
Influence of the Anthocyanin and Cofactor Structure on the Formation Efficiency of Naturally Derived Pyranoanthocyanins




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
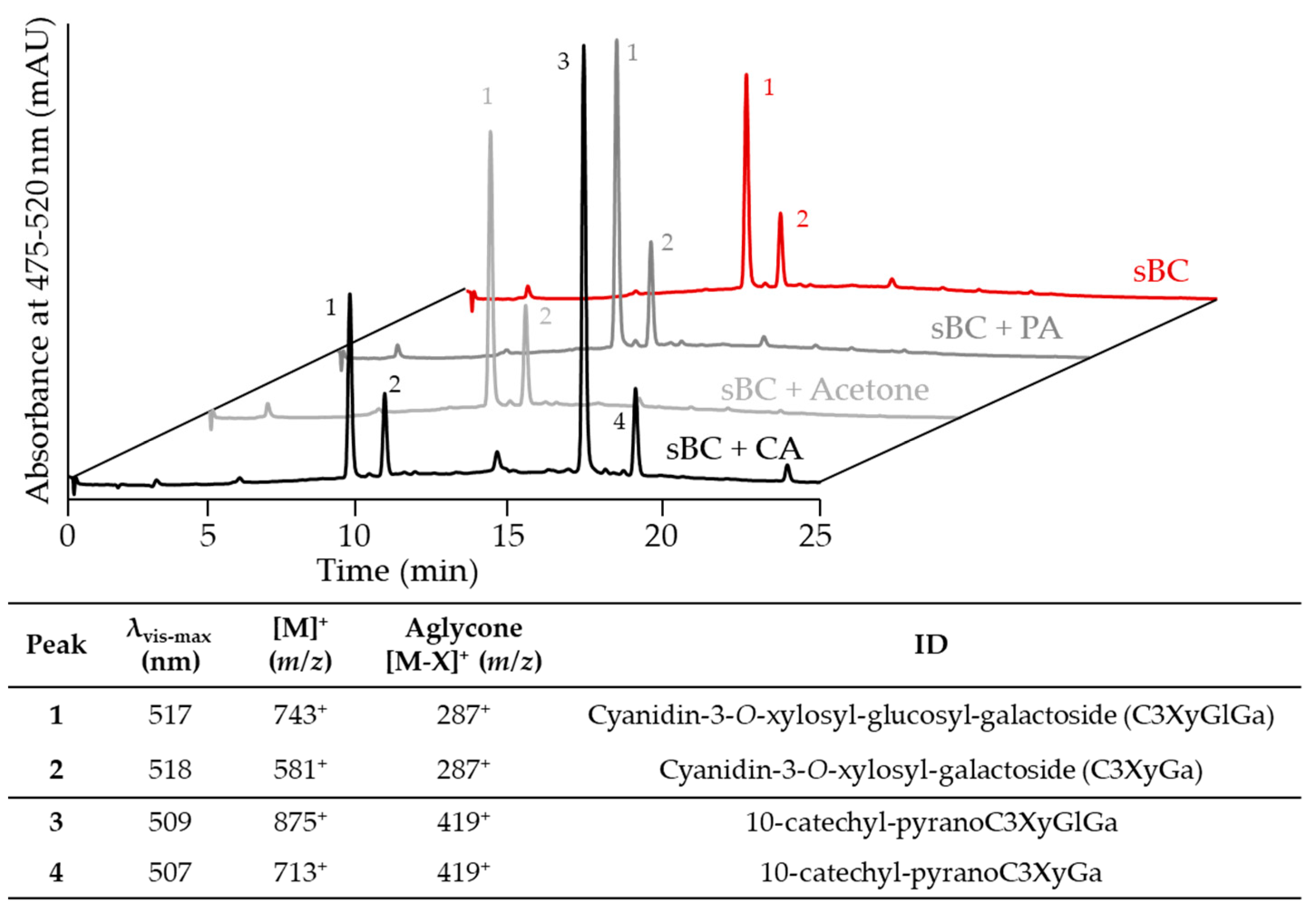
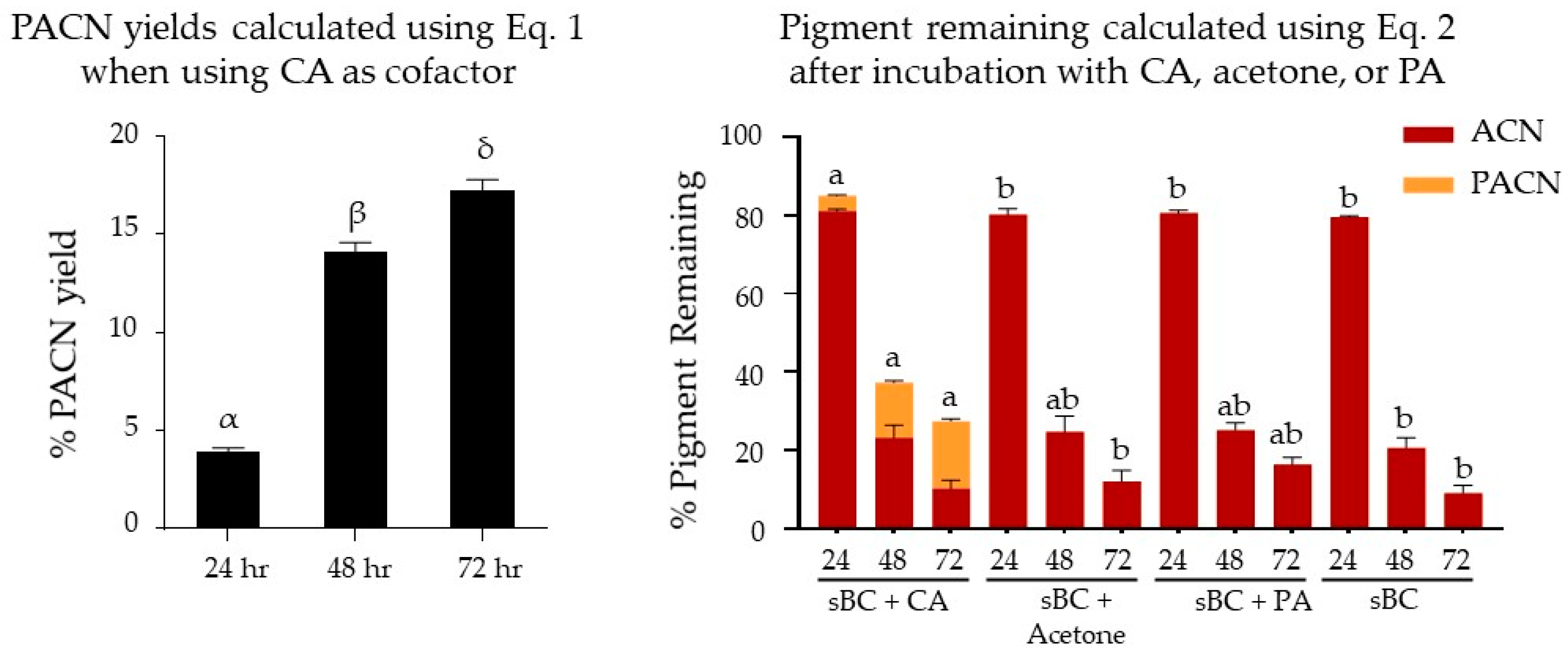
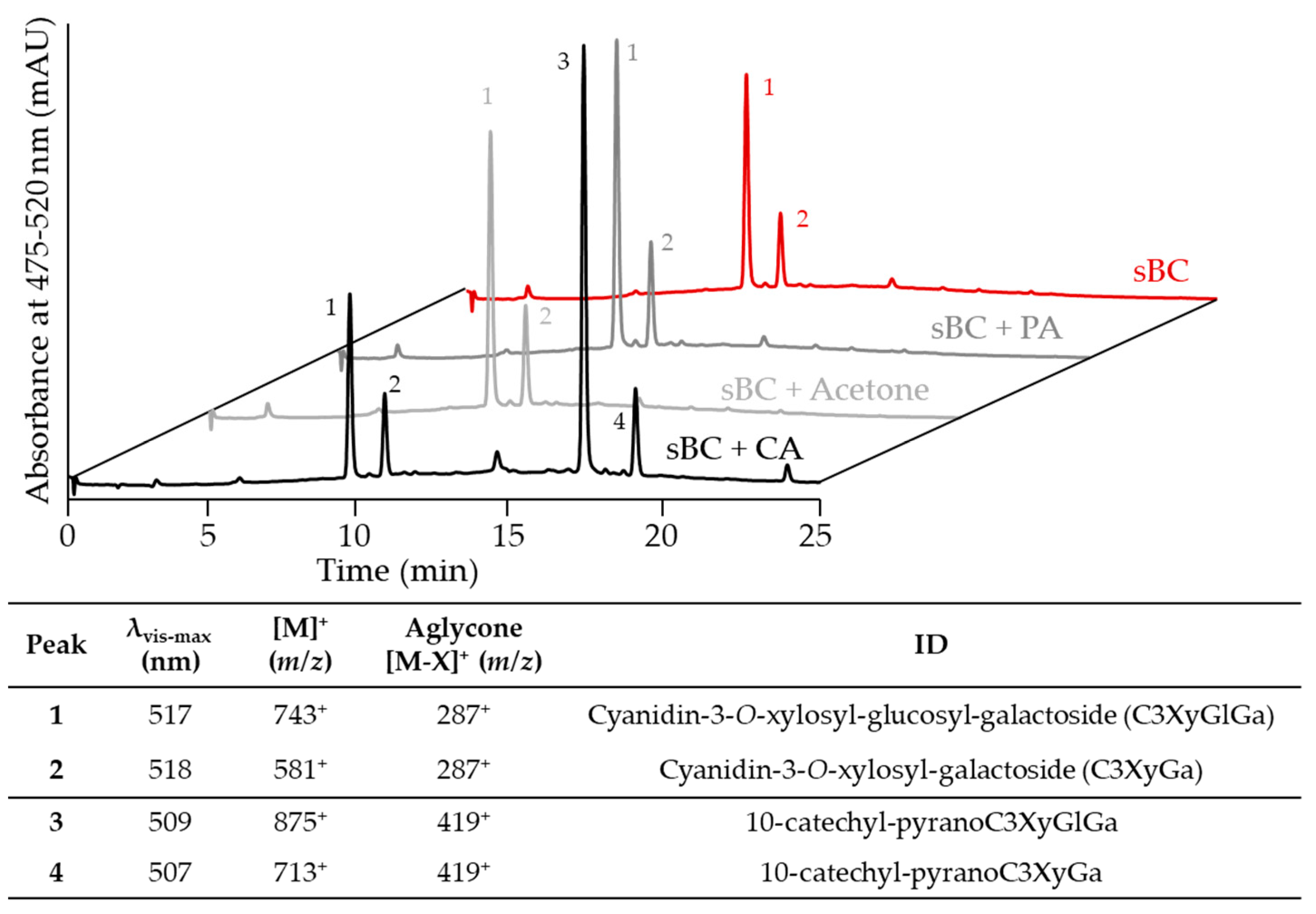
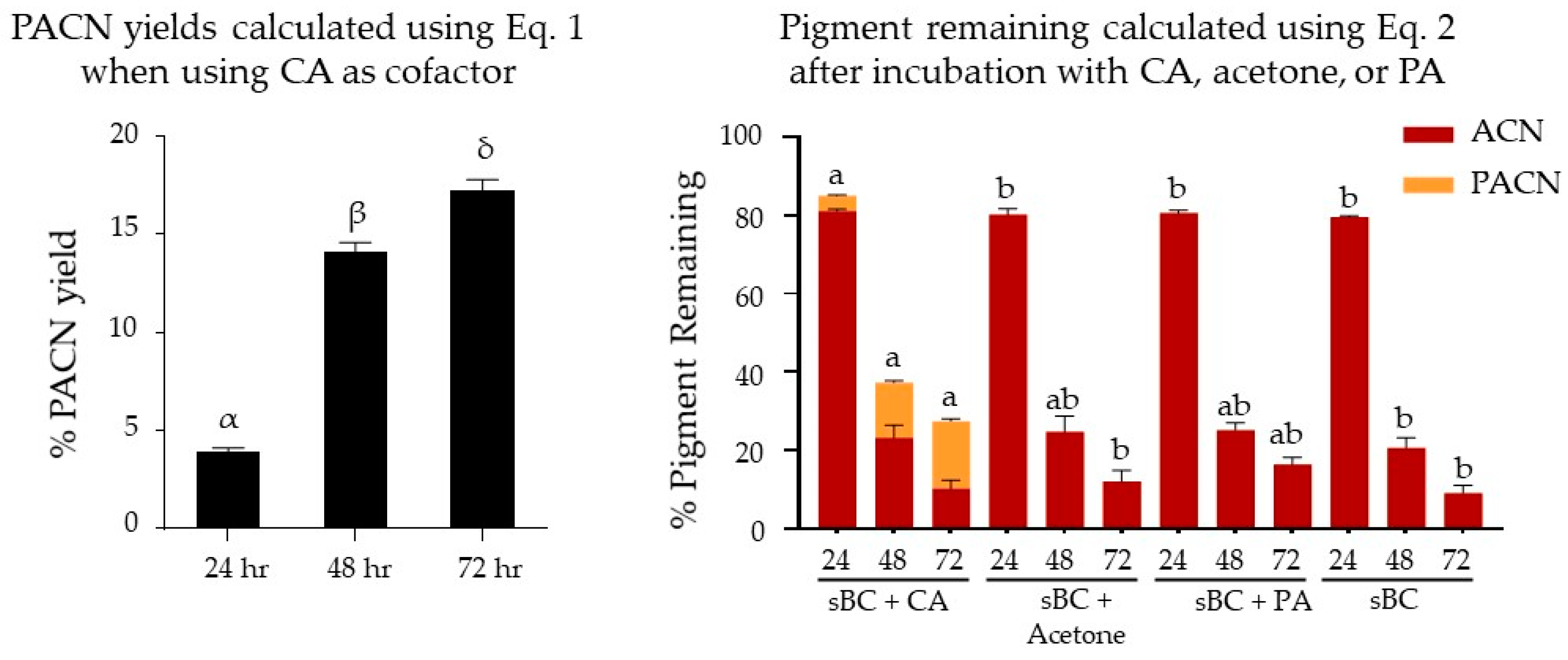
2.1. Evaluation of Pyranoanthocyanin Formation Efficiency Using Different Types of Cofactors
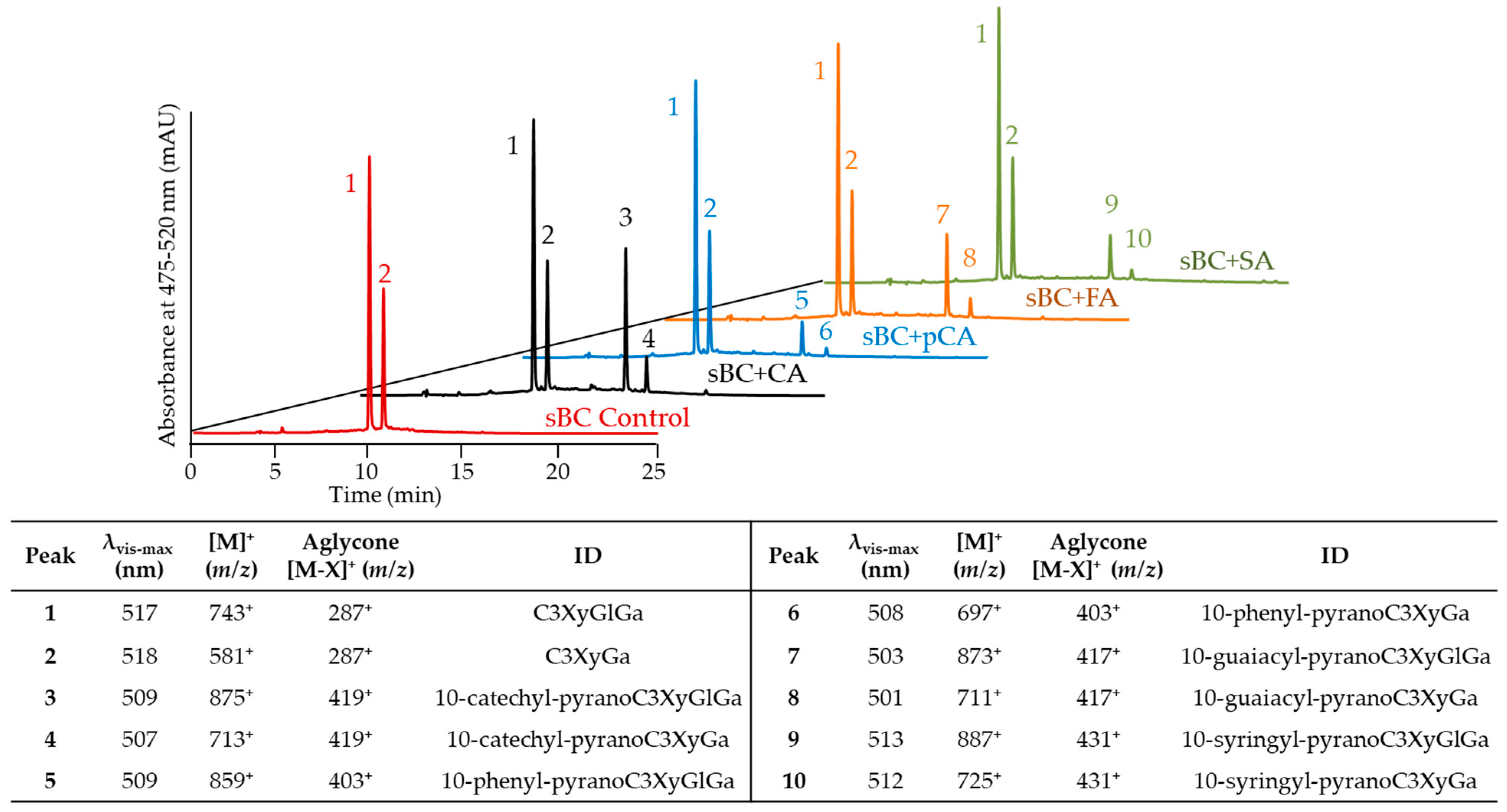
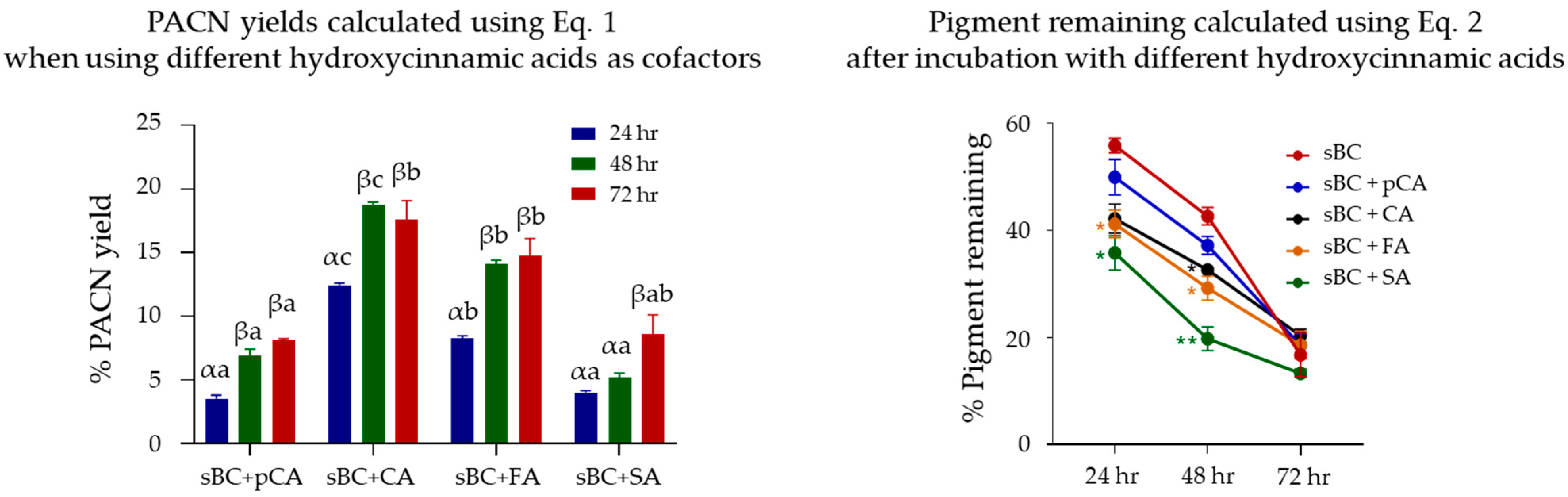
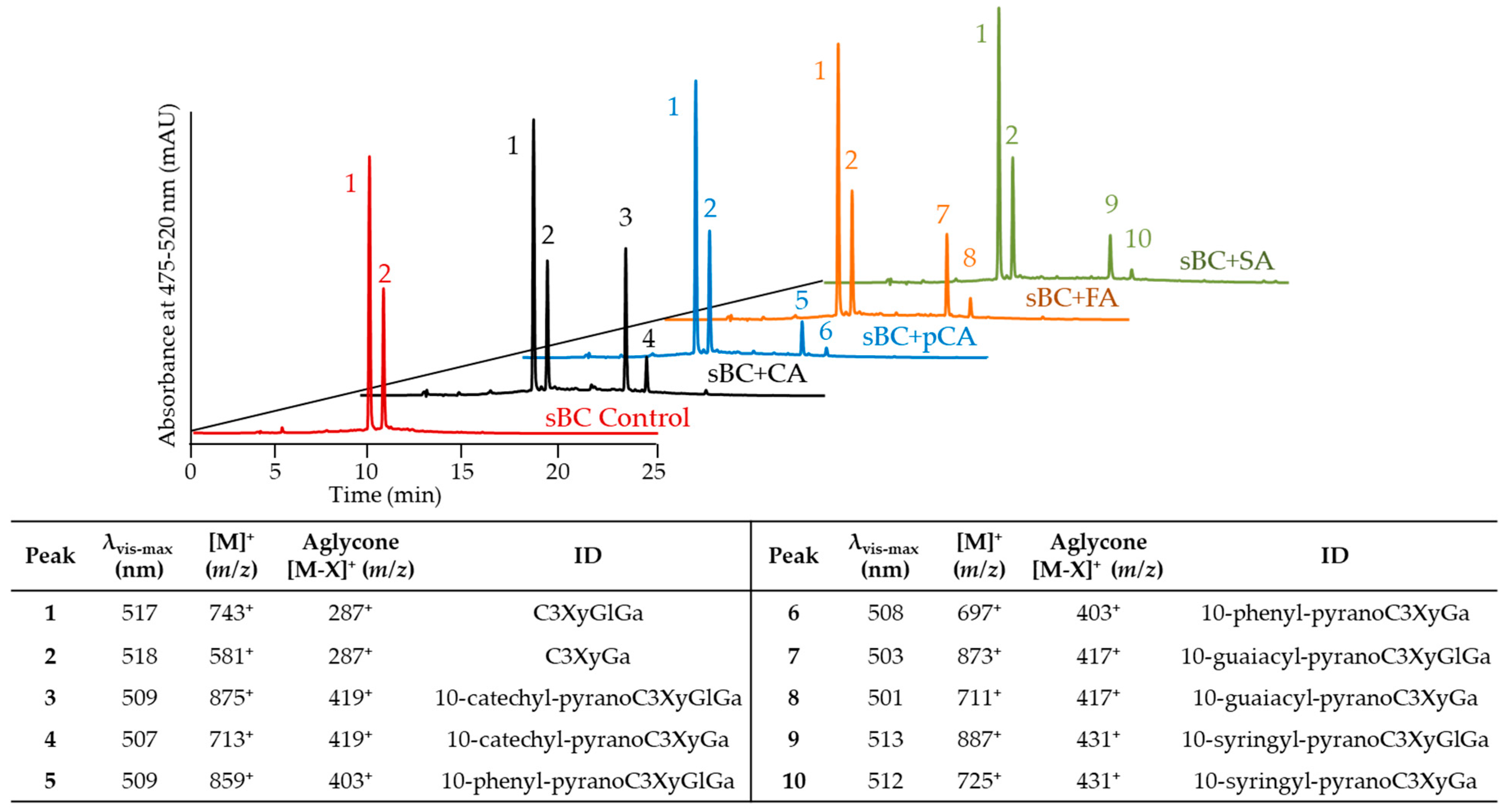
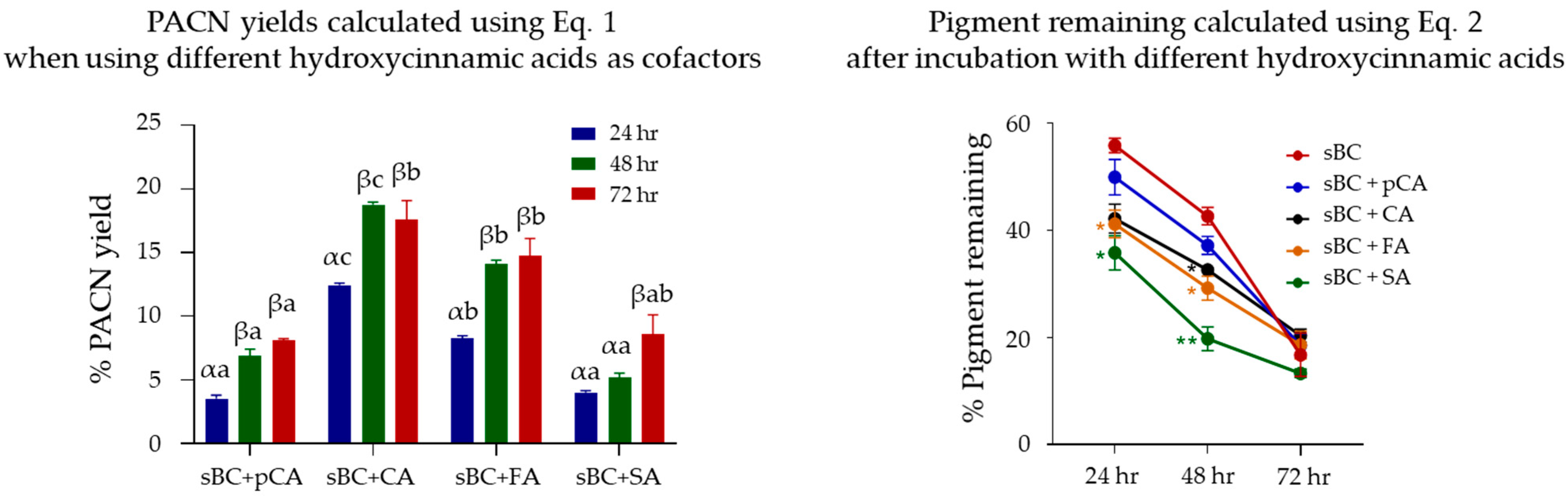
2.2. Evaluation of Pyranoanthocyanin Formation Efficiency Using Different Hydroxycinnamic Acids
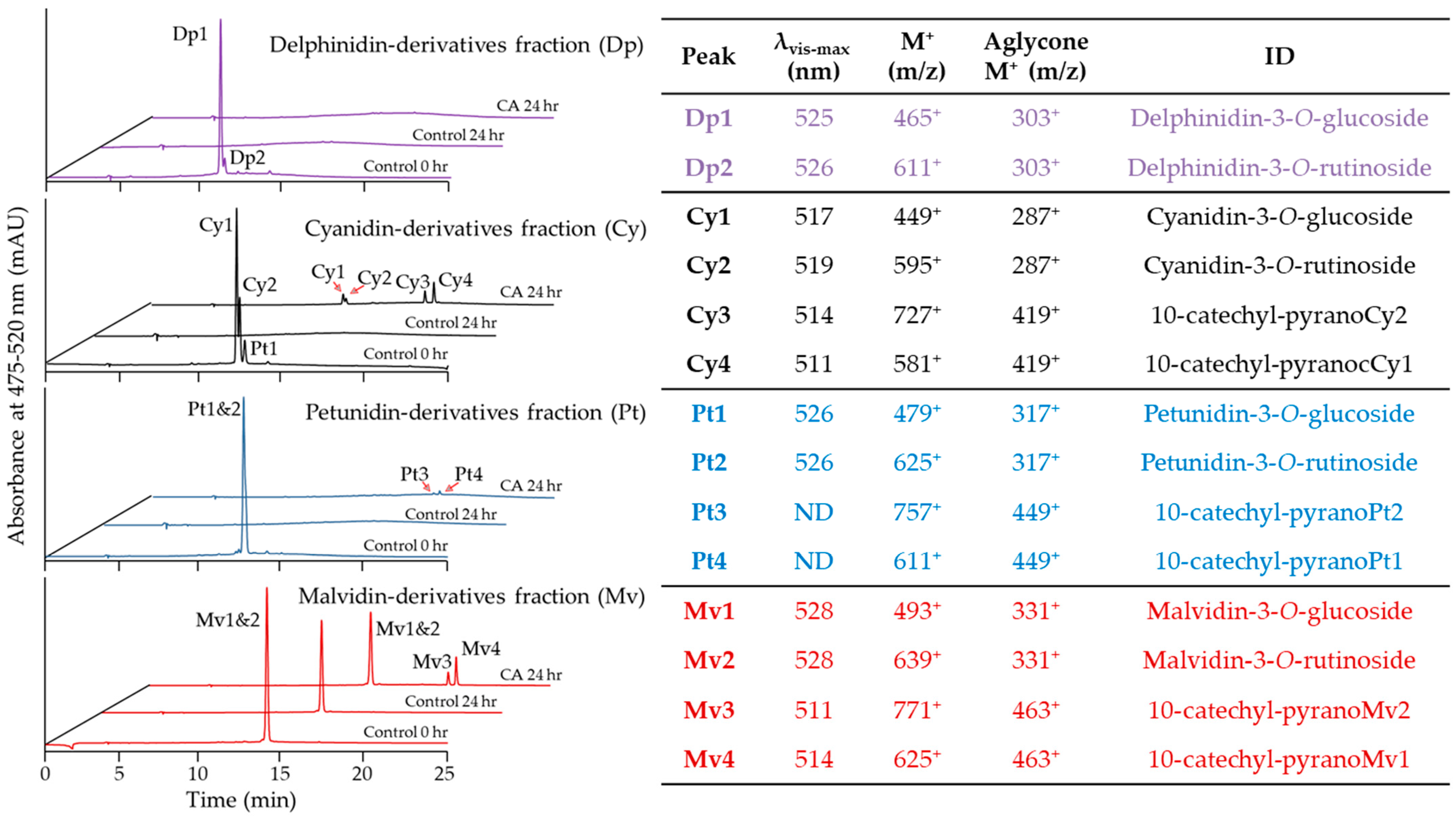
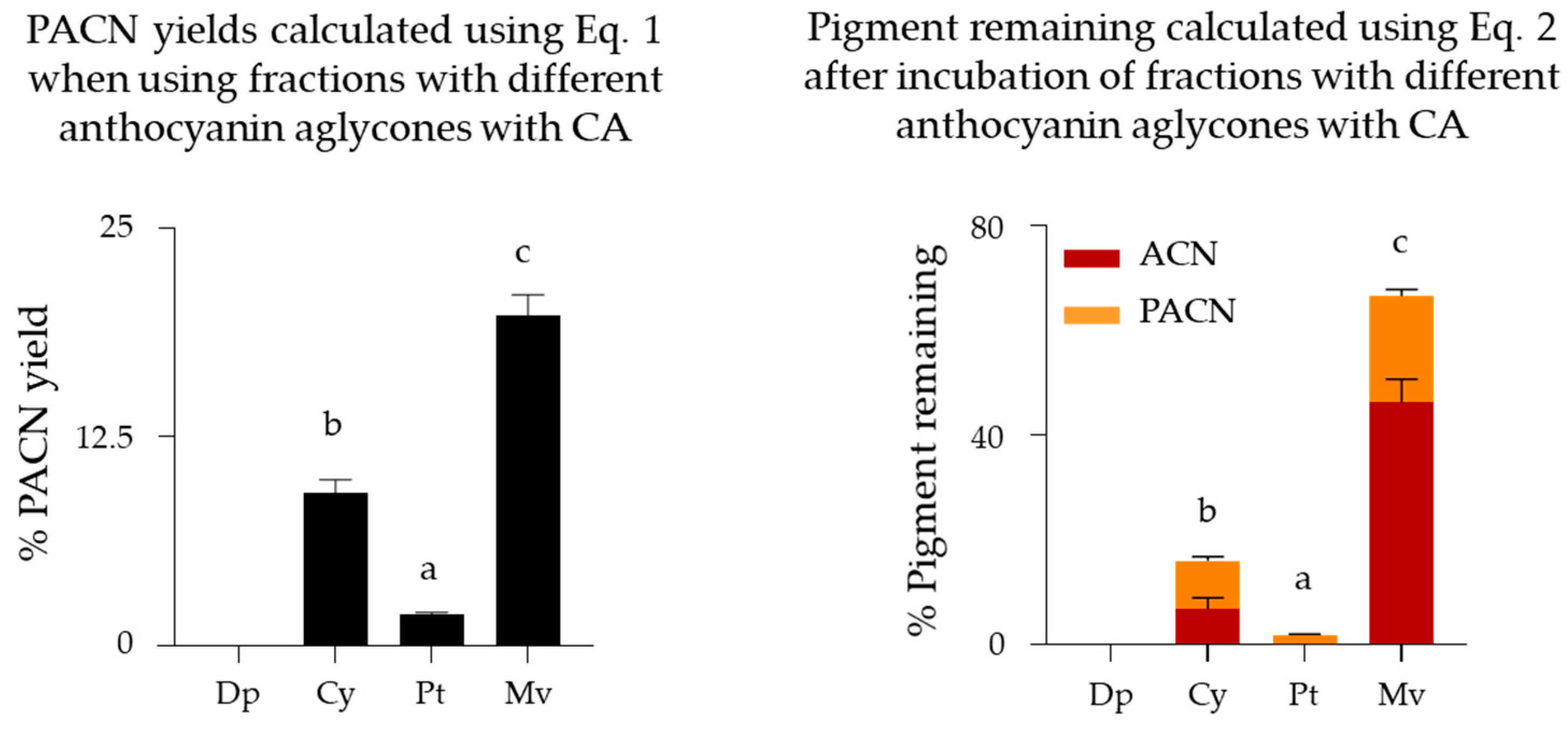
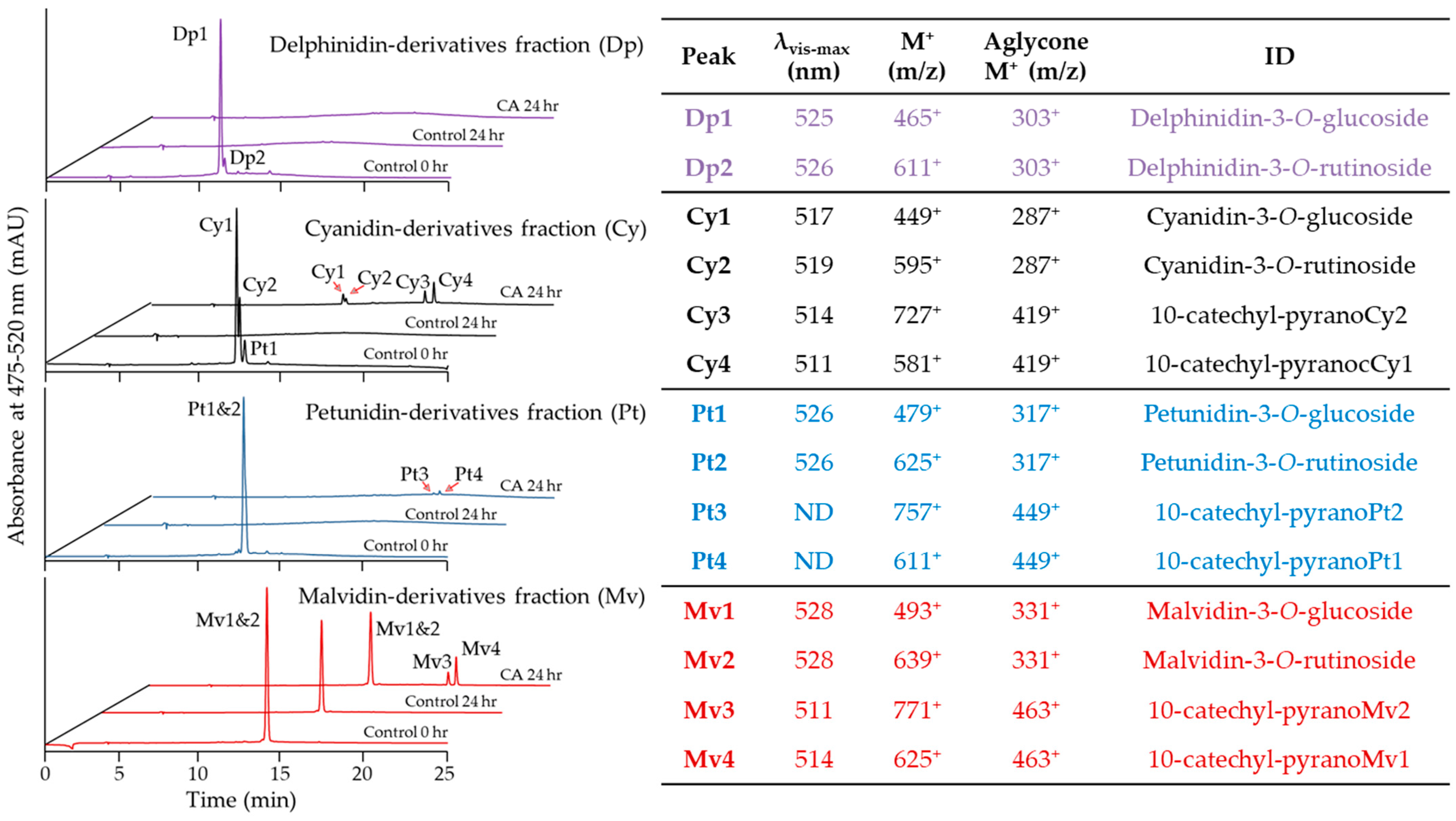
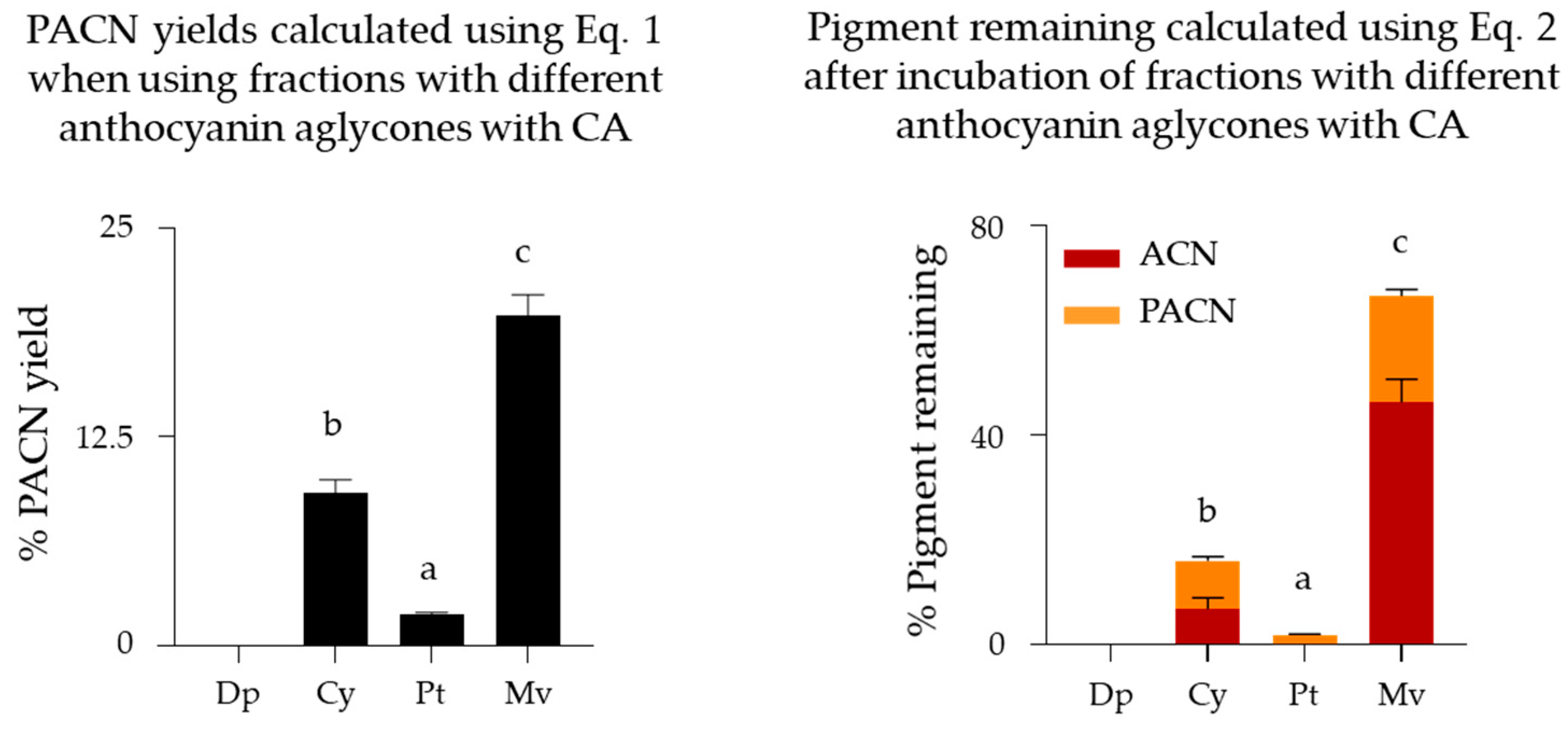
2.3. Evaluation of Pyranoanthocyanin Formation Efficiency Using Different Anthocyanins
3. Discussion
4. Materials and Methods
4.1. Plant Material, Chemical, and Reagents
4.2. Anthocyanin Preparation
4.3. Saponification of Black Carrot Anthocyanins
4.4. Pigment Semi-Purification
4.5. Fractionation of Different Anthocyanins
4.6. Anthocyanin and Pyranoanthoycanin Identification
4.7. Monomeric Anthocyanin Quantitation
4.8. Pyranoanthocyanin Formation—Comparing Different Types of Cofactors
4.9. Pyranoanthocyanin Formation—Comparing Different Hydroxycinnamic Acids
4.10. Pyranoanthocyanin Formation—Comparing Different Aglycones
4.11. Monitoring Pyranoanthocyanin Formation and Anthocyanin Changes Over Time
4.12. Statistical Evaluation of Data
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mazza, G.; Brouillard, R. Recent Developments in the Stabilization of Anthocyanins in Food Products. Food Chem. 1987, 25, 207–225. [Google Scholar] [CrossRef]
