
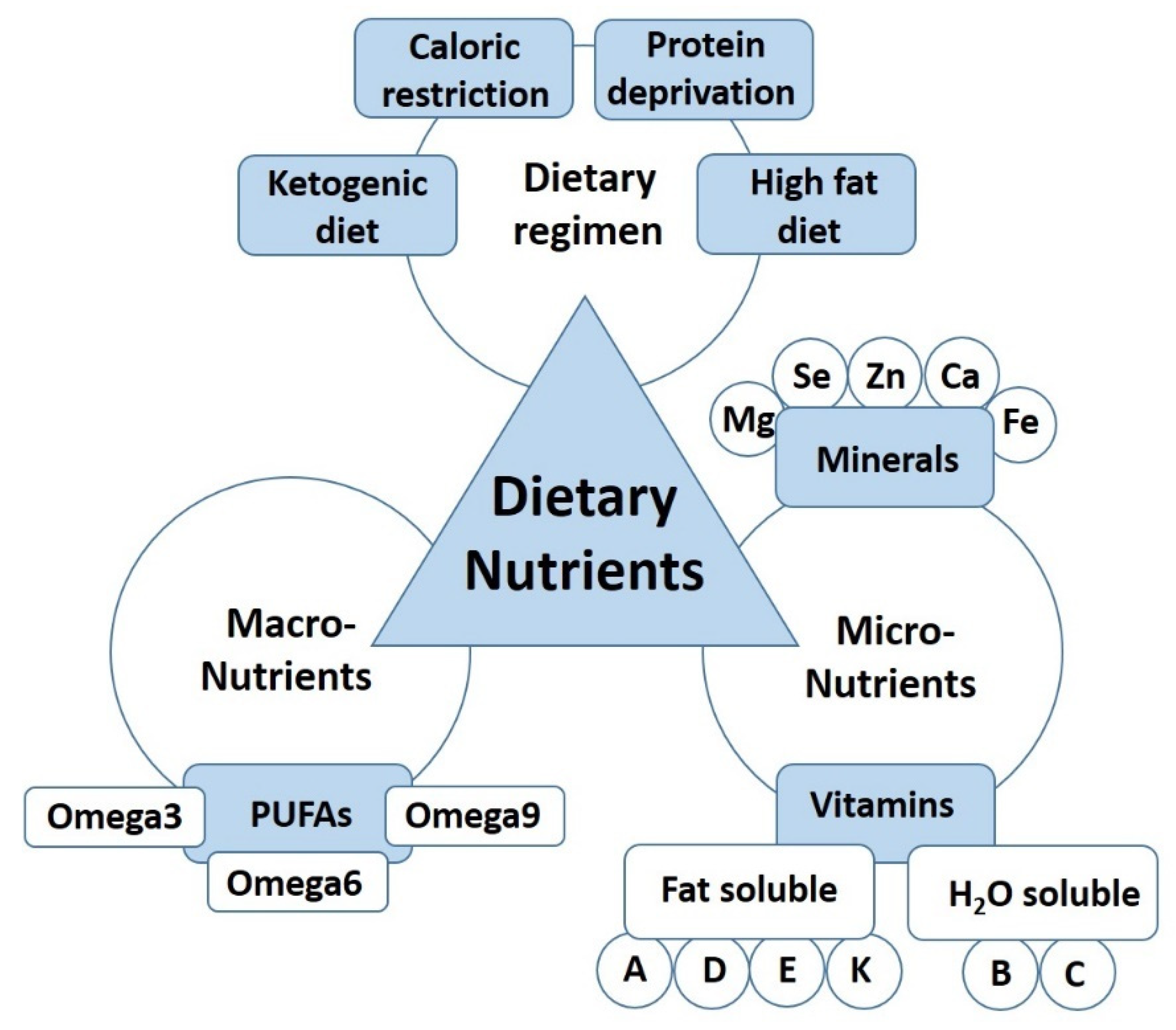
The Role of Dietary Nutrients in Peripheral Nerve Regeneration

 ,
,  , , , and
, , , and
Abstract
1. Introduction
2. Methods
3. Dietary Regimen
3.1. Ketogenic Diet
3.2. Caloric Restriction
3.3. Protein Deficient Diet (Protein Deprivation)
3.4. High Fat Diet
4. Dietary Macronutrients in Peripheral Nerve Regeneration
4.1. Polyunsaturated Fatty Acids (PUFAs)
4.1.1. Omega-3
4.1.2. Omega-6
4.1.3. Omega-9
5. Role of Dietary Micronutrients in Peripheral Nerve Regeneration
5.1. Vitamins
5.1.1. Lipid-Soluble Vitamins
Vitamin A and Retinoic Acid
Vitamin D (Calciferol)

{kind=link}
{kind=link}
| Reference | Type of Nerve Lesion and Animal Model | Type and Timing of Administration/Experimental Groups |
|---|---|---|
| Vitamin D | ||
| Albay et al., 2020 [96] | Sciatic nerve crush (Wistar–Hannover female rats) |
|
| Montava et al., 2015 [97] | Unilateral transected facial nerve repaired with autograft (New Zealand white rabbits) | From day 1 post-surgery weekly oral bolus of vitamin D3 (Uvedose® 200 IU/kg/day Crinex). Treatment for 12 weeks. |
| Chabas et al., 2013 [95] | 10 mm of the peroneal nerve cut out and repaired with autograft (Sprague–Dawley male adult rats) | Immediately after the injury and repair, rats were orally fed weekly (for eleven weeks) with boli containing:
|
| Chabas et al., 2008 [91] | 10 mm of the peroneal nerve cut out and repaired with autograft (Sprague–Dawley male adult rats) | 3 times for month oral vitamin D2 administration (Sterogyl® 100 IU/kg/day) for 12 weeks |
| Vitamin E | ||
| Azizi et al., 2014 [98] | 10 mm sciatic nerve transection unrepaired or repaired with chitosan conduit (Wistar male rats) | The chitosan conduit was filled with vitamin E (20 mg/kg, DL-all-rac-α-tocopherol) or pyrroloquinoline quinone (PPQ, 0.03 mmol/l) or a combination 1:1 of vitamin E and PPQ. Analysis were performed 4, 8, and 12 weeks after surgery. |
| Enrione et al., 1999 [99] | Sciatic nerve crush (Sprague–Dawley male rats) | Vitamin E-deficient diet: dl-α-tocopheryl acetate 0 mg/kg diet After 22 days of diet animals received nerve injury and continued feeding until sacrifice (15 days post-surgery). |
| Cecchini et al., 1994 [100] | Sciatic nerve crush (Sprague–Dawley male rats) | From day 30 after birth-deficient rats were fed a diet without vitamin E and injured at 3 months of age. Diet maintained until sacrifice (2 months from the injury). |
| Cuppini et al., 1993 [101] | Sciatic nerve crush (male Sprague–Dawley rats) | Vitamin E-deficient group: fed with a diet lacking vitamin E for 2 months before surgery. The dietetic treatment was maintained until the sacrifice (30 or 60 days after the lesion). |
| Cuppini et al., 1990 [102] | Sciatic nerve crush (Sprague–Dawley rats) |
|
Vitamin E
Vitamin K
5.1.2. Water-Soluble Vitamins
Vitamin B
Vitamin C (Ascorbic Acid)
5.2. Minerals
5.2.1. Magnesium
5.2.2. Selenium
5.2.3. Zinc
5.2.4. Calcium
5.2.5. Iron
6. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Huckhagel, T.; Nüchtern, J.; Regelsberger, J.; Gelderblom, M.; Lefering, R. Nerve trauma of the lower extremity: Evaluation of 60,422 leg injured patients from the TraumaRegister DGU® between 2002 and 2015. Scand. J. Trauma. Resusc. Emerg. Med. 2018, 26. [Google Scholar] [CrossRef] [PubMed]
- Huckhagel, T.; Nüchtern, J.; Regelsberger, J.; Lefering, R. Nerve injury in severe trauma with upper extremity involvement: Evaluation of 49,382 patients from the TraumaRegister DGU® between 2002 and 2015. Scand. J. Trauma. Resusc. Emerg. Med. 2018, 26, 76. [Google Scholar] [CrossRef] [PubMed]
- Daly, W.; Yao, L.; Zeugolis, D.; Windebank, A.; Pandit, A. A biomaterials approach to peripheral nerve regeneration: Bridging the peripheral nerve gap and enhancing functional recovery. J. R. Soc. Interface 2012, 9, 202–221. [Google Scholar] [CrossRef] [PubMed]
- Ciardelli, G.; Chiono, V. Materials for peripheral nerve regeneration. Macromol. Biosci. 2006, 6, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Grinsell, D.; Keating, C.P. Peripheral Nerve Reconstruction after Injury: A Review of Clinical and Experimental Therapies. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Hussain, G.; Wang, J.; Rasul, A.; Anwar, H.; Qasim, M.; Zafar, S.; Aziz, N.; Razzaq, A.; Hussain, R.; de Aguilar, J.L.G.; et al. Current status of therapeutic approaches against peripheral nerve injuries: A detailed story from injury to recovery. Int. J. Biol. Sci. 2020, 16, 116–134. [Google Scholar] [CrossRef]
- Modrak, M.; Talukder, M.A.H.; Gurgenashvili, K.; Noble, M.; Elfar, J.C. Peripheral nerve injury and myelination: Potential therapeutic strategies. J. Neurosci. Res. 2020, 98, 780–795. [Google Scholar] [CrossRef]
- Kubiak, C.A.; Kung, T.A.; Brown, D.L.; Cederna, P.S.; Kemp, S.W.P. State-of-The-Art techniques in treating peripheral nerve injury. Plast. Reconstr. Surg. 2018, 141, 702–710. [Google Scholar] [CrossRef]
- Sullivan, R.; Dailey, T.; Duncan, K.; Abel, N.; Borlongan, C. Peripheral nerve injury: Stem cell therapy and peripheral nerve transfer. Int. J. Mol. Sci. 2016, 17, 2101. [Google Scholar] [CrossRef]
- Basson, A.; Olivier, B.; Ellis, R.; Coppieters, M.; Stewart, A.; Mudzi, W. The effectiveness of neural mobilization for neuromusculoskeletal conditions: A systematic review and meta-Analysis. J. Orthop. Sports Phys. Ther. 2017, 47, 593–615. [Google Scholar] [CrossRef] [PubMed]
- Griffin, J.W.; Hogan, M.C.V.; Chhabra, A.B.; Deal, D.N. Peripheral nerve repair and reconstruction. J. Bone Jt. Surg. Ser. A 2013, 95, 2144–2151. [Google Scholar] [CrossRef] [PubMed]
- Micha, R.; Peñalvo, J.L.; Cudhea, F.; Imamura, F.; Rehm, C.D.; Mozaffarian, D. Association between dietary factors and mortality from heart disease, stroke, and type 2 diabetes in the United States. JAMA J. Am. Med. Assoc. 2017, 317, 912–924. [Google Scholar] [CrossRef]
- Wiseman, M.J. Nutrition and cancer: Prevention and survival. Br. J. Nutr. 2019, 122, 481–487. [Google Scholar] [CrossRef]
- Muñoz Fernández, S.S.; Lima Ribeiro, S.M. Nutrition and Alzheimer Disease. Clin. Geriatr. Med. 2018, 34, 677–697. [Google Scholar] [CrossRef] [PubMed]
- Boulos, C.; Yaghi, N.; El Hayeck, R.; Heraoui, G.N.H.A.; Fakhoury-Sayegh, N. Nutritional risk factors, microbiota and parkinson’s disease: What is the current evidence? Nutrients 2019, 11, 1896. [Google Scholar] [CrossRef]
- Granic, A.; Sayer, A.A.; Robinson, S.M. Dietary patterns, skeletal muscle health, and sarcopenia in older adults. Nutrients 2019, 11, 745. [Google Scholar] [CrossRef] [PubMed]
- Yildiran, H.; Macit, M.S.; Özata Uyar, G. New approach to peripheral nerve injury: Nutritional therapy. Nutr. Neurosci. 2020, 23, 744–755. [Google Scholar] [CrossRef] [PubMed]
- Dąbek, A.; Wojtala, M.; Pirola, L.; Balcerczyk, A. Modulation of cellular biochemistry, epigenetics and metabolomics by ketone bodies. Implications of the ketogenic diet in the physiology of the organism and pathological states. Nutrients 2020, 12, 788. [Google Scholar] [CrossRef]
- Li, R.J.; Liu, Y.; Liu, H.Q.; Li, J. Ketogenic diets and protective mechanisms in epilepsy, metabolic disorders, cancer, neuronal loss, and muscle and nerve degeneration. J. Food Biochem. 2020, 44, 13140. [Google Scholar] [CrossRef]
