpton, C.N.; Dasgupta, S.K.; Dong, J.F.; Burns, A.R. Endotoxin enhances microvascular thrombosis in mouse cremaster venules via a TLR4-dependent, neutrophil-independent mechanism. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H1671-9. [Google Scholar] [CrossRef]
- Martyanov, A.A.; Maiorov, A.S.; Filkova, A.A.; Ryabykh, A.A.; Svidelskaya, G.S.; Artemenko, E.O.; Gambaryan, S.P.; Panteleev, M.A.; Sveshnikova, A.N. Effects of bacterial lipopolysaccharides on platelet function: Inhibition of weak platelet activation. Sci. Rep. 2020, 10, 12296. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fang, S.; Li, X.; Feng, J.; Du, J.; Guo, L.; Su, Y.; Zhou, J.; Ding, G.; Bai, Y.; et al. Aspirin inhibits LPS-induced macrophage activation via the NF-κB pathway. Sci. Rep. 2017, 7, 11549. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, M.; Trampuz, A.; Kirschbaum, S.; Winkler, T.; Sass, F.A. Soluble Pecam-1 as a biomarker in periprosthetic joint infection. J. Clin. Med. 2021, 10, 612. [Google Scholar] [CrossRef] [PubMed]
- Paddock, C.; Zhou, D.; Lertkiatmongkol, P.; Newman, P.J.; Zhu, J. Structural basis for PECAM-1 homophilic binding. Blood 2016, 127, 1052–1061. [Google Scholar] [CrossRef] [Green Version]
- Misztal, T.; Golaszewska, A.; Branska-Januszewska, J.; Marcinczyk, N.; Chabielska, E.; Tomasiak, M.; Rusak, T. HAuCl4, putative general aquaporins blocker, reduces platelet spreading, filopodia formation, procoagulant response, and thrombus formation under flow. Front. Physiol. 2020, 11, 1025. [Google Scholar] [CrossRef]
- Zhou, X.; Wu, Y.; Ye, L.; Wang, Y.; Zhang, K.; Wang, L.; Huang, Y.; Wang, L.; Xian, S.; Zhang, Y.; et al. Aspirin alleviates endothelial gap junction dysfunction through inhibition of NLRP3 inflammasome activation in LPS-induced vascular injury. Acta Pharm. Sin. B 2019, 9, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Paniccia, R.; Priora, R.; Alessandrello Liotta, A.; Abbate, R. Platelet function tests: A comparative review. Vasc. Health Risk Manag. 2015, 133–148. [Google Scholar] [CrossRef] [Green Version]
- Dovizio, M.; Bruno, A.; Tacconelli, S.; Patrignani, P. Mode of action of aspirin as a chemopreventive agent. Recent Results Cancer Res. 2013, 191, 39–65. [Google Scholar] [CrossRef]
- Rosenkranz, B.; Frölich, J.C. Plasma concentrations and anti-platelet effects after low dose acetylsalicylic acid. Prostaglandins. Leukot. Med. 1985, 19, 289–300. [Google Scholar] [CrossRef]
- Serebruany, V.L.; Malinin, A.I.; Ziai, W.; Pokov, A.N.; Bhatt, D.L.; Alberts, M.J.; Hanley, D.F. Effects of clopidogrel and aspirin in combination versus aspirin alone on platelet activation and major receptor expression in patients after recent ischemic stroke. Stroke 2005, 36, 2289–2292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwaginga, J.J.; Nash, G.; King, M.R.; Heemskerk, J.W.M.; Frojmovic, M.; Hoylaerts, M.F.; Sakariassen, K.S. Flow-based assays for global assessment of hemostasis. Part 1: Biorheologic considerations. J. Thromb. Haemost. 2006, 4, 2486–2487. [Google Scholar] [CrossRef] [PubMed]
- Van Kruchten, R.; Cosemans, J.M.E.M.; Heemskerk, J.W.M. Measurement of whole blood thrombus formation using parallel-plate flow chambers–a practical guide. Platelets 2012, 23, 229–242. [Google Scholar] [CrossRef]
- Vital, S.A.; Senchenkova, E.Y.; Ansari, J.; Gavins, F.N.E. Targeting AnxA1/formyl peptide receptor 2 pathway affords protection against pathological thrombo-inflammation. Cells 2020, 9, 2473. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marcinczyk, N.; Misztal, T.; Gromotowicz-Poplawska, A.; Zebrowska, A.; Rusak, T.; Radziwon, P.; Chabielska, E. Utility of Platelet Endothelial Cell Adhesion Molecule 1 in the Platelet Activity Assessment in Mouse and Human Blood. Int. J. Mol. Sci. 2021, 22, 9611. https://doi.org/10.3390/ijms22179611
Marcinczyk N, Misztal T, Gromotowicz-Poplawska A, Zebrowska A, Rusak T, Radziwon P, Chabielska E. Utility of Platelet Endothelial Cell Adhesion Molecule 1 in the Platelet Activity Assessment in Mouse and Human Blood. International Journal of Molecular Sciences. 2021; 22(17):9611. https://doi.org/10.3390/ijms22179611
Chicago/Turabian StyleMarcinczyk, Natalia, Tomasz Misztal, Anna Gromotowicz-Poplawska, Agnieszka Zebrowska, Tomasz Rusak, Piotr Radziwon, and Ewa Chabielska. 2021. "Utility of Platelet Endothelial Cell Adhesion Molecule 1 in the Platelet Activity Assessment in Mouse and Human Blood" International Journal of Molecular Sciences 22, no. 17: 9611. https://doi.org/10.3390/ijms22179611
APA StyleMarcinczyk, N., Misztal, T., Gromotowicz-Poplawska, A., Zebrowska, A., Rusak, T., Radziwon, P., & Chabielska, E. (2021). Utility of Platelet Endothelial Cell Adhesion Molecule 1 in the Platelet Activity Assessment in Mouse and Human Blood. International Journal of Molecular Sciences, 22(17), 9611. https://doi.org/10.3390/ijms22179611