- Sigurdson, G.T.; Tang, P.; Giusti, M.M. Natural Colorants: Food Colorants from Natural Sources. Annu. Rev. Food Sci. Technol. 2017, 8, 261–280. [Google Scholar] [CrossRef] [PubMed]
- Bateman, B.; Warner, J.O.; Hutchinson, E.; Dean, T.; Rowlandson, P.; Gant, C.; Grundy, J.; Fitzgerald, C.; Stevenson, J. The Effects of a Double Blind, Placebo Controlled, Artificial Food Colourings and Benzoate Preservative Challenge on Hyperactivity in a General Population Sample of Preschool Children. Arch. Dis. Child. 2004, 89, 506–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCann, D.; Barrett, A.; Cooper, A.; Crumpler, D.; Dalen, L.; Grimshaw, K.; Kitchin, E.; Lok, K.; Porteous, L.; Prince, E.; et al. Food Additives and Hyperactive Behaviour in 3-Year-Old and 8/9-Year-Old Children in the Community: A Randomised, Double-Blinded, Placebo-Controlled Trial. Lancet 2007, 370, 1560–1567. [Google Scholar] [CrossRef]
- He, J.; Monica Giusti, M. Anthocyanins: Natural Colorants with Health-Promoting Properties. Annu. Rev. Food Sci. Technol. 2010, 1, 163–187. [Google Scholar] [CrossRef]
- Farr, J.E.; Giusti, M.M. Investigating the Interaction of Ascorbic Acid with Anthocyanins and Pyranoanthocyanins. Molecules 2018, 23, 744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pangestu, N.P.; Miyagusuku-Cruzado, G.; Giusti, M.M. Copigmentation with Chlorogenic and Ferulic Acid Affected Color and Anthocyanin Stability in Model Beverages Colored with Sambucus Peruviana, Sambucus Nigra and Daucus Carota during Storage. Foods 2020, 9, 1476. [Google Scholar] [CrossRef] [PubMed]
- Miyagusuku-Cruzado, G.; Jiménez-Flores, R.; Giusti, M.M. Whey Protein Addition and Its Increased Light Absorption and Tinctorial Strength of Model Solutions Colored with Anthocyanins. J. Dairy Sci. 2020, 104, 6449–6462. [Google Scholar] [CrossRef]