- Cooper, M.A.; McCoin, C.; Pei, D.; Thyfault, J.P.; Koestler, D.; Wright, D.E. Reduced mitochondrial reactive oxygen species production in peripheral nerves of mice fed a ketogenic diet. Exp. Physiol. 2018, 103, 1206–1212. [Google Scholar] [CrossRef] [PubMed]
- Sayadi, J.J.; Sayadi, L.; Satteson, E.; Chopan, M. Nerve injury and repair in a ketogenic milieu: A systematic review of traumatic injuries to the spinal cord and peripheral nervous tissue. PLoS ONE 2021, 16. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, Y.; Liu, H.; Chen, L.; Li, R. Ketogenic Diet Potentiates Electrical Stimulation–Induced Peripheral Nerve Regeneration after Sciatic Nerve Crush Injury in Rats. Mol. Nutr. Food Res. 2020, 64. [Google Scholar] [CrossRef] [PubMed]
- Mayr, K.A.; Kwok, C.H.T.; Eaton, S.E.A.; Baker, G.B.; Whelan, P.J. The effects of a ketogenic diet on sensorimotor function in a thoracolumbar mouse spinal cord injury model. eNeuro 2020, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Liśkiewicz, A.; Właszczuk, A.; Gendosz, D.; Larysz-Brysz, M.; Kapustka, B.; Łączyński, M.; Lewin-Kowalik, J.; Jędrzejowska-Szypułka, H. Sciatic nerve regeneration in rats subjected to ketogenic diet. Nutr. Neurosci. 2016, 19, 116–124. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, F.; Vacca, V.; Pavone, F.; Marinelli, S. Impact of caloric restriction on peripheral nerve injury-induced neuropathic pain during ageing in mice. Eur. J. Pain 2020, 24, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Coccurello, R.; Nazio, F.; Rossi, C.; De Angelis, F.; Vacca, V.; Giacovazzo, G.; Procacci, P.; Magnaghi, V.; Ciavardelli, D.; Marinelli, S. Effects of caloric restriction on neuropathic pain, peripheral nerve degeneration and inflammation in normometabolic and autophagy defective prediabetic Ambra1 mice. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Song, Z.; Xie, W.; Strong, J.A.; Berta, T.; Ulrich-Lai, Y.M.; Guo, Q.; Zhang, J.M. High-fat diet exacerbates postoperative pain and inflammation in a sex-dependent manner. Pain 2018, 159, 1731–1741. [Google Scholar] [CrossRef]
- Bekar, E.; Altunkaynak, B.Z.; Balc, K.; Aslan, G.; Ayyldz, M.; Kaplan, S. Effects of high fat diet induced obesity on peripheral nerve regeneration and levels of GAP 43 and TGF-β in rats. Biotech. Histochem. 2014, 89, 446–456. [Google Scholar] [CrossRef]
- Golbidi, S.; Daiber, A.; Korac, B.; Li, H.; Essop, M.F.; Laher, I. Health Benefits of Fasting and Caloric Restriction. Curr. Diab. Rep. 2017, 17. [Google Scholar] [CrossRef]
- Lee, S.; Notterpek, L. Dietary restriction supports peripheral nerve health by enhancing endogenous protein quality control mechanisms. Exp. Gerontol. 2013, 48, 1085–1090. [Google Scholar] [CrossRef]
- Rangaraju, S.; Hankins, D.; Madorsky, I.; Madorsky, E.; Lee, W.H.; Carter, C.S.; Leeuwenburgh, C.; Notterpek, L. Molecular architecture of myelinated peripheral nerves is supported by calorie restriction with aging. Aging Cell 2009, 8, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Oldfors, A. Nerve fibre degeneration of the central and peripheral nervous systems in severe protein deprivation in rats. Acta Neuropathol. 1981, 54, 121–127. [Google Scholar] [CrossRef]
- Gomes, S.P.; Nyengaard, J.R.; Misawa, R.; Girotti, P.A.; Castelucci, P.; Blazquez, F.H.J.; De Melo, M.P.; Ribeiro, A.A.C. Atrophy and neuron loss: Effects of a protein-deficient diet on sympathetic neurons. J. Neurosci. Res. 2009, 87, 3568–3575. [Google Scholar] [CrossRef] [PubMed]
- Chopra, J.S.; Dhand, U.K.; Mehta, S.; Bakshi, V.; Rana, S.; Mehta, J. Effect of protein calorie malnutrition on peripheral nerves: A clinical, electrophysiological and histopathological study. Brain 1986, 109, 307–323. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Strong, J.A.; Zhang, J.M. Active nerve regeneration with failed target reinnervation drives persistent neuropathic pain. eNeuro 2017, 4. [Google Scholar] [CrossRef]
- Zhang, A.C.; De Silva, M.E.H.; MacIsaac, R.J.; Roberts, L.; Kamel, J.; Craig, J.P.; Busija, L.; Downie, L.E. Omega-3 polyunsaturated fatty acid oral supplements for improving peripheral nerve health: A systematic review and meta-analysis. Nutr. Rev. 2020, 78, 323–341. [Google Scholar] [CrossRef]
- Gladman, S.J.; Huang, W.; Lim, S.N.; Dyall, S.C.; Boddy, S.; Kang, J.X.; Knight, M.M.; Priestley, J.V.; Michael-Titus, A.T. Improved outcome after peripheral nerve injury in mice with increased levels of endogenous Omega-3 polyunsaturated fatty acids. J. Neurosci. 2012, 32, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Shi, Z.; Su, H.; So, K.F.; Cui, Q. Increased production of omega-3 fatty acids protects retinal ganglion cells after optic nerve injury in mice. Exp. Eye Res. 2016, 148, 90–96. [Google Scholar] [CrossRef]
- Galán-Arriero, I.; Serrano-Muñoz, D.; Gómez-Soriano, J.; Goicoechea, C.; Taylor, J.; Velasco, A.; Ávila-Martín, G. The role of Omega-3 and Omega-9 fatty acids for the treatment of neuropathic pain after neurotrauma. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1629–1635. [Google Scholar] [CrossRef]
- Motta, J.R.; Jung, I.E.d.C.; Azzolin, V.F.; Teixeira, C.F.; Braun, L.E.; De Oliveira Nerys, D.A.; Motano, M.A.E.; Duarte, M.M.M.F.; Maia-Ribeiro, E.A.; da Cruz, I.B.M.; et al. Avocado oil (Persea americana) protects SH-SY5Y cells against cytotoxicity triggered by cortisol by the modulation of BDNF, oxidative stress, and apoptosis molecules. J. Food Biochem. 2021, 45. [Google Scholar] [CrossRef]
- Mitsou, E.; Pletsa, V.; Sotiroudis, G.T.; Panine, P.; Zoumpanioti, M.; Xenakis, A. Development of a microemulsion for encapsulation and delivery of gallic acid. The role of chitosan. Colloids Surf. B Biointerfaces 2020, 190, 110974. [Google Scholar] [CrossRef]
- Kosek, V.; Heczkova, M.; Novak, F.; Meisnerova, E.; Novákova, O.; Zelenka, J.; Bechynska, K.; Vrzacova, N.; Suttnar, J.; Hlavackova, A.; et al. The ω-3 Polyunsaturated Fatty Acids and Oxidative Stress in Long-Term Parenteral Nutrition Dependent Adult Patients: Functional Lipidomics Approach. Nutrients 2020, 12, 2351. [Google Scholar] [CrossRef]
- Pascoe, M.C.; Howells, D.W.; Crewther, D.P.; Carey, L.M.; Crewther, S.G. Fish oil supplementation associated with decreased cellular degeneration and increased cellular proliferation 6 weeks after middle cerebral artery occlusion in the rat. Neuropsychiatr. Dis. Treat. 2015, 11, 153–164. [Google Scholar] [CrossRef]
- Pascoe, M.C.; Howells, D.W.; Crewther, D.P.; Constantinou, N.; Carey, L.M.; Rewell, S.S.; Turchini, G.M.; Kaur, G.; Crewther, S.G. Fish oil diet associated with acute reperfusion related hemorrhage, and with reduced stroke-related sickness behaviors and motor impairment. Front. Neurol. 2014, 5 FEB. [Google Scholar] [CrossRef]
- Dyall, S.C.; Michael-Titus, A.T. Neurological benefits of omega-3 fatty acids. NeuroMolecular Med. 2008, 10, 219–235. [Google Scholar] [CrossRef] [PubMed]
- Hasadsri, L.; Wang, B.H.; Lee, J.V.; Erdman, J.W.; Llano, D.A.; Barbey, A.K.; Wszalek, T.; Sharrock, M.F.; Wang, H. Omega-3 Fatty Acids as a Putative Treatment for Traumatic Brain Injury. J. Neurotrauma 2013, 30, 897–906. [Google Scholar] [CrossRef]
- Michael-Titus, A.T.; Priestley, J.V. Omega-3 fatty acids and traumatic neurological injury: From neuroprotection to neuroplasticity? Trends Neurosci. 2014, 37, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Zendedel, A.; Habib, P.; Dang, J.; Lammerding, L.; Hoffmann, S.; Beyer, C.; Slowik, A. Omega-3 polyunsaturated fatty acids ameliorate neuroinflammation and mitigate ischemic stroke damage through interactions with astrocytes and microglia. J. Neuroimmunol. 2015, 278, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Erkayıran, U.; Isık, H.; Seven, A.; Batıoglu, A.S.; Kayaalp, D.; Caydere, M.; Ergun, Y. Omega-3 prevents myelin degeneration in rat foetuses exposed to radiation. J. Obstet. Gynaecol. 2016, 36, 444–449. [Google Scholar] [CrossRef]
- Liu, J.W.; Almaguel, F.G.; Bu, L.; De Leon, D.D.; De Leon, M. Expression of E-FABP in PC12 cells increases neurite extension during differentiation: Involvement of n-3 and n-6 fatty acids. J. Neurochem. 2008, 106, 2015–2029. [Google Scholar] [CrossRef] [PubMed]
- Unda, S.R.; Villegas, E.A.; Toledo, M.E.; Asis Onell, G.; Laino, C.H. Beneficial effects of fish oil enriched in omega-3 fatty acids on the development and maintenance of neuropathic pain. J. Pharm. Pharmacol. 2020, 72, 437–447. [Google Scholar] [CrossRef]
- Monroy Guízar, E.A.; García Benavides, L.; Ambriz Plascencia, A.R.; Pascoe González, S.; Totsuka Sutto, S.E.; Cardona Muñoz, E.G.; Méndez-del Villar, M. Effect of Alpha-Lipoic Acid on Clinical and Neurophysiologic Recovery of Carpal Tunnel Syndrome: A Double-Blind, Randomized Clinical Trial. J. Med. Food 2018, 21, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Oktay, K.N.K.; Akpinar, P.; Ozkan, F.U.; Aktas, I.; Canillioglu, Y.E.; Senturk, G.E. The effect of alpha lipoic acid on the recovery of sciatic nerve injury in rats: A prospective randomized controlled trial. Am. J. Phys. Med. Rehabil. 2020. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.V.; Oliveira, J.T.; Santos, B.L.R.; Dias, F.C.; Martinez, A.M.B.; Lima, C.K.F.; Miranda, A.L.P. Long-chain omega-3 fatty acids supplementation accelerates nerve regeneration and prevents neuropathic pain behavior in mice. Front. Pharmacol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Demir, R.; Yayla, M.; Akpinar, E.; Cakir, M.; Calikoglu, C.; Ozel, L.; Ozdemir, G.; Mercantepe, T. Protective effects of alpha-lipoic acid on experimental sciatic nerve crush injury in rats: Assessed with functional, molecular and electromicroscopic analyses. Int. J. Neurosci. 2014, 124, 935–943. [Google Scholar] [CrossRef]
- Senoglu, M.; Nacitarhan, V.; Kurutas, E.B.; Senoglu, N.; Altun, I.; Atli, Y.; Ozbag, D. Intraperitoneal Alpha-Lipoic Acid to prevent neural damage after crush injury to the rat sciatic nerve. J. Brachial Plex. Peripher. Nerve Inj. 2009, 4. [Google Scholar] [CrossRef] [PubMed]
- Ramli, D.; Aziz, I.; Mohamad, M.; Abdulahi, D.; Sanusi, J. The Changes in Rats with Sciatic Nerve Crush Injury Supplemented with Evening Primrose Oil: Behavioural, Morphologic, and Morphometric Analysis. Evid.-Based Complementary Altern. Med. 2017. [Google Scholar] [CrossRef]
- Bazinet, R.P.; Layé, S. Polyunsaturated fatty acids and their metabolites in brain function and disease. Nat. Rev. Neurosci. 2014, 15, 771–785. [Google Scholar] [CrossRef]
- Bazan, N.G. Neuroprotectin D1-mediated anti-inflammatory and survival signaling in stroke, retinal degenerations, and Alzheimer’s disease. J. Lipid Res. 2009, 50, S400–S405. [Google Scholar] [CrossRef]
- Bazan, N.G. Cellular and molecular events mediated by docosahexaenoic acid-derived neuroprotectin D1 signaling in photoreceptor cell survival and brain protection. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 205–211. [Google Scholar] [CrossRef]
- Sijben, J.W.C.; Calder, P.C. Differential immunomodulation with long-chain n-3 PUFA in health and chronic disease. Proc. Nutr. Soc. 2007, 66, 237–259. [Google Scholar] [CrossRef]
- Huang, C.T.; Tsai, Y.J. Docosahexaenoic acid confers analgesic effects after median nerve injury via inhibition of c-Jun N-terminal kinase activation in microglia. J. Nutr. Biochem. 2016, 29, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Robson, L.G.; Dyall, S.; Sidloff, D.; Michael-Titus, A.T. Omega-3 polyunsaturated fatty acids increase the neurite outgrowth of rat sensory neurones throughout development and in aged animals. Neurobiol. Aging 2010, 31, 678–687. [Google Scholar] [CrossRef]
- Pham, T.L.; He, J.; Kakazu, A.H.; Jun, B.; Bazan, N.G.; Bazan, H.E.P. Defining a mechanistic link between pigment epithelium–derived factor, docosahexaenoic acid, and corneal nerve regeneration. J. Biol. Chem. 2017, 292, 18486–18499. [Google Scholar] [CrossRef] [PubMed]
- Gerbi, A.; Sennoune, S.; Pierre, S.; Sampol, J.; Raccah, D.; Vague, P.; Maixent, J.M. Localization of Na,K-ATPase α/β isoforms in rat sciatic nerves: Effect of diabetes and fish oil treatment. J. Neurochem. 1999, 73, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.; Bender, D.; Botham, K.; Kennelly, P.; Rodwell, V.; Weill, P. Micronutrients, Vitamins & Minerals. In Harper’s Illustrated Biochemistry; McGraw Hill Medical: New York, NY, USA, 2012; pp. 467–478. [Google Scholar]
- de Oliveira, M.R. The neurotoxic effects of vitamin A and retinoids. An. Acad. Bras. Cienc. 2015, 87, 1361–1373. [Google Scholar] [CrossRef]
- Olson, C.R.; Mello, C.V. Significance of vitamin A to brain function, behavior and learning. Mol. Nutr. Food Res. 2010, 54, 489–495. [Google Scholar] [CrossRef]
- Al-saaeed, S.M.; Al-khalisy, M.H. The Regenerative Role of Vitamins B1, B6, B12 in Treatment of Peripheral Neuropathy. Int. J. Sci. Res. 2015, 6, 2319–7064. [Google Scholar] [CrossRef]
- Mey, J.; McCaffery, P. Retinoic acid signaling in the nervous system of adult vertebrates. Neuroscientist 2004, 10, 409–421. [Google Scholar] [CrossRef]
- Fragoso, Y.D.; Stoney, P.N.; McCaffery, P.J. The evidence for a beneficial role of vitamin A in multiple sclerosis. CNS Drugs 2014, 28, 291–299. [Google Scholar] [CrossRef]
- See, A.W.M.; Clagett-Dame, M. The temporal requirement for vitamin A in the developing eye: Mechanism of action in optic fissure closure and new roles for the vitamin in regulating cell proliferation and adhesion in the embryonic retina. Dev. Biol. 2009, 325, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Stoney, P.N.; McCaffery, P. A Vitamin on the mind: New discoveries on control of the brain by Vitamin, A. World Rev. Nutr. Diet. 2016, 115, 98–108. [Google Scholar] [CrossRef]
- Shearer, K.D.; Stoney, P.N.; Morgan, P.J.; McCaffery, P.J. A vitamin for the brain. Trends Neurosci. 2012, 35, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Latasa, M.J.; Cosgaya, J.M. Regulation of retinoid receptors by retinoic acid and axonal contact in Schwann cells. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Latasa, M.J.; Ituero, M.; Moran-Gonzalez, A.; Aranda, A.; Cosgaya, J.M. Retinoic acid regulates myelin formation in the peripheral nervous system. Glia 2010, 58, 1451–1464. [Google Scholar] [CrossRef]
- Zhelyaznik, N.; Schrage, K.; McCaffery, P.; Mey, J. Activation of retinoic acid signalling after sciatic nerve injury: Up-regulation of cellular retinoid binding proteins. Eur. J. Neurosci. 2003, 18, 1033–1040. [Google Scholar] [CrossRef]