- Weber, F.; Boch, K.; Schieber, A. Influence of Copigmentation on the Stability of Spray Dried Anthocyanins from Blackberry. LWT Food Sci. Technol. 2017, 75, 72–77. [Google Scholar] [CrossRef]
- Tang, P.; Giusti, M.M. Metal Chelates of Petunidin Derivatives Exhibit Enhanced Color and Stability. Foods 2020, 9, 1426. [Google Scholar] [CrossRef]
- Sun, J.; Li, X.; Luo, H.; Ding, L.; Jiang, X.; Li, X.; Jiao, R.; Bai, W. Comparative Study on the Stability and Antioxidant Activity of Six Pyranoanthocyanins Based on Malvidin-3-Glucoside. J. Agric. Food Chem. 2020, 68, 2783–2794. [Google Scholar] [CrossRef]
- Farr, J.E.; Sigurdson, G.T.; Giusti, M.M. Influence of Cyanidin Glycosylation Patterns on Carboxypyranoanthocyanin Formation. Food Chem. 2018, 259, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Velenosi, M.; Crupi, P.; Perniola, R.; Marsico, A.D.; Salerno, A.; Alexandre, H.; Archidiacono, N.; Ventura, M.; Cardone, M.F. Color Stabilization of Apulian Red Wines through the Sequential Inoculation of Starmerella Bacillaris and Saccharomyces Cerevisiae. Molecules 2021, 26, 907. [Google Scholar] [CrossRef] [PubMed]
- Marquez, A.; Serratosa, M.P.; Merida, J. Pyranoanthocyanin Derived Pigments in Wine: Structure and Formation during Winemaking. J. Chem. 2013, 2013, 713028. [Google Scholar] [CrossRef]
- Bakker, J.; Timberlake, C.F. Isolation, Identification, and Characterization of New Color-Stable Anthocyanins Occurring in Some Red Wines. J. Agric. Food Chem. 1997, 45, 35–43. [Google Scholar] [CrossRef]
- Mateus, N.; Silva, A.M.S.; Vercauteren, J.; de Freitas, V. Occurrence of Anthocyanin-Derived Pigments in Red Wines. J. Agric. Food Chem. 2001, 49, 4836–4840. [Google Scholar] [CrossRef]