- Zhelyaznik, N.; Mey, J. Regulation of retinoic acid receptors α, β and retinoid X receptor α after sciatic nerve injury. Neuroscience 2006, 141, 1761–1774. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, I. Vitamin D Metabolism and Guidelines for Vitamin D Supplementation. Clin. Biochem. Rev. 2020, 41, 103–126. [Google Scholar] [CrossRef] [PubMed]
- Gabaj, N.N.; Unic, A.; Miler, M.; Pavicic, T.; Culej, J.; Bolanca, I.; Mahecic, D.H.; Kopcinovic, L.M.; Vrtaric, A. In sickness and in health: Pivotal role of vitamin D. Biochem. Med. 2020, 30, 1–13. [Google Scholar]
- Jones, G. The discovery and synthesis of the nutritional factor vitamin D. Int. J. Paleopathol. 2018, 23, 96–99. [Google Scholar] [CrossRef]
- Lopez, A.G.; Kerlan, V.; Desailloud, R. Non-classical effects of vitamin D: Non-bone effects of vitamin D. Ann. Endocrinol. 2021, 82, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Zittermann, A.; Trummer, C.; Theiler-schwetz, V.; Lerchbaum, E.; März, W.; Pilz, S. Vitamin D and cardiovascular disease: An updated narrative review. Int. J. Mol. Sci. 2021, 22, 2896. [Google Scholar] [CrossRef]
- Zhang, Y.; Tan, H.; Tang, J.; Li, J.; Chong, W.; Hai, Y.; Feng, Y.; Lunsford, L.D.; Xu, P.; Jia, D.; et al. Effects of vitamin D supplementation on prevention of type 2 diabetes in patients with prediabetes: A systematic review and meta-analysis. Diabetes Care 2020, 43, 1650–1658. [Google Scholar] [CrossRef]
- Yammine, K.; Wehbe, R.; Assi, C. A systematic review on the efficacy of vitamin D supplementation on diabetic peripheral neuropathy. Clin. Nutr. 2020, 39, 2970–2974. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fang, F.; Tang, J.; Jia, L.; Feng, Y.; Xu, P.; Faramand, A. Association between Vitamin D supplementation and mortality: Systematic review and meta-analysis. BMJ 2019, 366, l4673. [Google Scholar] [CrossRef] [PubMed]
- Eleni, A.; Panagiotis, P. A systematic review and meta-analysis of vitamin D and calcium in preventing osteoporotic fractures. Clin. Rheumatol. 2020, 39, 3571–3579. [Google Scholar] [CrossRef]
- Jolliffe, D.A.; Camargo, C.A.; Sluyter, J.D.; Aglipay, M.; Aloia, J.F.; Ganmaa, D.; Bergman, P.; Bischoff-Ferrari, H.A.; Borzutzky, A.; Damsgaard, C.T.; et al. Vitamin D supplementation to prevent acute respiratory infections: A systematic review and meta-analysis of aggregate data from randomised controlled trials. Lancet Diabetes Endocrinol. 2021. [Google Scholar] [CrossRef]
- Głąbska, D.; Kołota, A.; Lachowicz, K.; Skolmowska, D.; Stachoń, M.; Guzek, D. The influence of vitamin d intake and status on mental health in children: A systematic review. Nutrients 2021, 13, 952. [Google Scholar] [CrossRef]
- Krishnan, A.V.; Feldman, D. Mechanisms of the anti-cancer and anti-inflammatory actions of vitamin D. Annu. Rev. Pharmacol. Toxicol. 2011, 51, 311–336. [Google Scholar] [CrossRef]
- Chabas, J.F.; Alluin, O.; Rao, G.; Garcia, S.; Lavaut, M.N.; Risso, J.J.; Legre, R.; Magalon, G.; Khrestchatisky, M.; Marqueste, T.; et al. Vitamin D2 potentiates axon regeneration. J. Neurotrauma 2008, 25, 1247–1256. [Google Scholar] [CrossRef]
- Naveilhan, P.; Neveu, I.; Wion, D.; Brachet, P. 1,25-Dihydroxyvitamin D3, an inducer of glial cell line-derived neurotrophic factor. Neuroreport 1996, 7, 2171–2175. [Google Scholar] [CrossRef]
- Neveu, I.; Naveilhan, P.; Jehan, F.; Baudet, C.; Wion, D.; De Luca, H.F.; Brachet, P. 1,25-Dihydroxyvitamin D3 regulates the synthesis of nerve growth factor in primary cultures of glial cells. Mol. Brain Res. 1994, 24, 70–76. [Google Scholar] [CrossRef]
- Wion, D.; Macgrogan, D.; Neveu, I.; Jehan, F.; Houlgatte, R.; Brachet, P. 1,25-Dihydroxyvitamin D3 is a potent inducer of nerve growth factor synthesis. J. Neurosci. Res. 1991, 28, 110–114. [Google Scholar] [CrossRef]
- Chabas, J.F.; Stephan, D.; Marqueste, T.; Garcia, S.; Lavaut, M.N.; Nguyen, C.; Legre, R.; Khrestchatisky, M.; Decherchi, P.; Feron, F. Cholecalciferol (Vitamin D3) Improves Myelination and Recovery after Nerve Injury. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Albay, C.; Adanir, O.; Kahraman Akkalp, A.; Dogan, V.B.; Gulec, M.A.; Beytemur, O. Cyanocobalamin and cholecalciferol synergistically improve functional and histopathological nerve healing in experimental rat model. Ideggyogy. Sz. 2020, 73, 113–120. [Google Scholar] [CrossRef]
- Montava, M.; Garcia, S.; Mancini, J.; Jammes, Y.; Courageot, J.; Lavieille, J.P.; Feron, F. Vitamin D3 potentiates myelination and recovery after facial nerve injury. Eur. Arch. Oto-Rhino-Laryngol. 2015, 272, 2815–2823. [Google Scholar] [CrossRef]
- Azizi, A.; Azizi, S.; Heshmatian, B.; Amini, K. Improvement of functional recovery of transected peripheral nerve by means of chitosan grafts filled with vitamin E, pyrroloquinoline quinone and their combination. Int. J. Surg. 2014, 12, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Enrione, E.B.; Weeks, O.I.; Kranz, S.; Shen, J. A vitamin E-deficient diet affects nerve regeneration in rats. Nutrition 1999, 15, 140–144. [Google Scholar] [CrossRef]
- Cecchini, T.; Cuppini, R.; Ciaroni, S.; De Matteis, R.; Grande, P. Del Increased number of sciatic sensory neurons in vitamin-e-deficient rats. Somatosens. Mot. Res. 1994, 11, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Cuppini, R.; Cecchini, T.; Ciaroni, S.; Ambrogini, P.; Grande, P. Del Nodal and terminal sprouting by regenerating nerve in vitamin E-deficient rats. J. Neurol. Sci. 1993, 117, 61–67. [Google Scholar] [CrossRef]
- Cuppini, R.; Cecchini, T.; Cuppini, C.; Ciaroni, S.; Del Grande, P. Time course of sprouting during muscle reinnervation in vitamin E-deficient rats. Muscle Nerve 1990, 13, 1027–1031. [Google Scholar] [CrossRef]
- Sundl, I.; Murkovic, M.; Bandoniene, D.; Winklhofer-Roob, B.M. Vitamin E content of foods: Comparison of results obtained from food composition tables and HPLC analysis. Clin. Nutr. 2007, 26, 145–153. [Google Scholar] [CrossRef]
- McLaughlin, P.J.; Weihrauch, J.L. Vitamin E content of foods. J. Am. Diet. Assoc. 1979, 75, 647–665. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.W. Vitamin E: Molecular and biological function. Proc. Nutr. Soc. 1994, 53, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Southam, E.; Thomas, P.K.; King, R.H.M.; Goss-sampson, M.A.; Muller, D.P.R. Experimental vitamin e deficiency in rats: Morphological and functional evidence of abnormal axonal transport secondary to free radical damage. Brain 1991, 114, 915–936. [Google Scholar] [CrossRef] [PubMed]
- Urano, S.; Asai, Y.; Makabe, S.; Matsuo, M.; Izumiyama, N.; Ohtsubo, K.; Endo, T. Oxidative injury of synapse and alteration of antioxidative defense systems in rats, and its prevention by vitamin E. Eur. J. Biochem. 1997, 245, 64–70. [Google Scholar] [CrossRef]
- Jiang, Q. Natural forms of vitamin E: Metabolism, antioxidant, and anti-inflammatory activities and their role in disease prevention and therapy. Free Radic. Biol. Med. 2014, 72, 76–90. [Google Scholar] [CrossRef]
- Traber, M.G. Vitamin E inadequacy in humans: Causes and consequences. Adv. Nutr. 2014, 5, 503–514. [Google Scholar] [CrossRef]
- Abraham, A.; Kattoor, A.J.; Saldeen, T.; Mehta, J.L. Vitamin E and its anticancer effects. Crit. Rev. Food Sci. Nutr. 2019, 59, 2831–2838. [Google Scholar] [CrossRef]