- He, J.; Carvalho, A.R.F.; Mateus, N.; de Freitas, V. Spectral Features and Stability of Oligomeric Pyranoanthocyanin-Flavanol Pigments Isolated from Red Wines. J. Agric. Food Chem. 2010, 58, 9249–9258. [Google Scholar] [CrossRef] [PubMed]
- de Freitas, V.; Mateus, N. Formation of Pyranoanthocyanins in Red Wines: A New and Diverse Class of Anthocyanin Derivatives. Anal. Bioanal. Chem. 2011, 401, 1467–1477. [Google Scholar] [CrossRef]
- Schwarz, M.; Wray, V.; Winterhalter, P. Isolation and Identification of Novel Pyranoanthocyanins from Black Carrot (Daucus Carota L.) Juice. J. Agric. Food Chem. 2004, 52, 5095–5101. [Google Scholar] [CrossRef]
- Blanco-Vega, D.; López-Bellido, F.J.; Alía-Robledo, J.M.; Hermosín-Gutiérrez, I. HPLC-DAD-ESI-MS/MS Characterization of Pyranoanthocyanins Pigments Formed in Model Wine. J. Agric. Food Chem. 2011, 59, 9523–9531. [Google Scholar] [CrossRef]
- Fossen, T.; Andersen, Ø.M. Anthocyanins from Red Onion, Allium Cepa, with Novel Aglycone. Phytochemistry 2003, 62, 1217–1220. [Google Scholar] [CrossRef]
- Andersen, Ø.M.; Fossen, T.; Torskangerpoll, K.; Fossen, A.; Hauge, U. Anthocyanin from Strawberry (Fragaria ananassa) with the Novel Aglycone, 5-Carboxypyranopelargonidin. Phytochemistry 2004, 65, 405–410. [Google Scholar] [CrossRef]
- Peng, Y.; Zhang, H.; Liu, R.; Mine, Y.; McCallum, J.; Kirby, C.; Tsao, R. Antioxidant and Anti-Inflammatory Activities of Pyranoanthocyanins and Other Polyphenols from Staghorn Sumac (Rhus Hirta L.) in Caco-2 Cell Models. J. Funct. Foods 2016, 20, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Laaksonen, O.; Yang, W.; Zhang, B.; Yang, B. Pyranoanthocyanins in Bilberry (Vaccinium Myrtillus L.) Wines Fermented with Schizosaccharomyces Pombe and Their Evolution during Aging. Food Chem. 2020, 305, 125438. [Google Scholar] [CrossRef]