- Lewis, E.D.; Meydani, S.N.; Wu, D. Regulatory role of vitamin E in the immune system and inflammation. IUBMB Life 2019, 71, 487–494. [Google Scholar] [CrossRef]
- Lee, G.Y.; Han, S.N. The role of vitamin E in immunity. Nutrients 2018, 10, 1614. [Google Scholar] [CrossRef]
- Cecchini, T.; Cuppini, R.; Ciaroni, S.; Grande, P. Del Increased number of dorsal root ganglion neurons in vitamin-e-deficient rats. Somatosens. Mot. Res. 1993, 10, 433–443. [Google Scholar] [CrossRef]
- Cecchini, T.; Cuppini, R.; Ciaroni, S.; Barili, P.; De Matteis, R.; Grande, P. Del Changes in the number of primary sensory neurons in normal and vitamin-e-deficient rats during aging. Somatosens. Mot. Res. 1995, 12, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Simes, D.C.; Viegas, C.S.B.; Araújo, N.; Marreiros, C. Vitamin K as a Diet Supplement with Impact in Human Health: Current Evidence in Age-Related Diseases. Nutrients 2020, 12, 138. [Google Scholar] [CrossRef] [PubMed]
- Willems, B.A.G.; Vermeer, C.; Reutelingsperger, C.P.M.; Schurgers, L.J. The realm of vitamin K dependent proteins: Shifting from coagulation toward calcification. Mol. Nutr. Food Res. 2014, 58, 1620–1635. [Google Scholar] [CrossRef]
- Simes, D.C.; Viegas, C.S.B.; Araújo, N.; Marreiros, C. Vitamin K as a Powerful Micronutrient in Aging and Age-Related Diseases: Pros and Cons from Clinical Studies. Int. J. Mol. Sci. 2019, 20, 4150. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, M.; Mereu, M.C.; Aghi, A.; Iervasi, G.; Gallieni, M. Vitamin K and bone. Clin. Cases Miner. Bone Metab. 2017, 14, 200–206. [Google Scholar] [CrossRef]
- Fusaro, M.; Cianciolo, G.; Brandi, M.L.; Ferrari, S.; Nickolas, T.L.; Tripepi, G.; Plebani, M.; Zaninotto, M.; Iervasi, G.; La Manna, G.; et al. Vitamin K and Osteoporosis. Nutrients 2020, 12, 3625. [Google Scholar] [CrossRef]
- Lasemi, R.; Kundi, M.; Moghadam, N.B.; Moshammer, H.; Hainfellner, J.A. Vitamin K2 in multiple sclerosis patients. Wien. Klin. Wochenschr. 2018, 130, 307–313. [Google Scholar] [CrossRef]
- Sakaue, M.; Mori, N.; Okazaki, M.; Kadowaki, E.; Kaneko, T.; Hemmi, N.; Sekiguchi, H.; Maki, T.; Ozawa, A.; Hara, S.; et al. Vitamin K has the potential to protect neurons from methylmercury-induced cell death In Vitro. J. Neurosci. Res. 2011, 89, 1052–1058. [Google Scholar] [CrossRef]
- Popescu, D.C.; Huang, H.; Singhal, N.K.; Shriver, L.; McDonough, J.; Clements, R.J.; Freeman, E.J. Vitamin K enhances the production of brain sulfatides during remyelination. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Goudarzi, S.; Rivera, A.; Butt, A.M.; Hafizi, S. Gas6 Promotes Oligodendrogenesis and Myelination in the Adult Central Nervous System and After Lysolecithin-Induced Demyelination. ASN Neuro 2016, 8. [Google Scholar] [CrossRef]
- Iwamoto, J.; Matsumoto, H.; Takeda, T.; Sato, Y.; Yeh, J.K. Effects of vitamin K2 on cortical and cancellous bone mass, cortical osteocyte and lacunar system, and porosity in sciatic neurectomized rats. Calcif. Tissue Int. 2010, 87, 254–262. [Google Scholar] [CrossRef]
- Tsang, C.K.; Kamei, Y. Novel effect of vitamin K1 (phylloquinone) and vitamin K2 (menaquinone) on promoting nerve growth factor-mediated neurite outgrowth from PC12D cells. Neurosci. Lett. 2002, 323, 9–12. [Google Scholar] [CrossRef]
- Fernández-Villa, D.; Jiménez Gómez-Lavín, M.; Abradelo, C.; Román, J.S.; Rojo, L. Molecular Sciences Tissue Engineering Therapies Based on Folic Acid and Other Vitamin B Derivatives. Functional Mechanisms and Current Applications in Regenerative Medicine. Int. J. Mol. Sci. 2018, 19, 4068. [Google Scholar] [CrossRef] [PubMed]
- Wendołowicz, A.; Stefańska, E.; Ostrowska, L. Influence of selected dietary components on the functioning of the human nervous system. Rocz. Panstw. Zakl. Hig. 2018, 69, 15–21. [Google Scholar]
- Altun, I.; Kurutaş, E.B. Vitamin B complex and vitamin B12 levels after peripheral nerve injury. Neural Regen. Res. 2016, 11, 842–845. [Google Scholar] [CrossRef] [PubMed]
- Al-Saaeed, S.M.; Ali, H.A.; Ali, S.M.; Ali, S.A. Vitamins B therapy in Regeneration of Peripheral Neuropathy Associated with Lipid Profile. In Proceedings of the Journal of Physics: Conference Series; Institute of Physics Publishing: Bristol, UK, 2019; Volume 1279. [Google Scholar]
- Ehmedah, A.; Nedeljkovic, P.; Dacic, S.; Repac, J.; Draskovic-Pavlovic, B.; Vučević, D.; Pekovic, S.; Nedeljkovic, B.B. Effect of Vitamin B Complex Treatment on Macrophages to Schwann Cells Association during Neuroinflammation after Peripheral Nerve Injury. Molecules 2020, 25, 5426. [Google Scholar] [CrossRef]
- Sağır, D.; Kuruoğlu, E.; Onger, M.E.; Yarar, E. The Effects of folic acid and topiramate on peripheral nerve regeneration. Biotech. Histochem. 2020, 95, 105–112. [Google Scholar] [CrossRef]
- Kang, W.B.; Chen, Y.J.; Lu, D.Y.; Yan, J.Z. Folic acid contributes to peripheral nerve injury repair by promoting Schwann cell proliferation, migration, and secretion of nerve growth factor. Neural Regen. Res. 2019, 14, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Nedeljković, P.; Dacić, S.; Kovačević, M.; Peković, S.; Vučević, D.; Božić-Nedeljković, B. Vitamin B complex as a potential therapeutical modality in combating peripheral nerve injury. Acta Med. Med. 2018, 57, 85–91. [Google Scholar] [CrossRef]
- Harma, A.; Sahin, M.S.; Zorludemir, S. Effects of intraperitoneally administered folic acid on the healing of repaired tibial nerves in rats. J. Reconstr. Microsurg. 2015, 31, 191–197. [Google Scholar] [CrossRef]
- Liao, W.C.; Wang, Y.J.; Huang, M.C.; Tseng, G.F. Methylcobalamin Facilitates Collateral Sprouting of Donor Axons and Innervation of Recipient Muscle in End-to-Side Neurorrhaphy in Rats. PLoS ONE 2013, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Yang, T.; Li, Q.; Zhu, Z.; Wang, L.; Bai, G.; Li, D.; Li, Q.; Wang, W. Dexamethasone and vitamin B12 synergistically promote peripheral nerve regeneration in rats by upregulating the expression of brain-derived neurotrophic factor. Arch. Med. Sci. 2012, 8, 924–930. [Google Scholar] [CrossRef]
- Okada, K.; Tanaka, H.; Temporin, K.; Okamoto, M.; Kuroda, Y.; Moritomo, H.; Murase, T.; Yoshikawa, H. Methylcobalamin increases Erk1/2 and Akt activities through the methylation cycle and promotes nerve regeneration in a rat sciatic nerve injury model. Exp. Neurol. 2010, 222, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xu, Y.; Wang, X.; Liu, J.; Hu, X.; Tan, D.; Li, Z.; Guo, J. Ascorbic acid accelerates Wallerian degeneration after peripheral nerve injury. Neural Regen. Res. 2021, 16, 1078–1085. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, Y.; Fan, Z.; Wang, X.; Li, Z.; Wen, J.; Deng, J.; Tan, D.; Pan, M.; Hu, X.; et al. Ascorbic acid facilitates neural regeneration after sciatic nerve crush injury. Front. Cell. Neurosci. 2019, 13. [Google Scholar] [CrossRef]
- Bâ, A. Metabolic and structural role of thiamine in nervous tissues. Cell. Mol. Neurobiol. 2008, 28, 923–931. [Google Scholar] [CrossRef]
- Calderón-Ospina, C.A.; Nava-Mesa, M.O. B Vitamins in the nervous system: Current knowledge of the biochemical modes of action and synergies of thiamine, pyridoxine, and cobalamin. CNS Neurosci. Ther. 2020, 26, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Hilal Ahmad, S.; Ahmad, S.B.; Jan, A. Thiamine deficiency related peripheral neuropathy in peripartum women of Kashmir, India: A hospital based study. Int. J. Adv. Med. 2019, 6, 662. [Google Scholar] [CrossRef]