- Rein, M.J.; Ollilainen, V.; Vahermo, M.; Yli-Kauhaluoma, J.; Heinonen, M. Identification of Novel Pyranoanthocyanins in Berry Juices. Eur. Food Res. Technol. 2005, 220, 239–244. [Google Scholar] [CrossRef]
- Mateus, N.; Oliveira, J.; Haettich-Motta, M.; de Freitas, V. New Family of Bluish Pyranoanthocyanins. J. Biomed. Biotechnol. 2004, 2004, 299–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Giusti, M.M. Pyranoanthocyanin Formation Rates and Yields as Affected by Cyanidin-3-Substitutions and Pyruvic or Caffeic Acids. Food Chem. 2021, 345, 128776. [Google Scholar] [CrossRef] [PubMed]
- Hoehn, M.E. Altering pH, Temperature and Cofactors to Increase the Formation of the More Stable Anthocyanin Derived Pyranoanthocyanin. Master’s Thesis, The Ohio State University, Columbus, OH, USA, 2019. [Google Scholar]
- Straathof, N.; Giusti, M.M. Improvement of Naturally Derived Food Colorant Performance with Efficient Pyranoanthocyanin Formation from Sambucus Nigra Anthocyanins Using Caffeic Acid and Heat. Molecules 2020, 25, 5998. [Google Scholar] [CrossRef]
- Oliveira, J.; de Freitas, V.; Silva, A.M.S.; Mateus, N. Reaction between Hydroxycinnamic Acids and Anthocyanin-Pyruvic Acid Adducts Yielding New Portisins. J. Agric. Food Chem. 2007, 55, 6349–6356. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, M.; Wabnitz, T.C.; Winterhalter, P. Pathway Leading to the Formation of Anthocyanin-Vinylphenol Adducts and Related Pigments in Red Wines. J. Agric. Food Chem. 2003, 51, 3682–3687. [Google Scholar] [CrossRef] [PubMed]