- Nemazannikova, N.; Mikkelsen, K.; Stojanovska, L.; Blatch, G.L.; Apostolopoulos, V. Is there a Link between Vitamin B and Multiple Sclerosis? Med. Chem. 2018, 14, 170–180. [Google Scholar] [CrossRef]
- Mooney, S.; Leuendorf, J.-E.; Hendrickson, C.; Hellmann, H. Vitamin B6: A Long Known Compound of Surprising Complexity. Molecules 2009, 14, 329–351. [Google Scholar] [CrossRef] [PubMed]
- Hellmann, H.; Mooney, S. Vitamin B6: A Molecule for Human Health? Molecules 2010, 15, 442–459. [Google Scholar] [CrossRef] [PubMed]
- Aufiero, E.; Stitik, T.P.; Foye, P.M.; Chen, B. Pyridoxine Hydrochloride Treatment of Carpal Tunnel Syndrome: A Review. Nutr. Rev. 2004, 62, 96–104. [Google Scholar] [CrossRef]
- Nishimoto, S.; Tanaka, H.; Okamoto, M.; Okada, K.; Murase, T.; Yoshikawa, H. Methylcobalamin promotes the differentiation of Schwann cells and remyelination in lysophosphatidylcholine-induced demyelination of the rat sciatic nerve. Front. Cell. Neurosci. 2015, 9, 1–13. [Google Scholar] [CrossRef]
- Eldridge, C.F.; Bunge, M.B.; Bunge, R.P.; Wood, P.M. Differentiation of axon-related Schwann cells in vitro. I. Ascorbic acid regulates basal lamina assembly and myelin formation. J. Cell Biol. 1987, 105, 1023–1034. [Google Scholar] [CrossRef]
- Huff, T.C.; Sant, D.W.; Camarena, V.; Van Booven, D.; Andrade, N.S.; Mustafi, S.; Monje, P.V.; Wang, G. Vitamin C regulates Schwann cell myelination by promoting DNA demethylation of pro-myelinating genes. J. Neurochem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Gess, B.; Röhr, D.; Fledrich, R.; Sereda, M.W.; Kleffner, I.; Humberg, A.; Nowitzki, J.; Strecker, J.K.; Halfter, H.; Young, P. Sodium-dependent vitamin C transporter 2 deficiency causes hypomyelination and extracellular matrix defects in the peripheral nervous system. J. Neurosci. 2011, 31, 17180–17192. [Google Scholar] [CrossRef] [PubMed]
- Gess, B.; Lohmann, C.; Halfter, H.; Young, P. Sodium-dependent vitamin C transporter 2 (SVCT2) is necessary for the uptake of L-ascorbic acid into schwann cells. Glia 2010, 58, 287–299. [Google Scholar] [CrossRef]
- Röhr, D.; Halfter, H.; Schulz, J.B.; Young, P.; Gess, B. Sodium-dependent Vitamin C transporter 2 deficiency impairs myelination and remyelination after injury: Roles of collagen and demethylation. Glia 2017, 65, 1186–1200. [Google Scholar] [CrossRef]
- Chin, K.-Y.; Ima-Nirwana, S. Vitamin C and Bone Health: Evidence from Cell, Animal and Human Studies. Curr. Drug Targets 2018, 19, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Granger, M.; Eck, P. Dietary Vitamin C in Human Health. In Advances in Food and Nutrition Research; Academic Press Inc.: Cambridge, MA, USA, 2018; Volume 83, pp. 281–310. [Google Scholar]
- Pareyson, D.; Reilly, M.M.; Schenone, A.; Fabrizi, G.M.; Cavallaro, T.; Manganelli, L.; Vita, G.; Quattrone, A.; Padua, L.; Gemignani, F.; et al. Ascorbic acid in charcot-marie-tooth disease type 1A (CMTTRIAAL and CMT-TRAUK): A double-blind randomised trial. Lancet Neurol. 2011, 10, 320–328. [Google Scholar] [CrossRef]
- Volpe, S.L. Magnesium in disease prevention and overall health. Adv. Nutr. 2013, 4. [Google Scholar] [CrossRef]
- de Baaij, J.H.F.; Hoenderop, J.G.J.; Bindels, R.J.M. Magnesium in man: Implications for health and disease. Physiol. Rev. 2015, 95, 1–46. [Google Scholar] [CrossRef]
- Pan, H.C.; Sheu, M.L.; Su, H.L.; Chen, Y.J.; Chen, C.J.; Yang, D.Y.; Chiu, W.T.; Cheng, F.C. Magnesium supplement promotes sciatic nerve regeneration and down-regulates inflammatory response. Magnes. Res. 2011, 24, 54–70. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Cheng, F.C.; Chen, C.J.; Su, H.L.; Sheu, M.L.; Sheehan, J.; Pan, H.C. Down-regulated expression of magnesium transporter genes following a high magnesium diet attenuates sciatic nerve crush injury. Neurosurgery 2019, 84, 965–976. [Google Scholar] [CrossRef]
- Kapoukranidou, D.; Gougoulias, N.; Hatzisotiriou, A.; Fardi, D.; Albani, M.; Kalpidis, I. Assessment of motoneuron death during development following neonatal nerve crush and Mg2+ treatment. Med. Sci. Monit. 2005, 11, BR373–BR379. [Google Scholar]
- Greensmith, L.; Mooney, E.C.; Waters, H.J.; Houlihan-Burne, D.G.; Lowrie, M.B. Magnesium ions reduce motoneuron death following nerve injury or exposure to N-methyl-d-aspartate in the developing rat. Neuroscience 1995, 68, 807–812. [Google Scholar] [CrossRef]
- Gougoulias, N.; Hatzisotiriou, A.; Kapoukranidou, D.; Albani, M. Magnesium administration provokes motor unit survival, after sciatic nerve injury in neonatal rats. BMC Musculoskelet. Disord. 2004, 5. [Google Scholar] [CrossRef]
- Hopkins, T.M.; Little, K.J.; Vennemeyer, J.J.; Triozzi, J.L.; Turgeon, M.K.; Heilman, A.M.; Minteer, D.; Marra, K.; Hom, D.B.; Pixley, S.K. Short and long gap peripheral nerve repair with magnesium metal filaments. J. Biomed. Mater. Res. Part A 2017, 105, 3148–3158. [Google Scholar] [CrossRef] [PubMed]
- Li, B.H.; Yang, K.; Wang, X. Biodegradable magnesium wire promotes regeneration of compressed sciatic nerves. Neural Regen. Res. 2016, 11, 2012–2017. [Google Scholar] [CrossRef]
- Vennemeyer, J.J.; Hopkins, T.; Hershcovitch, M.; Little, K.D.; Hagen, M.C.; Minteer, D.; Hom, D.B.; Marra, K.; Pixley, S.K. Initial observations on using magnesium metal in peripheral nerve repair. J. Biomater. Appl. 2015, 29, 1145–1154. [Google Scholar] [CrossRef]
- Kızılay, Z.; Erken, H.A.; Aktaş, S.; Ersoy, N.; Abas, B.İ.; Topcu, A.; Yenisey, C.; İsmailoğlu, Ö. Effects of selenium on axon and myelin healing in an experimental sciatic nerve injury model. İstanbul Tıp Fakültesi Derg. 2017, 80, 138–183. [Google Scholar] [CrossRef]
- Qian, Y.; Cheng, Y.; Song, J.; Xu, Y.; Yuan, W.E.; Fan, C.; Zheng, X. Mechano-Informed Biomimetic Polymer Scaffolds by Incorporating Self-Powered Zinc Oxide Nanogenerators Enhance Motor Recovery and Neural Function. Small 2020, 16. [Google Scholar] [CrossRef] [PubMed]
- Iman, M.; Araghi, M.; Panahi, Y.; Mohammadi, R. Effects of Chitosan-Zinc Oxide Nanocomposite Conduit on Transected Sciatic Nerve: An Animal Model Study. Bull. Emerg. Trauma 2017, 5, 240–248. [Google Scholar] [CrossRef]
- Salehi, M.; Naseri-Nosar, M.; Ebrahimi-Barough, S.; Nourani, M.; Vaez, A.; Farzamfar, S.; Ai, J. Regeneration of sciatic nerve crush injury by a hydroxyapatite nanoparticle-containing collagen type I hydrogel. J. Physiol. Sci. 2018, 68, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Burk, R.F. Selenium, an antioxidant nutrient. Nutr. Clin. Care 2002, 5, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Tapiero, H.; Townsend, D.M.; Tew, K.D. The antioxidant role of selenium and seleno-compounds. Biomed. Pharmacother. 2003, 57, 134–144. [Google Scholar] [CrossRef]
- Avery, J.C.; Hoffmann, P.R. Selenium, selenoproteins, and immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef]
- Zendedel, A.; Gharibi, Z.; Anbari, K.; Abbaszadeh, A.; khayat, Z.K.; Khorramabadi, R.M.; Soleymaninejad, M.; Gholami, M. Selenium Ameliorate Peripheral Nerve Ischemic-Reperfusion Injury via Decreased TNF-α. Biol. Trace Elem. Res. 2017, 176, 328–337. [Google Scholar] [CrossRef]
- Zendedel, A.; Delavari, S.; Ahmadvand, H.; Ghanadi, K.; Gholami, M. Effects of Selenium on Antioxidant Activity and Recovery From Sciatic Nerve Ischemia-Reperfusion in Adult Rats. Zahedan J. Res. Med. Sci. 2015, 17, 5200. [Google Scholar] [CrossRef]
- Levenson, C.W.; Morris, D. Zinc and neurogenesis: Making new neurons from development to adulthood. Adv. Nutr. 2011, 2, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Ünal, B.; Tan, H.; Orbak, Z.; Kiki, I.; Bilici, M.; Bilici, N.; Aslan, H.; Kaplan, S. Morphological alterations produced by zinc deficiency in rat sciatic nerve: A histological, electron microscopic, and stereological study. Brain Res. 2005, 1048, 228–234. [Google Scholar] [CrossRef]
- O’Dell, B.L.; Conley-Harrison, J.; Besch-Williford, C.; Browning, J.D.; O’Brien, D. Zinc status and peripheral nerve function in guinea pigs. FASEB J. 1990, 4, 2919–2922. [Google Scholar] [CrossRef]
- Aydemir Sezer, U.; Ozturk, K.; Aru, B.; Yanıkkaya Demirel, G.; Sezer, S.; Bozkurt, M.R. Zero valent zinc nanoparticles promote neuroglial cell proliferation: A biodegradable and conductive filler candidate for nerve regeneration. J. Mater. Sci. Mater. Med. 2017, 28. [Google Scholar] [CrossRef]
- Ekram, B.; Abd El-Hady, B.M.; El-Kady, A.M.; Fouad, M.T.; Sadek, Z.I.; Amr, S.M.; Gabr, H.; Waly, A.I.; Guirguis, O.W. Enhanced mesenchymal stem cells growth on antibacterial microgrooved electrospun zinc chloride/polycaprolactone conduits for peripheral nerve regeneration. J. Bioact. Compat. Polym. 2021, 36, 152–168. [Google Scholar] [CrossRef]
- Li, K.; Wang, X.F.; Li, D.Y.; Chen, Y.C.; Zhao, L.J.; Liu, X.G.; Guo, Y.F.; Shen, J.; Lin, X.; Deng, J.; et al. The good, the bad, and the ugly of calcium supplementation: A review of calcium intake on human health. Clin. Interv. Aging 2018, 13, 2443–2452. [Google Scholar] [CrossRef]
- Henley, J.; Poo, M.M. Guiding neuronal growth cones using Ca2+ signals. Trends Cell Biol. 2004, 14, 320–330. [Google Scholar] [CrossRef]
- Li, G.; Xiao, Q.; McNaughton, R.; Han, L.; Zhang, L.; Wang, Y.; Yang, Y. Nanoengineered porous chitosan/CaTiO3 hybrid scaffolds for accelerating Schwann cells growth in peripheral nerve regeneration. Colloids Surf. B Biointerfaces 2017, 158, 57–67. [Google Scholar] [CrossRef]
- Zargar Kharazi, A.; Dini, G.; Naser, R. Fabrication and evaluation of a nerve guidance conduit capable of Ca2+ ion release to accelerate axon extension in peripheral nerve regeneration. J. Biomed. Mater. Res. Part A 2018, 106, 2181–2189. [Google Scholar] [CrossRef]
- Nawrotek, K.; Tylman, M.; Rudnicka, K.; Balcerzak, J.; Kamiński, K. Chitosan-based hydrogel implants enriched with calcium ions intended for peripheral nervous tissue regeneration. Carbohydr. Polym. 2016, 136, 764–771. [Google Scholar] [CrossRef]
- Levi, S.; Taveggia, C. Iron homeostasis in peripheral nervous system, still a black box? Antioxidants Redox Signal. 2014, 21, 634–648. [Google Scholar] [CrossRef]
- Abbaspour, N.; Hurrell, R.; Kelishadi, R. Review on iron and its importance for human health. J. Res. Med. Sci. 2014, 19, 164–174. [Google Scholar] [PubMed]
- Santiago González, D.A.; Cheli, V.T.; Wan, R.; Paez, P.M. Iron Metabolism in the Peripheral Nervous System: The Role of DMT1, Ferritin, and Transferrin Receptor in Schwann Cell Maturation and Myelination. J. Neurosci. 2019, 39, 9940–9953. [Google Scholar] [CrossRef]
- Salis, C.; Setton, C.P.; Soto, E.F.; Pasquini, J.M. The mRNA of transferrin is expressed in Schwann cells during their maturation and after nerve injury. Exp. Neurol. 2007, 207, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Graeber, M.B.; Raivich, G.; Kreutzberg, G.W. Increase of transferrin receptors and iron uptake in regenerating motor neurons. J. Neurosci. Res. 1989, 23, 342–345. [Google Scholar] [CrossRef] [PubMed]
- Raivich, G.; Graeber, M.B.; Gehrmann, J.; Kreutzberg, G.W. Transferrin Receptor Expression and Iron Uptake in the Injured and Regenerating Rat Sciatic Nerve. Eur. J. Neurosci. 1991, 3, 919–927. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, S. Iron, hormesis, and protection in acute kidney injury. Kidney Int. 2016, 90, 16–17. [Google Scholar] [CrossRef]
- Amos-Kroohs, R.M.; Usach, V.; Piñero, G.; Vorhees, C.V.; Martinez Vivot, R.; Soto, P.A.; Williams, M.T.; Setton-Avruj, P. Metal bashing: Iron deficiency and manganese overexposure impact on peripheral nerves. J. Toxicol. Environ. Health Part A Curr. Issues 2019, 82, 99–112. [Google Scholar] [CrossRef] [PubMed]


| Reference | Type of Nerve Lesion and Animal Model | Description and Timing of the Diet |
|---|---|---|
| Ketogenic Diet | ||
| Li et al., 2020 [22] | Sciatic nerve crush (Sprague–Dawley rats) | 65.8% fat, 3.0% carbohydrate, and 18.1% protein for 8 weeks after injury |
| Mayr et al., 2020 [23] | Transection of the common peroneal and tibial nerves in mouse (C57/BL6 mice) | 75.1% fat, 3.2% carbohydrates, and 8.6% protein for 7 days before injury and up to 28 day after injury |
| Liśkiewicz et al., 2016 [24] | Sciatic nerve crush (Wistar rats) | 79% fat, 0.8% carbohydrates, and 9.5% protein for 6 weeks after injury. Preconditioned group received KD also for 3 weeks before injury |
| Caloric restriction | ||
| De Angelis et al., 2020 [25] | Chronic constriction injury of sciatic nerve in mouse (CD1 male mice 12 months old) | 40% less of daily consumption for 7 days after CCI |
| Coccurello et al., 2018 [26] | Chronic constriction injury (CCI) of sciatic nerve (Wild-type CD1 male mice, Ambra1+ transgenic CD1 male mice) | 40% less of daily consumption for 7 days after CCI |
| High-fat diet | ||
| Song et al., 2018 [27] | Plantar incision (Long-Evans rats) | 40% fat administrated with different timing: a group received a high-fat diet for 6 weeks before injury and 2 weeks after, another group was fed with high-fat diet for 7 weeks and then switched back to standard diet for 2 weeks before to plantar incision, while in the last group, only male rats were switched to high-fat diet 1 week before injury |
| Bekar et al., 2014 [28] | Sciatic nerve crush injury (Sprague–Dawley rats) | 40% fat for 3 months (started before injury and ended 4 weeks after injury) |
| Reference | Type of Nerve Lesion and Animal Model | Type and Timing of Administration/Experimental Groups |
|---|---|---|
| Omega-3 | ||
| Oktay et al., 2020 [53] | Sciatic nerve crush (Sprague–Dawley male rat) | 50 or 100 mg/kg intraperitoneal alpha-lipoic acid (ALA) injection was done daily after nerve injury for 14 days |
| Unda et al., 2020 [51] | Sciatic nerve chronic constriction injury (Wistar rats) | Saline solution with concentrated salmon oil ω-3 (0.72 g/kg) administered orally for 21 consecutive days |
| Silva et al., 2017 [54] | Partial sciatic nerve ligation model (Swiss mice) | DHA/EPA-concentrate fish oil 2.3 g/kg orally administered 5 days/week for 2 weeks after nerve injury |
| Demir et al., 2014 [55] | Sciatic nerve crush (adult female albino Wistar rats) | ALA was suspended in distilled water 1 mL (per rat and administered orally at a dosage of 25 or 50 mg/kg by gavage), daily for 30 days |
| Senoglu et al., 2009 [56] | Sciatic nerve crush (female Sprague–Dawley rats) | 100 mg/kg intraperitoneal ALA injection was done 24 and 1 h before crush injury |