- del Carpio Jiménez, C.; Serrano Flores, C.; He, J.; Tian, Q.; Schwartz, S.J.; Giusti, M.M. Characterisation and Preliminary Bioactivity Determination of Berberis Boliviana Lechler Fruit Anthocyanins. Food Chem. 2011, 128, 717–724. [Google Scholar] [CrossRef]
- Topić Božič, J.; Ćurko, N.; Kovačević Ganić, K.; Butinar, L.; Albreht, A.; Vovk, I.; Korte, D.; Mozetič Vodopivec, B. Synthesis of Pyranoanthocyanins from Pinot Noir Grape Skin Extract Using Fermentation with High Pyranoanthocyanin Producing Yeasts and Model Wine Storage as Potential Approaches in the Production of Stable Natural Food Colorants. Eur. Food Res. Technol. 2020, 246, 1141–1152. [Google Scholar] [CrossRef]
- Eiro, M.J.; Heinonen, M. Anthocyanin Color Behavior and Stability during Storage: Effect of Intermolecular Copigmentation. J. Agric. Food Chem. 2002, 50, 7461–7466. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Wang, Y.; Xie, P.; Zhang, L.; Li, Y.; Zhou, J. Copigmentation Effects of Phenolics on Color Enhancement and Stability of Blackberry Wine Residue Anthocyanins: Chromaticity, Kinetics and Structural Simulation. Food Chem. 2019, 275, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Boulton, R. The Copigmentation of Anthocyanins and Its Role in the Color of Red Wine: A Critical Review. Am. J. Enol. Vitic. 2001, 52, 67–87. [Google Scholar]
- Zheng, W.; Wang, S.Y. Oxygen Radical Absorbing Capacity of Phenolics in Blueberries, Cranberries, Chokeberries, and Lingonberries. J. Agric. Food Chem. 2003, 51, 502–509. [Google Scholar] [CrossRef]
- Frond, A.D.; Iuhas, C.I.; Stirbu, I.; Leopold, L.; Socaci, S.; Andreea, S.; Ayvaz, H.; Andreea, S.; Mihai, S.; Diaconeasa, Z.; et al. Phytochemical Characterization of Five Edible Purple-Reddish Vegetables: Anthocyanins, Flavonoids, and Phenolic Acid Derivatives. Molecules 2019, 24, 1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torre, P.; Aliakbarian, B.; Rivas, B.; Domínguez, J.M.; Converti, A. Release of Ferulic Acid from Corn Cobs by Alkaline Hydrolysis. Biochem. Eng. J. 2008, 40, 500–506. [Google Scholar] [CrossRef]