| Omega-6 | ||
| Ramli et al., 2017 [57] | Sciatic nerve crush injury (Sprague–Dawley rats) | 6000 mg/day of evening primrose oil through an esophageal feeding tube starting from day 1 after nerve crush for 4 weeks |
| Reference | Type of Nerve Lesion and Animal Model | Type and Timing of Administration/Experimental Groups |
|---|---|---|
| Vitamin B complex | ||
| Ehmedah et al., 2020 [130] | Transection of the motor branch of femoral nerve followed by end-to-end repair (Albino Oxford adult male rats) | Intraperitoneal injection of vitamin B complex ampoules (2 mL) of Beviplex (Beviplex®, Galenika a.d. Belgrade, Serbia), each containing B1 (40 mg), B2 (4 mg), B3 (100 mg), B5 (10 mg), B6 (8 mg), and B12 (4 µg). Dose: 1.85 mL/kg/day. The first was injected immediately (15 min) after the surgery and then every 24 h from the day of the operation until the day of sacrifice. Sacrifice: 1, 3, 7, and 14 days after surgery. |
| Sağır et al., 2020 [131] | Sciatic nerve crush (Wistar male rats) | Daily folic acid intraperitoneal injection for 6 weeks (0.5 mg/kg/day) |
| Al-Saaeed et al., 2019 [129] | Sciatic nerve crush (Albino male rats) |
|
| Kang et al., 2019 [132] | End-to-end repair of the sciatic nerve (Sprague–Dawley male rats) | Intraperitoneal administration of 80 μg/kg folic acid for 7 days following nerve injury |
| Nedeljković et al., 2018 [133] | Transection of the motor branch of the femoral nerve with immediate reconstruction using a termino-terminal anastomosis (Albino Oxford rats) | Treatment with vitamin B complex (Beviplex®, Galenika a.d. Belgrade, Serbia). Each ampoule (2 mL) contains B1 (40 mg), B2 (4 mg), B3 (100 mg), B5 (10 mg), B6 (8 mg), and B12 (4 µg). The dose was 1.85 mL/kg/day. Administration by intraperitoneal injection daily from the day of injury until the day of sacrifice. Examination at 1, 3, 7, and 14 days after injury. |
| Al-Saaeed et al., 2015 [69] | Sciatic nerve crush (Albino rats) |
|
| Harma et al., 2015 [134] | Tibial nerves transection repaired by end-to-end repair (adult male Wistar Albino rats) | Intraperitoneal administration of 80 μg/kg folic acid daily for 6 weeks |
| Liao et al., 2013 [135] | End-to-side neurorrhaphy between ulnar and musculocutaneous nerves (Wistar male rats) | Methylcobalamin (500 µg/mL; Methylcobal® Eisai, Tokyo, Japan) 250 μg/day Daily intraperitoneal injection for 3 months |
| Sun et al., 2012 [136] | Sciatic nerve crush of the peroneal branch (Wistar male rats) | Injection into the injured site of vitamin B12 alone (2 mg/kg) or with dexamethasone (1 mg/kg), daily for 28 days |
| Okada et al., 2010 [137] | Sciatic nerve cut followed by direct end-to-end suture (Wistar female rats) | Continuous administration of methylcobalamin (1 mg/kg/day) in PBS for 12 weeks using an osmotic minipump placed subcutaneously in the right side of the back |
| Vitamin C | ||
| Li et al., 2021 [138] | Sciatic nerve transection (Specific-pathogen-free female Sprague–Dawley rats) | Intragastric administration of ascorbic acid solution (suspension with saline at a concentration of 20 mg/mL): 400 mg/kg the day of injury; 200 mg/kg the following days for 5 or 8 days |
| Li et al., 2019 [139] | Sciatic nerve crush (C57BL/6 mice) | Daily intragastric administration of ascorbic acid solution (suspension with saline at a concentration of 13.33 mg/mL): 400 mg/kg the day of injury; 200 mg/kg the following days for 3 or 28 days |
| Reference | Type of Nerve Lesion and Animal Model | Type of Administration/Experimental Groups |
|---|---|---|
| Magnesium | ||
| Cheng et al., 2019 [159] | Sciatic nerve crush (Sprague–Dawley rats) | High magnesium water (magnesium salt solution (1 mg/mL) after nerve injury) for 4 weeks |
| Hopkins et al., 2017 [163] | 6 mm sciatic nerve defect repaired with 8 mm long poly(caprolactone) (PCL) nerve conduit AND 15 mm sciatic nerve defect repaired with 17 mm long PCL conduit (Lewis male rats) | 10 mm long or 20 mm long Mg2+ filament was inserted inside conduits, extending two mm into both stumps |
| Li et al., 2016 [164] | Acute sciatic nerve compression performed using an 8 mm long silastic tube for 3 h. (Sprague–Dawley rats) | A biodegradable Mg-3%Al-1%Zn wire (diameter 3 mm) was put through the sciatic nerve epineurium |
| Vennemeyer et al., 2015 [165] | 6 mm sciatic nerve defect repaired with 8 mm long poly(caprolactone) (PCL) nerve conduit (Lewis male rats) | Mg filaments were embedded into the nerve stumps (2 mm deep) with the conduit threaded over all. The conduit lumens were filled via syringe with a keratin hydrogel or sterile saline. |
| Pan et al., 2011 [158] | Sciatic nerve crush (Imprinting control region mice) |
|
| Gougoulias et al., 2004 [162] | Sciatic nerve crush at the 2nd day postnatal (Wistar Albino rats) | Subcutaneous injection of MgSO47H2O (0.05 mL of 1 M solution/10 g body weight) daily from operation day until rats were 14 days old |
| Greensmith et al., 1995 [161] | Sciatic nerve crush at the day of birth (Wistar Albino rats) | Subcutaneous injection of MgSO4 (0.05 mL 1 M MgSO4/10 g body weight) daily from operation day until rats were 10 days old |
| Selenium | ||
| Kizilay et al., 2017 [166] | Sciatic nerve crush (Wistar albino rats) | Selenium was given at a dose of 1.5 mg/kg (dissolved in sterile saline) by oral gavage at 1st, 24th, 48th, and 72nd h after surgery |
| Zinc | ||
| Qian et al., 2020 [167] | 15 mm sciatic nerve gap repaired with ZnO-loaded PCL piezoelectric nanogenerator scaffold (Sprague–Dawley male rats) | Zinc oxide nanoparticles (purity 99%)-loaded PCL piezoelectric nanogenerator scaffold fabricated using 3D injectable multilayer moulding technique |
| Iman et al., 2017 [168] | 15 mm sciatic nerve gap repaired with 14 mm chitosan–zinc oxide nanocomposite conduit (White Wistar male rats) | Chitosan–zinc oxide nanocomposite conduit. Analysis at 4, 8, and 12 weeks after surgery |
| Calcium | ||
| Salehi et al., 2018 [169] | Sciatic nerve crush (Wistar male rats) | Local injection of 0.50 mL of a single batch of collagen + hydroxyapatite nanoparticles hydrogel to the site of crush injury using a 16-gauge needle |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Soury, M.; Fornasari, B.E.; Carta, G.; Zen, F.; Haastert-Talini, K.; Ronchi, G. The Role of Dietary Nutrients in Peripheral Nerve Regeneration. Int. J. Mol. Sci. 2021, 22, 7417. https://doi.org/10.3390/ijms22147417
El Soury M, Fornasari BE, Carta G, Zen F, Haastert-Talini K, Ronchi G. The Role of Dietary Nutrients in Peripheral Nerve Regeneration. International Journal of Molecular Sciences. 2021; 22(14):7417. https://doi.org/10.3390/ijms22147417
Chicago/Turabian StyleEl Soury, Marwa, Benedetta Elena Fornasari, Giacomo Carta, Federica Zen, Kirsten Haastert-Talini, and Giulia Ronchi. 2021. "The Role of Dietary Nutrients in Peripheral Nerve Regeneration" International Journal of Molecular Sciences 22, no. 14: 7417. https://doi.org/10.3390/ijms22147417
APA StyleEl Soury, M., Fornasari, B. E., Carta, G., Zen, F., Haastert-Talini, K., & Ronchi, G. (2021). The Role of Dietary Nutrients in Peripheral Nerve Regeneration. International Journal of Molecular Sciences, 22(14), 7417. https://doi.org/10.3390/ijms22147417