- Schwarz, M.; Winterhalter, P. A Novel Synthetic Route to Substituted Pyranoanthocyanins with Unique Colour Properties. Tetrahedron Lett. 2003, 44, 7583–7587. [Google Scholar] [CrossRef]
- Gaspar, A.; Garrido, E.M.; Esteves, M.; Quezada, E.; Milhazes, N.; Garrido, J.; Borges, F. New Insights into the Antioxidant Activity of Hydroxycinnamic Acids: Synthesis and Physicochemical Characterization of Novel Halogenated Derivatives. Eur. J. Med. Chem. 2009, 44, 2092–2099. [Google Scholar] [CrossRef]
- Song, B.J.; Sapper, T.N.; Burtch, C.E.; Brimmer, K.; Goldschmidt, M.; Ferruzzi, M.G. Photo- and Thermodegradation of Anthocyanins from Grape and Purple Sweet Potato in Model Beverage Systems. J. Agric. Food Chem. 2013, 61, 1364–1372. [Google Scholar] [CrossRef]
- Wang, F.; Li, H.; Qin, Y.; Mao, Y.; Zhang, B.; Deng, Z. Effects of Heat, Ultrasound, and Microwave Processing on the Stability and Antioxidant Activity of Delphinidin and Petunidin. J. Food Biochem. 2019, 43, e12818. [Google Scholar] [CrossRef] [PubMed]
- Benito, S.; Morata, A.; Palomero, F.; González, M.C.; Suárez-Lepe, J.A. Formation of Vinylphenolic Pyranoanthocyanins by Saccharomyces Cerevisiae and Pichia Guillermondii in Red Wines Produced Following Different Fermentation Strategies. Food Chem. 2011, 124, 15–23. [Google Scholar] [CrossRef]
- Hillebrand, S.; Schwarz, M.; Winterhalter, P. Characterization of Anthocyanins and Pyranoanthocyanins from Blood Orange [Citrus Sinensis (L.) Osbeck] Juice. J. Agric. Food Chem. 2004, 52, 7331–7338. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Saona, L.E.; Wrolstad, R.E. Extraction, Isolation, and Purifification of Anthocyanins. Curr. Protoc. Food Anal. Chem. 2001, F1.1.1–F1.1.11. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Characterization of Red Radish Anthocyanins. J. Food Sci. 1996, 61, 322–326. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Characterization and Measurement of Anthocyanins by UV-Visible Spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1–13. [Google Scholar] [CrossRef]
- Cabrita, L.; Fossen, T.; Andersen, Ø.M. Colour and Stability of the Six Common Anthocyanidin 3-Glucosides in Aqueous Solutions. Food Chem. 2000, 68, 101–107. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miyagusuku-Cruzado, G.; Voss, D.M.; Giusti, M.M. Influence of the Anthocyanin and Cofactor Structure on the Formation Efficiency of Naturally Derived Pyranoanthocyanins. Int. J. Mol. Sci. 2021, 22, 6708. https://doi.org/10.3390/ijms22136708
Miyagusuku-Cruzado G, Voss DM, Giusti MM. Influence of the Anthocyanin and Cofactor Structure on the Formation Efficiency of Naturally Derived Pyranoanthocyanins. International Journal of Molecular Sciences. 2021; 22(13):6708. https://doi.org/10.3390/ijms22136708
Chicago/Turabian StyleMiyagusuku-Cruzado, Gonzalo, Danielle M. Voss, and M. Monica Giusti. 2021. "Influence of the Anthocyanin and Cofactor Structure on the Formation Efficiency of Naturally Derived Pyranoanthocyanins" International Journal of Molecular Sciences 22, no. 13: 6708. https://doi.org/10.3390/ijms22136708
APA StyleMiyagusuku-Cruzado, G., Voss, D. M., & Giusti, M. M. (2021). Influence of the Anthocyanin and Cofactor Structure on the Formation Efficiency of Naturally Derived Pyranoanthocyanins. International Journal of Molecular Sciences, 22(13), 6708. https://doi.org/10.3390/ijms22136708